
Congo Red as a Supramolecular Carrier System for Doxorubicin: An Approach to Understanding the Mechanism of Action

 , , , , , and
, , , , , and 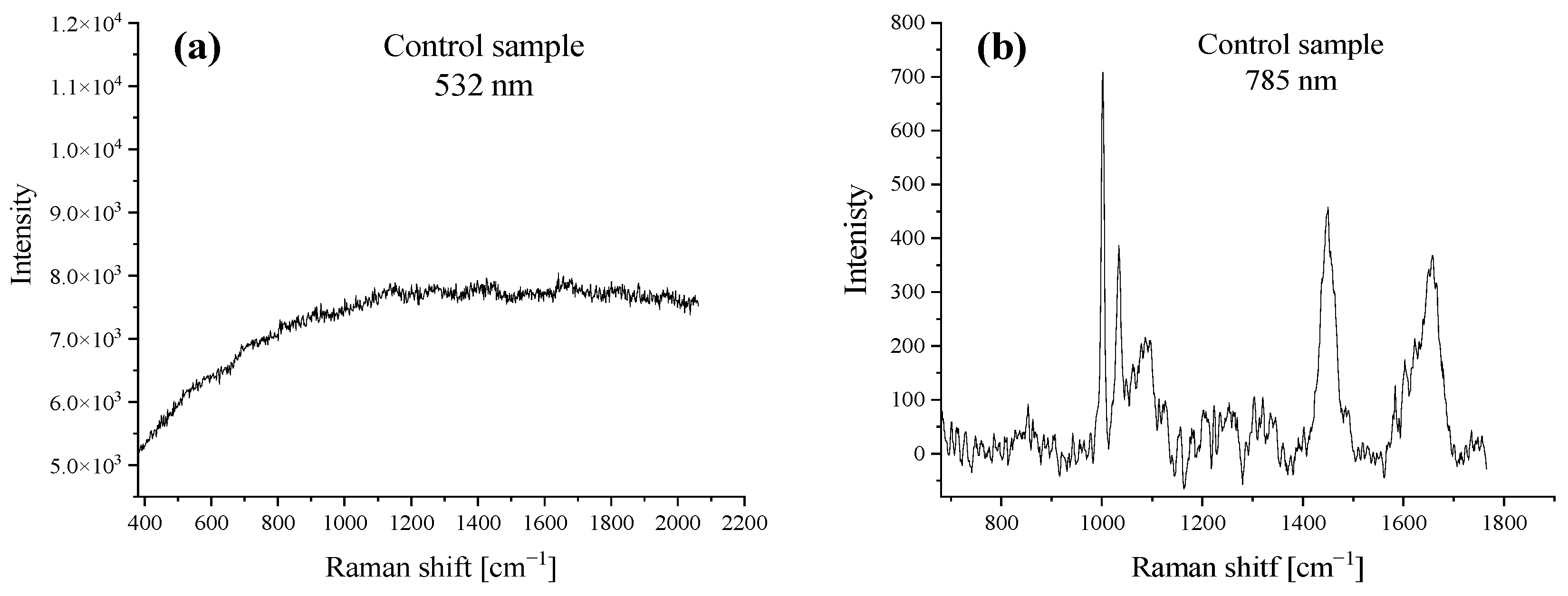{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Raman Spectroscopy
2.2. Monolayer Experiments
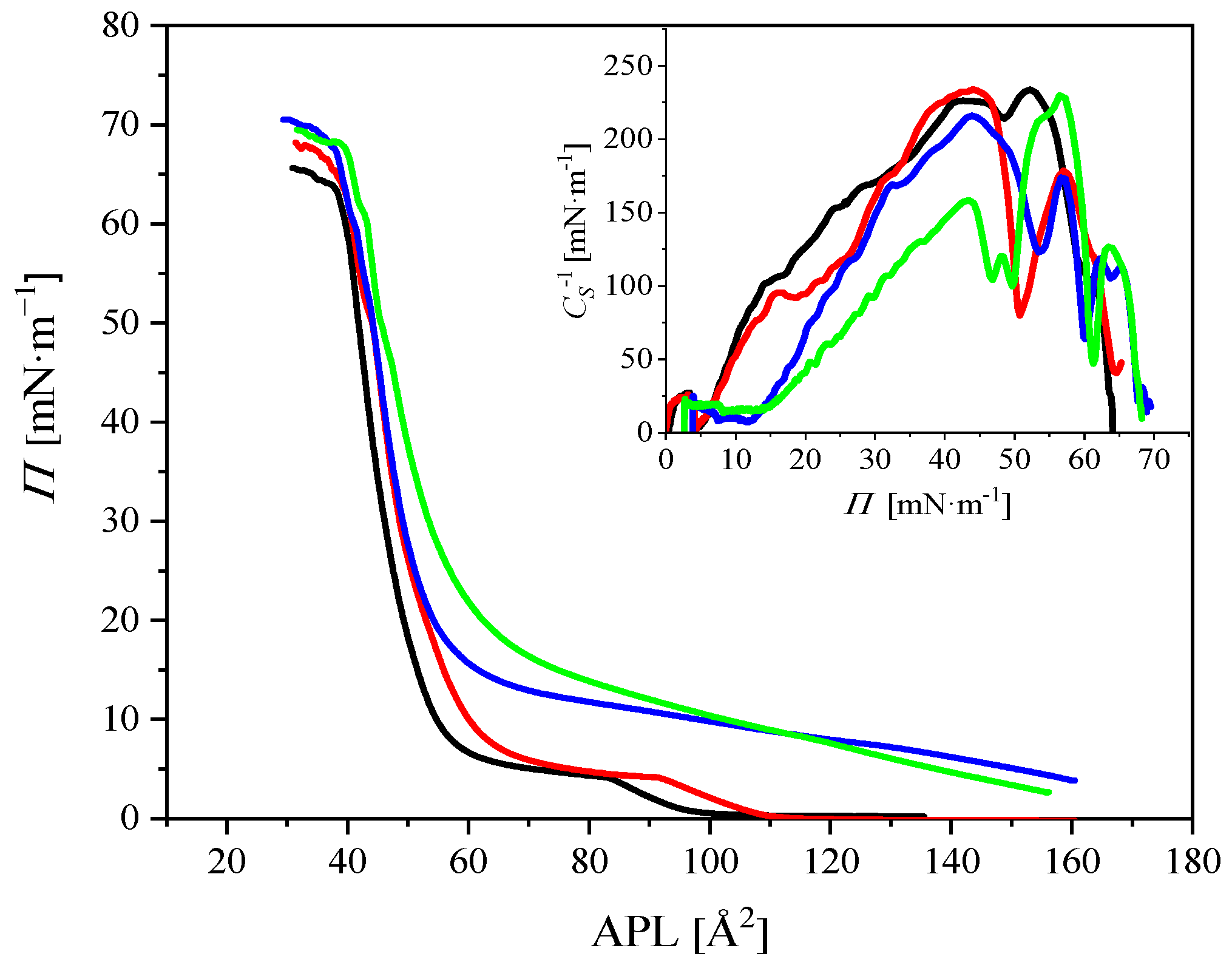
2.2.1. Compression Isotherms
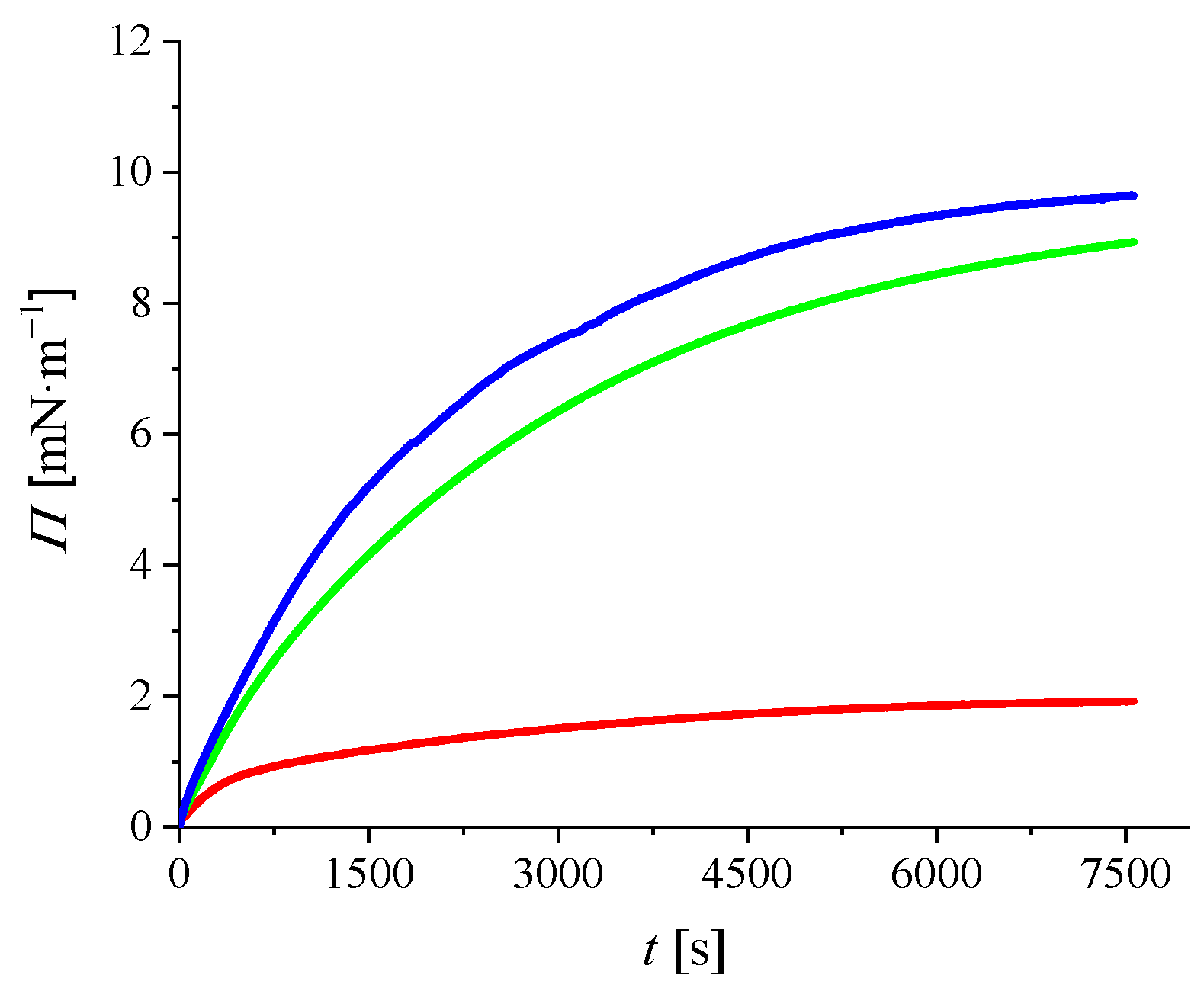
2.2.2. Adsorption Kinetics
2.3. Molecular Dynamics Simulations
2.3.1. Potential of Mean Force Calculations
2.3.2. Unbiased MD Simulations
3. Materials and Methods
3.1. Materials
3.2. Raman Spectroscopy
3.3. Surface Pressure–Area Isotherms
3.4. Adsorption Kinetics Measurement
3.5. MD Simulations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arcamone, F.; Cassinelli, G.; Fantini, G.; Grein, A.; Orezzi, P.; Pol, C.; Spalla, C. Adriamycin, 14-hydroxydaunomycin, a new antitumor antibiotic from S. peucetius var. Caesius. Biotechnol. Bioeng. 2000, 67, 704–713. [Google Scholar] [CrossRef]
- Blum, R.H.; Carter, S.K. Adriamycin—New anticancer drug with significant clinical activity. Ann. Intern. Med. 1974, 80, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Bender, R.A.; Zwelling, L.A.; Doroshow, J.H.; Locker, G.Y.; Hande, K.R.; Murinson, D.S.; Cohen, M.; Myers, C.E.; Chabner, B.A. Anti-neoplastic drugs—Clinical pharmacology and therapeutic use. Drugs 1978, 16, 46–87. [Google Scholar] [CrossRef] [PubMed]
- Gewirtz, D.A. A critical evaluation of the mechanisms of action proposed for the antitumor effects of the anthracycline antibiotics adriamycin and daunorubicin. Biochem. Pharmacol. 1999, 57, 727–741. [Google Scholar] [CrossRef]
- Mohan, U.P.; Pichiah, P.B.T.; Iqbal, S.T.A.; Arunachalam, S. Mechanisms of doxorubicin-mediated reproductive toxicity—A review. Reprod. Toxicol. 2021, 102, 80–89. [Google Scholar] [CrossRef]
- Umlauf, J.; Horky, M. Molecular biology of doxorubicin-induced cardiomyopathy. Exp. Clin. Cardiol. 2002, 7, 35–39. [Google Scholar]
- Pasvolsky, O.; Morelli, O.; Rozovski, U.; Vaturi, M.; Wolach, O.; Amitai, I.; Vaxman, I.; Ratzon, R.; Yeshurun, M.; Kornowski, R.; et al. Anthracycline-induced cardiotoxicity in acute myeloid leukemia patients who undergo allogeneic hematopoietic stem cell transplantation. Clin. Lymphoma Myeloma Leuk. 2019, 19, E343–E348. [Google Scholar] [CrossRef]
- Jawad, B.; Poudel, L.; Podgornik, R.; Steinmetz, N.F.; Ching, W.Y. Molecular mechanism and binding free energy of doxorubicin intercalation in DNA. Phys. Chem. Chem. Phys. 2019, 21, 3877–3893. [Google Scholar] [CrossRef]
- Hasinoff, B.B.; Patel, D.; Wu, X. The role of topoisomerase II beta in the mechanisms of action of the doxorubicin cardioprotective agent dexrazoxane. Cardiovasc. Toxicol. 2020, 20, 312–320. [Google Scholar] [CrossRef]
- Khan, S.A.; Akhtar, M.J. Structural modification and strategies for the enhanced doxorubicin drug delivery. Bioorg. Chem. 2022, 120, 105599. [Google Scholar] [CrossRef]
- Senapati, S.; Mahanta, A.K.; Kumar, S.; Maiti, P. Controlled drug delivery vehicles for cancer treatment and their performance. Signal Transduct. Target. Ther. 2018, 3, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soppimath, K.S.; Aminabhavi, T.M.; Kulkarni, A.R.; Rudzinski, W.E. Biodegradable polymeric nanoparticles as drug delivery devices. J. Control. Release 2001, 70, 1–20. [Google Scholar] [CrossRef]
- Kumar, S.; Singh, S.; Senapati, S.; Singh, A.P.; Ray, B.; Maiti, P. Controlled drug release through regulated biodegradation of poly(lactic acid) using inorganic salts. Int. J. Biol. Macromol. 2017, 104, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Mo, R.; Jiang, T.Y.; Gu, Z. Recent progress in multidrug delivery to cancer cells by liposomes. Nanomedicine 2014, 9, 1117–1120. [Google Scholar] [CrossRef]
- Peng, P.P.; Yang, K.; Tong, G.S.; Ma, L. Polysaccharide nanoparticles for targeted cancer therapies. Curr. Drug Metab. 2018, 19, 781–792. [Google Scholar] [CrossRef]
- Patra, C.R.; Bhattacharya, R.; Mukhopadhyay, D.; Mukherjee, P. Application of gold nanoparticles for targeted therapy in cancer. J. Biomed. Nanotech. 2008, 4, 99–132. [Google Scholar] [CrossRef]
- Kim, H.; Lee, Y.; Kang, S.; Choi, M.; Lee, S.; Kim, S.; Gujrati, V.; Kim, J.; Jon, S. Self-assembled nanoparticles comprising aptide-SN38 conjugates for use in targeted cancer therapy. Nanotechnology 2016, 27, 48LT01. [Google Scholar] [CrossRef]
- Sailaja, G.S.; Gondi, C.S.; Bhoopathi, P.; Gorantla, B.; Rao, J.S. Self-assembled superparamagnetic iron oxide nanoparticles for targeted cancer therapy. Cancer Res. 2012, 72, 4758. [Google Scholar] [CrossRef]
- Jagusiak, A.; Chlopas, K.; Zemanek, G.; Koscik, I.; Roterman, I. Interaction of supramolecular Congo red and Congo red-doxorubicin complexes with proteins for drug carrier design. Pharmaceutics 2021, 13, 2027. [Google Scholar] [CrossRef]
- Roterman, I.; Konieczny, L. (Eds.) Self-Assembled Molecules—New Kind of Protein Ligands; Springer: Cham, Switzerland, 2018; p. 21. [Google Scholar]
- Frid, P.; Anisimov, S.V.; Popovic, N. Congo red and protein aggregation in neurodegenerative diseases. Brain Res. Rev. 2007, 53, 135–160. [Google Scholar] [CrossRef]
- Ptak-Kaczor, M.; Kwiecinska, K.; Korchowiec, J.; Chlopas, K.; Banach, M.; Roterman, I.; Jagusiak, A. Structure and location of protein sites binding self-associated Congo red molecules with intercalated drugs as compact ligands-theoretical studies. Biomolecules 2021, 11, 501. [Google Scholar]
- Bosio, V.E.; Lopez, A.G.; Mukherjee, A.; Mechetti, M.; Castro, G.R. Tailoring doxorubicin sustainable release from biopolymeric smart matrix using congo red as molecular helper. J. Mater. Chem. B 2014, 2, 5178–5186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szlasa, W.; Zendran, I.; Zalesinska, A.; Tarek, M.; Kulbacka, J. Lipid composition of the cancer cell membrane. J. Bioenerg. Biomembr. 2020, 52, 321–342. [Google Scholar] [CrossRef] [PubMed]
- Zech, T.; Ejsing, C.S.; Gaus, K.; de Wet, B.; Shevchenko, A.; Simons, K.; Harder, T. Accumulation of raft lipids in T-cell plasma membrane domains engaged in TCR signalling. EMBO J. 2009, 28, 466–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freudenthal, O.; Quiles, F.; Francius, G.; Wojszko, K.; Gorczyca, M.; Korchowiec, B.; Rogalska, E. Nanoscale investigation of the interaction of colistin with model phospholipid membranes by Langmuir technique, and combined infrared and force spectroscopies. Biochim. Biophys. Acta Biomembr. 2016, 1858, 2592–2602. [Google Scholar] [CrossRef]
- Gravier, J.; Korchowiec, B.; Schneider, R.; Rogalska, E. Interaction of amphiphilic chlorin-based photosensitizers with 1,2-dipalmitoyl-sn-glycero-3-phosphocholine monolayers. Chem. Phys. Lipids 2009, 158, 102–109. [Google Scholar] [CrossRef]
- Gorczyca, M.; Korchowiec, B.; Korchowiec, J.; Trojan, S.; Rubio-Magnieto, J.; Luis, S.V.; Rogalska, E. A study of the interaction between a family of gemini amphiphilic pseudopeptides and model monomolecular film membranes formed with a cardiolipin. J. Phys. Chem. B 2015, 119, 6668–6679. [Google Scholar] [CrossRef]
- Korchowiec, B.M.; Baba, T.; Minamikawa, H.; Hato, M. Forces that control pH-dependent aggregation of nonionic glycolipid vesicles. Langmuir 2001, 17, 1853–1859. [Google Scholar] [CrossRef]
- Korchowiec, B.; Gorczyca, M.; Rogalska, E.; Regnouf-de-Vains, J.-B.; Mourer, M.; Korchowiec, J. The selective interactions of cationic tetra-p-guanidinoethylcalix [4] arene with lipid membranes: Theoretical and experimental model studies. Soft Matter 2016, 12, 181–190. [Google Scholar] [CrossRef]
- Wieclaw, K.; Korchowiec, B.; Corvis, Y.; Korchowiec, J.; Guermouche, H.; Rogalska, E. Meloxicam and meloxicam-beta-cyclodextrin complex in model membranes: Effects on the properties and enzymatic lipolysis of phospholipid monolayers in relation to anti-inflammatory activity. Langmuir 2009, 25, 1417–1426. [Google Scholar] [CrossRef]
- Marsh, D. Lateral pressure in membranes. Biochim. Biophys. Acta-Rev. Biomembr. 1996, 1286, 183–223. [Google Scholar] [CrossRef]
- Stachowicz-Kusnierz, A.; Korchowiec, B.; Rogalska, E.; Korchowiec, J. The lung surfactant activity probed with molecular dynamics simulations. Adv. Colloid Interface Sci. 2022, 304, 102659. [Google Scholar] [CrossRef] [PubMed]
- Stachowicz-Kusnierz, A.; Trojan, S.; Cwiklik, L.; Korchowiec, B.; Korchowiec, J. Modeling lung surfactant interactions with benzo a pyrene. Chem. Eur. J. 2017, 23, 5307–5316. [Google Scholar] [CrossRef] [PubMed]
- Korchowiec, B.; Korchowiec, J.; Gorczyca, M.; de Vains, J.-B.R.; Rogalska, E. Molecular organization of nalidixate conjugated calixarenes in bacterial model membranes probed by molecular dynamics simulation and Langmuir monolayer studies. J. Phys. Chem. B 2015, 119, 2990–3000. [Google Scholar] [CrossRef]
- Korchowiec, J.; Korchowiec, B.; Priebe, W.; Rogalska, E. DFT study on the selectivity of complexation of metal cations with a dioxadithia crown ether ligand. J. Phys. Chem. A 2008, 112, 13633–13640. [Google Scholar] [CrossRef]
- Davies, J.T.; Rideal, E.K. Interfacial Phenomena, 2nd ed.; Academic Press: Cambridge, MA, USA, 1963; p. 480. [Google Scholar]
- Girardon, M.; Korchowiec, B.; Korchowiec, J.; Rogalska, E.; Canilho, N.; Pasc, A. A way to introducing a hydrophilic bioactive agent into model lipid membranes. The role of cetyl palmitate in the interaction of curcumin with 1,2-dioleoyl-sn-glycero-3-phosphatidylcholine monolayers. J. Mol. Liq. 2020, 308, 113040. [Google Scholar] [CrossRef]
- Korchowiec, B.; Gorczyca, M.; Korchowiec, J.; Rubio-Magnieto, J.; Lotfallah, A.H.; Luis, S.V.; Rogalska, E. Structure—Membrane activity relationship in a family of peptide-based gemini amphiphiles: An insight from experimental and theoretical model systems. Colloids Surf. B Biointerfaces 2016, 146, 54–62. [Google Scholar] [CrossRef]
- Klauda, J.B.; Venable, R.M.; Freites, J.A.; O’Connor, J.W.; Tobias, D.J.; Mondragon-Ramirez, C.; Vorobyov, I.; MacKerell, A.D.; Pastor, R.W. Update of the CHARMM all-atom additive force field for lipids: Validation on six lipid types. J. Phys. Chem. B 2010, 114, 7830–7843. [Google Scholar] [CrossRef] [Green Version]
- Pastor, R.W.; MacKerell, A.D., Jr. Development of the CHARMM Force Field for Lipids. J. Phys. Chem. Lett. 2011, 2, 1526–1532. [Google Scholar] [CrossRef] [Green Version]
- Kwiecinska, K.; Stachowicz-Kusnierz, A.; Jagusiak, A.; Roterman, I.; Korchowiec, J. Impact of doxorubicin on self-organization of Congo red: Quantum chemical calculations and molecular dynamics simulations. ACS Omega 2020, 5, 19377–19384. [Google Scholar] [CrossRef]
- Price, D.J.; Brooks, C.L. A modified TIP3P water potential for simulation with Ewald summation. J. Chem. Phys. 2004, 121, 10096–10103. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.C.; Braun, R.; Wang, W.; Gumbart, J.; Tajkhorshid, E.; Villa, E.; Chipot, C.; Skeel, R.D.; Kale, L.; Schulten, K. Scalable molecular dynamics with NAMD. J. Comput. Chem. 2005, 26, 1781–1802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darden, T.; York, D.; Pedersen, L. Particle mesh Ewald—An N∙log(N) method for Ewald sums in large systems. J. Chem. Phys. 1993, 98, 10089–10092. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. Model. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Leach, A.R. Molecular Modeling. Principles and Applications; Pearson Educations: London, UK, 2001. [Google Scholar]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwiecińska, K.; Stachowicz-Kuśnierz, A.; Korchowiec, B.; Roman, M.; Kwiatek, W.M.; Jagusiak, A.; Roterman, I.; Korchowiec, J. Congo Red as a Supramolecular Carrier System for Doxorubicin: An Approach to Understanding the Mechanism of Action. Int. J. Mol. Sci. 2022, 23, 8935. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23168935
Kwiecińska K, Stachowicz-Kuśnierz A, Korchowiec B, Roman M, Kwiatek WM, Jagusiak A, Roterman I, Korchowiec J. Congo Red as a Supramolecular Carrier System for Doxorubicin: An Approach to Understanding the Mechanism of Action. International Journal of Molecular Sciences. 2022; 23(16):8935. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23168935
Chicago/Turabian StyleKwiecińska, Klaudia, Anna Stachowicz-Kuśnierz, Beata Korchowiec, Maciej Roman, Wojciech M. Kwiatek, Anna Jagusiak, Irena Roterman, and Jacek Korchowiec. 2022. "Congo Red as a Supramolecular Carrier System for Doxorubicin: An Approach to Understanding the Mechanism of Action" International Journal of Molecular Sciences 23, no. 16: 8935. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23168935