
Palmitoylation of Voltage-Gated Ion Channels

 , and
, and
Abstract
:1. Introduction
2. Protein Acylation
- S-palmitoylation or thioesterification is the major form of protein palmitoylation and consists of the addition of a palmitic acid (C16:0) to cysteines via a thioester bond [7].
3. S-Palmitoylation of Ion Channels
3.1. Palmitoylation of Voltage-Gated Ion Channels
3.1.1. Voltage-Gated Sodium Channels
3.1.2. Large Conductance Calcium-Activated Potassium (BK) Channels
3.1.3. Voltage-Dependent Potassium Channels
4. Palmitoylation of Channelosomes
4.1. Palmitoylation Affects Membrane Association and Protein Stability
4.2. Palmitoylation Affects Electrophysiological Properties
5. Interplay with Other Posttranslational Modifications
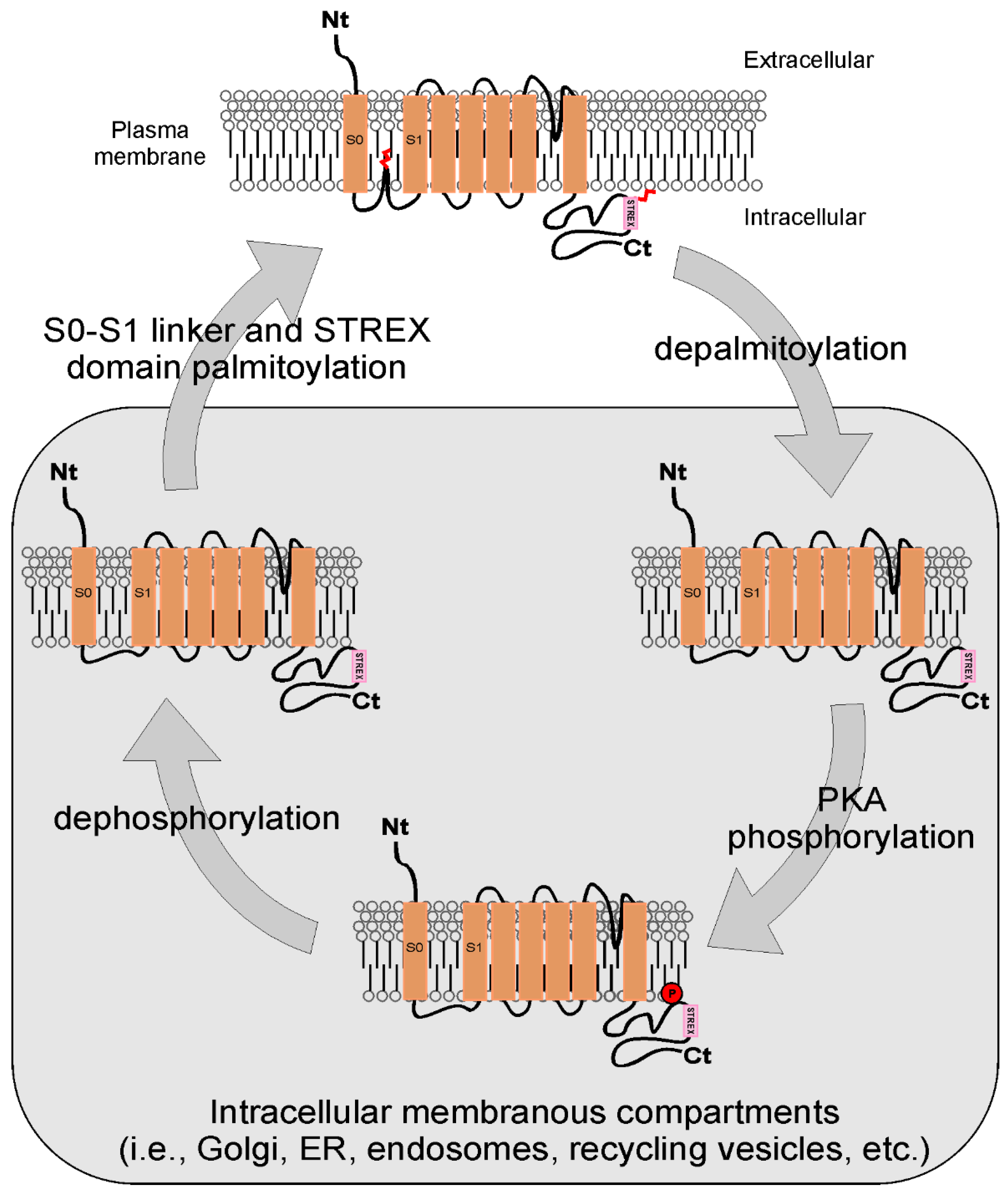
5.1. Phosphorylation
5.2. N-myristoylation and Polyisoprenilation
6. VGIC Palmitoylation and Diseases
6.1. Palmitoylation in Diabetes
6.2. Palmitoylation and BK/Kv Channel-Associated Disorders
6.3. Alzheimer’s and Cystic Fibrosis Diseases
6.4. Mitochondrial Activity and Cancer Development
7. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meinnel, T.; Giglione, C. Protein lipidation meets proteomics. Front. Biosci. 2008, 13, 6326–6340. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, L.H.; Shipston, M.J. The Physiology of Protein S-acylation. Physiol. Rev. 2015, 95, 341–376. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Zhang, X.; Chen, X.; Aramsangtienchai, P.; Tong, Z.; Lin, H. Protein Lipidation: Occurrence, Mechanisms, Biological Functions, and Enabling Technologies. Chem. Rev. 2018, 118, 919–988. [Google Scholar] [CrossRef] [PubMed]
- Nadolski, M.J.; Linder, M.E. Protein lipidation. FEBS J. 2007, 274, 5202–5210. [Google Scholar] [CrossRef] [PubMed]
- Linder, M.; Deschenes, R. Protein palmitoylation. Methods 2006, 40, 125–126. [Google Scholar] [CrossRef]
- Shipston, M.J. Ion Channel Regulation by Protein Palmitoylation. J. Biol. Chem. 2011, 286, 8709–8716. [Google Scholar] [CrossRef] [PubMed]
- Smotrys, J.E.; Linder, M.E. Palmitoylation of Intracellular Signaling Proteins: Regulation and Function. Annu. Rev. Biochem. 2004, 73, 559–587. [Google Scholar] [CrossRef]
- Yousefi-Salakdeh, E.; Johansson, J.; Strömberg, R. A method for S- and O-palmitoylation of peptides: Synthesis of pulmonary surfactant protein-C models. Biochem. J. 1999, 343, 557–562. [Google Scholar] [CrossRef]
- Zou, C.; Ellis, B.M.; Smith, R.M.; Chen, B.B.; Zhao, Y.; Mallampalli, R.K. Acyl-CoA:Lysophosphatidylcholine Acyltransferase I (Lpcat1) Catalyzes Histone Protein O-Palmitoylation to Regulate mRNA Synthesis. J. Biol. Chem. 2011, 286, 28019–28025. [Google Scholar] [CrossRef]
- Gutierrez, J.A.; Solenberg, P.J.; Perkins, D.R.; Willency, J.A.; Knierman, M.D.; Jin, Z.; Witcher, D.R.; Luo, S.; Onyia, J.E.; Hale, J.E. Ghrelin octanoylation mediated by an orphan lipid transferase. Proc. Natl. Acad. Sci. USA 2008, 105, 6320–6325. [Google Scholar] [CrossRef]
- Pepinsky, R.B.; Zeng, C.; Wen, D.; Rayhorn, P.; Baker, D.P.; Williams, K.P.; Bixler, S.A.; Ambrose, C.M.; Garber, E.A.; Miatkowski, K.; et al. Identification of a Palmitic Acid-modified Form of Human Sonic hedgehog. J. Biol. Chem. 1998, 273, 14037–14045. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Bachschmid, M.M.; Costello, C.E.; Lin, C. S- to N-Palmitoyl Transfer During Proteomic Sample Preparation. J. Am. Soc. Mass Spectrom. 2016, 27, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Fukata, Y.; Fukata, M. Protein palmitoylation in neuronal development and synaptic plasticity. Nat. Rev. Neurosci. 2010, 11, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.; Walder, K.; Puri, B.; Berk, M.; Maes, M. The Deleterious Effects of Oxidative and Nitrosative Stress on Palmitoylation, Membrane Lipid Rafts and Lipid-Based Cellular Signalling: New Drug Targets in Neuroimmune Disorders. Mol. Neurobiol. 2016, 53, 4638–4658. [Google Scholar] [CrossRef]
- Ohno, Y.; Kihara, A.; Sano, T.; Igarashi, Y. Intracellular localization and tissue-specific distribution of human and yeast DHHC cysteine-rich domain-containing proteins. Biochim. Biophys. Acta 2006, 1761, 474–483. [Google Scholar] [CrossRef]
- Greaves, J.; Carmichael, J.A.; Chamberlain, L.H. The palmitoyl transferase DHHC2 targets a dynamic membrane cycling pathway: Regulation by a C-terminal domain. Mol. Biol. Cell 2011, 22, 1887–1895. [Google Scholar] [CrossRef]
- Laude, A.J.; Prior, I.A. Palmitoylation and localisation of RAS isoforms are modulated by the hypervariable linker domain. J. Cell Sci. 2008, 121, 421–427. [Google Scholar] [CrossRef]
- Greaves, J.; Chamberlain, L.H. Differential palmitoylation regulates intracellular patterning of SNAP25. J. Cell Sci. 2011, 124, 1351–1360. [Google Scholar] [CrossRef]
- Qanbar, R.; Bouvier, M. Role of palmitoylation/depalmitoylation reactions in G-protein-coupled receptor function. Pharmacol. Ther. 2003, 97, 1–33. [Google Scholar] [CrossRef]
- Greaves, J.; Gorleku, O.A.; Salaun, C.; Chamberlain, L.H. Palmitoylation of the SNAP25 Protein Family: Specificity and Regulation by Dhhc Palmitoyl Transferases. J. Biol. Chem. 2010, 285, 24629–24638. [Google Scholar] [CrossRef]
- Goddard, A.D.; Watts, A. Regulation of G protein-coupled receptors by palmitoylation and cholesterol. BMC Biol. 2012, 10, 27. [Google Scholar] [CrossRef]
- Lemonidis, K.; Gorleku, O.A.; Sanchez-Perez, M.C.; Grefen, C.; Chamberlain, L.H. The Golgi S-acylation machinery comprises zDHHC enzymes with major differences in substrate affinity and S-acylation activity. Mol. Biol. Cell 2014, 25, 3870–3883. [Google Scholar] [CrossRef] [PubMed]
- Martin, B.R.; Cravatt, B.F. Large-scale profiling of protein palmitoylation in mammalian cells. Nat. Methods 2009, 6, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Chisari, M.; Maktabi, M.H.; Sobieski, C.; Zhou, H.; Konopko, A.M.; Martin, B.R.; Mennerick, S.J.; Blumer, K.J. A Mechanism Regulating G Protein-coupled Receptor Signaling That Requires Cycles of Protein Palmitoylation and Depalmitoylation. J. Biol. Chem. 2014, 289, 6249–6257. [Google Scholar] [CrossRef] [PubMed]
- Sanders, S.S.; Martin, D.D.O.; Butland, S.L.; Lavallée-Adam, M.; Calzolari, D.; Kay, C.; Yates, J.R.; Hayden, M.R. Curation of the Mammalian Palmitoylome Indicates a Pivotal Role for Palmitoylation in Diseases and Disorders of the Nervous System and Cancers. PLoS Comput. Biol. 2015, 11, e1004405. [Google Scholar] [CrossRef]
- Wu, Z.; Tan, R.; Zhu, L.; Yao, P.; Hu, Q. Protein S-Palmitoylation and Lung Diseases. Adv. Exp. Med. Biol. 2021, 1304, 165–186. [Google Scholar] [CrossRef]
- Young, F.B.; Butland, S.L.; Sanders, S.S.; Sutton, L.M.; Hayden, M.R. Putting proteins in their place: Palmitoylation in Huntington disease and other neuropsychiatric diseases. Prog. Neurobiol. 2012, 97, 220–238. [Google Scholar] [CrossRef]
- Milligan, G.; Parenti, M.; Magee, A.I. The dynamic role of palmitoylation in signal transduction. Trends Biochem. Sci. 1995, 20, 181–187. [Google Scholar] [CrossRef]
- Thompson, J.W.; Forrester, M.T.; Moseley, M.A.; Foster, M.W. Solid-phase capture for the detection and relative quantification of S-nitrosoproteins by mass spectrometry. Methods 2013, 62, 130–137. [Google Scholar] [CrossRef]
- Drisdel, R.C.; Green, W.N. Labeling and quantifying sites of protein palmitoylation. BioTechniques 2004, 36, 276–285. [Google Scholar] [CrossRef]
- Drisdel, R.C.; Alexander, J.K.; Sayeed, A.; Green, W.N. Assays of protein palmitoylation. Methods 2006, 40, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Robinson, C.V.; Rohacs, T.; Hansen, S.B. Tools for Understanding Nanoscale Lipid Regulation of Ion Channels. Trends Biochem. Sci. 2019, 44, 795–806. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, C.M.; Hille, B. Voltage-Gated Ion Channels and Electrical Excitability. Neuron 1998, 20, 371–380. [Google Scholar] [CrossRef]
- Shipston, M.J. Ion channel regulation by protein S-acylation. J. Gen. Physiol. 2014, 143, 659–678. [Google Scholar] [CrossRef] [PubMed]
- Kashlan, O.B.; Kleyman, T.R. ENaC structure and function in the wake of a resolved structure of a family member. Am. J. Physiol. Ren. Physiol. 2011, 301, F684–F696. [Google Scholar] [CrossRef]
- Mueller, G.M.; Maarouf, A.B.; Kinlough, C.L.; Sheng, N.; Kashlan, O.B.; Okumura, S.; Luthy, S.; Kleyman, T.R.; Hughey, R.P. Cys Palmitoylation of the β Subunit Modulates Gating of the Epithelial Sodium Channel. J. Biol. Chem. 2010, 285, 30453–30462. [Google Scholar] [CrossRef]
- Mukherjee, A.; Mueller, G.M.; Kinlough, C.L.; Sheng, N.; Wang, Z.; Mustafa, S.A.; Kashlan, O.B.; Kleyman, T.R.; Hughey, R.P. Cysteine Palmitoylation of the γ Subunit Has a Dominant Role in Modulating Activity of the Epithelial Sodium Channel. J. Biol. Chem. 2014, 289, 14351–14359. [Google Scholar] [CrossRef]
- Hayashi, T.; Rumbaugh, G.; Huganir, R.L. Differential Regulation of AMPA Receptor Subunit Trafficking by Palmitoylation of Two Distinct Sites. Neuron 2005, 47, 709–723. [Google Scholar] [CrossRef]
- DeSouza, S.; Fu, J.; States, B.A.; Ziff, E.B. Differential Palmitoylation Directs the AMPA Receptor-Binding Protein ABP to Spines or to Intracellular Clusters. J. Neurosci. 2002, 22, 3493–3503. [Google Scholar] [CrossRef]
- Steinke, K.V.; Gorinski, N.; Wojciechowski, D.; Todorov, V.; Guseva, D.; Ponimaskin, E.; Fahlke, C.; Fischer, M. Human CLC-K Channels Require Palmitoylation of Their Accessory Subunit Barttin to Be Functional. J. Biol. Chem. 2015, 290, 17390–17400. [Google Scholar] [CrossRef]
- Gorinski, N.; Wojciechowski, D.; Guseva, P.D.; Galil, D.A.; Mueller, F.E.; Wirth, A.; Thiemann, S.; Zeug, A.; Schmidt, S.; Zareba-Kozioł, M.; et al. DHHC7-mediated palmitoylation of the accessory protein barttin critically regulates the functions of ClC-K chloride channels. J. Biol. Chem. 2020, 295, 5970–5983. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Nishikawa, K.; Hiroaki, Y.; Fujiyoshi, Y. Formation of aquaporin-4 arrays is inhibited by palmitoylation of N-terminal cysteine residues. Biochim. Biophys. Acta 2008, 1778, 1181–1189. [Google Scholar] [CrossRef]
- Hayashi, T. Conservation and phylogenetic stepwise changes of aquaporin (AQP) 4 palmitoylation in vertebrate evolution. Neurotransmitter 2017, 4, e1608. [Google Scholar] [CrossRef]
- Gök, C.; Plain, F.; Robertson, A.D.; Howie, J.; Baillie, G.S.; Fraser, N.J.; Fuller, W. Dynamic Palmitoylation of the Sodium-Calcium Exchanger Modulates Its Structure, Affinity for Lipid-Ordered Domains, and Inhibition by XIP. Cell Rep. 2020, 31, 107697. [Google Scholar] [CrossRef]
- Gök, C.; Main, A.; Gao, X.; Kerekes, Z.; Plain, F.; Kuo, C.-W.; Robertson, A.D.; Fraser, N.J.; Fuller, W. Insights into the molecular basis of the palmitoylation and depalmitoylation of NCX1. Cell Calcium 2021, 97, 102408. [Google Scholar] [CrossRef] [PubMed]
- Bezanilla, F. Voltage-Gated Ion Channels. IEEE Trans. NanoBiosci. 2005, 4, 34–48. [Google Scholar] [CrossRef]
- Capera, J.; Serrano-Novillo, C.; Navarro-Perez, M.; Cassinelli, S.; Felipe, A. The Potassium Channel Odyssey: Mechanisms of Traffic and Membrane Arrangement. Int. J. Mol. Sci. 2019, 20, 734. [Google Scholar] [CrossRef]
- Jiang, Y.; Lee, A.; Chen, J.; Cadene, M.; Chait, B.T.; MacKinnon, R. The open pore conformation of potassium channels. Nature 2002, 417, 523–526. [Google Scholar] [CrossRef]
- Coleman, S.K.; Newcombe, J.; Pryke, J.; Dolly, J.O. Subunit composition of Kv1 channels in human CNS. J. Neurochem. 1999, 73, 849–858. [Google Scholar] [CrossRef]
- Beacham, D.; Ahn, M.; Catterall, W.A.; Scheuer, T. Sites and Molecular Mechanisms of Modulation of Na(V)1.2 Channels by Fyn Tyrosine Kinase. J. Neurosci. 2007, 27, 11543–11551. [Google Scholar] [CrossRef]
- Schmidt, J.W.; Catterall, W.A. Palmitylation, sulfation, and glycosylation of the alpha subunit of the sodium channel. Role of post-translational modifications in channel assembly. J. Biol. Chem. 1987, 262, 13713–13723. [Google Scholar] [CrossRef]
- Bosmans, F.; Milescu, M.; Swartz, K.J. Palmitoylation influences the function and pharmacology of sodium channels. Proc. Natl. Acad. Sci. USA 2011, 108, 20213–20218. [Google Scholar] [CrossRef] [PubMed]
- Pei, Z.; Xiao, Y.; Meng, J.; Hudmon, A.; Cummins, T.R. Cardiac sodium channel palmitoylation regulates channel availability and myocyte excitability with implications for arrhythmia generation. Nat. Commun. 2016, 7, 12035. [Google Scholar] [CrossRef]
- Pan, Y.; Xiao, Y.; Pei, Z.; Cummins, T.R. S-Palmitoylation of the sodium channel Nav1.6 regulates its activity and neuronal excitability. J. Biol. Chem. 2020, 295, 6151–6164. [Google Scholar] [CrossRef] [PubMed]
- Shipston, M.J. S-acylation dependent post-translational cross-talk regulates large conductance calcium- and voltage- activated potassium (BK) channels. Front. Physiol. 2014, 5, 281. [Google Scholar] [CrossRef]
- Jeffries, O.; Geiger, N.; Rowe, I.C.M.; Tian, L.; McClafferty, H.; Chen, L.; Bi, D.; Knaus, H.G.; Ruth, P.; Shipston, M.J. Palmitoylation of the S0-S1 Linker Regulates Cell Surface Expression of Voltage- and Calcium-activated Potassium (BK) Channels. J. Biol. Chem. 2010, 285, 33307–33314. [Google Scholar] [CrossRef]
- Zhou, X.; Wulfsen, I.; Korth, M.; McClafferty, H.; Lukowski, R.; Shipston, M.; Ruth, P.; Dobrev, D.; Wieland, T. Palmitoylation and Membrane Association of the Stress Axis Regulated Insert (STREX) Controls BK Channel Regulation by Protein Kinase C. J. Biol. Chem. 2012, 287, 32161–32171. [Google Scholar] [CrossRef]
- Jeffries, O.; Tian, L.; McClafferty, H.; Shipston, M.J. An Electrostatic Switch Controls Palmitoylation of the Large Conductance Voltage- and Calcium-activated Potassium (BK) Channel. J. Biol. Chem. 2012, 287, 1468–1477. [Google Scholar] [CrossRef]
- Tian, L.; McClafferty, H.; Jeffries, O.; Shipston, M.J. Multiple Palmitoyltransferases Are Required for Palmitoylation-dependent Regulation of Large Conductance Calcium- and Voltage-activated Potassium Channels. J. Biol. Chem. 2010, 285, 23954–23962. [Google Scholar] [CrossRef]
- Tian, L.; McClafferty, H.; Knaus, H.-G.; Ruth, P.; Shipston, M.J. Distinct Acyl Protein Transferases and Thioesterases Control Surface Expression of Calcium-activated Potassium Channels. J. Biol. Chem. 2012, 287, 14718–14725. [Google Scholar] [CrossRef]
- Attali, B.; Chandy, K.G.; Giese, M.H.; Grissmer, S.; Gutman, G.A.; Jan, L.Y.; Lazdunski, M.; Mckinnon, D.; Nerbonne, J.; Pardo, L.A.; et al. Voltage-gated potassium channels (Kv) in GtoPdb v.2021.3. IUPHAR/BPS Guid. Pharmacol. CITE 2021, 2021. [Google Scholar] [CrossRef]
- Sanders, S.S.; Hernandez, L.M.; Soh, H.; Karnam, S.; Walikonis, R.S.; Tzingounis, A.V.; Thomas, G.M. The palmitoyl acyltransferase ZDHHC14 controls Kv1-family potassium channel clustering at the axon initial segment. eLife 2020, 9, e56058. [Google Scholar] [CrossRef] [PubMed]
- Gubitosi-Klug, R.A.; Mancuso, D.J.; Gross, R.W. The human Kv1.1 channel is palmitoylated, modulating voltage sensing: Identification of a palmitoylation consensus sequence. Proc. Natl. Acad. Sci. USA 2005, 102, 5964–5968. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Foster, K.; Li, Q.; Martens, J.R. S-acylation regulates Kv1.5 channel surface expression. Am. J. Physiol. Cell Physiol. 2007, 293, C152–C161. [Google Scholar] [CrossRef] [PubMed]
- Jindal, H.K.; Folco, E.J.; Liu, G.X.; Koren, G. Posttranslational modification of voltage-dependent potassium channel Kv1.5: COOH-terminal palmitoylation modulates its biological properties. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, H2012–H2021. [Google Scholar] [CrossRef]
- Thomas, G.M.; Hayashi, T.; Chiu, S.-L.; Chen, C.-M.; Huganir, R.L. Palmitoylation by DHHC5/8 Targets GRIP1 to Dendritic Endosomes to Regulate AMPA-R Trafficking. Neuron 2012, 73, 482–496. [Google Scholar] [CrossRef]
- Dong, H.; Zhang, P.; Song, I.; Petralia, R.S.; Liao, D.; Huganir, R.L. Characterization of the Glutamate Receptor-Interacting Proteins GRIP1 and GRIP2. J. Neurosci. 1999, 19, 6930–6941. [Google Scholar] [CrossRef]
- Sheng, M.; Sala, C. PDZ Domains and the Organization of Supramolecular Complexes. Annu. Rev. Neurosci. 2001, 24, 1–29. [Google Scholar] [CrossRef]
- Setou, M.; Seog, D.-H.; Tanaka, Y.; Kanai, Y.; Takei, Y.; Kawagishi, M.; Hirokawa, N. Glutamate-receptor-interacting protein GRIP1 directly steers kinesin to dendrites. Nature 2002, 417, 83–87. [Google Scholar] [CrossRef]
- Brigidi, G.S.; Santyr, B.; Shimell, J.; Jovellar, B.; Bamji, S.X. Activity-regulated trafficking of the palmitoyl-acyl transferase DHHC5. Nat. Commun. 2015, 6, 8200. [Google Scholar] [CrossRef]
- McEwen, D.P.; Isom, L.L. Heterophilic Interactions of Sodium Channel β1 Subunits with Axonal and Glial Cell Adhesion Molecules. J. Biol. Chem. 2004, 279, 52744–52752. [Google Scholar] [CrossRef] [PubMed]
- Bouza, A.A.; Philippe, J.M.; Edokobi, N.; Pinsky, A.M.; Offord, J.; Calhoun, J.D.; Lopez-Florán, M.; Lopez-Santiago, L.F.; Jenkins, P.M.; Isom, L.L. Sodium channel β1 subunits are post-translationally modified by tyrosine phosphorylation, S-palmitoylation, and regulated intramembrane proteolysis. J. Biol. Chem. 2020, 295, 10380–10393. [Google Scholar] [CrossRef] [PubMed]
- Long, J.-F.; Tochio, H.; Wang, P.; Fan, J.-S.; Sala, C.; Niethammer, M.; Sheng, M.; Zhang, M. Supramodular structure and synergistic target binding of the N-terminal tandem PDZ domains of PSD-95. J. Mol. Biol. 2003, 327, 203–214. [Google Scholar] [CrossRef]
- Zhang, J.; Lewis, S.M.; Kuhlman, B.; Lee, A.L. Supertertiary Structure of the MAGUK Core from PSD-95. Structure 2013, 21, 402–413. [Google Scholar] [CrossRef]
- Martínez-Mármol, R.; Styrczewska, K.; Pérez-Verdaguer, M.; Vallejo-Gracia, A.; Comes, N.; Sorkin, A.; Felipe, A. Ubiquitination mediates Kv1.3 endocytosis as a mechanism for protein kinase C-dependent modulation. Sci. Rep. 2017, 7, 42395. [Google Scholar] [CrossRef]
- Szilagyi, O.; Boratko, A.; Panyi, G.; Hajdu, P. The role of PSD-95 in the rearrangement of Kv1.3 channels to the immunological synapse. Pflügers Arch. Eur. J. Physiol. 2013, 465, 1341–1353. [Google Scholar] [CrossRef]
- Topinka, J.R.; Bredt, D.S. N-Terminal Palmitoylation of PSD-95 Regulates Association with Cell Membranes and Interaction with K+ Channel Kv1.4. Neuron 1998, 20, 125–134. [Google Scholar] [CrossRef]
- El-Husseini, A.E.; Craven, S.E.; Chetkovich, D.M.; Firestein, B.; Schnell, E.; Aoki, C.; Bredt, D.S. Dual Palmitoylation of Psd-95 Mediates Its Vesiculotubular Sorting, Postsynaptic Targeting, and Ion Channel Clustering. J. Cell Biol. 2000, 148, 159–172. [Google Scholar] [CrossRef]
- Roig, S.R.; Cassinelli, S.; Navarro-Pérez, M.; Pérez-Verdaguer, M.; Estadella, I.; Capera, J.; Felipe, A. S-acylation-dependent membrane microdomain localization of the regulatory Kvβ2.1 subunit. Cell. Mol. Life Sci. 2022, 79, 230. [Google Scholar] [CrossRef]
- Uebele, V.N.; England, S.K.; Chaudhary, A.; Tamkun, M.M.; Snyders, D.J. Functional Differences in Kv1.5 Currents Expressed in Mammalian Cell Lines Are Due to the Presence of Endogenous Kvβ2.1 Subunits. J. Biol. Chem. 1996, 271, 2406–2412. [Google Scholar] [CrossRef]
- Martínez-Mármol, R.; Villalonga, N.; Solé, L.; Vicente, R.; Tamkun, M.M.; Soler, C.; Felipe, A. Multiple Kv1.5 targeting to membrane surface microdomains. J. Cell. Physiol. 2008, 217, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Heinemann, S.H.; Rettig, J.; Graack, H.R.; Pongs, O. Functional characterization of Kv channel beta-subunits from rat brain. J. Physiol. 1996, 493, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Grande, M.; Suàrez, E.; Vicente, R.; Canto, C.; Coma, M.; Tamkun, M.M.; Zorzano, A.; Gumà, A.; Felipe, A. Voltage-dependent K+channel β subunits in muscle: Differential regulation during postnatal development and myogenesis. J. Cell. Physiol. 2003, 195, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Takimoto, K.; Yang, E.-K.; Conforti, L. Palmitoylation of KChIP Splicing Variants Is Required for Efficient Cell Surface Expression of Kv4.3 Channels. J. Biol. Chem. 2002, 277, 26904–26911. [Google Scholar] [CrossRef]
- Murthy, A.; Workman, S.W.; Jiang, M.; Hu, J.; Sifa, I.; Bernas, T.; Tang, W.; Deschenes, I.; Tseng, G.-N. Dynamic palmitoylation regulates trafficking of K channel interacting protein 2 (KChIP2) across multiple subcellular compartments in cardiac myocytes. J. Mol. Cell. Cardiol. 2019, 135, 1–9. [Google Scholar] [CrossRef]
- Vassar, R.; Kandalepas, P.C. The β-secretase enzyme BACE1 as a therapeutic target for Alzheimer’s disease. Alzheimer’s Res. Ther. 2011, 3, 20. [Google Scholar] [CrossRef]
- Hernández, C.C.; Zaika, O.; Tolstykh, G.P.; Shapiro, M.S. Regulation of neural KCNQ channels: Signalling pathways, structural motifs and functional implications. J. Physiol. 2008, 586, 1811–1821. [Google Scholar] [CrossRef]
- Wang, H.-S.; Pan, Z.; Shi, W.; Brown, B.S.; Wymore, R.S.; Cohen, I.S.; Dixon, J.E.; McKinnon, D. KCNQ2 and KCNQ3 Potassium Channel Subunits: Molecular Correlates of the M-Channel. Science 1998, 282, 1890–1893. [Google Scholar] [CrossRef]
- Dai, G. Neuronal KCNQ2/3 channels are recruited to lipid raft microdomains by palmitoylation of BACE1. J. Gen. Physiol. 2022, 154, e202112888. [Google Scholar] [CrossRef]
- Chen, L.; Bi, D.; Tian, L.; McClafferty, H.; Steeb, F.; Ruth, P.; Knaus, H.G.; Shipston, M.J. Palmitoylation of the β4-Subunit Regulates Surface Expression of Large Conductance Calcium-activated Potassium Channel Splice Variants. J. Biol. Chem. 2013, 288, 13136–13144. [Google Scholar] [CrossRef]
- Buraei, Z.; Yang, J. Structure and function of the β subunit of voltage-gated Ca2+ channels. Biochim. Biophys. Acta 2012, 1828, 1530–1540. [Google Scholar] [CrossRef] [PubMed]
- Chien, A.J.; Gao, T.; Perez-Reyes, E.; Hosey, M.M. Membrane Targeting of L-type Calcium Channels. Role of palmitoylation in the subcellular localization of the β2a subunit. J. Biol. Chem. 1998, 273, 23590–23597. [Google Scholar] [CrossRef] [PubMed]
- Qin, N.; Platano, D.; Olcese, R.; Costantin, J.L.; Stefani, E.; Birnbaumer, L. Unique regulatory properties of the type 2a Ca2+ channel β subunit caused by palmitoylation. Proc. Natl. Acad. Sci. USA 1998, 95, 4690–4695. [Google Scholar] [CrossRef] [PubMed]
- Hurley, J.H.; Cahill, A.L.; Currie, K.P.; Fox, A.P. The role of dynamic palmitoylation in Ca2+ channel inactivation. Proc. Natl. Acad. Sci. USA 2000, 97, 9293–9298. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.-Q.; Martinez-Ortiz, W.; Hwang, J.; Fan, X.; Cardozo, T.J.; Coetzee, W.A. Palmitoylation of the KATP channel Kir6.2 subunit promotes channel opening by regulating PIP2 sensitivity. Proc. Natl. Acad. Sci. USA 2020, 117, 10593–10602. [Google Scholar] [CrossRef]
- Ardito, F.; Giuliani, M.; Perrone, D.; Troiano, G.; Lo Muzio, L. The crucial role of protein phosphorylation in cell signaling and its use as targeted therapy (Review). Int. J. Mol. Med. 2017, 40, 271–280. [Google Scholar] [CrossRef]
- Samanta, A.; Hughes, T.E.T.; Moiseenkova-Bell, V.Y. Transient Receptor Potential (TRP) Channels. Subcell Biochem. 2018, 87, 141–165. [Google Scholar] [CrossRef]
- Zheng, W.; Yang, J.; Beauchamp, E.; Cai, R.; Hussein, S.; Hofmann, L.; Li, Q.; Flockerzi, V.; Berthiaume, L.G.; Tang, J.; et al. Regulation of TRPP3 Channel Function by N-terminal Domain Palmitoylation and Phosphorylation. J. Biol. Chem. 2016, 291, 25678–25691. [Google Scholar] [CrossRef]
- Mouillac, B.; Caron, M.; Bonin, H.; Dennis, M.; Bouvier, M. Agonist-modulated palmitoylation of beta 2-adrenergic receptor in Sf9 cells. J. Biol. Chem. 1992, 267, 21733–21737. [Google Scholar] [CrossRef]
- Adachi, N.; Hess, D.T.; McLaughlin, P.; Stamler, J.S. S-Palmitoylation of a Novel Site in the β2-Adrenergic Receptor Associated with a Novel Intracellular Itinerary. J. Biol. Chem. 2016, 291, 20232–20246. [Google Scholar] [CrossRef]
- Moffett, S.; Mouillac, B.; Bonin, H.; Bouvier, M. Altered phosphorylation and desensitization patterns of a human beta 2-adrenergic receptor lacking the palmitoylated Cys341. EMBO J. 1993, 12, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Solis, G.P.; Kazemzadeh, A.; Abrami, L.; Valnohova, J.; Alvarez, C.; van der Goot, F.G.; Katanaev, V.L. Local and substrate-specific S-palmitoylation determines subcellular localization of Gαo. Nat. Commun. 2022, 13, 2072. [Google Scholar] [CrossRef]
- Tian, L.; Jeffries, O.; McClafferty, H.; Molyvdas, A.; Rowe, I.C.M.; Saleem, F.; Chen, L.; Greaves, J.; Chamberlain, L.H.; Knaus, H.-G.; et al. Palmitoylation gates phosphorylation-dependent regulation of BK potassium channels. Proc. Natl. Acad. Sci. USA 2008, 105, 21006–21011. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Song, Z.-H.; Ying, M.-D.; Zhu, H.; He, Q.-J.; Yang, B.; Cao, J. N-myristoylation: From cell biology to translational medicine. Acta Pharmacol. Sin. 2020, 41, 1005–1015. [Google Scholar] [CrossRef] [PubMed]
- Resh, M.D. Use of analogs and inhibitors to study the functional significance of protein palmitoylation. Methods 2006, 40, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Nitabach, M.N.; Llamas, D.A.; Thompson, I.J.; Collins, K.A.; Holmes, T.C. Phosphorylation-Dependent and Phosphorylation-Independent Modes of Modulation of Shaker Family Voltage-Gated Potassium Channels by Src Family Protein Tyrosine Kinases. J. Neurosci. 2002, 22, 7913–7922. [Google Scholar] [CrossRef]
- Pochynyuk, O.; Stockand, J.D.; Staruschenko, A. Ion Channel Regulation by Ras, Rho, and Rab Small GTPases. Exp. Biol. Med. 2007, 232, 1258–1265. [Google Scholar] [CrossRef]
- Wright, L.P.; Philips, M.R. Thematic review series: Lipid Posttranslational Modifications CAAX modification and membrane targeting of Ras. J. Lipid Res. 2006, 47, 883–891. [Google Scholar] [CrossRef]
- Hancock, J.F.; Magee, A.I.; Childs, J.E.; Marshall, C.J. All ras proteins are polyisoprenylated but only some are palmitoylated. Cell 1989, 57, 1167–1177. [Google Scholar] [CrossRef]
- Olofsson, C.S.; Salehi, A.; Holm, C.; Rorsman, P. Palmitate increases L-type Ca2+ currents and the size of the readily releasable granule pool in mouse pancreatic β-cells. J. Physiol. 2004, 557, 935–948. [Google Scholar] [CrossRef]
- Bähring, R. Kv channel-interacting proteins as neuronal and non-neuronal calcium sensors. Channels 2018, 12, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Hoa, N.; Myers, M.P.; Douglass, T.G.; Zhang, J.G.; Delgado, C.; Driggers, L.; Callahan, L.L.; VanDeusen, G.; Pham, J.T.H.; Bhakta, N.; et al. Molecular Mechanisms of Paraptosis Induction: Implications for a Non-Genetically Modified Tumor Vaccine. PLoS ONE 2009, 4, e4631. [Google Scholar] [CrossRef] [PubMed]
- Villa, C.; Combi, R. Potassium Channels and Human Epileptic Phenotypes: An Updated Overview. Front. Cell. Neurosci. 2016, 10, 81. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.; Hoa, N.T.; Wilson, Z.; Arismendi-Morillo, G.; Kong, X.-T.; Tajhya, R.B.; Beeton, C.; Jadus, M.R. Big Potassium (BK) ion channels in biology, disease and possible targets for cancer immunotherapy. Int. Immunopharmacol. 2014, 22, 427–443. [Google Scholar] [CrossRef]
- Dixit, G.; Sahu, I.D.; Reynolds, W.D.; Wadsworth, T.M.; Harding, B.D.; Jaycox, C.K.; Dabney-Smith, C.; Sanders, C.R.; Lorigan, G.A. Probing the Dynamics and Structural Topology of the Reconstituted Human KCNQ1 Voltage Sensor Domain (Q1-VSD) in Lipid Bilayers Using Electron Paramagnetic Resonance Spectroscopy. Biochemistry 2019, 58, 965–973. [Google Scholar] [CrossRef]
- Demontis, G.C.; Pezzini, F.; Margari, E.; Bianchi, M.; Longoni, B.; Doccini, S.; Lalowski, M.M.; Santorelli, F.M.; Simonati, A. Electrophysiological Profile Remodeling via Selective Suppression of Voltage-Gated Currents by CLN1/PPT1 Overexpression in Human Neuronal-Like Cells. Front. Cell. Neurosci. 2020, 14, 569598. [Google Scholar] [CrossRef]
- Cho, E.; Park, M. Palmitoylation in Alzheimer’s disease and other neurodegenerative diseases. Pharmacol. Res. 2016, 111, 133–151. [Google Scholar] [CrossRef]
- Kelleher, R.J., 3rd; Shen, J. Presenilin-1 mutations and Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2017, 114, 629–631. [Google Scholar] [CrossRef]
- McClure, M.L.; Barnes, S.; Brodsky, J.L.; Sorscher, E.J. Trafficking and function of the cystic fibrosis transmembrane conductance regulator: A complex network of posttranslational modifications. Am. J. Physiol. Lung Cell. Mol. Physiol. 2016, 311, L719–L733. [Google Scholar] [CrossRef]
- Fhu, C.W.; Ali, A. Protein Lipidation by Palmitoylation and Myristoylation in Cancer. Front. Cell Dev. Biol. 2021, 9, 673647. [Google Scholar] [CrossRef]
- Spinelli, M.; Fusco, S.; Grassi, C. Nutrient-Dependent Changes of Protein Palmitoylation: Impact on Nuclear Enzymes and Regulation of Gene Expression. Int. J. Mol. Sci. 2018, 19, 3820. [Google Scholar] [CrossRef] [PubMed]
- Planey, S.L.; Keay, S.K.; Zhang, C.-O.; Zacharias, D.A. Palmitoylation of Cytoskeleton Associated Protein 4 by DHHC2 Regulates Antiproliferative Factor-mediated Signaling. Mol. Biol. Cell 2009, 20, 1454–1463. [Google Scholar] [CrossRef] [PubMed]
- Harada, T.; Sada, R.; Osugi, Y.; Matsumoto, S.; Matsuda, T.; Hayashi-Nishino, M.; Nagai, T.; Harada, A.; Kikuchi, A. Palmitoylated CKAP4 regulates mitochondrial functions through an interaction with VDAC2 at ER–mitochondria contact sites. J. Cell Sci. 2020, 133, jcs249045. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Channel Family | Channel | Palmitoylated Residue | Correlated Diseases | Reference |
|---|---|---|---|---|
| Epithelial sodium channels | ENaCs | [34,35,36] | ||
| ATP-sensitive potassium channels | Kir6.2 | Cys166 | [95] | |
| Transient receptor potential (TRP) channels | TRPP3 | Cys38 | [97] | |
| Chloride channels | CFTR | Cys524 | Cystic fibrosis | [119] |
| Voltage-gated calcium channels | Cav1, 2 | [93] | ||
| Large conductance calcium-activated potassium channels | BK | Cys53,54,56 STREX | Cancer CLN1 disease | [56,57] [116] |
| Sodium-calcium exchangers | NCX1 | [42] | ||
| Voltage-gated potassium channels | Kv1.1 Kv1.2 Kv1.4 Kv1.5 | Cys243 | [63,64] [62] [62] [64,65] | |
| Voltage-gated sodium channels | Nav1.2 Nav1.5 Nav1.6 | Cys981 Cys1169-1170 Cys1978 | [50,55] [50,55] [50,55] | |
| Aquaporins | AQ4 | Cys609 | [42,43] | |
| GluR1-4 | AMPARs | [38,39] |
| Protein Family | Regulatory Subunit/Ancillary proteins | Palmitoylated Residue | Correlated Diseases | Reference |
|---|---|---|---|---|
| Glutamate receptor-interacting proteins | GRIP1b/2b | [66,67] | ||
| Clc-k accessory subunit, Barttin, family | BARTTIN | [40,41] | ||
| Cytoskeleton-associated protein 4 family | CKAP4 | Cancer Mitochondria disorders | [122] | |
| Voltage-gated sodium channels | Navβ1 | Cys162 | [72] | |
| Voltage-gated calcium channels | Cavβ2a | Cys3, 4 | Diabetes | [92,94,110] |
| Voltage-gated potassium channels | Kvβ2.1 | Cys51, 212, 248, 301, 311 | [79] | |
| Alzheimer’s | ||||
| Kv channel-interacting proteins | KChIP2 | Long QT syndrome | [85,86] | |
| Cancer | ||||
| β-secretase precursor | BACE1 | Cys474, 478, 482, 485 | Alzheimer’s | [87,118] |
| G protein-coupled receptors | β2AR | Cys265 | [100] | |
| Ras proteins | p21-Ras | Cys188 | Cancer | [107] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cassinelli, S.; Viñola-Renart, C.; Benavente-Garcia, A.; Navarro-Pérez, M.; Capera, J.; Felipe, A. Palmitoylation of Voltage-Gated Ion Channels. Int. J. Mol. Sci. 2022, 23, 9357. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23169357
Cassinelli S, Viñola-Renart C, Benavente-Garcia A, Navarro-Pérez M, Capera J, Felipe A. Palmitoylation of Voltage-Gated Ion Channels. International Journal of Molecular Sciences. 2022; 23(16):9357. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23169357
Chicago/Turabian StyleCassinelli, Silvia, Carla Viñola-Renart, Anna Benavente-Garcia, María Navarro-Pérez, Jesusa Capera, and Antonio Felipe. 2022. "Palmitoylation of Voltage-Gated Ion Channels" International Journal of Molecular Sciences 23, no. 16: 9357. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23169357