
Chronic N-Acetylcysteine Treatment Prevents Amphetamine-Induced Hyperactivity in Heterozygous Disc1 Mutant Mice, a Putative Prodromal Schizophrenia Animal Model


 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Chronic NAC Normalized the Amph-Induced Activity in Het Disc1 Mice
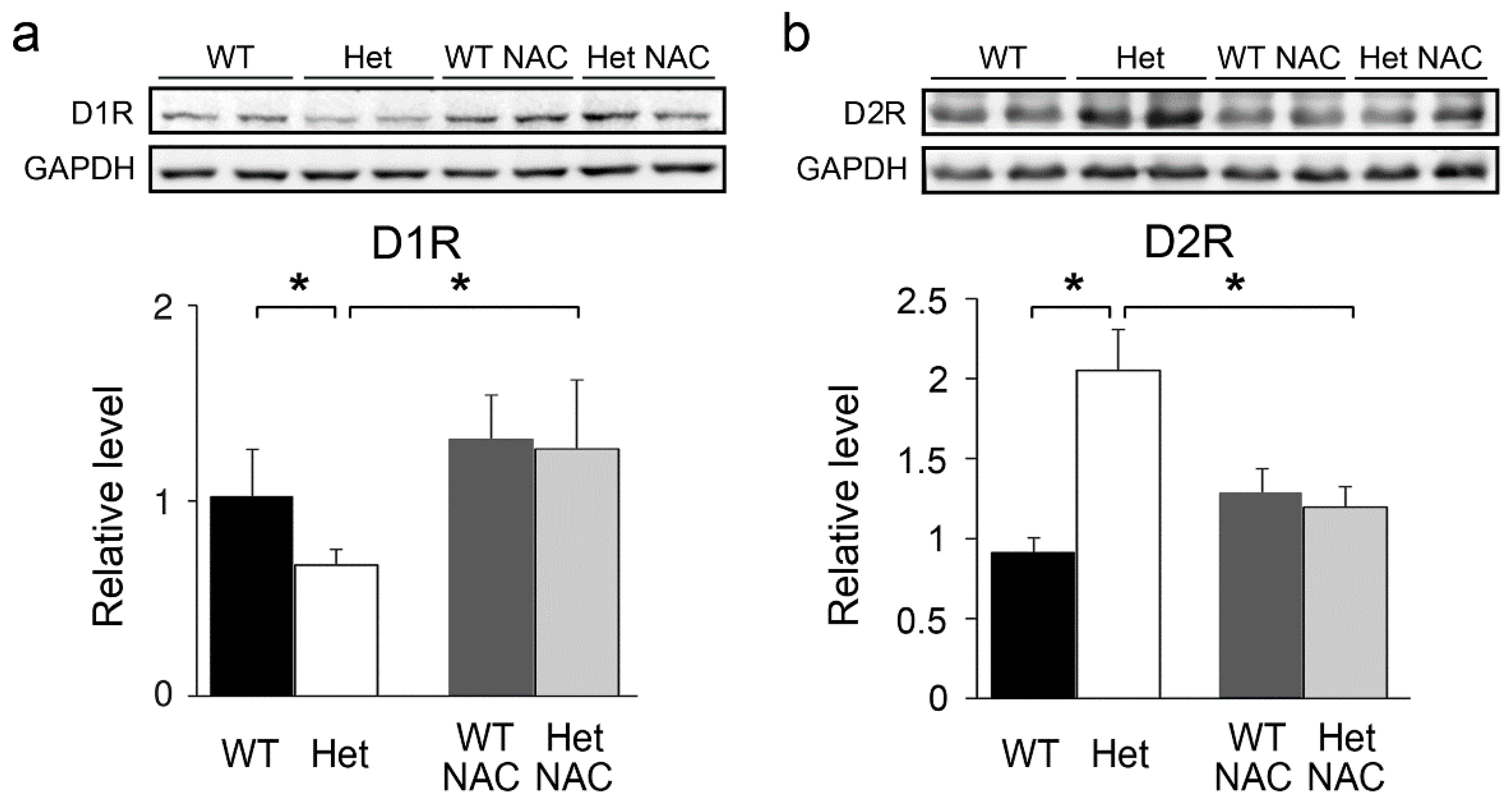
2.2. Chronic NAC Adjusted the Striatum DA System in Het Disc1 Mice
2.3. Chronic NAC Increased the Expression of Striatal GSK3s in Het Disc1 Mice
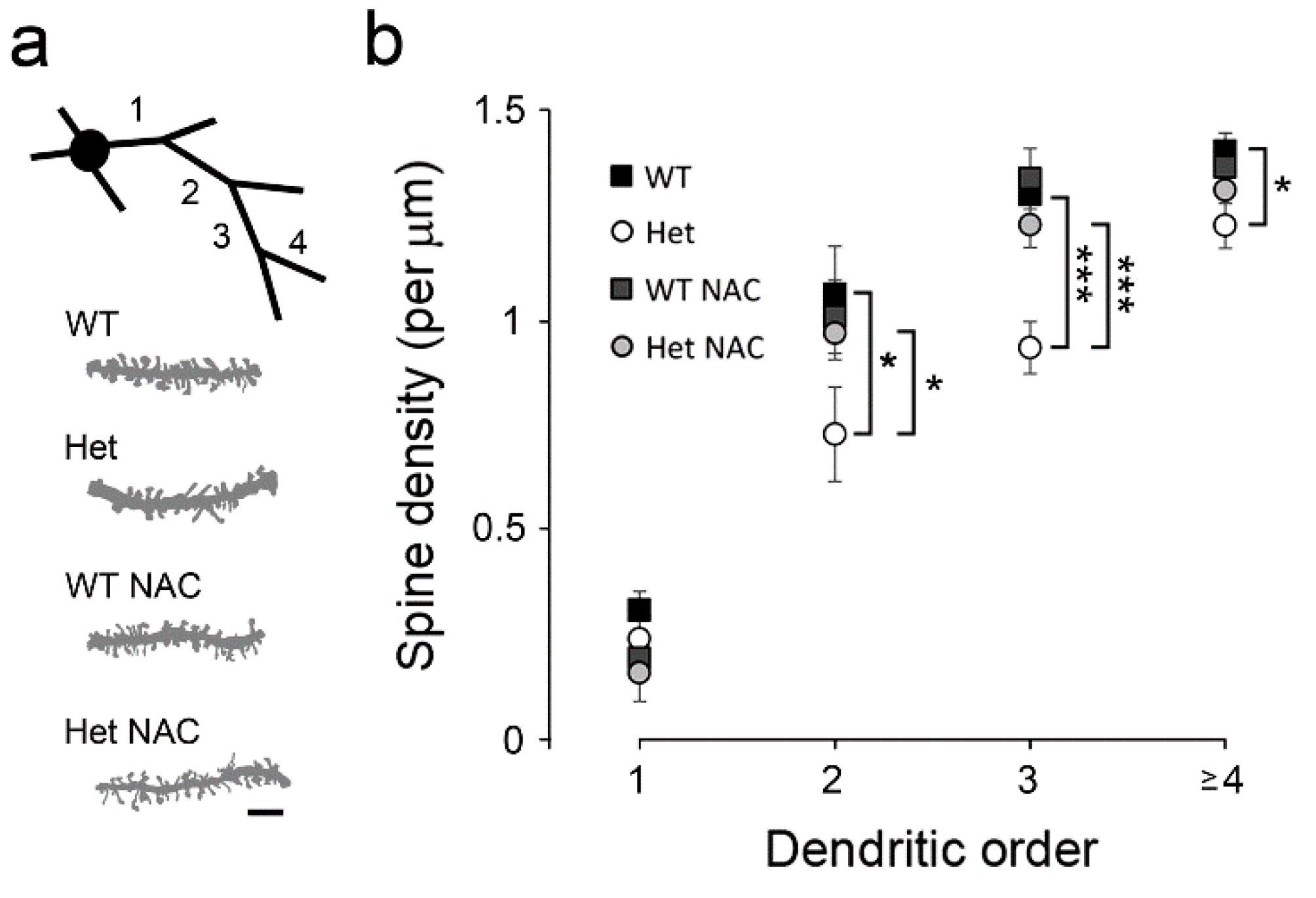
2.4. Chronic NAC Rescued the MSN Dendritic Impairments in Het Disc1 Mice
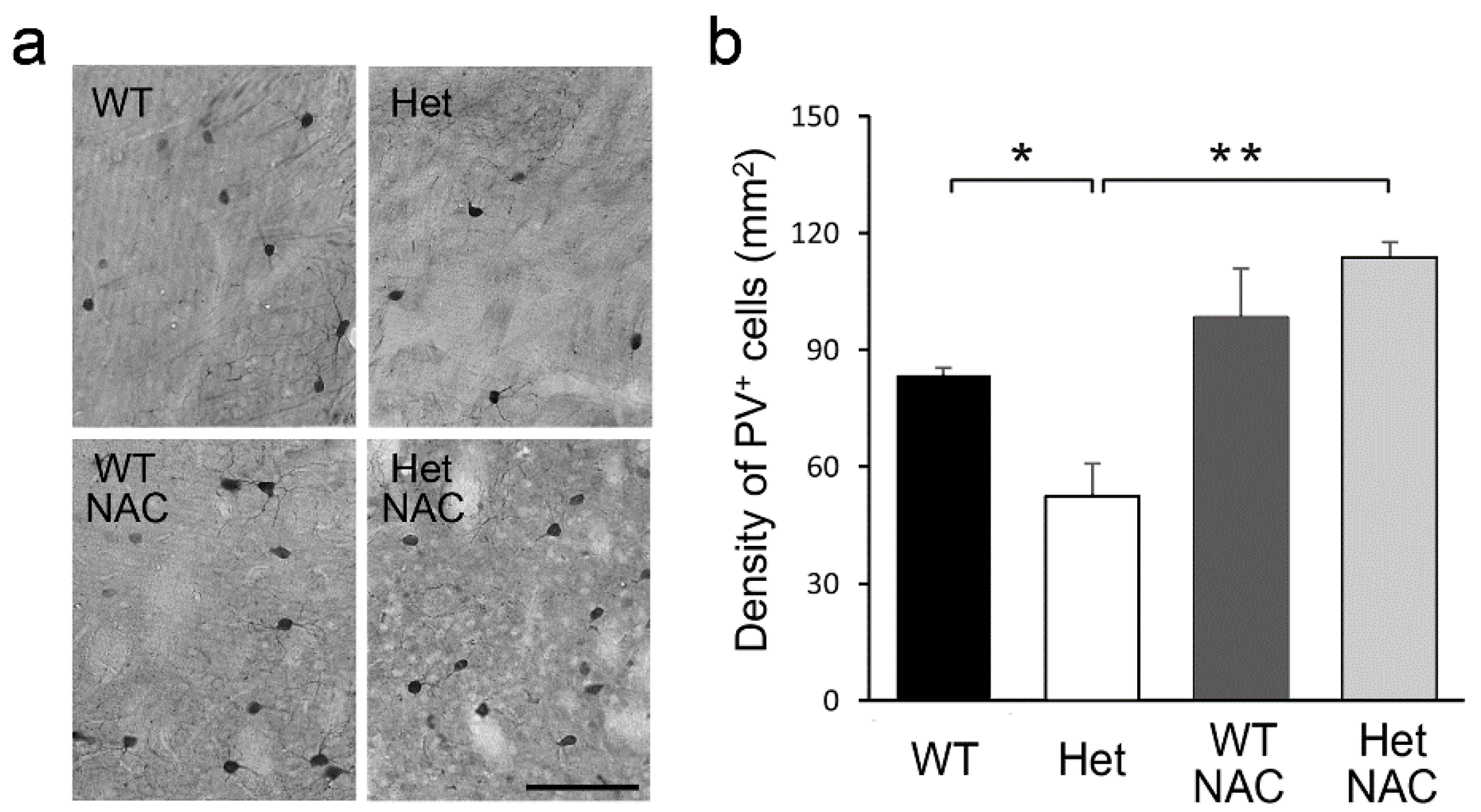
2.5. Chronic NAC Restored Striatal PV Neurons Density in Het Disc1 Mice
3. Discussion
3.1. Animal Models of Prodromal SZ
3.2. Multifaceted Therapeutic Potentials of NAC
3.3. GSK3-Mediated Protective Effects
3.4. Limitations
4. Materials and Methods
4.1. Animals
4.2. NAC Treatment
4.3. Amphetamine Challenge Test
4.4. Western Blot Analysis
4.5. Immunohistochemistry
4.6. Golgi Stain and Morphometric Analyses
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lewis, D.A.; Lieberman, J.A. Catching up on schizophrenia: Natural history and neurobiology. Neuron 2000, 28, 325–334. [Google Scholar] [CrossRef] [Green Version]
- Ross, C.A.; Margolis, R.L.; Reading, S.A.; Pletnikov, M.; Coyle, J.T. Neurobiology of schizophrenia. Neuron 2006, 52, 139–153. [Google Scholar] [CrossRef] [Green Version]
- McGlashan TH, Johannessen JO Early detection and intervention with schizophrenia: Rationale. Schizophr. Bull. 1996, 22, 201–222. [CrossRef]
- Yung, A.R.; McGorry, P.D. The prodromal phase of first-episode psychosis: Past and current conceptualizations. Schizophr. Bull. 1996, 22, 353–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Addington, J.; Heinssen, R. Prediction and prevention of psychosis in youth at clinical high risk. Annu. Rev. Clin. Psychol. 2012, 8, 269–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCutcheon, R.A.; Reis Marques, T.; Howes, O.D. Schizophrenia-An overview. JAMA Psychiatry 2020, 77, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K. Recent advances in the early intervention in schizophrenia: Future direction from preclinical findings. Curr. Psychiatry Rep. 2019, 21, 75. [Google Scholar] [CrossRef]
- Lin, C.H.; Lane, H.Y. Early identification and intervention of schizophrenia: Insight from hypotheses of glutamate dysfunction and oxidative stress. Front. Psychiatry 2019, 10, 93. [Google Scholar] [CrossRef]
- Heinssen, R.K.; Insel, T.R. Preventing the onset of psychosis: Not quite there yet. Schizophr. Bull. 2015, 41, 28–29. [Google Scholar] [CrossRef] [Green Version]
- Wessels, H.; Wagner, M.; Kuhr, K.; Berning, J.; Pützfeld, V.; Janssen, B.; Bottlender, R.; Maurer, K.; Möller, H.J.; Gaebel, W.; et al. Predictors of treatment response to psychological interventions in people at clinical high risk of first-episode psychosis. Early Interv. Psychiatry 2019, 13, 120–127. [Google Scholar] [CrossRef] [Green Version]
- Gordon, J.A.; Heinssen, R.K. Collaborative approaches to the clinical high-risk state: From data to mechanism to intervention. Biol. Psychiatry 2020, 88, 287–288. [Google Scholar] [CrossRef] [PubMed]
- Tenn, C.C.; Fletcher, P.J.; Kapur, S. A putative animal model of the “prodromal” state of schizophrenia. Biol. Psychiatry 2005, 57, 586–593. [Google Scholar] [CrossRef] [PubMed]
- Petty, A.; Cui, X.; Tesiram, Y.; Kirik, D.; Howes, O.; Eyles, D. Enhanced Dopamine in Prodromal Schizophrenia (EDiPS): A new animal model of relevance to schizophrenia. NPJ Schizophr. 2019, 5, 6. [Google Scholar] [CrossRef] [PubMed]
- Simpson, E.H.; Kellendonk, C.; Kandel, E. A possible role for the striatum in the pathogenesis of the cognitive symptoms of schizophrenia. Neuron 2010, 65, 585–596. [Google Scholar] [CrossRef] [Green Version]
- Dahoun, T.; Trossbach, S.V.; Brandon, N.J.; Korth, C.; Howes, O.D. The impact of Disrupted-in-Schizophrenia 1 (DISC1) on the dopaminergic system: A systematic review. Transl. Psychiatry 2017, 7, e1015. [Google Scholar] [CrossRef]
- Abi-Dargham, A.; Rodenhiser, J.; Printz, D.; Zea-Ponce, Y.; Gil, R.; Kegeles, L.S.; Weiss, R.; Cooper, T.B.; Mann, J.J.; Van Heertum, R.L.; et al. Increased baseline occupancy of D2 receptors by dopamine in schizophrenia. Proc. Natl. Acad. Sci. USA 2000, 97, 8104–8109. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.L.; Urban, N.; Slifstein, M.; Xu, X.; Kegeles, L.S.; Girgis, R.R.; Beckerman, Y.; Harkavy-Friedman, J.M.; Gil, R.; Abi-Dargham, A. Striatal dopamine release in schizophrenia comorbid with substance dependence. Mol. Psychiatry 2013, 18, 909–915. [Google Scholar] [CrossRef] [Green Version]
- Urs, N.M.; Peterson, S.M.; Caron, M.G. New concepts in dopamine D2 receptor biased signaling and implications for schizophrenia therapy. Biol. Psychiatry 2017, 81, 78–85. [Google Scholar] [CrossRef] [Green Version]
- Abi-Dargham, A. From “bedside” to “bench” and back: A translational approach to studying dopamine dysfunction in schizophrenia. Neurosci. Biobehav. Rev. 2020, 110, 174–179. [Google Scholar] [CrossRef]
- Su, P.; Li, S.; Chen, S.; Lipina, T.V.; Wang, M.; Lai, T.K.; Lee, F.H.; Zhang, H.; Zhai, D.; Ferguson, S.S.; et al. A dopamine D2 receptor-DISC1 protein complex may contribute to antipsychotic-like effects. Neuron 2014, 84, 1302–1316. [Google Scholar] [CrossRef] [Green Version]
- Millar, J.K.; Christie, S.; Anderson, S.; Lawson, D.; Hsiao-Wei Loh, D.; Devon, R.S.; Arveiler, B.; Muir, W.J.; Blackwood, D.H.; Porteous, D.J. Genomic structure and localisation within a linkage hotspot of Disrupted In Schizophrenia 1, a gene disrupted by a translocation segregating with schizophrenia. Mol. Psychiatry 2001, 6, 173–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duff, B.J.; Macritchie, K.A.N.; Moorhead, T.W.J.; Lawrie, S.M.; Blackwood, D.H.R. Human brain imaging studies of DISC1 in schizophrenia, bipolar disorder and depression: A systematic review. Schizophr. Res. 2013, 147, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Y.; Liu, Y.; Yan, J.W.; Hu, X.L.; Zhu, D.M.; Xu, X.T.; Li, X.S. Gene polymorphisms of DISC1 is associated with schizophrenia: Evidence from a meta-analysis. Prog. Neuropsychopharmacol. Biol. Psychiatry 2018, 81, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.M.; Liu, Y.L.; Hwu, H.G.; Fann, C.S.; Yang, U.C.; Hsu, P.C.; Chang, C.C.; Chen, W.J.; Hwang, T.J.; Hsieh, M.H.; et al. Genetic associations and expression of extra-short isoforms of disrupted-in-schizophrenia 1 in a neurocognitive subgroup of schizophrenia. J. Hum. Genet. 2019, 64, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.M.; Lin, C.H.; Lane, H.Y. Distinctively lower DISC1 mRNA levels in patients with schizophrenia, especially in those with higher positive, negative, and depressive symptoms. Pharmacol. Biochem. Behav. 2022, 213, 173335. [Google Scholar] [CrossRef]
- Brandon, N.J.; Sawa, A. Linking neurodevelopmental and synaptic theories of mental illness through DISC1. Nat. Rev. Neurosci. 2011, 12, 707–722. [Google Scholar] [CrossRef]
- Bradshaw, N.J.; Porteous, D.J. DISC1-binding proteins in neural development, signalling and schizophrenia. Neuropharmacology 2012, 62, 1230–1241. [Google Scholar] [CrossRef] [Green Version]
- Tropea, D.; Hardingham, N.; Millar, K.; Fox, K. Mechanisms underlying the role of DISC1 in synaptic plasticity. J. Physiol. 2018, 596, 2747–2771. [Google Scholar] [CrossRef] [Green Version]
- Juan, L.W.; Liao, C.C.; Lai, W.S.; Chang, C.Y.; Pei, J.C.; Wong, W.R.; Liu, C.M.; Hwu, H.G.; Lee, L.J. Phenotypic characterization of C57BL/6J mice carrying the Disc1 gene from the 129S6/SvEv strain. Brain Struct. Funct. 2014, 219, 1417–1431. [Google Scholar] [CrossRef]
- Baskaran, R.; Lai, C.C.; Li, W.Y.; Tuan, L.H.; Wang, C.C.; Lee, L.J.; Liu, C.M.; Hwu, H.G.; Lee, L.J. Characterization of striatal phenotypes in heterozygous Disc1 mutant mice, a model of haploinsufficiency. J. Comp. Neurol. 2020, 528, 1157–1172. [Google Scholar] [CrossRef]
- Ralph, R.J.; Paulus, M.P.; Geyer, M.A. Strain-specific effects of amphetamine on prepulse inhibition and patterns of locomotor behavior in mice. J. Pharmacol. Exp. Ther. 2001, 298, 148–155. [Google Scholar] [PubMed]
- Dean, O.; Giorlando, F.; Berk, M. N-acetylcysteine in psychiatry: Current therapeutic evidence and potential mechanisms of action. J. Psychiatry Neurosci. 2011, 36, 78–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berk, M.; Malhi, G.S.; Gray, L.J.; Dean, O.M. The promise of N-acetylcysteine in neuropsychiatry. Trends Pharmacol. Sci. 2013, 34, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, A.P.; Benvenutti, R.; Pilz, L.K.; Elisabetsky, E. N-acetylcysteine prevents increased amphetamine sensitivity in social isolation-reared mice. Schizophr. Res. 2014, 155, 109–111. [Google Scholar] [CrossRef]
- Deepmala Slattery, J.; Kumar, N.; Delhey, L.; Berk, M.; Dean, O.; Spielholz, C.; Frye, R. Clinical trials of N-acetylcysteine in psychiatry and neurology: A systematic review. Neurosci. Biobehav. Rev. 2015, 55, 294–321. [Google Scholar] [CrossRef] [Green Version]
- Yolland, C.O.; Hanratty, D.; Neill, E.; Rossell, S.L.; Berk, M.; Dean, O.M.; Castle, D.J.; Tan, E.J.; Phillipou, A.; Harris, A.W.; et al. Meta-analysis of randomised controlled trials with N-acetylcysteine in the treatment of schizophrenia. Aust. N. Z. J. Psychiatry 2020, 54, 453–466. [Google Scholar] [CrossRef]
- Raghu, G.; Berk, M.; Campochiaro, P.A.; Jaeschke, H.; Marenzi, G.; Richeldi, L.; Wen, F.Q.; Nicoletti, F.; Calverley, P.M.A. The multifaceted therapeutic role of N-acetylcysteine (NAC) in disorders characterized by oxidative stress. Curr. Neuropharmacol. 2021, 19, 1202–1224. [Google Scholar] [CrossRef]
- Smaga, I.; Frankowska, M.; Filip, M. N-acetylcysteine as a new prominent approach for treating psychiatric disorders. Br. J. Pharmacol. 2021, 178, 2569–2594. [Google Scholar] [CrossRef]
- Bradlow, R.C.J.; Berk, M.; Kalivas, P.W.; Back, S.E.; Kanaan, R.A. The potential of N-Acetyl-L-Cysteine (NAC) in the treatment of psychiatric disorders. CNS Drugs 2022, 36, 451–482. [Google Scholar] [CrossRef]
- Asevedo, E.; Cunha, G.R.; Zugman, A.; Mansur, R.B.; Brietzke, E. N-acetylcysteine as a potentially useful medication to prevent conversion to schizophrenia in at-risk individuals. Rev. Neurosci. 2012, 23, 353–362. [Google Scholar] [CrossRef]
- Schmidt, S.J.; Hurlemann, R.; Schultz, J.; Wasserthal, S.; Kloss, C.; Maier, W.; Meyer-Lindenberg, A.; Hellmich, M.; Muthesius-Digón, A.; ESPRIT-B1 Group; et al. Multimodal prevention of first psychotic episode through N-acetyl-l-cysteine and integrated preventive psychological intervention in individuals clinically at high risk for psychosis: Protocol of a randomized, placebo-controlled, parallel-group trial. Early Interv. Psychiatry 2019, 13, 1404–1415. [Google Scholar] [CrossRef] [PubMed]
- Lovestone, S.; Killick, R.; Di Forti, M.; Murray, R. Schizophrenia as a GSK-3 dysregulation disorder. Trends Neurosci. 2007, 30, 142–149. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, O.; Nolan, Y. Glycogen synthase kinase-3 as a therapeutic target for cognitive dysfunction in neuropsychiatric disorders. CNS Drugs 2015, 29, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Cymerman, I.A.; Gozdz, A.; Urbanska, M.; Milek, J.; Dziembowska, M.; Jaworski, J. Structural plasticity of dendritic spines requires GSK3α and GSK3β. PLoS ONE 2015, 10, e0134018. [Google Scholar]
- Cingolani, L.A.; Goda, Y. Actin in action: The interplay between the actin cytoskeleton and synaptic efficacy. Nat. Rev. Neurosci. 2008, 9, 344–356. [Google Scholar] [CrossRef]
- Tepper, J.M.; Koós, T.; Ibanez-Sandoval, O.; Tecuapetla, F.; Faust, T.W.; Assous, M. Heterogeneity and Diversity of Striatal GABAergic Interneurons: Update 2018. Front. Neuroanat. 2018, 12, 91. [Google Scholar] [CrossRef] [Green Version]
- Stansfield, K.H.; Ruby, K.N.; Soares, B.D.; McGlothan, J.L.; Liu, X.; Guilarte, T.R. Early-life lead exposure recapitulates the selective loss of parvalbumin-positive GABAergic interneurons and subcortical dopamine system hyperactivity present in schizophrenia. Transl. Psychiatry 2015, 5, e522. [Google Scholar] [CrossRef] [Green Version]
- Hamburg, H.; Trossbach, S.V.; Bader, V.; Chwiesko, C.; Kipar, A.; Sauvage, M.; Crum, W.R.; Vernon, A.C.; Bidmon, H.J.; Korth, C. Simultaneous effects on parvalbumin-positive interneuron and dopaminergic system development in a transgenic rat model for sporadic schizophrenia. Sci. Rep. 2016, 6, 34946. [Google Scholar] [CrossRef] [Green Version]
- Wiltschko, A.B.; Pettibone, J.R.; Berke, J.D. Opposite effects of stimulant and antipsychotic drugs on striatal fast-spiking interneurons. Neuropsychopharmacology 2010, 35, 1261–1270. [Google Scholar] [CrossRef]
- Woods, S.W.; Miller, T.J.; McGlashan, T.H. The “prodromal” patient: Both symptomatic and at-risk. CNS Spectr. 2001, 6, 223–232. [Google Scholar] [CrossRef]
- Miller, T.J.; McGlashan, T.H.; Rosen, J.L.; Somjee, L.; Markovich, P.J.; Stein, K.; Woods, S.W. Prospective diagnosis of the initial prodrome for schizophrenia based on the Structured Interview for Prodromal Syndromes: Preliminary evidence of interrater reliability and predictive validity. Am. J. Psychiatry 2002, 159, 863–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yung, A.R.; Phillips, L.J.; Yuen, H.P.; Francey, S.M.; McFarlane, C.A.; Hallgren, M.; McGorry, P.D. Psychosis prediction: 12-month follow up of a high-risk (“prodromal”) group. Schizophr. Res. 2003, 60, 21–32. [Google Scholar] [CrossRef]
- Howes, O.D.; McCutcheon, R.; Owen, M.J.; Murray, R.M. The role of genes, stress, and dopamine in the development of schizophrenia. Biol. Psychiatry 2017, 81, 9–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trossbach, S.V.; Bader, V.; Hecher, L.; Pum, M.E.; Masoud, S.T.; Prikulis, I.; Schäble, S.; de Souza Silva, M.A.; Su, P.; Boulat, B.; et al. Misassembly of full-length Disrupted-in-Schizophrenia 1 protein is linked to altered dopamine homeostasis and behavioral deficits. Mol. Psychiatry 2016, 21, 1561–1572. [Google Scholar] [CrossRef] [Green Version]
- Kellendonk, C.; Simpson, E.H.; Polan, H.J.; Malleret, G.; Vronskaya, S.; Winiger, V.; Moore, H.; Kandel, E.R. Transient and selective overexpression of dopamine D2 receptors in the striatum causes persistent abnormalities in prefrontal cortex functioning. Neuron 2006, 49, 603–615. [Google Scholar] [CrossRef] [Green Version]
- Cazorla, M.; Shegda, M.; Ramesh, B.; Harrison, N.L.; Kellendonk, C. Striatal D2 receptors regulate dendritic morphology of medium spiny neurons via Kir2 channels. J. Neurosci. 2012, 32, 2398–2409. [Google Scholar] [CrossRef]
- O’Hare, J.K.; Li, H.; Kim, N.; Gaidis, E.; Ade, K.; Beck, J.; Yin, H.; Calakos, N. Striatal fast-spiking interneurons selectively modulate circuit output and are required for habitual behavior. eLife 2017, 6, e26231. [Google Scholar] [CrossRef]
- Lee, F.H.; Zai, C.C.; Cordes, S.P.; Roder, J.C.; Wong, A.H. Abnormal interneuron development in disrupted-in-schizophrenia-1 L100P mutant mice. Mol. Brain 2013, 6, 20. [Google Scholar] [CrossRef] [Green Version]
- Nakai, T.; Nagai, T.; Wang, R.; Yamada, S.; Kuroda, K.; Kaibuchi, K.; Yamada, K. Alterations of GABAergic and dopaminergic systems in mutant mice with disruption of exons 2 and 3 of the Disc1 gene. Neurochem. Int. 2014, 74, 74–83. [Google Scholar] [CrossRef] [Green Version]
- Cabungcal, J.H.; Steullet, P.; Kraftsik, R.; Cuenod, M.; Do, K.Q. Early-life insults impair parvalbumin interneurons via oxidative stress: Reversal by N-acetylcysteine. Biol. Psychiatry 2013, 73, 574–582. [Google Scholar] [CrossRef]
- Cardis, R.; Cabungcal, J.H.; Dwir, D.; Do, K.Q.; Steullet, P. A lack of GluN2A-containing NMDA receptors confers a vulnerability to redox dysregulation: Consequences on parvalbumin interneurons, and their perineuronal nets. Neurobiol. Dis. 2018, 109, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Dwir, D.; Cabungcal, J.H.; Xin, L.; Giangreco, B.; Parietti, E.; Cleusix, M.; Jenni, R.; Klauser, P.; Conus, P.; Cuénod, M.; et al. Timely N-Acetyl-Cysteine and environmental enrichment rescue oxidative stress-induced parvalbumin interneuron impairments via MMP9/RAGE pathway: A translational approach for early intervention in psychosis. Schizophr. Bull. 2021, 47, 1782–1794. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Cabungcal, J.H.; Cuenod, M.; Uliana, D.L.; Do, K.Q.; Grace, A.A. Thalamic reticular nucleus impairments and abnormal prefrontal control of dopamine system in a developmental model of schizophrenia: Prevention by N-acetylcysteine. Mol. Psychiatry 2021, 26, 7679–7689. [Google Scholar] [CrossRef] [PubMed]
- Kulak, A.; Steullet, P.; Cabungcal, J.H.; Werge, T.; Ingason, A.; Cuenod, M.; Do, K.Q. Redox dysregulation in the pathophysiology of schizophrenia and bipolar disorder: Insights from animal models. Antioxid. Redox Signal. 2013, 18, 1428–1443. [Google Scholar] [CrossRef]
- Cabungcal, J.H.; Counotte, D.S.; Lewis, E.; Tejeda, H.A.; Piantadosi, P.; Pollock, C.; Calhoon, G.G.; Sullivan, E.; Presgraves, E.; Kil, J.; et al. Juvenile antioxidant treatment prevents adult deficits in a developmental model of schizophrenia. Neuron 2014, 83, 1073–1084. [Google Scholar] [CrossRef] [Green Version]
- Phensy, A.; Driskill, C.; Lindquist, K.; Guo, L.; Jeevakumar, V.; Fowler, B.; Du, H.; Kroener, S. Antioxidant treatment in male mice prevents mitochondrial and synaptic changes in an NMDA receptor dysfunction model of schizophrenia. eNeuro 2017, 4, ENEURO.0081-17.2017. [Google Scholar] [CrossRef] [Green Version]
- Salim, S. Oxidative stress and psychological disorders. Curr. Neuropharmacol. 2014, 12, 140–147. [Google Scholar] [CrossRef] [Green Version]
- Steullet, P.; Cabungcal, J.H.; Coyle, J.; Didriksen, M.; Gill, K.; Grace, A.A.; Hensch, T.K.; LaMantia, A.S.; Lindemann, L.; Maynard, T.M.; et al. Oxidative stress-driven parvalbumin interneuron impairment as a common mechanism in models of schizophrenia. Mol. Psychiatry 2017, 22, 936–943. [Google Scholar] [CrossRef]
- Perkins, D.O.; Jeffries, C.D.; Do, K.Q. Potential roles of redox dysregulation in the development of schizophrenia. Biol. Psychiatry 2020, 88, 326–336. [Google Scholar] [CrossRef]
- Curpan, A.S.; Luca, A.C.; Ciobica, A. Potential novel therapies for neurodevelopmental diseases targeting oxidative stress. Oxid. Med. Cell. Longev. 2021, 2021, 6640206. [Google Scholar] [CrossRef]
- Kim, Y.; Vadodaria, K.C.; Lenkei, Z.; Kato, T.; Gage, F.H.; Marchetto, M.C.; Santos, R. Mitochondria, metabolism, and redox mechanisms in psychiatric disorders. Antioxid. Redox Signal. 2019, 31, 275–317. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.W.; Jaaro-Peled, H.; Shahani, N.; Sedlak, T.W.; Zoubovsky, S.; Burruss, D.; Emiliani, F.; Sawa, A.; Gallagher, M. Cognitive and motivational deficits together with prefrontal oxidative stress in a mouse model for neuropsychiatric illness. Proc. Natl. Acad. Sci. USA 2013, 110, 12462–12467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bridges, R.J.; Natale, N.R.; Patel, S.A. System xc⁻ cystine/glutamate antiporter: An update on molecular pharmacology and roles within the CNS. Br. J. Pharmacol. 2012, 165, 20–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moran, M.M.; McFarland, K.; Melendez, R.I.; Kalivas, P.W.; Seamans, J.K. Cystine/glutamate exchange regulates metabotropic glutamate receptor presynaptic inhibition of excitatory transmission and vulnerability to cocaine seeking. J. Neurosci. 2005, 25, 6389–6393. [Google Scholar] [CrossRef] [Green Version]
- Yi, S.Y.; Barnett, B.R.; Poetzel, M.J.; Stowe, N.A.; Yu, J.J. Clinical translational neuroimaging of the antioxidant effect of N-acetylcysteine on neural microstructure. Magn. Reson. Med. 2022, 87, 820–836. [Google Scholar] [CrossRef]
- Markoutsa, E.; Xu, P. Redox potential-sensitive N-Acetyl Cysteine-prodrug nanoparticles inhibit the activation of microglia and improve neuronal survival. Mol. Pharm. 2017, 14, 1591–1600. [Google Scholar] [CrossRef] [Green Version]
- Beasley, C.; Cotter, D.; Khan, N.; Pollard, C.; Sheppard, P.; Varndell, I.; Lovestone, S.; Anderton, B.; Everall, I. Glycogen synthase kinase-3beta immunoreactivity is reduced in the prefrontal cortex in schizophrenia. Neurosci. Lett. 2001, 302, 117–120. [Google Scholar] [CrossRef]
- Kozlovsky, N.; Shanon-Weickert, C.; Tomaskovic-Crook, E.; Kleinman, J.E.; Belmaker, R.H.; Agam, G. Reduced GSK-3beta mRNA levels in postmortem dorsolateral prefrontal cortex of schizophrenic patients. J. Neural Transm. 2004, 111, 1583–1592. [Google Scholar] [CrossRef] [Green Version]
- Hur, E.M.; Zhou, F.Q. GSK3 signalling in neural development. Nat. Rev. Neurosci. 2010, 11, 539–551. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.Y.; Snider, W.D. Functions of GSK-3 signaling in development of the nervous system. Front. Mol. Neurosci. 2011, 4, 44. [Google Scholar] [CrossRef] [Green Version]
- Hajka, D.; Budziak, B.; Pietras, Ł.; Duda, P.; McCubrey, J.A.; Gizak, A. GSK3 as a regulator of cytoskeleton architecture: Consequences for health and disease. Cells 2021, 10, 2092. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Zhao, Z.; Wang, D.; Xiao, J. Glycogen synthase kinase-3 as a key regulator of cognitive function. Acta Biochim. Biophys. Sin. 2020, 52, 219–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vita, A.; Barlati, S.; Ceraso, A.; Nibbio, G.; Ariu, C.; Deste, G.; Wykes, T. Effectiveness, core elements, and moderators of response of cognitive remediation for schizophrenia: A systematic review and meta-analysis of randomized clinical trials. JAMA Psychiatry 2021, 78, 848–858. [Google Scholar] [CrossRef] [PubMed]
- Veselinović, T.; Neuner, I. Progress and pitfalls in developing agents to treat neurocognitive deficits associated with schizophrenia. CNS Drugs 2022, 36, 819–858. [Google Scholar] [CrossRef]
- Ito, H.; Ichiyanagi, O.; Naito, S.; Bilim, V.N.; Tomita, Y.; Kato, T.; Nagaoka, A.; Tsuchiya, N. GSK-3 directly regulates phospho-4EBP1 in renal cell carcinoma cell-line: An intrinsic subcellular mechanism for resistance to mTORC1 inhibition. BMC Cancer 2016, 16, 393. [Google Scholar] [CrossRef] [Green Version]
- Moy, J.K.; Khoutorsky, A.; Asiedu, M.N.; Dussor, G.; Price, T.J. eIF4E phosphorylation influences Bdnf mRNA translation in mouse dorsal root ganglion neurons. Front. Cell. Neurosci. 2018, 12, 29. [Google Scholar] [CrossRef] [Green Version]
- Scharfman, H.; Goodman, J.; Macleod, A.; Phani, S.; Antonelli, C.; Croll, S. Increased neurogenesis and the ectopic granule cells after intrahippocampal BDNF infusion in adult rats. Exp. Neurol. 2005, 192, 348–356. [Google Scholar] [CrossRef]
- Bath, K.G.; Schilit, A.; Lee, F.S. Stress effects on BDNF expression: Effects of age, sex, and form of stress. Neuroscience 2013, 239, 149–156. [Google Scholar] [CrossRef]
- Park, H.; Poo, M.M. Neurotrophin regulation of neural circuit development and function. Nat. Rev. Neurosci. 2013, 14, 7–23. [Google Scholar] [CrossRef]
- Calabrese, F.; Rossetti, A.C.; Racagni, G.; Gass, P.; Riva, M.A.; Molteni, R. Brain-derived neurotrophic factor: A bridge between inflammation and neuroplasticity. Front. Cell. Neurosci. 2014, 8, 430. [Google Scholar] [CrossRef]
- Tsao, C.Y.; Tuan, L.H.; Lee, L.J.; Liu, C.M.; Hwu, H.G.; Lee, L.J. Impaired response to sleep deprivation in heterozygous Disc1 mutant mice. World J. Biol. Psychiatry 2022, 23, 55–66. [Google Scholar] [CrossRef] [PubMed]
- das Neves Duarte, J.M.; Kulak, A.; Gholam-Razaee, M.M.; Cuenod, M.; Gruetter, R.; Do, K.Q. N-acetylcysteine normalizes neurochemical changes in the glutathione-deficient schizophrenia mouse model during development. Biol. Psychiatry 2012, 71, 1006–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.C.; Ho, U.C.; Ko, M.C.; Liao, C.C.; Lee, L.J. Differential neuronal changes in medial prefrontal cortex, basolateral amygdala and nucleus accumbens after postweaning social isolation. Brain Struct. Funct. 2012, 217, 337–351. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, C.-C.; Baskaran, R.; Tsao, C.-Y.; Tuan, L.-H.; Siow, P.-F.; Palani, M.; Lee, L.J.-H.; Liu, C.-M.; Hwu, H.-G.; Lee, L.-J. Chronic N-Acetylcysteine Treatment Prevents Amphetamine-Induced Hyperactivity in Heterozygous Disc1 Mutant Mice, a Putative Prodromal Schizophrenia Animal Model. Int. J. Mol. Sci. 2022, 23, 9419. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23169419
Lai C-C, Baskaran R, Tsao C-Y, Tuan L-H, Siow P-F, Palani M, Lee LJ-H, Liu C-M, Hwu H-G, Lee L-J. Chronic N-Acetylcysteine Treatment Prevents Amphetamine-Induced Hyperactivity in Heterozygous Disc1 Mutant Mice, a Putative Prodromal Schizophrenia Animal Model. International Journal of Molecular Sciences. 2022; 23(16):9419. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23169419
Chicago/Turabian StyleLai, Chuan-Ching, Rathinasamy Baskaran, Chih-Yu Tsao, Li-Heng Tuan, Pei-Fen Siow, Mahalakshmi Palani, Lukas Jyuhn-Hsiarn Lee, Chih-Min Liu, Hai-Gwo Hwu, and Li-Jen Lee. 2022. "Chronic N-Acetylcysteine Treatment Prevents Amphetamine-Induced Hyperactivity in Heterozygous Disc1 Mutant Mice, a Putative Prodromal Schizophrenia Animal Model" International Journal of Molecular Sciences 23, no. 16: 9419. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23169419