
Differential Ganglioside and Cholesterol Depletion by Various Cyclodextrin Derivatives and Their Effect on Synaptosomal Glutamate Release


Abstract
:1. Introduction
2. Results and Discussion
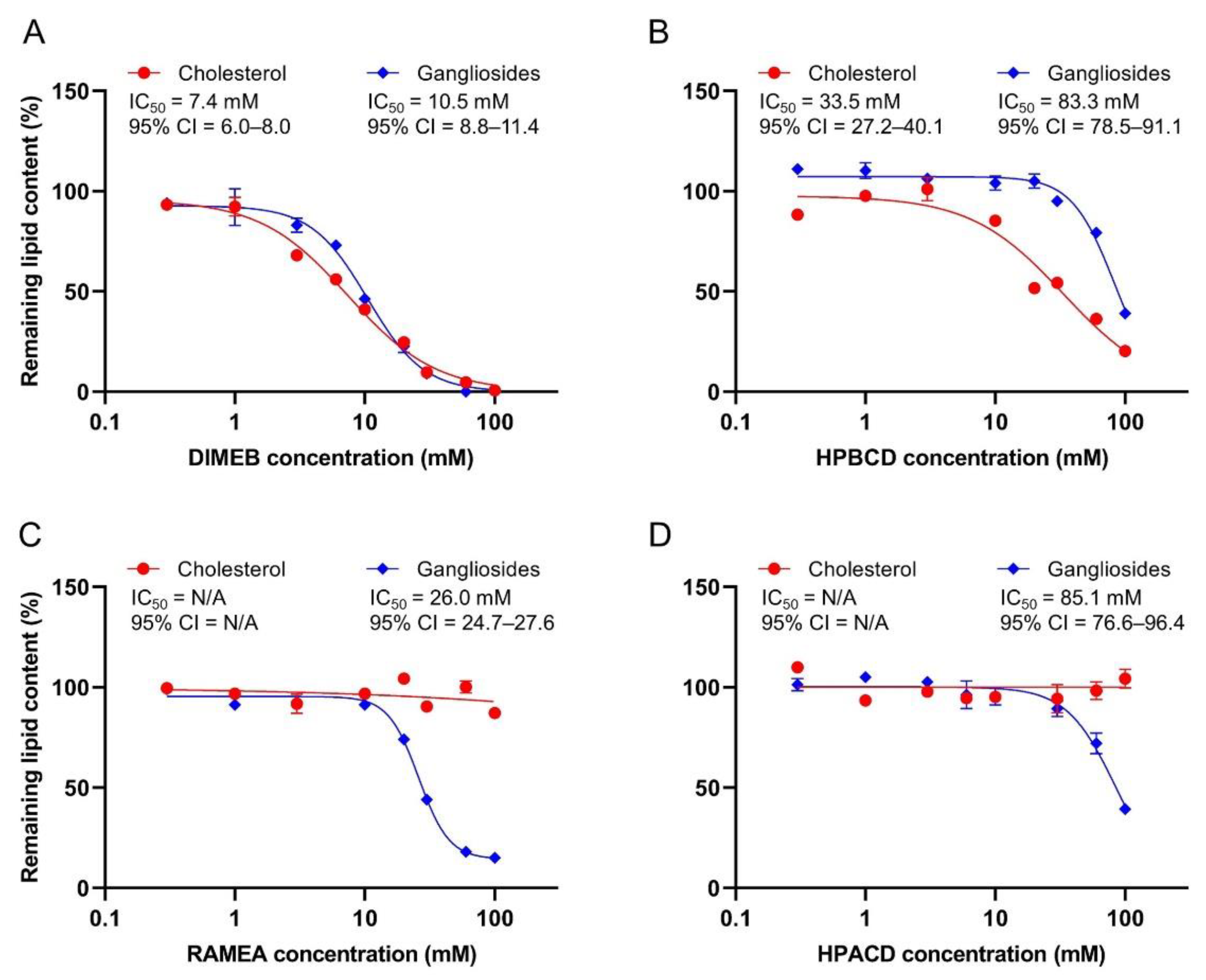
2.1. Effect of CDs on Ganglioside and Cholesterol Content of Synaptosomes
2.2. Effect of CDs on Membrane Integrity and Viability of Synaptosomes
2.3. Effect of CDs on Basal and 4-Aminopyridine (4-AP)-Evoked Glutamate Release from Synaptosomes
3. Materials and Methods
3.1. Chemicals
3.2. Preparation and Treatment of Synaptosomes with Cyclodextrins
3.3. Ganglioside Analysis
3.4. Cholesterol Assay
3.5. Resazurin Reduction Viability Assay
3.6. Membrane Integrity Measurement by LDH Release Assay
3.7. Glutamate Release Measurement
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Kwak, D.H.; Seo, B.B.; Chang, K.T.; Choo, Y.K. Roles of gangliosides in mouse embryogenesis and embryonic stem cell differentiation. Exp. Mol. Med. 2011, 43, 379–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vajn, K.; Viljetic, B.; Degmecic, I.V.; Schnaar, R.L.; Heffer, M. Differential Distribution of Major Brain Gangliosides in the Adult Mouse Central Nervous System. PLoS ONE 2013, 8, e75720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.C.; Lee, W.S.; Park, C.S.; Juhng, S.W. The biologic role of ganglioside in neuronal differentiation—Effects of GM1 ganglioside on human neuroblastoma SH-SY5Y cells. J. Korean Med. Sci. 1994, 9, 179–187. [Google Scholar] [CrossRef] [Green Version]
- Roisen, F.J.; Bartfeld, H.; Nagele, R.; Yorke, G. Ganglioside stimulation of axonal sprouting in vitro. Science 1981, 214, 577–578. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Kobayashi, M.; Sugiura, M.; Ozaki, N.; Nishio, K.; Shiraishi, Y.; Furukawa, K.; Furukawa, K.; Sugiura, Y. Morphological study of disordered myelination and the degeneration of nerve fibers in the spinal cord of mice lacking complex gangliosides. Arch. Histol. Cytol. 2003, 66, 37–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jennemann, R.; Sandhoff, R.; Wang, S.; Kiss, E.; Gretz, N.; Zuliani, C.; Martin-Villalba, A.; Jager, R.; Schorle, H.; Kenzelmann, M.; et al. Cell-specific deletion of glucosylceramide synthase in brain leads to severe neural defects after birth. Proc. Natl. Acad. Sci. USA 2005, 102, 12459–12464. [Google Scholar] [CrossRef] [Green Version]
- Schengrund, C.L. Lipid rafts: Keys to neurodegeneration. Brain Res. Bull. 2010, 82, 7–17. [Google Scholar] [CrossRef]
- Sonnino, S.; Mauri, L.; Chigorno, V.; Prinetti, A. Gangliosides as components of lipid membrane domains. Glycobiology 2007, 17, 1R–13R. [Google Scholar] [CrossRef] [Green Version]
- Quinn, P.J.; Wolf, C. The liquid-ordered phase in membranes. Biochim. Biophys. Acta 2009, 1788, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Munro, S. Lipid rafts: Elusive or illusive? Cell 2003, 115, 377–388. [Google Scholar] [CrossRef] [Green Version]
- Sevcsik, E.; Schutz, G.J. With or without rafts? Alternative views on cell membranes. BioEssays News Rev. Mol. Cell. Dev. Biol. 2016, 38, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Horvath, A.; Biro-Suto, T.; Kantas, B.; Payrits, M.; Skoda-Foldes, R.; Szanti-Pinter, E.; Helyes, Z.; Szoke, E. Antinociceptive Effects of Lipid Raft Disruptors, a Novel Carboxamido-Steroid and Methyl beta-Cyclodextrin, in Mice by Inhibiting Transient Receptor Potential Vanilloid 1 and Ankyrin 1 Channel Activation. Front. Physiol. 2020, 11, 559109. [Google Scholar] [CrossRef]
- Santha, P.; Oszlacs, O.; Dux, M.; Dobos, I.; Jancso, G. Inhibition of glucosylceramide synthase reversibly decreases the capsaicin-induced activation and TRPV1 expression of cultured dorsal root ganglion neurons. Pain 2010, 150, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.A.; Halverson-Tamboli, R.A.; Rasenick, M.M. Lipid raft microdomains and neurotransmitter signalling. Nat. Rev. Neurosci. 2007, 8, 128–140. [Google Scholar] [CrossRef]
- Xu, Y.; Sun, J.; Yang, L.; Zhao, S.; Liu, X.; Su, Y.; Zhang, J.; Zhao, M. Gangliosides play important roles in the nervous system by regulating ion concentrations. Neurochem. Res. 2022, 47, 1791–1798. [Google Scholar] [CrossRef] [PubMed]
- European Medicines Agency (EMA). Cyclodextrins Used as Excipients Report Published in Support of the ‘Questions and Answers on Cyclodextrins Used as Excipients in Medicinal Products for Human Use’ (EMA/CHMP/495747/2013). Available online: https://www.ema.europa.eu/en/documents/scientific-guideline/questions-answers-cyclodextrins-used-excipients-medicinal-products-human-use_en.pdf (accessed on 18 July 2022).
- Saokham, P.; Muankaew, C.; Jansook, P.; Loftsson, T. Solubility of Cyclodextrins and Drug/Cyclodextrin Complexes. Molecules 2018, 23, 1161. [Google Scholar] [CrossRef] [Green Version]
- Singhal, A.; Szente, L.; Hildreth, J.E.K.; Song, B. Hydroxypropyl-beta and -gamma cyclodextrins rescue cholesterol accumulation in Niemann-Pick C1 mutant cell via lysosome-associated membrane protein 1. Cell Death Dis. 2018, 9, 1019. [Google Scholar] [CrossRef]
- Kilsdonk, E.P.; Yancey, P.G.; Stoudt, G.W.; Bangerter, F.W.; Johnson, W.J.; Phillips, M.C.; Rothblat, G.H. Cellular cholesterol efflux mediated by cyclodextrins. J. Biol. Chem. 1995, 270, 17250–17256. [Google Scholar] [CrossRef] [Green Version]
- Ohtani, Y.; Irie, T.; Uekama, K.; Fukunaga, K.; Pitha, J. Differential effects of alpha-, beta- and gamma-cyclodextrins on human erythrocytes. Eur. J. Biochem. 1989, 186, 17–22. [Google Scholar] [CrossRef]
- Zidovetzki, R.; Levitan, I. Use of cyclodextrins to manipulate plasma membrane cholesterol content: Evidence, misconceptions and control strategies. Biochim. Biophys. Acta 2007, 1768, 1311–1324. [Google Scholar] [CrossRef] [Green Version]
- Davidson, C.D.; Fishman, Y.I.; Puskas, I.; Szeman, J.; Sohajda, T.; McCauliff, L.A.; Sikora, J.; Storch, J.; Vanier, M.T.; Szente, L.; et al. Efficacy and ototoxicity of different cyclodextrins in Niemann-Pick C disease. Ann. Clin. Transl. Neurol. 2016, 3, 366–380. [Google Scholar] [CrossRef]
- Maeda, Y.; Motoyama, K.; Higashi, T.; Onodera, R.; Takeo, T.; Nakagata, N.; Kurauchi, Y.; Katsuki, H.; Ishitsuka, Y.; Kondo, Y.; et al. Lowering effect of dimethyl—Cyclodextrin on GM1-ganglioside accumulation in GM1-gangliosidosis model cells and in brain of -galactosidase-knockout mice. J. Incl. Phenom. Macrocycl. Chem. 2019, 93, 53–66. [Google Scholar] [CrossRef]
- Geda, O.; Tabi, T.; Szoko, E. Development and validation of capillary electrophoresis method for quantification of gangliosides in brain synaptosomes. J. Pharm. Biomed. Anal. 2021, 205, 114329. [Google Scholar] [CrossRef] [PubMed]
- Szente, L.; Fenyvesi, E. Cyclodextrin-Lipid Complexes: Cavity Size Matters. Struct. Chem. 2017, 28, 479–492. [Google Scholar] [CrossRef]
- Ottico, E.; Prinetti, A.; Prioni, S.; Giannotta, C.; Basso, L.; Chigorno, V.; Sonnino, S. Dynamics of membrane lipid domains in neuronal cells differentiated in culture. J. Lipid Res. 2003, 44, 2142–2151. [Google Scholar] [CrossRef] [Green Version]
- Irie, T.; Otagiri, M.; Sunada, M.; Uekama, K.; Ohtani, Y.; Yamada, Y.; Sugiyama, Y. Cyclodextrin-induced hemolysis and shape changes of human erythrocytes in vitro. J. Pharm.-Dyn. 1982, 5, 741–744. [Google Scholar] [CrossRef] [Green Version]
- Nakanishi, K.; Nadai, T.; Masada, M.; Miyajima, K. Effect of cyclodextrins on biological membrane. II. Mechanism of enhancement on the intestinal absorption of non-absorbable drug by cyclodextrins. Chem. Pharm. Bull. 1992, 40, 1252–1256. [Google Scholar] [CrossRef] [Green Version]
- Irie, T.; Uekama, K. Pharmaceutical applications of cyclodextrins. III. Toxicological issues and safety evaluation. J. Pharm. Sci. 1997, 86, 147–162. [Google Scholar] [CrossRef]
- Kiss, T.; Fenyvesi, F.; Bacskay, I.; Varadi, J.; Fenyvesi, E.; Ivanyi, R.; Szente, L.; Tosaki, A.; Vecsernyes, M. Evaluation of the cytotoxicity of beta-cyclodextrin derivatives: Evidence for the role of cholesterol extraction. Eur. J. Pharm. Sci. 2010, 40, 376–380. [Google Scholar] [CrossRef]
- Arima, H.; Motoyama, K.; Irie, T. Recent Findings on Safety Profiles of Cyclodextrins, Cyclodextrin Conjugates, and Polypseudorotaxanes. In Cyclodextrins in Pharmaceutics, Cosmetics, and Biomedicine; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2011; pp. 91–122. [Google Scholar]
- Monnaert, V.; Tilloy, S.; Bricout, H.; Fenart, L.; Cecchelli, R.; Monflier, E. Behavior of alpha-, beta-, and gamma-cyclodextrins and their derivatives on an in vitro model of blood-brain barrier. J. Pharmacol. Exp. Ther. 2004, 310, 745–751. [Google Scholar] [CrossRef] [Green Version]
- Rusznyak, A.; Malanga, M.; Fenyvesi, E.; Szente, L.; Varadi, J.; Bacskay, I.; Vecsernyes, M.; Vasvari, G.; Haimhoffer, A.; Feher, P.; et al. Investigation of the Cellular Effects of Beta- Cyclodextrin Derivatives on Caco-2 Intestinal Epithelial Cells. Pharmaceutics 2021, 13, 157. [Google Scholar] [CrossRef] [PubMed]
- Totterman, A.M.; Schipper, N.G.; Thompson, D.O.; Mannermaa, J.P. Intestinal safety of water-soluble beta-cyclodextrins in paediatric oral solutions of spironolactone: Effects on human intestinal epithelial Caco-2 cells. J. Pharm. Pharmacol. 1997, 49, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Roka, E.; Ujhelyi, Z.; Deli, M.; Bocsik, A.; Fenyvesi, E.; Szente, L.; Fenyvesi, F.; Vecsernyes, M.; Varadi, J.; Feher, P.; et al. Evaluation of the Cytotoxicity of alpha-Cyclodextrin Derivatives on the Caco-2 Cell Line and Human Erythrocytes. Molecules 2015, 20, 20269–20285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Churchward, M.A.; Coorssen, J.R. Cholesterol, regulated exocytosis and the physiological fusion machine. Biochem. J. 2009, 423, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfrieger, F.W. Role of cholesterol in synapse formation and function. Biochim. Biophys. Acta 2003, 1610, 271–280. [Google Scholar] [CrossRef] [Green Version]
- Lang, T.; Bruns, D.; Wenzel, D.; Riedel, D.; Holroyd, P.; Thiele, C.; Jahn, R. SNAREs are concentrated in cholesterol-dependent clusters that define docking and fusion sites for exocytosis. Embo J. 2001, 20, 2202–2213. [Google Scholar] [CrossRef] [Green Version]
- Salaun, C.; Gould, G.W.; Chamberlain, L.H. Lipid raft association of SNARE proteins regulates exocytosis in PC12 cells. J. Biol. Chem. 2005, 280, 19449–19453. [Google Scholar] [CrossRef] [Green Version]
- Taverna, E.; Saba, E.; Rowe, J.; Francolini, M.; Clementi, F.; Rosa, P. Role of lipid microdomains in P/Q-type calcium channel (Ca(v)2.1) clustering and function in presynaptic membranes. J. Biol. Chem. 2004, 279, 5127–5134. [Google Scholar] [CrossRef] [Green Version]
- Xia, F.Z.; Gao, X.D.; Kwan, E.; Lam, P.P.L.; Chan, L.L.; Sy, K.Y.; Sheu, L.; Wheeler, M.B.; Gaisano, H.Y.; Tsushima, R.G. Disruption of pancreatic beta-cell lipid rafts modifies K(v)2.1 channel gating and insulin exocytosis. J. Biol. Chem. 2004, 279, 24685–24691. [Google Scholar] [CrossRef] [Green Version]
- Xia, F.Z.; Leung, Y.M.; Gaisano, G.; Gao, X.D.; Chen, Y.; Fox, J.E.M.; Bhattacharjee, A.; Wheeler, M.B.; Gaisano, H.Y.; Tsushima, R.G. Targeting of voltage-gated K+ and Ca2+ channels and soluble N-ethylmaleimide-sensitive factor attachment protein receptor proteins to cholesterol-rich lipid rafts in pancreatic alpha-cells: Effects on glucagon stimulus-secretion coupling. Endocrinology 2007, 148, 2157–2167. [Google Scholar] [CrossRef] [Green Version]
- Gil, C.; Soler-Jover, A.; Blasi, J.; Aguilera, J. Synaptic proteins and SNARE complexes are localized in lipid rafts from rat brain synaptosomes. Biochem. Biophys. Res. Commun. 2005, 329, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Tarasenko, A.S.; Sivko, R.V.; Krisanova, N.V.; Himmelreich, N.H.; Borisova, T.A. Cholesterol Depletion from the Plasma Membrane Impairs Proton and Glutamate Storage in Synaptic Vesicles of Nerve Terminals. J. Mol. Neurosci. 2010, 41, 358–367. [Google Scholar] [CrossRef]
- Tanaka, Y.; Waki, H.; Kon, K.; Ando, S. Gangliosides enhance KCl-induced Ca2+ influx and acetylcholine release in brain synaptosomes. Neuroreport 1997, 8, 2203–2207. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Higashi, H.; Ogawa, H.; Takamori, K.; Iwabuchi, K. Involvement of ganglioside GT1b in glutamate release from neuroblastoma cells. Neurosci. Lett. 2012, 517, 140–143. [Google Scholar] [CrossRef] [PubMed]
- Modi, J.; Prentice, H.; Wu, J.-Y. Preparation, Stimulation and Other Uses of Adult Rat Brain Synaptosomes. Bio-Protocol 2017, 7, e2664. [Google Scholar] [CrossRef] [Green Version]
- Begley, J.G.; Butterfield, D.A.; Keller, J.N.; Koppal, T.; Drake, J.; Mattson, M.P. Cryopreservation of rat cortical synaptosomes and analysis of glucose and glutamate transporter activities, and mitochondrial function. Brain Res. Brain Res. Protoc. 1998, 3, 76–82. [Google Scholar] [CrossRef]
- Daniel, J.A.; Malladi, C.S.; Kettle, E.; McCluskey, A.; Robinson, P.J. Analysis of synaptic vesicle endocytosis in synaptosomes by high-content screening. Nat. Protoc. 2012, 7, 1439–1455. [Google Scholar] [CrossRef]
- Shimamoto, K.; LeBrun, B.; Yasuda-Kamatani, Y.; Sakaitani, M.; Shigeri, Y.; Yumoto, N.; Nakajima, T. DL-threo-beta-benzyloxyaspartate, a potent blocker of excitatory amino acid transporters. Mol. Pharmacol. 1998, 53, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Wagner, Z.; Tabi, T.; Jako, T.; Zachar, G.; Csillag, A.; Szoko, E. Chiral separation and determination of excitatory amino acids in brain samples by CE-LIF using dual cyclodextrin system. Anal. Bioanal. Chem. 2012, 404, 2363–2368. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| DIMEB | HPBCD | RAMEA | HPACD | |||||
|---|---|---|---|---|---|---|---|---|
| IC50 (mM) | 95% CI | IC50 (mM) | 95% CI | IC50 (mM) | 95% CI | IC50 (mM) | 95% CI | |
| GM1 | 15.0 | 12.7–17.6 | 80.8 | 76.0–89.5 | 29.1 | 27.9–30.5 | 86.7 | 79.0–100.7 |
| GD1a | 9.7 | 7.3–15.0 | 83.7 | 78.6–90.4 | 25.1 | 23.4–27.2 | 72.9 | 65.6–82.7 |
| GD1b | 14.5 | 10.7–23.9 | 86.6 | 80.2–98.0 | 26.4 | 24.6–28.3 | 90.6 | 68.2–139.9 |
| GT1b | 8.7 | 7.9–9.6 | 80.1 | 74.6–90.5 | 24.2 | 22.8–25.6 | 104.8 | 93.9–133.5 |
| GQ1b | 11.4 | 9.8–13.4 | 85.1 | 71.3–105.0 | 30.3 | 26.3–42.0 | 61.2 | 45.5–72.2 |
| Total | 10.5 | 8.8–11.4 | 83.3 | 78.5–91.1 | 26.0 | 24.7–27.6 | 85.1 | 76.6–96.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geda, O.; Tábi, T.; Lakatos, P.P.; Szökő, É. Differential Ganglioside and Cholesterol Depletion by Various Cyclodextrin Derivatives and Their Effect on Synaptosomal Glutamate Release. Int. J. Mol. Sci. 2022, 23, 9460. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23169460
Geda O, Tábi T, Lakatos PP, Szökő É. Differential Ganglioside and Cholesterol Depletion by Various Cyclodextrin Derivatives and Their Effect on Synaptosomal Glutamate Release. International Journal of Molecular Sciences. 2022; 23(16):9460. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23169460
Chicago/Turabian StyleGeda, Orsolya, Tamás Tábi, Péter P. Lakatos, and Éva Szökő. 2022. "Differential Ganglioside and Cholesterol Depletion by Various Cyclodextrin Derivatives and Their Effect on Synaptosomal Glutamate Release" International Journal of Molecular Sciences 23, no. 16: 9460. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23169460