
Metal Ion Periplasmic-Binding Protein YfeA of Glaesserella parasuis Induces the Secretion of Pro-Inflammatory Cytokines of Macrophages via MAPK and NF-κB Signaling through TLR2 and TLR4

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Bioinformatics Analysis of YfeA
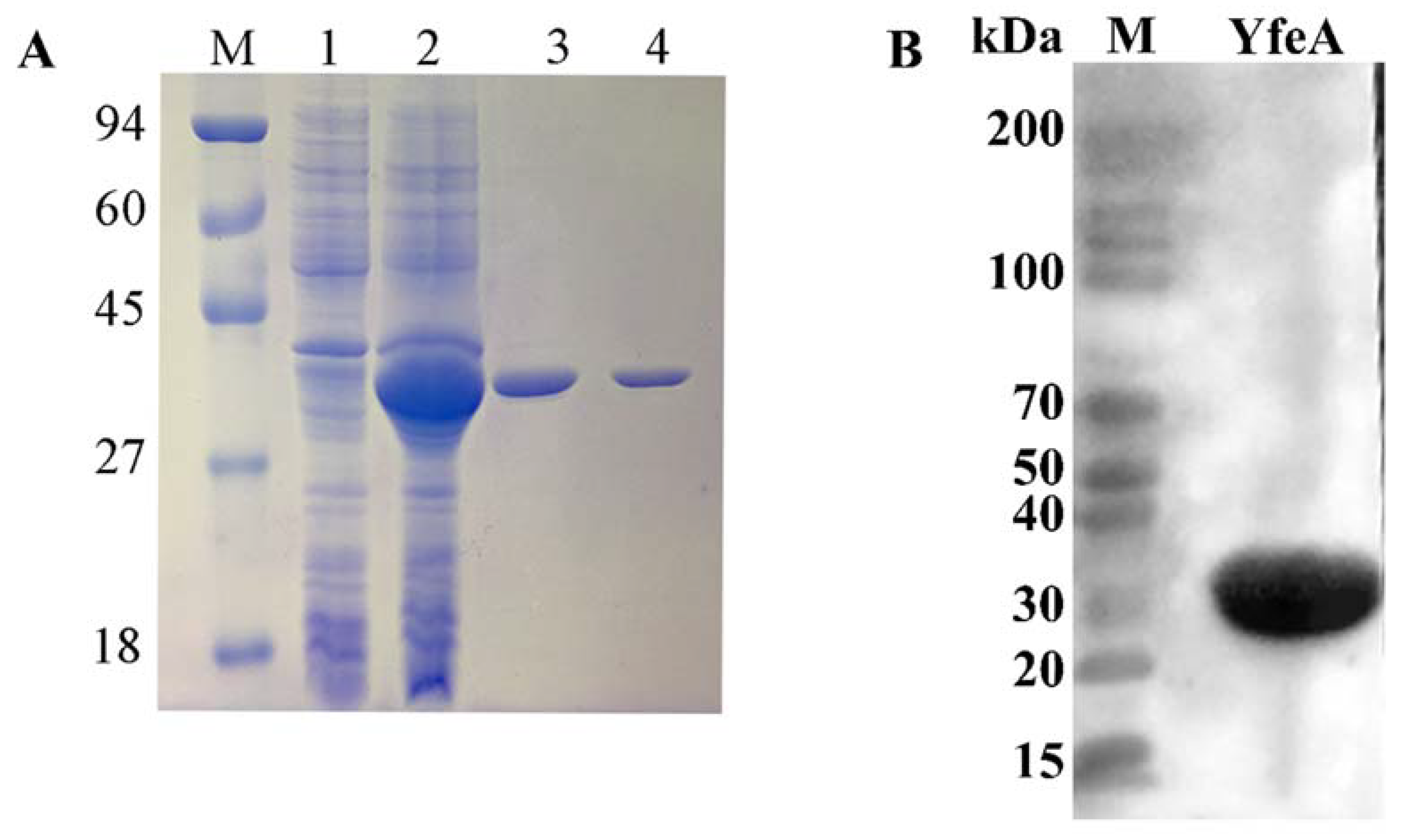
2.2. Recombinant YfeA Protein Could Be Applied to Cell Treatment
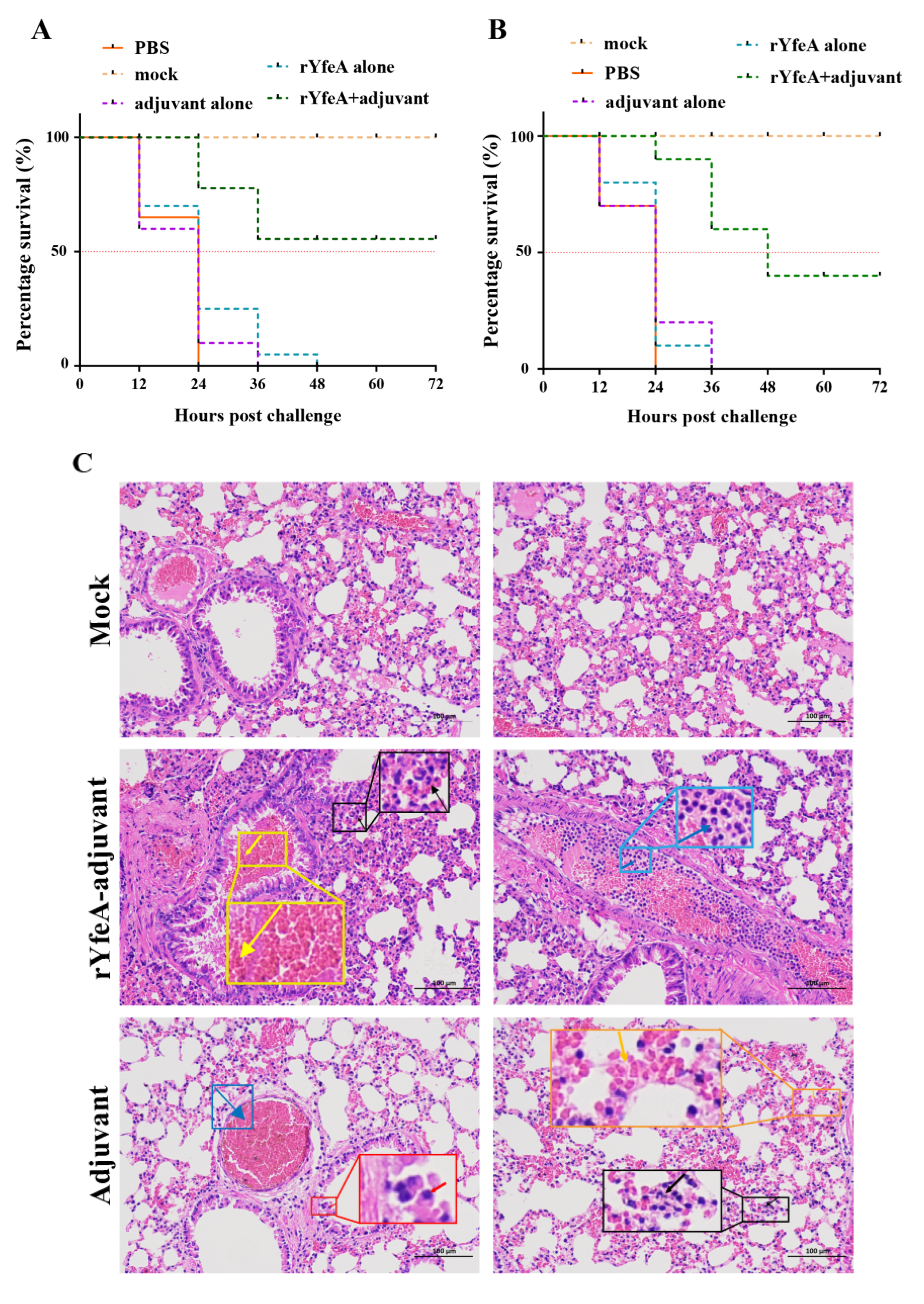
2.3. The Immunity of rYfeA-Vaccinated Mice to G. parasuis
2.4. YfeA Actively Induces the Secretion of Pro-Inflammatory Cytokines IL-1β, IL-6, and TNF-α
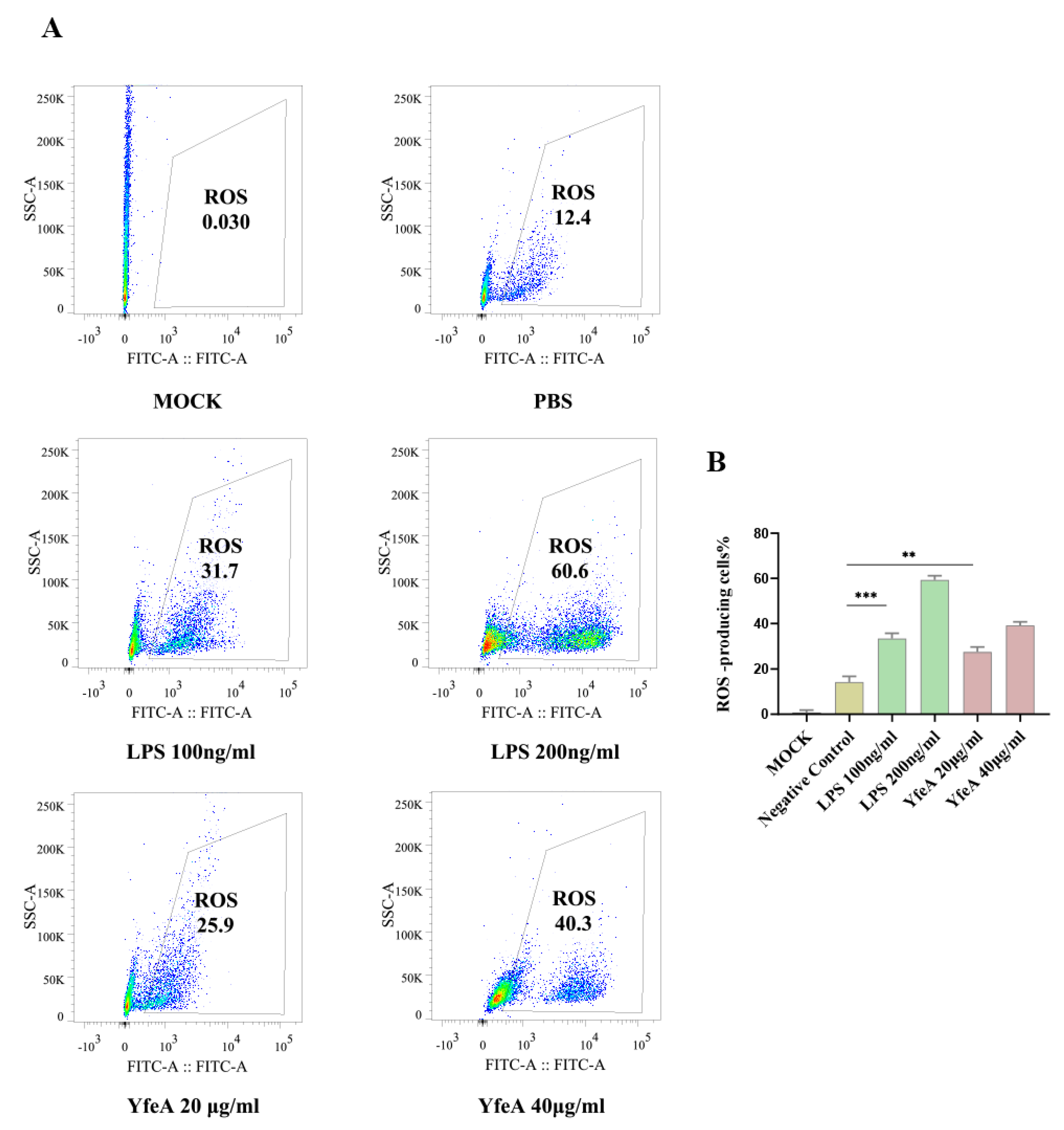
2.5. YfeA Stimulates Reactive Oxygen Species (ROS) Production in RAW 264.7 Cells
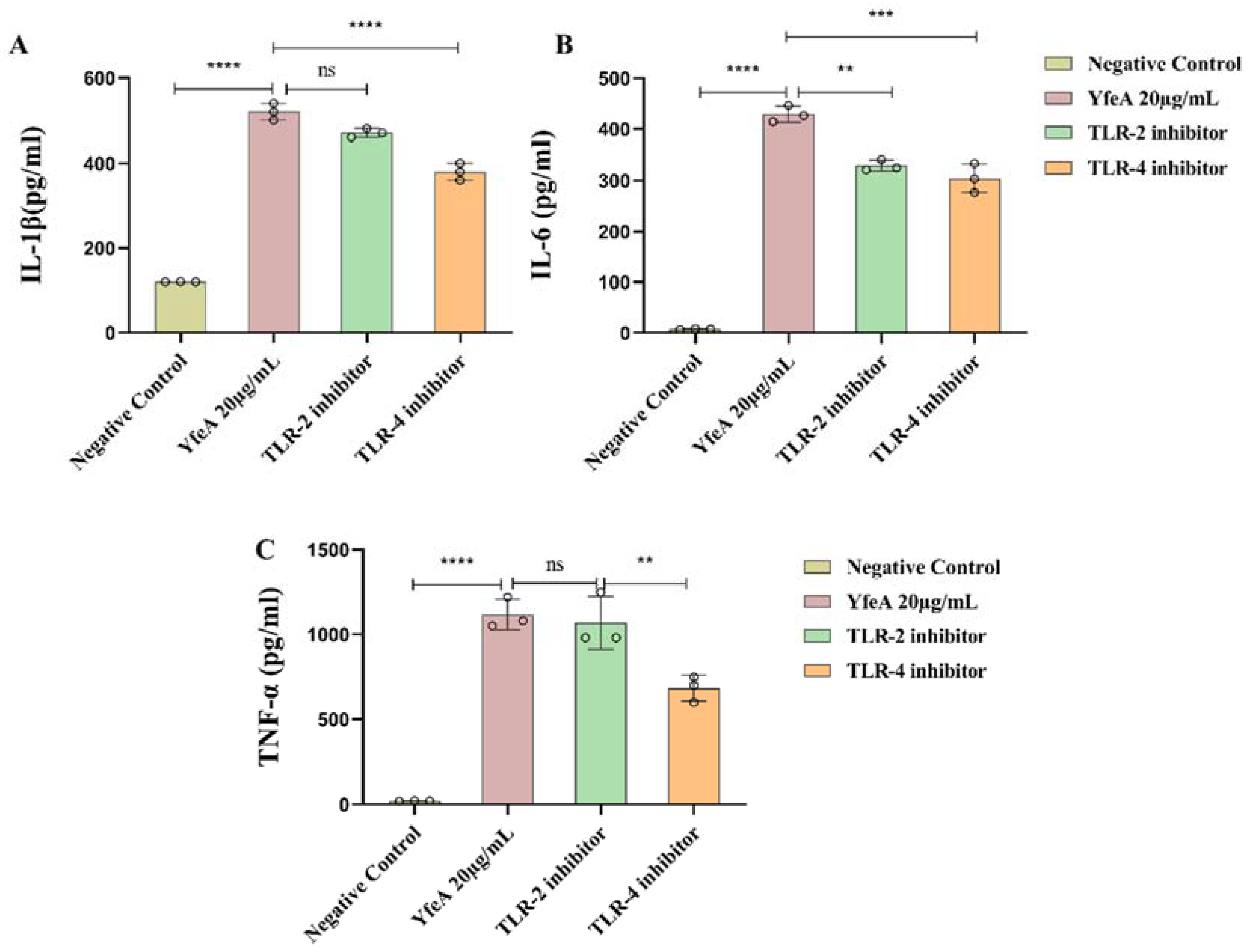
2.6. TLR2 and TLR4 Play a Regulatory Role in the Secretion of Pro-Inflammatory Cytokines in Macrophages
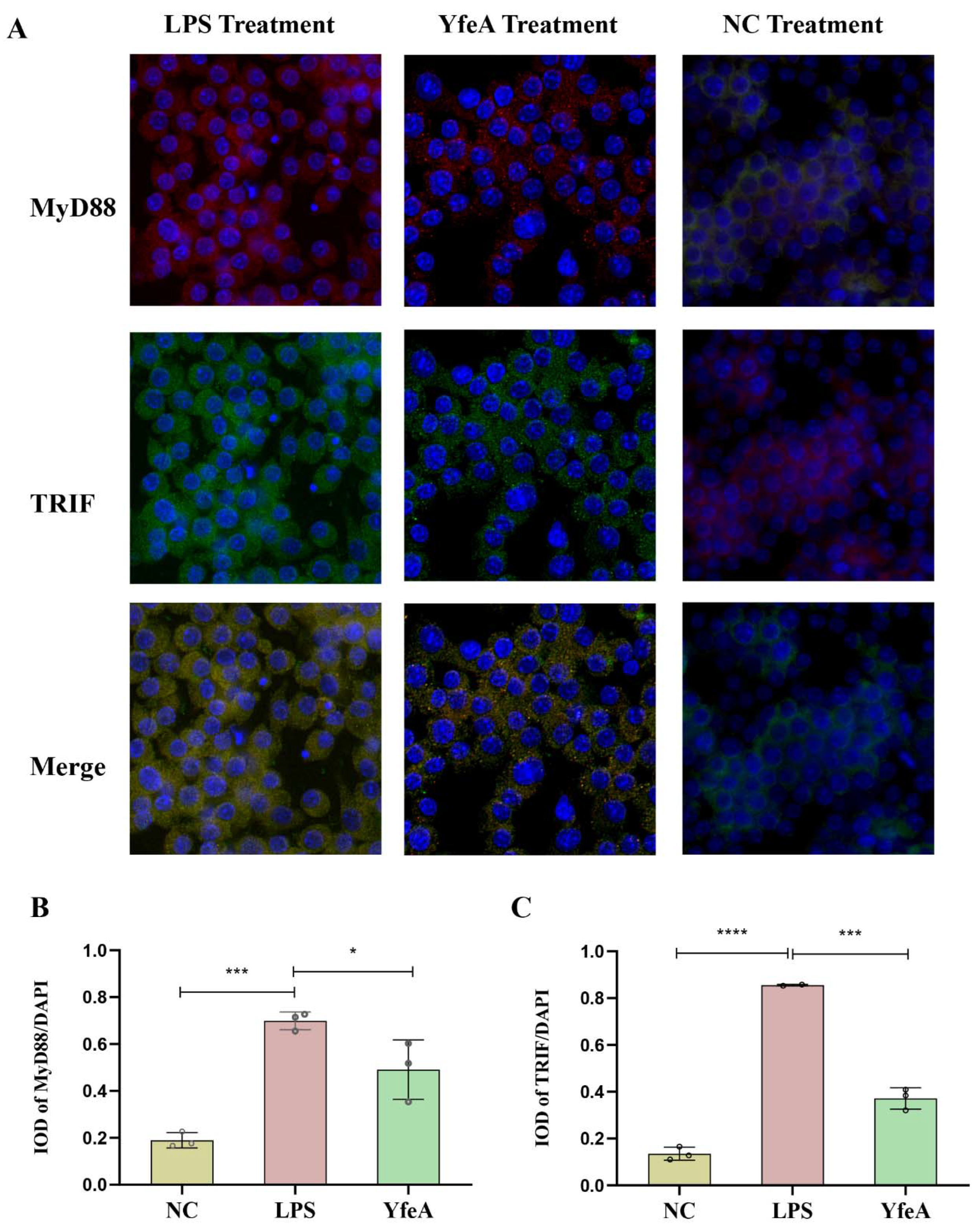
2.7. YfeA Induces Upregulation and Concentration of the Adaptor Protein MyD88 and TRIF in Macrophages
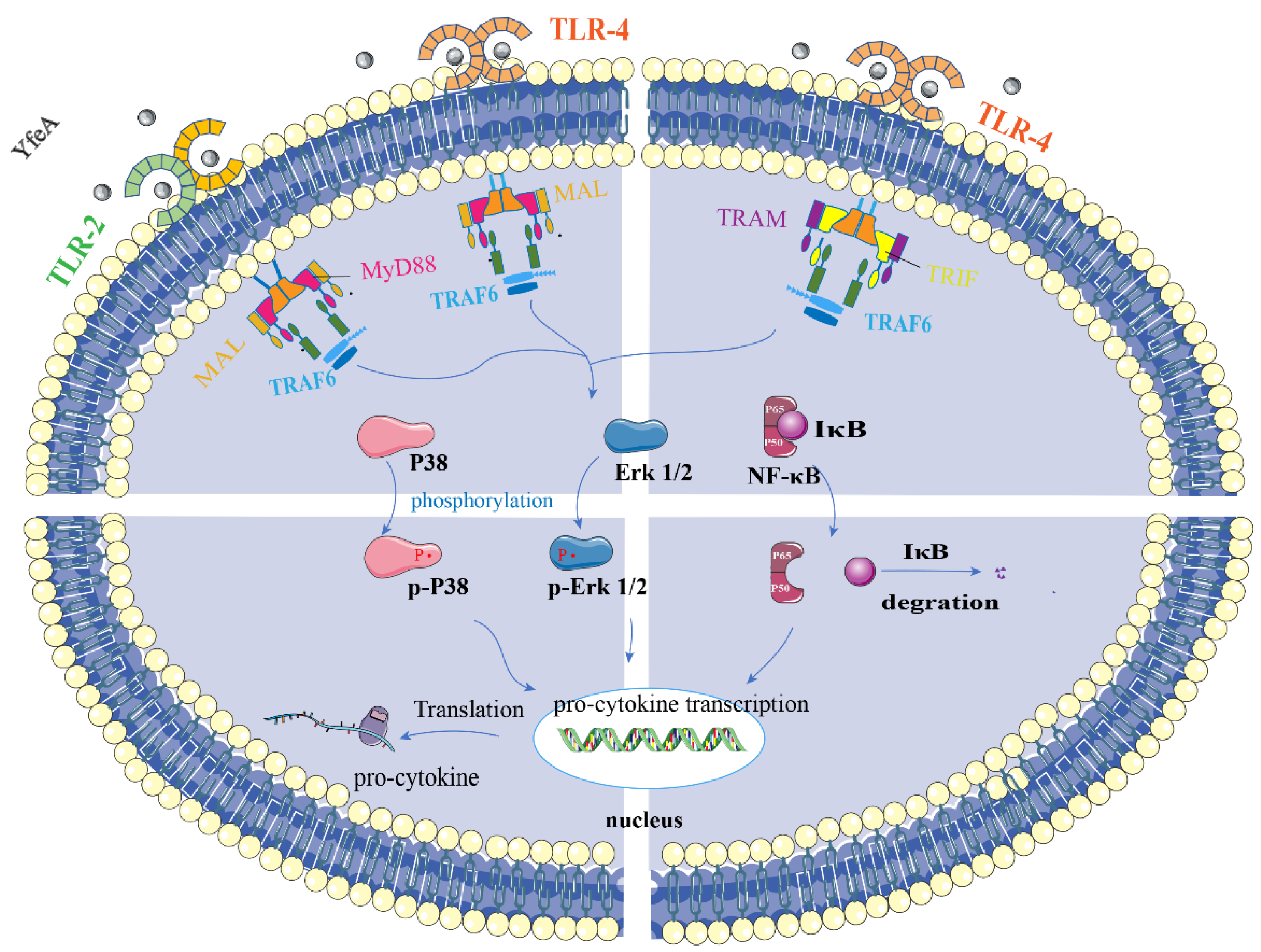
2.8. YfeA Activates Both MAP Kinases and NF-κB Signal Pathways of RAW 264.7 Cells
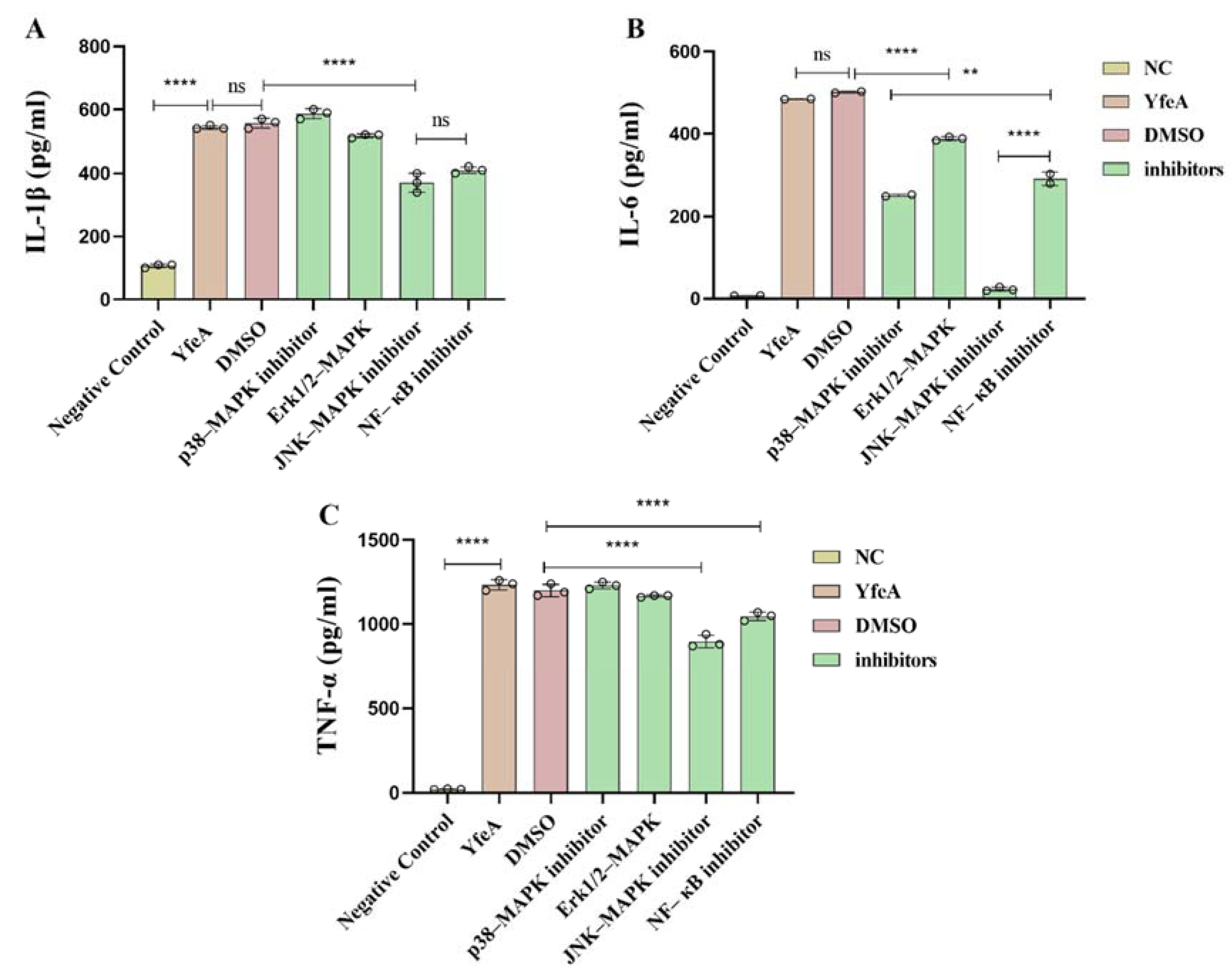
2.9. YfeA Induces Pro-Inflammatory Cytokines Production through MAPKs and NF-κB Signal Pathways
3. Discussion
4. Materials and Methods
4.1. Bacterial and Cell Culture Condition
4.2. Expression/Purification of Recombinant YfeA Protein and Removal of Endotoxins
4.3. Immunization and Mouse Challenge
4.4. The Capacity of YfeA to Stimulate Cytokine Production
4.5. Measurement of ROS Production
4.6. Toll-like Receptors 2 (TLR2) and 4 (TLR4) Blocking Assay
4.7. Indirect Immunofluorescence (IF) Assay for Detection of Adaptor Protein MyD88 and TRIF
4.8. Western Blotting Assay for Induction of Pro-Inflammatory Cytokines by YfeA
4.9. Effect of Inhibitors on the MAPK and NF-κB Signal Pathways
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dai, K.; He, L.; Chang, Y.-F.; Cao, S.; Zhao, Q.; Huang, X.; Wu, R.; Huang, Y.; Yan, Q.; Han, X.; et al. Basic Characterization of Natural Transformation in a Highly Transformable Haemophilus parasuis Strain SC1401. Front. Cell. Infect. Microbiol. 2018, 8, 32. [Google Scholar] [CrossRef] [PubMed]
- Espindola, J.P.; Balbinott, N.; Gressler, L.T.; Machado, G.; Klein, C.S.; Rebelatto, R.; Gutierrez Martin, C.B.; Kreutz, L.C.; Schryvers, A.B.; Frandoloso, R. Molecular serotyping of clinical strains of Haemophilus (Glaesserella) parasuis brings new insights regarding Glasser’s disease outbreaks in Brazil. PeerJ 2019, 7, e6817. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Zhou, N.; An, J.; Zhang, J.; Wang, M.; Li, Y.; Jiang, P. Haemophilus parasuis infection in 3D4/21 cells induces autophagy through the AMPK pathway. Cell. Microbiol. 2019, 21, e13031. [Google Scholar] [CrossRef]
- Zhao, Y.; Guo, L.; Li, J.; Huang, X.; Fang, B. Characterization of antimicrobial resistance genes in Haemophilus parasuis isolated from pigs in China. PeerJ 2018, 6, e4613. [Google Scholar] [CrossRef]
- Hensel, M.; Shea, J.E.; Gleeson, C.; Jones, M.; Holden, D.W.; Hensel, M.; Shea, J.E.; Gleeson, C.; Jones, M.D.; Dalton, E.; et al. Simultaneous identification of bacterial virulence genes by negative selection. Science 1995, 269, 400–403. [Google Scholar] [CrossRef] [PubMed]
- Garmory, H.S.; Titball, R.W. ATP-binding cassette transporters are targets for the development of antibacterial vaccines and therapies. Infect. Immun. 2004, 72, 6757–6763. [Google Scholar] [CrossRef] [PubMed]
- Crosa, J.H.; Mey, A.R.; Payne, S.M. Iron Transport in Bacteria; ASM Press: Washington, DC, USA, 2004. [Google Scholar]
- Bearden, S.W.; Perry, R.D. The Yfe system of Yersinia pestis transports iron and manganese and is required for full virulence of plague. Mol. Microbiol. 2010, 32, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Radka, C.D.; Delucas, L.J.; Wilson, L.S.; Lawrenz, M.B.; Aller, S.G. Crystal structure of Yersinia pestis virulence factor YfeA reveals two polyspecific metal-binding sites. Acta Crystallogr. Sect. D Struct. Biol. 2017, 73, 557–572. [Google Scholar] [CrossRef]
- Zaharik, M.L.; Cullen, V.L.; Fung, A.M.; Libby, S.J.; Choy, S.K.; Coburn, B.; Kehres, D.G.; Maguire, M.E.; Fang, F.C.; Finlay, B.B. The Salmonella enterica serovar typhimurium divalent cation transport systems MntH and SitABCD are essential for virulence in an Nramp1G169 murine typhoid model. Infect. Immun. 2004, 72, 5522–5525. [Google Scholar] [CrossRef]
- Janakiraman, A.; Slauch, J.M. The putative iron transport system SitABCD encoded on SPI1 is required for full virulence of Salmonella typhimurium. Mol. Microbiol. 2000, 35, 1146–1155. [Google Scholar] [CrossRef] [Green Version]
- Sack, M.; Baltes, N. Identification of novel potential virulence-associated factors in Haemophilus parasuis. Vet. Microbiol. 2009, 136, 382–386. [Google Scholar] [CrossRef] [PubMed]
- Papian, S.; Mobarez, A.M.; Khoramabadi, N.; Abdol, M.M.; Abadi, A. Investigating the role of L. pnuemophila LPS derivatives in formation of specific cell-mediated immune responses against the pathogen—ScienceDirect. Microb. Pathog. 2020, 147, 104396. [Google Scholar] [CrossRef] [PubMed]
- Jarma, E.; Regassa, L.B. Growth phase mediated regulation of the Actinobacillus pleuropneumoniae ApxI and ApxII toxins. Microb. Pathog. 2004, 36, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.; Ahn, J.; Yoo, S.; Hong, J.; Lee, E.; Yoon, I.; Jung, J.-K.; Lee, H. Identification of novel immunogenic proteins in pathogenic Haemophilus parasuis based on genome sequence analysis. Vet. Microbiol. 2011, 148, 89–92. [Google Scholar] [CrossRef]
- Iwasaki, A.; Medzhitov, R. Regulation of Adaptive Immunity by the Innate Immune System. Science 2010, 327, 291–295. [Google Scholar] [CrossRef]
- Gordon, A.S.; Lin, H.H.; Stacey, M.; Taylor, R.; Martínezmota, L.; Brown, M.G. Macrophage receptors and immune recognition. Annu. Rev. Immunol. 2004, 23, 901–944. [Google Scholar]
- Chu, H.; Mazmanian, S.K. Innate immune recognition of the microbiota promotes host-microbial symbiosis. Nat. Immunol. 2013, 14, 668–675. [Google Scholar] [CrossRef]
- Benoit, M.; Desnues, B.; Mege, J.L. Macrophage polarization in bacterial infections. J. Immunol. 2008, 181, 3733–3739. [Google Scholar] [CrossRef]
- Tan, H.Y.; Wang, N.; Li, S.; Hong, M.; Wang, X.; Feng, Y. The Reactive Oxygen Species in Macrophage Polarization: Reflecting Its Dual Role in Progression and Treatment of Human Diseases. Oxidative Med. Cell. Longev. 2016, 2016, 2795090. [Google Scholar] [CrossRef]
- Iwasaki, A.; Medzhitov, R. Toll-like receptor control of the adaptive immune responses. Nat. Immunol. 2004, 5, 987–995. [Google Scholar] [CrossRef]
- Wi, H.J.; Yan, J.; Choi, M.H.; Hong, S.T.; Bae, Y.M. Predominance of IL-10 and TGF-β production from the mouse macrophage cell line, RAW264.7, in response to crude antigens from Clonorchis sinensis. Cytokine 2012, 59, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid Redox Signal 2014, 20, 7. [Google Scholar] [CrossRef]
- Xl, A.; Xc, A.; Hong, Z.A.; Hl, A.; Hfa, B. Glaesserella parasuis induces inflammatory response in 3D4/21 cells through activation of NLRP3 inflammasome signaling pathway via ROS-ScienceDirect. Vet. Microbiol. 2021, 256, 109057. [Google Scholar]
- Yamamoto, M.; Sato, S.; Hemmi, H.; Hoshino, K.; Akira, S. Role of Adaptor TRIF in the MyD88-Independent Toll-Like Receptor Signaling Pathway. Science 2003, 301, 640–643. [Google Scholar] [PubMed]
- Dai, K.; Ma, X.; Yang, Z.; Chang, Y.F.; Cao, S.; Zhao, Q.; Huang, X.; Wu, R.; Huang, Y.; Yan, Q.; et al. Polyamine Transport Protein PotD Protects Mice against Haemophilus parasuis and Elevates the Secretion of Pro-Inflammatory Cytokines of Macrophage via JNK-MAPK and NF-κB Signal Pathways through TLR4. Vaccines 2019, 7, 216. [Google Scholar] [CrossRef]
- Lin, W.H.; Shih, H.C.; Lin, C.F.; Yang, C.Y.; Lin, C.N.; Chiou, M.T. Genotypic analyses and virulence characterization of Glaesserella parasuis isolates from Taiwan. Other 2019, 7, e6960. [Google Scholar]
- Mccaig, W.D.; Loving, C.L.; Hughes, H.R.; Brockmeier, S.L.; Alain, C. Characterization and Vaccine Potential of Outer Membrane Vesicles Produced by Haemophilus parasuis. PLoS ONE 2016, 11, e0149132. [Google Scholar] [CrossRef]
- Fetherston, J.D.; Mier, I.; Truszczynska, H.; Perry, R.D. The Yfe and Feo Transporters Are Involved in Microaerobic Growth and Virulence of Yersinia pestis in Bubonic Plague. Infect. Immun. 2012, 80, 3880. [Google Scholar] [CrossRef]
- Wang, Z.; Qin, Z.; Wei, H.; Wen, X.; Wen, Y. Prevalence and seroepidemiology of Haemophilus parasuis in Sichuan province, China. PeerJ 2017, 5, e3379. [Google Scholar] [CrossRef]
- Dai, K.; Yang, Z.; Ma, X.; Chang, Y.F.; Cao, S.; Zhao, Q.; Huang, X.; Wu, R.; Huang, Y.; Xia, J. Deletion of Polyamine Transport Protein PotD Exacerbates Virulence in Glaesserella (Haemophilus) parasuis in the Form of Non-biofilm-generated Bacteria in a Murine Acute Infection Model. Virullence 2021, 12, 540–546. [Google Scholar] [CrossRef]
- Zhou, M.; Guo, Y.; Zhao, J.; Hu, Q.; Hu, Y.; Zhang, A.; Chen, H.; Jin, M. Identification and characterization of novel immunogenic outer membrane proteins of Haemophilus parasuis serovar 5. Vaccine 2009, 27, 5271–5277. [Google Scholar] [CrossRef]
- Na, G.; Hje, B.; Hjk, B.; Ghp, B.; Hjs, B.; Jin, B. Immune-enhancing activity of Hydrangea macrophylla subsp. serrata leaves through TLR4/ROS-dependent activation of JNK and NF-κB in RAW264.7 cells and immunosuppressed mice-ScienceDirect. J. Funct. Foods 2020, 73, 104139. [Google Scholar]
- Jing, W.; Zhang, X.; Sun, W.; Hou, X.; Yao, Z.; Zhu, Y. CRISPR/CAS9-Mediated Genome Editing of miRNA-155 Inhibits Proinflammatory Cytokine Production by RAW264.7 Cells. Biomed Res. Int. 2015, 2015, 326042. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PCR Primers | Primer Sequences (50→30) | Products (bp) |
|---|---|---|
| YfeA Expression Primers | ||
| P1 (YfeA-pET28a-F) | cagcaaatgggtcgcggatccCAGCAGTTTAAAGTGGTCACCAC | |
| P2 (YfeA-pET28aR) | gtggtggtggtggtgctcgagTTATTTTTCAAATCCAGCAACAAT | 819 |
| qPCR primers | ||
| P3 (GAPDH-F) | GTGTTCCTACCCCCAATGTG | |
| P4 (GAPDH-R) | CATCGAAGGTGGAAGAGTGG | 189 |
| P5 (IL-1β-F) | GGGCCTCAAAGGAAAGAATC | |
| P6 (IL-1β-R) | TACCAGTTGGGGAACTCTGC | 183 |
| P7 (IL-4-F) | TCTTGATAAACTTAATTGTCTCT | |
| P8 (IL-4-R) | GCAGGATGACAACTAGCTGGG | 160 |
| P9 (IL-6-F) | GGGACTGATGCTGGTGACAA | |
| P10 (IL-6-R) | TCCACGATTTCCCAGAGAACA | 147 |
| P11 (TNF-α-F) | CGTCAGCCGATTTGCTATCT | |
| P12 (TNF-α-F) | CTTGGGCAGATTGACCTCAG | 184 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.; Tang, X.; Wang, K.; Dai, K.; Chang, Y.-F.; Du, S.; Zhao, Q.; Huang, X.; Wu, R.; Yan, Q.; et al. Metal Ion Periplasmic-Binding Protein YfeA of Glaesserella parasuis Induces the Secretion of Pro-Inflammatory Cytokines of Macrophages via MAPK and NF-κB Signaling through TLR2 and TLR4. Int. J. Mol. Sci. 2022, 23, 9627. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23179627
Yang Z, Tang X, Wang K, Dai K, Chang Y-F, Du S, Zhao Q, Huang X, Wu R, Yan Q, et al. Metal Ion Periplasmic-Binding Protein YfeA of Glaesserella parasuis Induces the Secretion of Pro-Inflammatory Cytokines of Macrophages via MAPK and NF-κB Signaling through TLR2 and TLR4. International Journal of Molecular Sciences. 2022; 23(17):9627. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23179627
Chicago/Turabian StyleYang, Zhen, Xinwei Tang, Kang Wang, Ke Dai, Yung-Fu Chang, Senyan Du, Qin Zhao, Xiaobo Huang, Rui Wu, Qigui Yan, and et al. 2022. "Metal Ion Periplasmic-Binding Protein YfeA of Glaesserella parasuis Induces the Secretion of Pro-Inflammatory Cytokines of Macrophages via MAPK and NF-κB Signaling through TLR2 and TLR4" International Journal of Molecular Sciences 23, no. 17: 9627. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23179627