
Structural Characteristics of the 5′-Terminal Region of Mouse p53 mRNA and Identification of Proteins That Bind to This mRNA Region

Abstract
:1. Introduction
2. Results
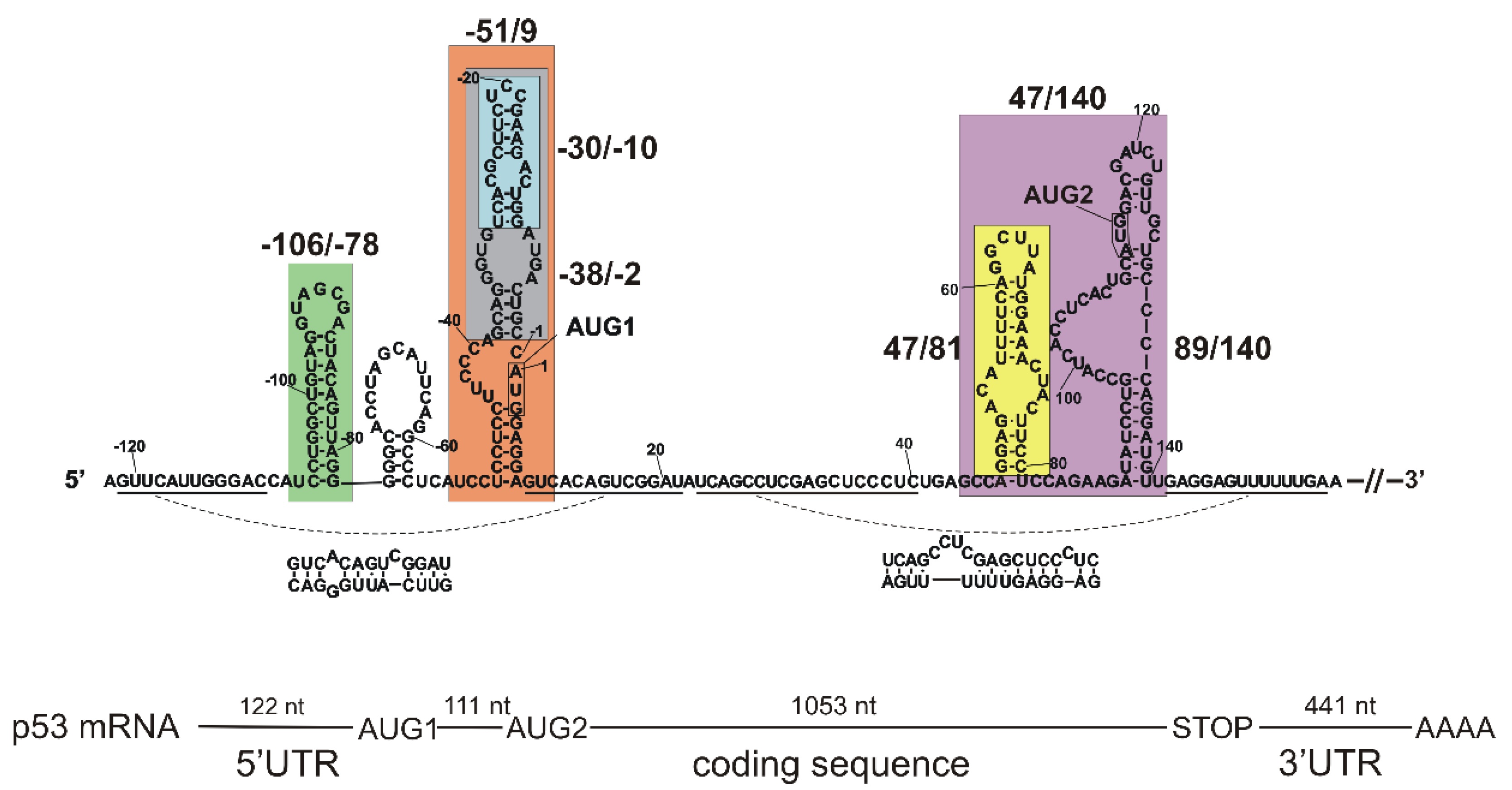
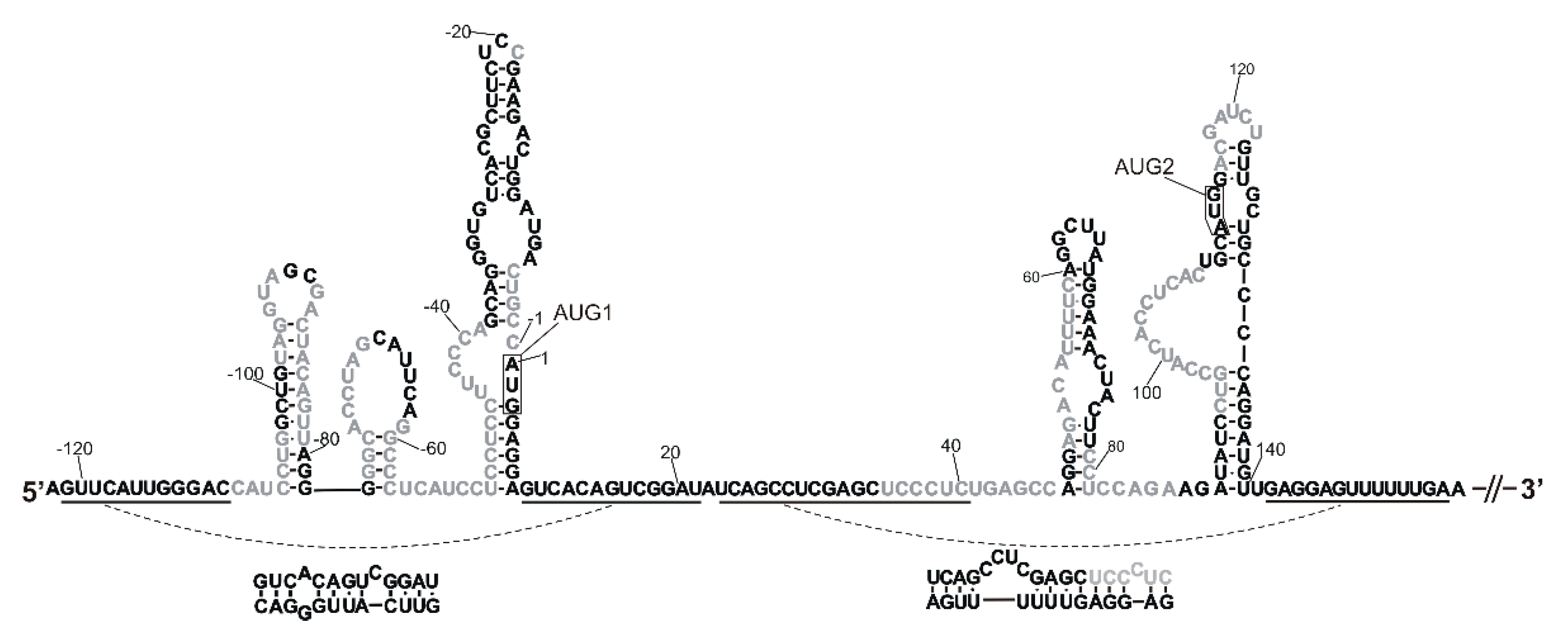
2.1. Tertiary Structure of the 5′-Terminal Region of Mouse p53 mRNA
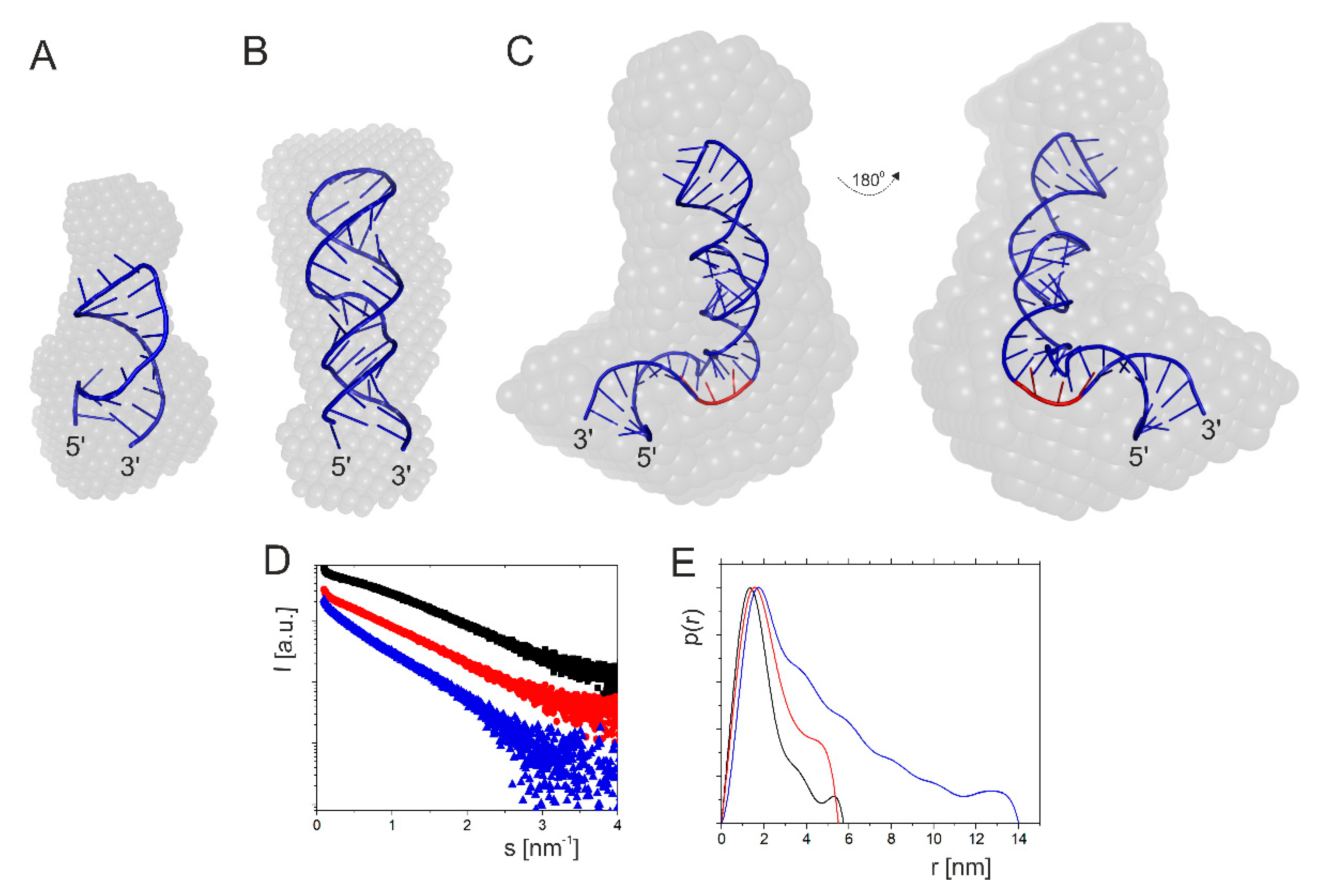
2.1.1. Hairpin Domain with AUG1 Initiation Codon
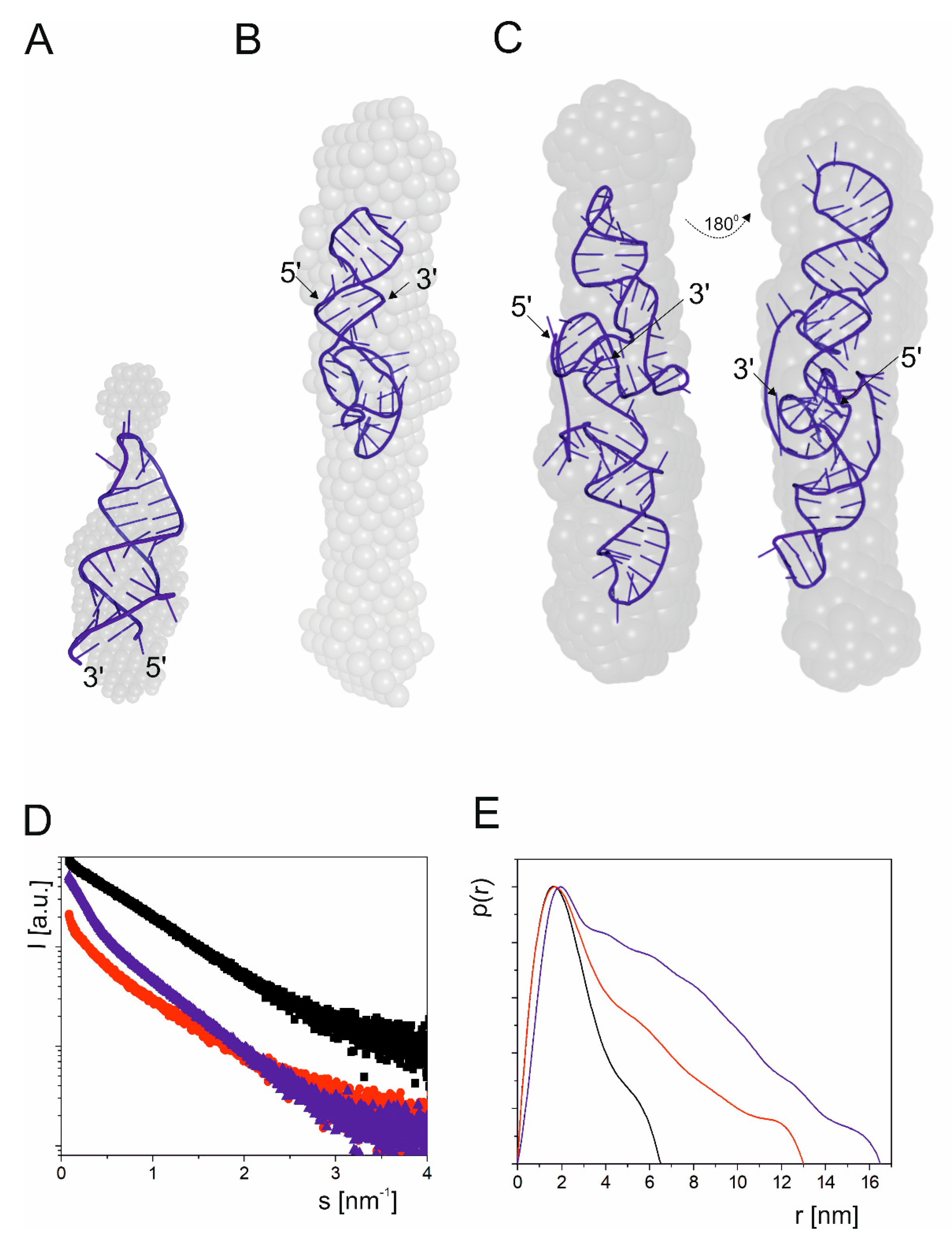
2.1.2. Two-Hairpins Domain with AUG2 Initiation Codon
2.1.3. Influence of Magnesium Ions on the Structure of Short Hairpin Motifs
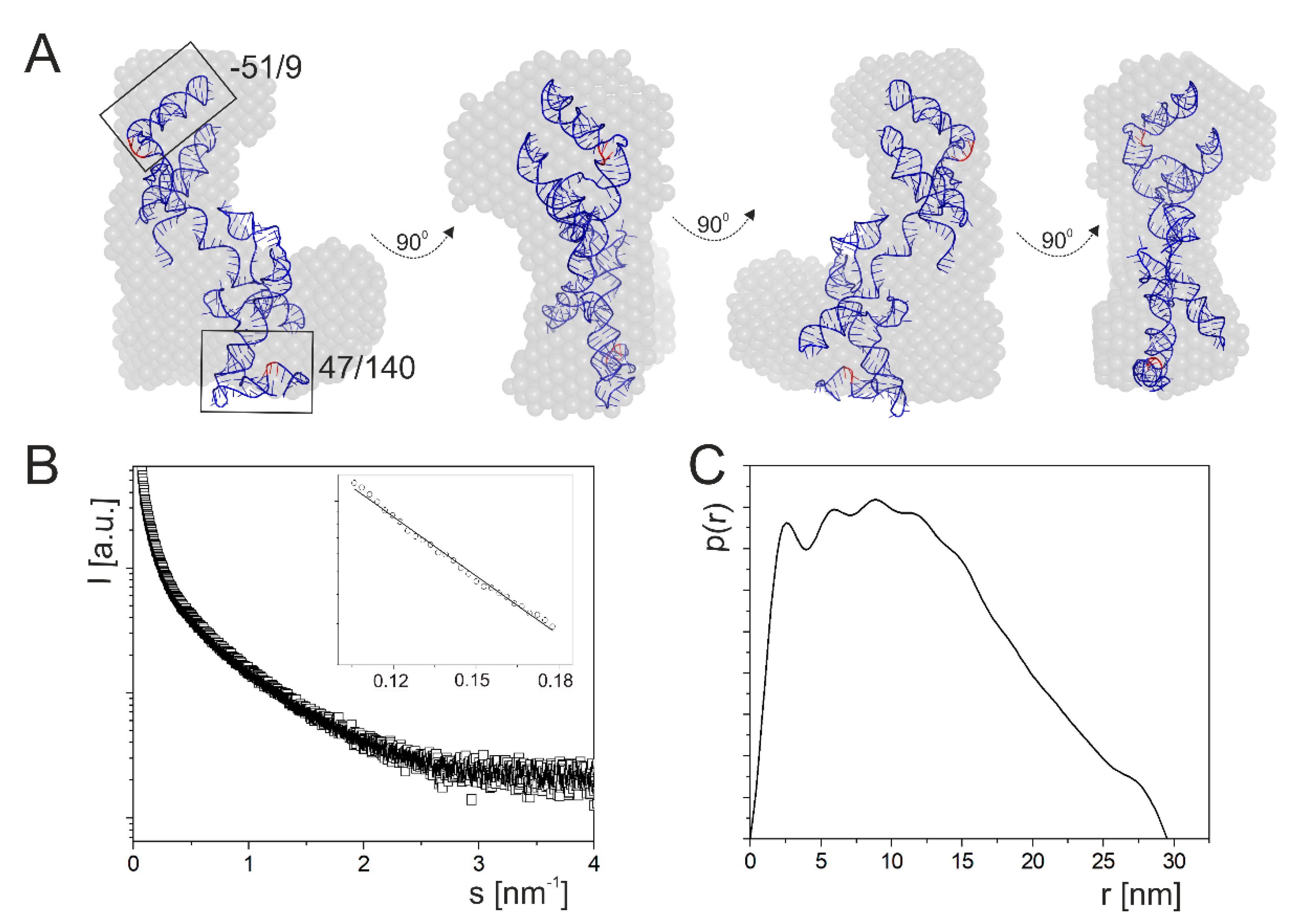
2.2. A Tertiary Structure Model of the 5′-Terminus of p53 mRNA
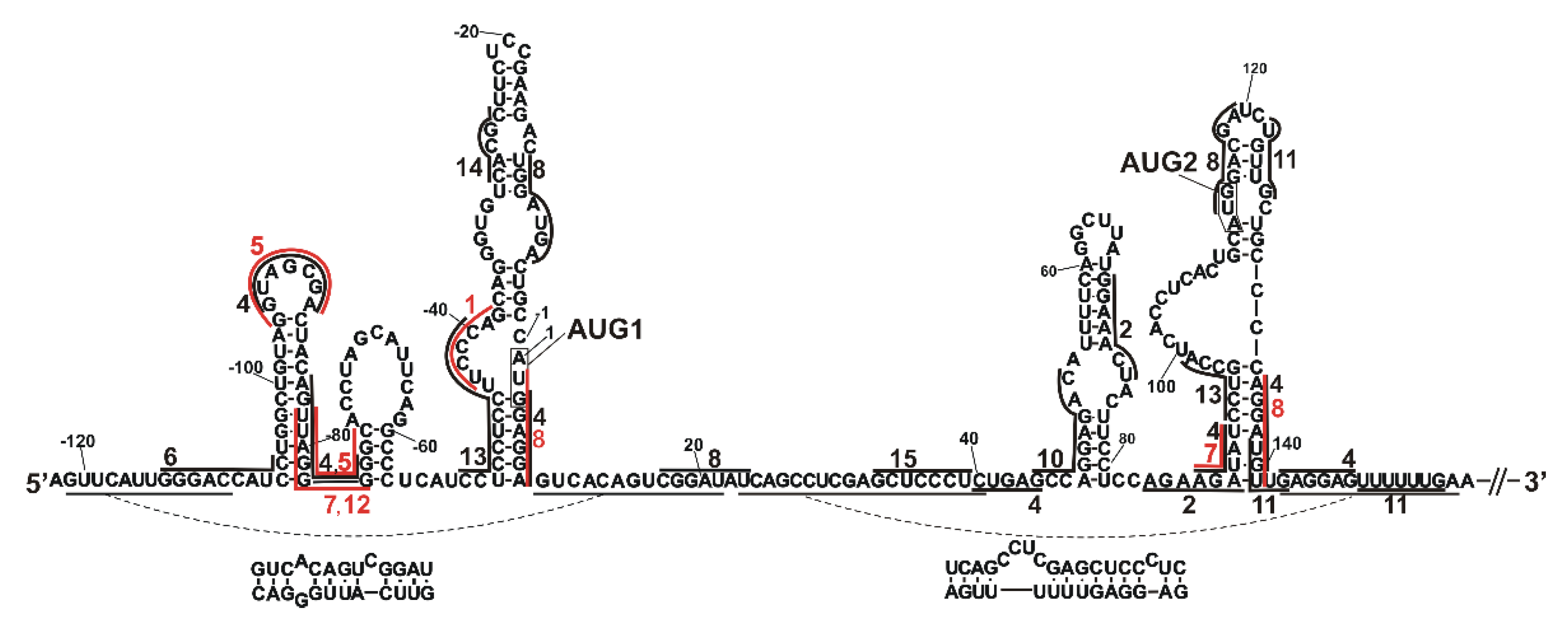
2.3. Determination of Sites Accessible to Hybridization of Complementary Oligonucleotides within the 5′-Terminal Region of Mouse p53 mRNA
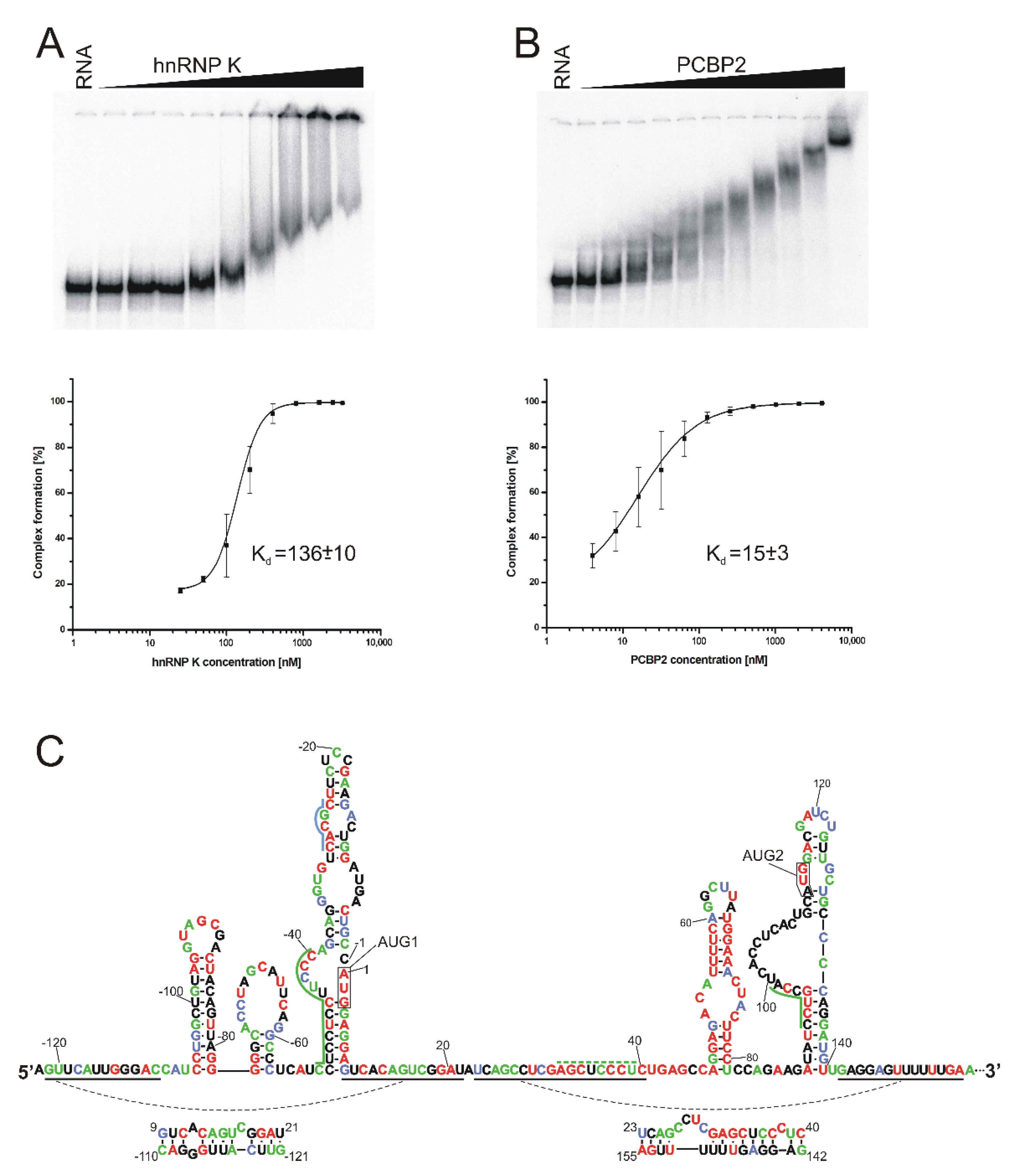
2.4. Identification of Proteins That Bind to the 5′-Terminal Region of Mouse p53 mRNA
2.5. hnRNP K and PCBP2 Proteins Efficiently Bind to the 5′-Terminal Region of p53 mRNA
3. Discussion
4. Materials and Methods
4.1. DNA Template Constructs and RNA Synthesis
4.2. Mapping of RNA Accessibility to Hybridization with DNA 6-Mer Libraries and RNase H Cleavage
4.3. Preparation of Cytoplasmic Lysate
4.4. RNA-Centric Affinity Chromatography
4.5. Mass Spectrometry Analysis
4.6. CD Measurements
4.7. Small Angle X-ray Scattering
4.8. Generation of RNA Fragments’ Tertiary Structure Models
4.9. Protein Overexpression and Purification
4.10. Electrophoretic Mobility Shift Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, J.; Zhang, C.; Hu, W.; Feng, Z. Tumor suppressor p53 and metabolism. J. Mol. Cell. Biol. 2019, 11, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Sammons, M.A.; Nguyen, T.T.; McDade, S.S.; Fischer, M. Tumor suppressor p53: From engaging DNA to target gene regulation. Nucleic Acids Res. 2020, 48, 8848–8869. [Google Scholar] [CrossRef] [PubMed]
- Vadivel Gnanasundram, S.; Bonczek, O.; Wang, L.; Chen, S.; Fahraeus, R. p53 mRNA Metabolism Links with the DNA Damage Response. Genes 2021, 12, 1446. [Google Scholar] [CrossRef] [PubMed]
- Marcel, V.; Hainaut, P. p53 isoforms—A conspiracy to kidnap p53 tumor suppressor activity? Cell. Mol. Life Sci. 2009, 66, 391–406. [Google Scholar] [CrossRef]
- Engelmann, D.; Pützer, B.M. Emerging from the shade of p53 mutants: N-terminally truncated variants of the p53 family in EMT signalling and cancer progression. Sci. Signal. 2014, 7, re9. [Google Scholar] [CrossRef]
- Anbarasan, T.; Bourdon, J.C. The Emerging Landscape of p53 Isoforms in Physiology, Cancer and Degenerative Diseases. Int. J. Mol. Sci. 2019, 20, 6257. [Google Scholar] [CrossRef]
- Hafner, A.; Bulyk, M.L.; Jambhekar, A.; Lahav, G. The multiple mechanisms that regulate p53 activity and cell fate. Nat. Rev. Mol. Cell. Biol. 2019, 20, 199–210. [Google Scholar] [CrossRef]
- Bourdon, J.C. p53 Family isoforms. Curr. Pharm. Biotechnol. 2007, 8, 332–336. [Google Scholar] [CrossRef]
- Marcel, V.; Dichtel-Danjoy, M.L.; Sagne, C.; Hafsi, H.; Ma, D.; Ortiz-Cuaran, S.; Olivier, M.; Hall, J.; Mollereau, B.; Hainaut, P.; et al. Biological functions of p53 isoforms through evolution: Lessons from animal and cellular models. Cell Death Differ. 2011, 18, 1815–1824. [Google Scholar] [CrossRef]
- Joruiz, S.M.; Bourdon, J.C. p53 Isoforms: Key Regulators of the Cell Fate Decision. Cold Spring Harb. Perspect. Med. 2016, 6, a026039. [Google Scholar] [CrossRef] [Green Version]
- Ungewitter, E.; Scrable, H. Delta40p53 controls the switch from pluripotency to differentiation by regulating IGF signaling in ESCs. Genes Dev. 2010, 24, 2408–2419. [Google Scholar] [CrossRef] [PubMed]
- Terrier, O.; Bourdon, J.C.; Rosa-Calatrava, M. p53 protein isoforms: Key regulators in the front line of pathogen infections? PLoS Pathog. 2013, 9, e1003246. [Google Scholar] [CrossRef] [PubMed]
- Halaby, M.J.; Yang, D.Q. p53 translational control: A new facet of p53 regulation and its implication for tumorigenesis and cancer therapeutics. Gene 2007, 395, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Vilborg, A.; Wilhelm, M.T.; Wiman, K.G. Regulation of tumor suppressor p53 at the RNA level. J. Mol. Med. 2010, 88, 645–652. [Google Scholar] [CrossRef]
- Marcel, V.; Nguyen Van Long, F.; Diaz, J.J. 40 Years of Research Put p53 in Translation. Cancers 2018, 10, 152. [Google Scholar] [CrossRef]
- Swiatkowska, A.; Dutkiewicz, M.; Zydowicz-Machtel, P.; Szpotkowska, J.; Janecki, D.M.; Ciesiolka, J. Translational Control in p53 Expression: The Role of 5′-Terminal Region of p53 mRNA. Int. J. Mol. Sci. 2019, 20, 5382. [Google Scholar] [CrossRef]
- Yang, D.Q.; Halaby, M.J.; Zhang, Y. The identification of an internal ribosomal entry site in the 5′-untranslated region of p53 mRNA provides a novel mechanism for the regulation of its translation following DNA damage. Oncogene 2006, 25, 4613–4619. [Google Scholar] [CrossRef]
- Ray, P.S.; Grover, R.; Das, S. Two internal ribosome entry sites mediate the translation of p53 isoforms. EMBO Rep. 2006, 7, 404–410. [Google Scholar] [CrossRef]
- Zydowicz-Machtel, P.; Swiatkowska, A.; Popenda, L.; Gorska, A.; Ciesiolka, J. Variants of the 5′-terminal region of p53 mRNA influence the ribosomal scanning and translation efficiency. Sci. Rep. 2018, 8, 1533. [Google Scholar] [CrossRef]
- Gorska, A.; Swiatkowska, A.; Dutkiewicz, M.; Ciesiolka, J. Modulation of p53 expression using antisense oligonucleotides complementary to the 5′-terminal region of p53 mRNA in vitro and in the living cells. PLoS ONE 2013, 8, e78863. [Google Scholar] [CrossRef] [Green Version]
- Swiatkowska, A.; Zydowicz, P.; Gorska, A.; Suchacka, J.; Dutkiewicz, M.; Ciesiolka, J. The Role of Structural Elements of the 5′-Terminal Region of p53 mRNA in Translation under Stress Conditions Assayed by the Antisense Oligonucleotide Approach. PLoS ONE 2015, 10, e0141676. [Google Scholar] [CrossRef]
- Blaszczyk, L.; Ciesiolka, J. Secondary structure and the role in translation initiation of the 5′-terminal region of p53 mRNA. Biochemistry 2011, 50, 7080–7092. [Google Scholar] [CrossRef] [PubMed]
- Gorska, A.; Blaszczyk, L.; Dutkiewicz, M.; Ciesiolka, J. Length variants of the 5′ untranslated region of p53 mRNA and their impact on the efficiency of translation initiation of p53 and its N-truncated isoform DeltaNp53. RNA Biol. 2013, 10, 1726–1740. [Google Scholar] [CrossRef] [PubMed]
- Swiatkowska, A.; Zydowicz, P.; Sroka, J.; Ciesiolka, J. The role of the 5′ terminal region of p53 mRNA in the p53 gene expression. Acta Biochim. Pol. 2016, 63, 645–651. [Google Scholar] [CrossRef]
- Haronikova, L.; Olivares-Illana, V.; Wang, L.; Karakostis, K.; Chen, S.; Fahraeus, R. The p53 mRNA: An integral part of the cellular stress response. Nucleic Acids Res. 2019, 47, 3257–3271. [Google Scholar] [CrossRef] [PubMed]
- Farnebo, M. Wrap53, a novel regulator of p53. Cell Cycle 2009, 8, 2343–2346. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, S.; Henriksson, S.; Corcoran, M.; Mendez-Vidal, C.; Wiman, K.G.; Farnebo, M. Wrap53, a natural p53 antisense transcript required for p53 induction upon DNA damage. Mol. Cell 2009, 33, 462–471. [Google Scholar] [CrossRef]
- Swiatkowska, A.; Dutkiewicz, M.; Machtel, P.; Janecki, D.M.; Kabacinska, M.; Zydowicz-Machtel, P.; Ciesiolka, J. Regulation of the p53 expression profile by hnRNP K under stress conditions. RNA Biol. 2020, 17, 1402–1415. [Google Scholar] [CrossRef]
- Janecki, D.M.; Swiatkowska, A.; Szpotkowska, J.; Urbanowicz, A.; Kabacińska, M.; Szpotkowski, K.; Ciesiołka, J. Poly(C)-binding Protein 2 Regulates the p53 Expression via Interactions with the 5′-Terminal Region of p53 mRNA. Int. J. Mol. Sci. 2021, 22, 13306. [Google Scholar] [CrossRef]
- Donehower, L.A.; Harvey, M.; Slagle, B.L.; McArthur, M.J.; Montgomery, C.A., Jr.; Butel, J.S.; Bradley, A. Mice deficient for p53 are developmentally normal but susceptible to spontaneous tumours. Nature 1992, 356, 215–221. [Google Scholar] [CrossRef]
- Kazantseva, M.; Mehta, S.; Eiholzer, R.A.; Hung, N.; Wiles, A.; Slatter, T.L.; Braithwaite, A.W. A mouse model of the Δ133p53 isoform: Roles in cancer progression and inflammation. Mamm. Genome 2018, 29, 831–842. [Google Scholar] [CrossRef]
- Szpotkowska, J.; Swiatkowska, A.; Ciesiolka, J. Length and secondary structure of the 5′ non-coding regions of mouse p53 mRNA transcripts—Mouse as a model organism for p53 gene expression studies. RNA Biol. 2019, 16, 25–41. [Google Scholar] [CrossRef]
- Graewert, M.A.; Svergun, D.I. Impact and progress in small and wide angle X-ray scattering (SAXS and WAXS). Curr. Opin. Struct. Biol. 2013, 23, 748–754. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Lois Pollack, L. SAXS studies of RNA: Structures, dynamics, and interactions with partners. Wiley Interdiscip. Rev. RNA 2016, 7, 512–526. [Google Scholar] [CrossRef]
- Popenda, M.; Szachniuk, M.; Antczak, M.; Purzycka, K.J.; Lukasiak, P.; Bartol, N.; Blazewicz, J.; Adamiak, R.W. Automated 3D structure composition for large RNAs. Nucleic Acids Res. 2012, 40, e112. [Google Scholar] [CrossRef] [PubMed]
- Antczak, M.; Popenda, M.; Zok, T.; Sarzynska, J.; Ratajczak, T.; Tomczyk, K.; Adamiak, R.W.; Szachniuk, M. New functionality of RNAComposer: An application to shape the axis of miR160 precursor structure. Acta Biochim. Pol. 2016, 63, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Michlewski, G.; Caceres, J.F. RNase-assisted RNA chromatography. RNA 2010, 16, 1673–1678. [Google Scholar] [CrossRef] [PubMed]
- Bakun, M.; Karczmarski, J.; Poznanski, J.; Rubel, T.; Rozga, M.; Malinowska, A.; Sands, D.; Hennig, E.; Oledzki, J.; Ostrowski, J.; et al. An integrated LC-ESI-MS platform for quantitation of serum peptide ladders. Application for colon carcinoma study. Proteom. Clin. Appl. 2009, 3, 932–946. [Google Scholar] [CrossRef]
- Candeias, M.M.; Malbert-Colas, L.; Powell, D.J.; Daskalogianni, C.; Maslon, M.M.; Naski, N.; Bourougaa, K.; Calvo, F.; Fåhraeus, R. P53 mRNA controls p53 activity by managing Mdm2 functions. Nat. Cell Biol. 2008, 10, 1098–1105. [Google Scholar] [CrossRef]
- Wrzesinski, J.; Legiewicz, M.; Ciesiołka, J. Mapping of accessible sites for oligonucleotide hybridization on hepatitis delta virus ribozymes. Nucleic Acids Res. 2000, 28, 1785–1793. [Google Scholar] [CrossRef] [Green Version]
- Swiatkowska, A.; Dutkiewicz, M.; Ciesiołka, J. Structural features of target RNA molecules greatly modulate the cleavage efficiency of trans-acting delta ribozymes. Biochemistry 2007, 46, 5523–5533. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Stewart, D.; Matlashewski, G. Regulation of human p53 activity and cell localization by alternative splicing. Mol. Cell. Biol. 2004, 24, 7987–7997. [Google Scholar] [CrossRef] [PubMed]
- Hoksza, D.; Svozil, D. Efficient RNA pairwise structure comparison by SETTER method. Bioinformatics 2012, 28, 1858–1864. [Google Scholar] [CrossRef]
- Reva, B.A.; Finkelstein, A.V.; Skolnick, J. What is the probability of a chance prediction of a protein structure with an rmsd of 6 A? Fold Des. 1998, 3, 141–147. [Google Scholar] [CrossRef]
- Lemkul, J.A.; Huang, J.; Roux, B.; MacKerell, A.D., Jr. An Empirical Polarizable Force Field Based on the Classical Drude Oscillator Model: Development History and Recent Applications. Chem. Rev. 2016, 116, 4983–5013. [Google Scholar] [CrossRef] [PubMed]
- Draper, D.E. A guide to ions and RNA structure. RNA 2004, 10, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Draper, D.E. RNA folding: Thermodynamic and molecular descriptions of the roles of ions. Biophys. J. 2008, 95, 5489–5495. [Google Scholar] [CrossRef]
- Wrzesinski, J.; Michałowski, D.; Ciesiołka, J.; Krzyzosiak, W.J. Specific RNA cleavages induced by manganese ions. FEBS Lett. 1995, 374, 62–68. [Google Scholar] [CrossRef]
- Szabat, M.; Pendzinski, T.; Czapik, T.; Kierzek, E.; Kierzek, R. Structural Aspects of the antiparallel and parallel duplexes formed by DNA, 2’-o-methyl RNA and RNA oligonucleotides. PLoS ONE 2015, 10, e0143354. [Google Scholar] [CrossRef]
- Yang, S.; Parisien, M.; Major, F.; Roux, B. RNA structure determination using SAXS data. J. Phys. Chem. B 2010, 114, 10039–10048. [Google Scholar] [CrossRef] [Green Version]
- Mir, K.U.; Southern, E.M. Determining the influence of structure on hybridization using oligonucleotide arrays. Nat Biotechnol. 1999, 17, 788–792. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Kim, W.; Lee, K.H.; Kim, S.H.; Lee, H.R.; Kim, H.J.; Jung, Y.; Choi, J.H.; Kim, K.T. hnRNP Q regulates translation of p53 in normal and stress conditions. Cell Death Differ. 2013, 20, 226–234. [Google Scholar] [CrossRef]
- Seo, J.Y.; Kim, D.Y.; Kim, S.H.; Kim, H.J.; Ryu, H.G.; Lee, J.; Lee, K.H.; Kim, K.T. Heterogeneous nuclear ribonucleoprotein (hnRNP) L promotes DNA damage-induced cell apoptosis by enhancing the translation of p53. Oncotarget 2017, 8, 51108–51122. [Google Scholar] [CrossRef] [PubMed]
- Takagi, M.; Absalon, M.J.; McLure, K.G.; Kastan, M.B. Regulation of p53 translation and induction after DNA damage by ribosomal protein L26 and nucleolin. Cell 2005, 123, 49–63. [Google Scholar] [CrossRef]
- Paz, I.; Kosti, I.; Ares, M., Jr.; Cline, M.; Mandel-Gutfreund, Y. RBPmap: A web server for mapping binding sites of RNA-binding proteins. Nucleic Acids Res. 2014, 42, W361-7. [Google Scholar] [CrossRef] [PubMed]
- Giudice, G.; Sánchez-Cabo, F.; Torroja, C.; Lara-Pezzi, E. ATtRACT-a database of RNA-binding proteins and associated motifs. Database 2016, 2016, baw035. [Google Scholar] [CrossRef]
- Ghisolfi-Nieto, L.; Joseph, G.; Puvion-Dutilleul, F.; Amalric, F.; Bouvet, P. Nucleolin is a sequence-specific RNA-binding protein: Characterization of targets on pre-ribosomal RNA. J. Mol. Biol. 1996, 260, 34–53. [Google Scholar] [CrossRef]
- Grover, R.; Ray, P.S.; Das, S. Polypyrimidine tract binding protein regulates IRES-mediated translation of p53 isoforms. Cell Cycle 2008, 7, 2189–2198. [Google Scholar] [CrossRef]
- Sharathchandra, A.; Lal, R.; Khan, D.; Das, S. Annexin A2 and PSF proteins interact with p53 IRES and regulate translation of p53 mRNA. RNA Biol. 2012, 9, 1429–1439. [Google Scholar] [CrossRef]
- Geuens, T.; Bouhy, D.; Timmerman, V. The hnRNP family: Insights into their role in health and disease. Hum. Genet. 2016, 135, 851–867. [Google Scholar] [CrossRef] [Green Version]
- Dahlberg, J.E.; Lund, E. Functions of the GTPase Ran in RNA export from the nucleus. Curr. Opin. Cell. Biol. 1998, 10, 400–408. [Google Scholar] [CrossRef]
- Roy, G.; De Crescenzo, G.; Khaleghpour, K.; Kahvejian, A.; O’Connor-McCourt, M.; Sonenberg, N. Paip1 interacts with poly(A) binding protein through two independent binding motifs. Mol. Cell. Biol. 2002, 22, 3769–3782. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, M.Q. Far upstream element binding protein 1: A commander of transcription, translation and beyond. Oncogene 2013, 32, 2907–2916. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Liu, Z.; Xu, Z.; Liu, J.; Zhang, J. High mobility group box 1 (HMGB1): A pivotal regulator of hematopoietic malignancies. J. Hematol. Oncol. 2020, 13, 91. [Google Scholar] [CrossRef]
- Stevenson, B.W.; Gorman, M.A.; Koach, J.; Cheung, B.B.; Marshall, G.M.; Parker, M.W.; Holien, J.K. A structural view of PA2G4 isoforms with opposing functions in cancer. J. Biol. Chem. 2020, 295, 16100–16112. [Google Scholar] [CrossRef]
- Attardi, L.D.; Jacks, T. The role of p53 in tumour suppression: Lessons from mouse models. Cell. Mol. Life Sci. 1999, 55, 48–63. [Google Scholar] [CrossRef]
- Lozano, G. Mouse models of p53 functions. Cold Spring Harb. Perspect. Biol. 2010, 2, a001115. [Google Scholar] [CrossRef]
- Jones, C.P.; Cantara, W.A.; Olson, E.D.; Musier-Forsyth, K. Small-angle X-ray scattering-derived structure of the HIV-1 5′ UTR reveals 3D tRNA mimicry. Proc. Natl. Acad. Sci. USA 2014, 111, 3395–3400. [Google Scholar] [CrossRef]
- Beckham, S.A.; Matak, M.Y.; Belousoff, M.J.; Venugopal, H.; Shah, N.; Vankadari, N.; Elmlund, H.; Nguyen, J.H.C.; Semler, B.L.; Wilce, M.C.J.; et al. Structure of the PCBP2/stem-loop IV complex underlying translation initiation mediated by the poliovirus type I IRES. Nucleic Acids Res. 2020, 48, 8006–8021. [Google Scholar] [CrossRef]
- Castillo-Martínez, J.; Fan, L.; Szewczyk, M.P.; Wang, Y.X.; Gallego, J. The low-resolution structural models of hepatitis C virus RNA subdomain 5BSL3.2 and its distal complex with domain 3′X point to conserved regulatory mechanisms within the Flaviviridae family. Nucleic Acids Res. 2022, 50, 2287–2301. [Google Scholar] [CrossRef]
- Schneider, C.; Sühnel, J. A molecular dynamics simulation study of coaxial stacking in RNA. J. Biomol. Struct. Dyn. 2000, 18, 345–352. [Google Scholar] [CrossRef]
- Choi, H.S.; Hwang, C.K.; Song, K.Y.; Law, P.Y.; Wei, L.N.; Loh, H.H. Poly(C)-binding proteins as transcriptional regulators of gene expression. Biochem. Biophys. Res. Commun. 2009, 380, 431–436. [Google Scholar] [CrossRef]
- Du, Z.; Fenn, S.; Tjhen, R.; James, T.L. Structure of a construct of a human poly(C)-binding protein containing the first and second KH domains reveals insights into its regulatory mechanisms. J. Biol. Chem. 2008, 283, 28757–28766. [Google Scholar] [CrossRef] [PubMed]
- Yoga, Y.M.; Traore, D.A.; Sidiqi, M.; Szeto, C.; Pendini, N.R.; Barker, A.; Leedman, P.J.; Wilce, J.A.; Wilce, M.C. Contribution of the first K-homology domain of poly(C)-binding protein 1 to its affinity and specificity for C-rich oligonucleotides. Nucleic Acids Res. 2012, 40, 5101–5114. [Google Scholar] [CrossRef] [PubMed]
- Manalastas-Cantos, K.; Konarev, P.V.; Hajizadeh, N.R.; Kikhney, A.G.; Petoukhov, M.V.; Molodenskiy, D.S.; Panjkovich, A.; Mertens, H.D.T.; Gruzinov, A.; Borges, C.; et al. ATSAS 3.0: Expanded functionality and new tools for small-angle scattering data analysis. J. Appl. Crystallogr. 2021, 54, 343–355. [Google Scholar] [CrossRef] [PubMed]
- Franke, D.; Svergun, D.I. DAMMIF, a program for rapid ab-initio shape determination in small-angle scattering. J. Appl. Crystallogr. 2009, 42, 342–346. [Google Scholar] [CrossRef]
- Konarev, P.V.; Petoukhov, M.V.; Svergun, D.I. Rapid automated superposition of shapes and macromolecular models using spherical harmonics. J. Appl. Crystallogr. 2016, 49, 953–960. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | -30/-10 | -38/-2 | -51/9 | 47/81 | 89/140 | 47/140 | -122/201 |
| Data collection | |||||||
| Instrument | P12, PETRA III | ||||||
| s range (nm−1) | 0.0088–5.0 | ||||||
| Wavelength (Å) | 1.24 | ||||||
| Temperature | 15 °C | ||||||
| Structural parameters | |||||||
| Rg (from p(r)) (nm) | 1.5 ± 0.05 | 1.94 ± 0.02 | 4.10 ± 0.02 | 2.078 ± 0.1 | 4.34 ± 0.02 | 6.17 ± 0.05 | 9.84 ± 0.04 |
| Rg (from Guinier) (nm) | 1.46 ± 0.03 | 1.96 ± 0.23 | 3.87 ± 0.44 | 2.099 ± 0.1 | 4.01 ± 0.31 | 6.02 ± 0.134 | 9.78 ± 0.34 |
| Rg (theoretical) (nm) | 1.275 | 1.744 | 2.484 | 1.73 | 1.985 | 5.498 | 9.535 |
| Dmax (nm) | 5.07 | 6.32 | 16.21 | 7.1 | 17.65 | 26.88 | 30.00 |
| Dmax (theoretical) | 4.169 | 6.145 | 8.006 | 6.404 | 6.674 | 19.87 | 34.84 |
| Porod volume estimate (nm3) | 9697 | 14,286 | 21,908 | 11,593 | 20,641 | 38,167 | 126,618 |
| Dry volume calculated from model (nm3) | 8081 | 11,905 | 19,051 | 9661 | 16,782 | 31,806 | 101,295 |
| Molecular mass determination | |||||||
| Contrast (Δρ × 1010 cm−2) | 3.047 | ||||||
| Experimental molecular weight (Da) | 7125 | 12,759 | 20,187 | 11,238 | 17,342 | 32,014 | 10,128 |
| Theoretical molecular weight (Da) | 6351 | 11,623.4 | 18,575.7 | 10,919.9 | 16,289.3 | 30,997.6 | 98,792.2 |
| Software used | |||||||
| Primary data reduction | PRIMUS | ||||||
| Data processing | PRIMUS | ||||||
| Quaternary structure modelling | SASREF | ||||||
| Computation od model intensities | CRYSOL | ||||||
| 3D graphics representation | PyMOL | ||||||
| No. | ID | Protein | Aliases | MS Spectra Number | Cell Line | MCF7 | HepG2 | HT-29 | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Doxo | - | + | - | + | - | + | |||||
| 1 | P09405 | Nucleolin | - | 40 | + | + | + | + | + | + | |
| 2 | P29341 | Polyadenylate-binding protein 1 | PABP1 | 24 | + | + | - | + | - | - | |
| 3 | P62827 | GTP-binding nuclear protein Ran | RAN | 22 | - | + | + | + | + | + | |
| 4 | P49312 | Heterogeneous nuclear ribonucleoprotein A1 | hnRNP A1 | 20 | + | + | + | + | + | + | |
| 5 | O88569 | Heterogeneous nuclear ribonucleoproteins A2/B1 | hnRNP A2/B1 | 18 | + | - | + | + | + | + | |
| 6 | P50580 | Proliferation-associated protein 2G4 | PA2G4, EBP1 | 17 | + | + | - | - | + | + | |
| 7 | Q60668 | Heterogeneous nuclear ribonucleoprotein D0 | hnRNP D0 | 13 | + | + | + | + | + | + | |
| 8 | P97855 | Ras GTPase-activating protein-binding protein 1 | G3BP1 | 12 | + | + | - | - | + | - | |
| 9 | P63158 | High mobility group protein B1 | HMGB1 | 9 | + | + | - | + | + | + | |
| 10 | Q8BG05 | Heterogeneous nuclear ribonucleoprotein A3 | hnRNP A3 | 8 | + | - | + | + | - | + | |
| 11 | Q91WJ8 | Far upstream element-binding protein 1 | FUBP1, FUP | 7 | - | - | + | + | + | + | |
| 12 | P17225 | Polypyrimidine tract-binding protein 1 | PTBP1 | 7 | + | + | + | + | + | + | |
| 13 | Q61990 | Poly(rC)-binding protein 2 | PCBP2 | 4 | + | + | - | - | + | + | |
| 14 | P61979 | Heterogeneous nuclear ribonucleoprotein K | hnRNP K | 4 | + | + | + | + | + | + | |
| 15 | Q8BHD7 | Polypyrimidine tract-binding protein 3 | PTBP3, ROD1 | 2 | - | + | + | + | + | + | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szpotkowska, J.; Szpotkowski, K.; Ciesiołka, J. Structural Characteristics of the 5′-Terminal Region of Mouse p53 mRNA and Identification of Proteins That Bind to This mRNA Region. Int. J. Mol. Sci. 2022, 23, 9709. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23179709
Szpotkowska J, Szpotkowski K, Ciesiołka J. Structural Characteristics of the 5′-Terminal Region of Mouse p53 mRNA and Identification of Proteins That Bind to This mRNA Region. International Journal of Molecular Sciences. 2022; 23(17):9709. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23179709
Chicago/Turabian StyleSzpotkowska, Joanna, Kamil Szpotkowski, and Jerzy Ciesiołka. 2022. "Structural Characteristics of the 5′-Terminal Region of Mouse p53 mRNA and Identification of Proteins That Bind to This mRNA Region" International Journal of Molecular Sciences 23, no. 17: 9709. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23179709