
Association of MMP-2 and MMP-9 Polymorphisms with Diabetes and Pathogenesis of Diabetic Complications

Abstract
:1. Properties, Functions and Regulation of the MMP
1.1. Matrix Metalloproteinases—General Characteristics
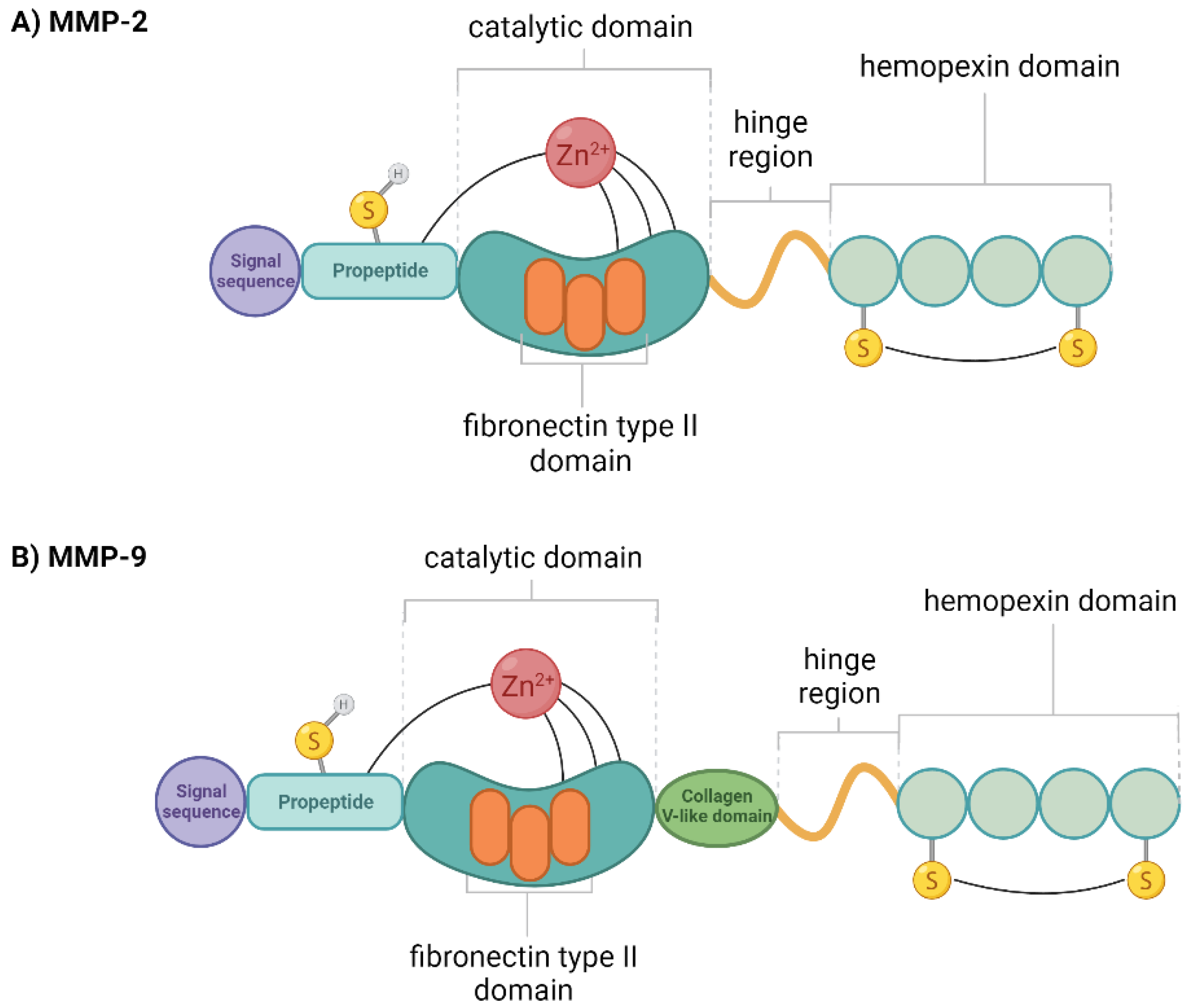
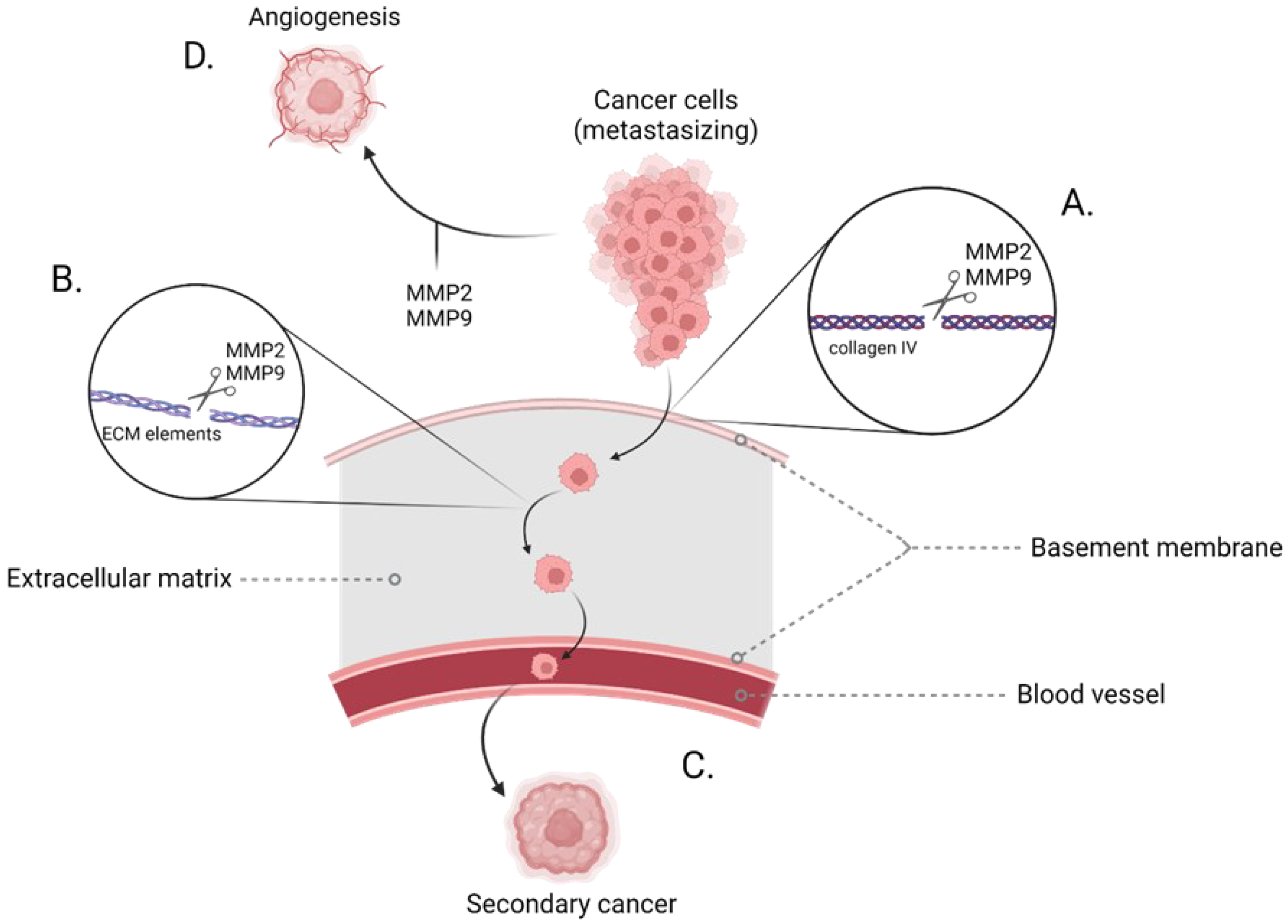
1.2. Gelatinase
1.3. MMP-2 and MMP-9
2. MMPs Polymorphisms Associated with Type 2 Diabetes
2.1. MMP-2
2.2. MMP-9
3. Gelatinase Polymorphisms in Diseases Accompanying Type 2 Diabetes
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cui, N.; Hu, M.; Khalil, R.A. Biochemical and biological attributes of matrix metalloproteinases. Prog. Mol. Biol. Transl. Sci. 2017, 147, 1–73. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Werb, Z. The many faces of metalloproteases: Cell growth, invasion, angiogenesis and metastasis. Trends. Cell. Biol. 2001, 11, 37–43. [Google Scholar] [CrossRef]
- Mittal, R.; Patel, A.P.; Debs, L.H.; Nguyen, D.; Patel, K.; Grati, M.; Mittal, J.; Yan, D.; Chapagain, P.; Liu, X.Z. Intricate functions of matrix metalloproteinases in physiological and pathological conditions. J. Cell. Physiol. 2016, 231, 2599–2621. [Google Scholar] [CrossRef] [PubMed]
- Shipley, J.M.; Doyle, G.A.R.; Fliszar, C.J.; Ye, Q.Z.; Johnson, L.L.; Shapiro, S.D.; Welgus, H.G.; Senior, R.M. The structural basis for the elastolytic activity of the 92-KDa and 72-KDa gelatinases: Role of the fibronectin type II-like repeats. J. Biol. Chem. 1996, 271, 4335–4341. [Google Scholar] [CrossRef] [PubMed]
- Koyama, H.; Iwata, H.; Kawahara, Y.; Iwase, H.; Kobayashi, S.; Fujii, Y.Y. Gelatinolytic activity of matrix metalloproteinase-2 and -9 in oesophageal carcinoma; A study using in situ zymography. Eur. J. Cancer. 2000, 36, 2164–2170. [Google Scholar] [CrossRef]
- Steffensen, B.; Wallon, U.M.; Overall, C.M. Extracellular matrix binding properties of recombinant fibronectin type II-like modules of human 72-KDa gelatinase/type IV collagenase: High affinity binding to native type I collagen but not native type IV collagen. J. Biol. Chem. 1995, 270, 11555–11566. [Google Scholar] [CrossRef]
- Mook, O.R.F.; Frederiks, W.M.; Van Noorden, C.J.F. The role of gelatinases in colorectal cancer progression and metastasis. Biochim. Biophys. Acta Rev. Cancer 2004, 1705, 69–89. [Google Scholar] [CrossRef]
- Fernandez-Garcia, B.; Eiró, N.; Marín, L.; González-Reyes, S.; González, L.O.; Lamelas, M.L.; Vizoso, F.J. Expression and prognostic significance of fibronectin and matrix metalloproteases in breast cancer metastasis. Histopathology 2014, 64, 512–522. [Google Scholar] [CrossRef]
- Li, H.; Huang, J.; Yang, B.; Xiang, T.; Yin, X.; Peng, W.; Cheng, W.; Wan, J.; Luo, F.; Li, H.; et al. Mangiferin exerts antitumor activity in breast cancer cells by regulating matrix metalloproteinases, epithelial to mesenchymal transition, and β-catenin signaling pathway. Toxicol. Appl. Pharmacol. 2013, 272, 180–190. [Google Scholar] [CrossRef]
- Liabakk, N.B.; Talbot, I.; Smith, R.A.; Wilkinson, K.; Balkwill, F. Matrix metalloprotease 2 (MMP-2) and matrix metalloprotease 9 (MMP-9) type IV collagenases in colorectal cancer. Cancer Res. 1996, 56, 190–196. [Google Scholar]
- Schnaper, H.W.; Kleinman, H.K. Regulation of cell function by extracellular matrix. Pediatric Nephrol. 1993, 7, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Lewalle, J.M.; Munaut, C.; Pichot, B.; Cataldo, D.; Baramova, E.; Foidart, J.M. Plasma membrane-dependent activation of gelatinase A in human vascular endothelial cells. J. Cell. Physiol. 1995, 165, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Bekes, E.M.; Schweighofer, B.; Kupriyanova, T.A.; Zajac, E.; Ardi, V.C.; Quigley, J.P.; Deryugina, E.I. Tumor-recruited neutrophils and neutrophil TIMP-free MMP-9 regulate coordinately the levels of tumor angiogenesis and efficiency of malignant cell intravasation. Am. J. Pathol. 2011, 179, 1455–1470. [Google Scholar] [CrossRef] [PubMed]
- Iochmann, S.; Bléchet, C.; Chabot, V.; Saulnier, A.; Amini, A.; Gaud, G.; Gruel, Y.; Reverdiau, P. Transient RNA silencing of tissue factor pathway inhibitor-2 modulates lung cancer cell invasion. Clin. Exp. Metastasis 2009, 26, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Safranek, J.; Pesta, M.; Holubec, L.; Kulda, V.; Dreslerova, J.; Vrzalova, J.; Topolcan, O.; Pesek, M.; Finek, J.; Treska, V. Expression of MMP-7, MMP-9, TIMP-1 and TIMP-2 MRNA in lung tissue of patients with non-small cell lung cancer (NSCLC) and benign pulmonary disease. Anticancer Res. 2009, 29, 2513–2517. [Google Scholar]
- Bastian, F.; Parmentier, G.; Roux, J.; Moretti, S.; Laudet, V.; Robinson-Rechavi, M. Bgee: Integrating and comparing heterogeneous transcriptome data among species. In Proceedings of the 5th International Workshop, DILS 2008, Evry, France, 25–27 June 2008; pp. 124–131. [Google Scholar]
- Su, A.I.; Wiltshire, T.; Batalov, S.; Lapp, H.; Ching, K.A.; Block, D.; Zhang, J.; Soden, R.; Hayakawa, M.; Kreiman, G.; et al. A gene atlas of the mouse and human protein-encoding transcriptomes. Proc. Natl. Acad. Sci. USA 2004, 101, 6062–6067. [Google Scholar] [CrossRef] [PubMed]
- Fagerberg, L.; Hallström, B.M.; Oksvold, P.; Kampf, C.; Djureinovic, D.; Odeberg, J.; Habuka, M.; Tahmasebpoor, S.; Danielsson, A.; Edlund, K.; et al. Analysis of the human tissue-specific expression by genome-wide integration of transcriptomics and antibody-based proteomics. Mol. Cell. Proteom. 2014, 13, 397–406. [Google Scholar] [CrossRef]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Martignetti, J.A.; Aqeel, A.A.; Sewairi, W.A.; Boumah, C.E.; Kambouris, M.; Mayouf, S.A.; Sheth, K.V.; Eid, W.A.; Dowling, O.; Harris, J.; et al. Mutation of the matrix metalloproteinase 2 gene (MMP2) causes a multicentric osteolysis and arthritis syndrome. Nat. Genet. 2001, 28, 261–265. [Google Scholar] [CrossRef]
- Elsebaie, H.; Mansour, M.A.; Elsayed, S.M.; Mahmoud, S.; El-Sobky, T.A. Multicentric osteolysis, nodulosis, and arthropathy in two unrelated children with matrix metalloproteinase 2 variants: Genetic-skeletal correlations. Bone Rep. 2021, 15, 101106. [Google Scholar] [CrossRef]
- Yadav, S.S.; Mandal, R.K.; Singh, M.K.; Dwivedi, P.; Sethi, R.; Usman, K.; Khattri, S. Cumulative risk of metabolic syndrome correlated with the coexistence of (-1306C/T) and altered circulating MMP2 level. Exp. Clin. Endocrinol. Diabetes 2016, 124, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.S.; Mandal, R.K.; Singh, M.K.; Usman, K.; Khattri, S. Genetic variants of matrix metalloproteinase (MMP2) gene influence metabolic syndrome susceptibility. Genet. Test. Mol. Biomark. 2014, 18, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Belo, V.A.; Luizon, M.R.; Carneiro, P.C.; Gomes, V.A.; Lacchini, R.; Lanna, C.M.M.; Souza-Costa, D.C.; Tanus-Santos, J.E. Effect of metabolic syndrome risk factors and MMP-2 genetic variations on circulating MMP-2 levels in childhood obesity. Mol. Biol. Rep. 2013, 40, 2697–2704. [Google Scholar] [CrossRef] [PubMed]
- Wilson, P.W.F.; D’Agostino, R.B.; Parise, H.; Sullivan, L.; Meigs, J.B. Metabolic syndrome as a precursor of cardiovascular disease and type 2 diabetes mellitus. Circulation 2005, 112, 3066–3072. [Google Scholar] [CrossRef] [PubMed]
- Sharony, R.; Borochowitz, Z.; Cohen, L.; Shtorch-Asor, A.; Rosenfeld, R.; Modai, S.; Reinstein, E. Prenatal course of metaphyseal anadysplasia associated with homozygous mutation in MMP9 identified by exome sequencing. Clin. Genet. 2017, 92, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Lausch, E.; Keppler, R.; Hilbert, K.; Cormier-Daire, V.; Nikkel, S.; Nishimura, G.; Unger, S.; Spranger, J.; Superti-Furga, A.; Zabel, B. Mutations in MMP9 and MMP13 determine the mode of inheritance and the clinical spectrum of metaphyseal anadysplasia. Am. J. Hum. Genet. 2009, 85, 168–178. [Google Scholar] [CrossRef]
- Yadav, S.S.; Mandal, R.K.; Singh, M.K.; Verma, A.; Dwivedi, P.; Sethi, R.; Usman, K.; Khattri, S. High serum level of matrix metalloproteinase 9 and promoter polymorphism—1562 C:T as a new risk factor for metabolic syndrome. DNA Cell Biol. 2014, 33, 816–822. [Google Scholar] [CrossRef] [PubMed]
- Sarray, S.; Dallel, M.; Lamine, L.B.; Jairajpuri, D.; Sellami, N.; Turki, A.; Malalla, Z.; Brock, R.; Ghorbel, M.; Mahjoub, T. Association of matrix metalloproteinase-2 gene polymorphisms with susceptibility to type 2 diabetes: A Case Control Study. J. Diabetes Its Complicat. 2021, 35, 107908. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Liu, C.; Liu, G.; Zuo, C.; Chen, H. Cumulative evidence for association between genetic polymorphisms and esophageal cancer susceptibility: A review with evidence from meta-analysis and genome-wide association studies. Cancer Med. 2019, 8, 1289–1305. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Hao, X.; Zhang, Z. Risk of macular degeneration affected by polymorphisms in matrix metalloproteinase-2: A case-control study in Chinese Han population. Medicine 2017, 96, e8190. [Google Scholar] [CrossRef]
- Tarki, S.E.; Far, I.S.; Aminimoghaddam, S.; Fooladi, B.; Sarhangi, N.; Farahani, M.S.; Klashami, Z.N.; Hamidi, A.K.; Amoli, M.M. Investigating the association of matrix metalloproteinase-2 gene variants with endometriosis in an Iranian population. Eur. J. Obstet. Gynecol. 2021, 258, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Price, S.J.; Greaves, D.R.; Watkins, H. Identification of novel, functional genetic variants in the human matrix metalloproteinase-2 gene: Role of Sp1 in allele-specific transcriptional regulation. J. Biol. Chem. 2001, 276, 7549–7558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Fan, X.-H.; Guan, Y.-Q.; Li, Y.; Sun, W.; Yang, X.-Z.; Liu, R. MMP-2 gene polymorphisms in type 2 diabetes mellitus diabetic retinopathy. Int. J. Ophthalmol. 2010, 3, 137–140. [Google Scholar] [CrossRef]
- Pérez-Hernández, N.; Vargas-Alarcón, G.; Martínez-Rodríguez, N.; Martínez-Ríos, M.A.; Peña-Duque, M.A.; Peña-Díaz, A.; Valente-Acosta, B.; Posadas-Romero, C.; Medina, A.; Rodríguez-Pérez, J.M. The matrix metalloproteinase 2-1575 gene polymorphism is associated with the risk of developing myocardial infarction in Mexican patients. J. Atheroscler. Thromb. 2012, 19, 718–727. [Google Scholar] [CrossRef] [PubMed]
- Harendza, S.; Lovett, D.H.; Panzer, U.; Lukacs, Z.; Kuhnl, P.; Stahl, R.A.K. Linked common polymorphisms in the gelatinase a promoter are associated with diminished transcriptional response to estrogen and genetic fitness. J. Biol. Chem. 2003, 278, 20490–20499. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Sun, Y.; Benveniste, E.N. The transcription factors Sp1, Sp3, and AP-2 are required for constitutive matrix metalloproteinase-2 gene expression in astroglioma cells. J. Biol. Chem. 1999, 274, 29130–29137. [Google Scholar] [CrossRef]
- Sherry, S.T.; Ward, M.H.; Kholodov, M.; Baker, J.; Phan, L.; Smigielski, E.M.; Sirotkin, K. DbSNP: The NCBI database of genetic variation. Nucleic Acids Res. 2001, 29, 308–311. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Agrawal, N.K.; Gupta, S.K.; Singh, K. A functional single nucleotide polymorphism -1562c>t in the matrix metalloproteinase-9 promoter is associated with type 2 diabetes and diabetic foot ulcers. Int. J. Low. Extrem. Wounds 2013, 12, 199–204. [Google Scholar] [CrossRef]
- Saravani, S.; Yari, D.; Saravani, R.; Ahmadabadi, C.A. Association of COL4A3 (Rs55703767), MMP-9 (Rs17576) and TIMP-1 (Rs6609533) gene polymorphisms with susceptibility to type 2 diabetes. Biomed. Rep. 2017, 6, 329–334. [Google Scholar] [CrossRef]
- Yang, H.; Liu, J.; Fan, Y.; Guo, Q.; Ge, L.; Yu, N.; Zheng, X.; Dou, Y.; Zheng, S. Associations between various possible promoter polymorphisms of MMPs genes and endometriosis risk: A meta-analysis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2016, 205, 174–188. [Google Scholar] [CrossRef]
- Xu, T.; Zhang, S.; Qiu, D.; Li, X.; Fan, Y. Association between matrix metalloproteinase 9 polymorphisms and breast cancer risk: An updated meta-analysis and trial sequential analysis. Gene 2020, 759, 144972. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.-F.; Mi, Y.-Y.; Cao, Q.; Wang, W.; Qin, C.; Wei, J.-F.; Zhou, Y.-J.; Li, Y.-F.; Tang, M.; Liu, W.-M.; et al. Update analysis of studies on the MMP-9 -1562 C>T polymorphism and cancer risk. Mol. Biol. Rep. 2012, 39, 3435–3441. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.-Y.; Wu, Y.; Zhang, Y.; Liu, C.-Y.; Deng, C.-Y.; Peng, L.; Zhou, L. Associations between matrix metalloproteinase gene polymorphisms and glaucoma susceptibility: A meta-analysis. BMC Ophthalmol. 2017, 17, 48. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Zhou, H.; Zhu, J. MMP-9-C1562T polymorphism and susceptibility to chronic obstructive pulmonary disease: A meta-analysis. Medicine 2020, 99, e21479. [Google Scholar] [CrossRef]
- Mohammadhosayni, M.; Khosrojerdi, A.; Lorian, K.; Aslani, S.; Imani, D.; Razi, B.; Babaie, F.; Torkamandi, S. Matrix metalloproteinases (MMPs) family gene polymorphisms and the risk of multiple sclerosis: Systematic review and meta-analysis. BMC Neurol. 2020, 20, 218. [Google Scholar] [CrossRef]
- Zou, F.; Zhang, J.; Xiang, G.; Jiao, H.; Gao, H. Association of matrix metalloproteinase 9 (MMP-9) polymorphisms with asthma risk: A meta-analysis. Can. Respir. J. 2019, 2019, 9260495. [Google Scholar] [CrossRef]
- Wu, G.; Cai, H.; Li, G.; Meng, S.; Huang, J.; Xu, H.; Chen, M.; Hu, M.; Yang, W.; Wang, C.; et al. Influence of the matrix metalloproteinase 9 Geners3918242 polymorphism on development of ischemic stroke: A meta-analysis. World Neurosurg. 2020, 133, e31–e61. [Google Scholar] [CrossRef] [PubMed]
- Van den Steen, P.E.; Proost, P.; Grillet, B.; Brand, D.D.; Kang, A.H.; Van Damme, J.; Opdenakker, G. Cleavage of denatured natural collagen type II by neutrophil gelatinase B reveals enzyme specificity, post-translational modifications in the substrate, and the formation of remnant epitopes in rheumatoid arthritis. FASEB J. 2002, 16, 379–389. [Google Scholar] [CrossRef]
- Zhang, B.; Ye, S.; Herrmann, S.M.; Eriksson, P.; de Maat, M.; Evans, A.; Arveiler, D.; Luc, G.; Cambien, F.; Hamsten, A.; et al. Functional polymorphism in the regulatory region of gelatinase B gene in relation to severity of coronary atherosclerosis. Circulation 1999, 99, 1788–1794. [Google Scholar] [CrossRef]
- Ahluwalia, T.S.; Khullar, M.; Ahuja, M.; Kohli, H.S.; Bhansali, A.; Mohan, V.; Venkatesan, R.; Rai, T.S.; Sud, K.; Singal, P.K. Common variants of inflammatory cytokine genes are associated with risk of nephropathy in type 2 diabetes among Asian Indians. PLoS ONE 2009, 4, e5168. [Google Scholar] [CrossRef]
- Maral, S.; Acar, M.; Balcik, O.S.; Uctepe, E.; Hatipoglu, O.F.; Akdeniz, D.; Altun, H.U.; Kosar, A.; Gunduz, M.; Gunduz, E. Matrix metalloproteinases 2 and 9 polymorphism in patients with myeloproliferative diseases: A STROBE-compliant observational study. Medicine 2015, 94, e732. [Google Scholar] [CrossRef] [PubMed]
- Nazir, N.; Siddiqui, K.; Al-Qasim, S.; Al-Naqeb, D. Meta-analysis of diabetic nephropathy associated genetic variants in inflammation and angiogenesis involved in different biochemical pathways. BMC Med. Genet. 2014, 15, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamal, A.; Elgengehy, F.T.; Abd Elaziz, M.M.; Gamal, S.M.; Sobhy, N.; Medhat, A.; El Dakrony, A.H.M. Matrix metalloproteinase-9 Rs17576 gene polymorphism and Behçet’s disease: Is there an association? Immunol. Investig. 2017, 46, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Goyal, P.; Singh, M.; Deshmukh, S.; Upadhyay, D.; Kant, S.; Agrawal, N.K.; Gupta, S.K.; Singh, K. Association of functional SNP-1562C > T in MMP9 promoter with proliferative diabetic retinopathy in North Indian type 2 diabetes mellitus patients. J. Diabetes Its Complicat. 2017, 31, 1648–1651. [Google Scholar] [CrossRef] [PubMed]
- Sarray, S.; Lamine, L.B.; Dallel, M.; Jairajpuri, D.; Turki, A.; Sellami, N.; Ezzidi, I.; Abdelhadi, M.; Brock, R.; Ghorbel, M.; et al. Association of MMP-2 genes variants with diabetic retinopathy in Tunisian population with type 2 diabetes. J. Diabetes Its Complicat. 2022, 36, 108182. [Google Scholar] [CrossRef]
- Beránek, M.; Kolar, P.; Tschoplova, S.; Kankova, K.; Vasku, A. Genetic variations and plasma levels of gelatinase A (matrix metalloproteinase-2) and gelatinase B (matrix metalloproteinase-9) in proliferative diabetic retinopathy. Mol. Vis. 2008, 14, 1114–1121. [Google Scholar]
- Albert, C.; Kube, J.; Albert, A.; Schanze, D.; Zenker, M.; Mertens, P.R. Cubilin single nucleotide polymorphism variants are associated with macroangiopathy while a matrix metalloproteinase-9 single nucleotide polymorphism flip-flop may indicate susceptibility of diabetic nephropathy in type-2 diabetic patients. Nephron 2019, 141, 156–165. [Google Scholar] [CrossRef]
- Feng, S.; Ye, G.; Bai, S.; Wei, H.; Liao, X.; Li, L. Matrix metalloproteinase-9 -1562C/T gene polymorphism is associated with diabetic nephropathy. BioMed Res. Int. 2016, 2016, 1627143. [Google Scholar] [CrossRef]
- Gantala, S.R.; Kondapalli, M.S.; Kummari, R.; Padala, C.; Tupurani, M.A.; Kupsal, K.; Galimudi, R.K.; Gundapaneni, K.K.; Puranam, K.; Shyamala, N.; et al. Collagenase-1 (-1607 1G/2G), gelatinase-A (-1306 C/T), stromelysin-1 (-1171 5A/6A) functional promoter polymorphisms in risk prediction of type 2 diabetic nephropathy. Gene 2018, 673, 22–31. [Google Scholar] [CrossRef]
- Zhang, Z.; Wu, X.; Cai, T.; Gao, W.; Zhou, X.; Zhao, J.; Yao, J.; Shang, H.; Dong, J.; Liao, L. Matrix metalloproteinase 9 gene promoter (rs 3918242) mutation reduces the risk of diabetic microvascular complications. Int. J. Environ. Res. Public Health 2015, 12, 8023–8033. [Google Scholar] [CrossRef]
- Wang, Y.; Su, Y.; Xu, Y.; Pan, S.H.; Liu, G.D. Genetic polymorphism c.1562C>T of the MMP-9 is associated with macroangiopathy in type 2 diabetes mellitus. Biochem. Biophys. Res. Commun. 2010, 391, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Buraczynska, M.; Dragan, M.; Buraczynska, K.; Orlowska-Kowalik, G.; Ksiazek, A. Matrix metalloproteinase-2 (MMP-2) gene polymorphism and cardiovascular comorbidity in type 2 diabetes patients. J. Diabetes Its Complicat. 2015, 29, 829–833. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Frequency of Alternative Allele among Population (=1−the Frequency of the Reference Allele) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gelatinase | SNP | Alternative Allele | Total | All Africans | African American | Individuals with African Ancestry | Asian Individuals Excluding South Asian | East Asian | South Asian | Asian Individiuals Excluding South or East Asian | European | Latin American Individiuals with Afro-Caribbean Ancestry | Latin American Individiuals with Mostly European and Native American Ancestry |
| MMP-2 | |||||||||||||
| −1306 C/T (rs243865) [38] | T | 0.20139 | 0.0597 | 0.0608 | 0.025 | 0.073 | 0.055 | 0.129 | 0.12 | 0.24479 | 0.204 | 0.2152 | |
| −1575 G/A (rs243866) [38] | A | 0.229833 | 0.0562 | 0.0574 | 0.022 | 0.087 | 0.079 | 0.1594 | 0.112 | 0.242161 | 0.203 | 0.2182 | |
| MMP-9 | |||||||||||||
| −1562 C/T (rs3918242) [38] | T | 0.17402 | 0.2275 | 0.2330 | 0.070 | 0.226 | 0.267 | 0.327 | 0.04 | 0.16158 | 0.110 | 0.084 | |
| +279 A/G (rs17576) [38] | G | 0.355297 | 0.34381 | 0.34448 | 0.324 | 0.755 | 0.755 | 0.4824 | 0.755 | 0.355310 | 0.3325 | 0.2256 | |
| Gelatinase | SNP | Control, n (%) | Patients, n (%) | χ2 | p-Value | OR (95%CI) |
|---|---|---|---|---|---|---|
| MMP-2 | ||||||
| S. Sarray et al. (2021) [29] | −1306 C/T (rs243865) | Σn = 310 | Σn = 791 | |||
| genotype | ||||||
| CC | (64.1) | (73) | 0.006 | 1.00 | ||
| CT | (27.3) | (23.3) | 0.75 (0.54–1.03) | |||
| TT | (8.6) | (3.7) | 0.38 (0.21–0.7) | |||
| CT + TT | (35.9) | (27) | 0.66 (0.49–0.89) | |||
| CC + CT | (91.5) | (96.3) | 0.003 | 1.00 | ||
| allele frequency | ||||||
| C | (77.75) * | (84.65) * | ||||
| T | (22.25) * | (15.35) * | ||||
| S. Sarray et al. (2021) [29] | −1575 G/A (rs243866) | Σn = 310 | Σn = 791 | |||
| genotype | ||||||
| GG | (66.1) | (74.1) | 0.012 | 1.00 | ||
| GA | (30.4) | (23.7) | 0.70 (0.51–0.95) | |||
| AA | (3.5) | (2.2) | 0.56 (0.25–1.25) | |||
| GA + AA | (33.9) | (25.9) | 0.68 (0.51–0.92) | |||
| GG + GA | (96.5) | (97.8) | 0.25 | 1.00 | ||
| allele frequency | ||||||
| G | (81.3) * | (85.95) * | ||||
| A | (18.7) * | (14.05) * | ||||
| MMP-9 | ||||||
| K. Singh et al. (2013) [39] | −1562 C/T (rs3918242) | Σn = 267 | Σn = 353 | |||
| genotype | ||||||
| CC | 196 (73.40) | 204 (57.8) | Ref. | |||
| CT | 69 (25.84) | 137 (57.8) | 13.28 * | 0.00027 * | 1.91 (1.34–2.71) * | |
| TT | 2 (0.75) | 12 (3.4) | 6.53 * | 0.01061 * | 5.76 (1.27–26.09) * | |
| CT + TT | 71 (26.6) | 149 (42.2) | 16.2 * | 0.00006 * | 2.02 (1.43–2.84) * | |
| allele frequency | ||||||
| C | 461 (86.33) | 545 (77.2) | Ref. | |||
| T | 73 (13.67) | 161 (22.8) | 16.57 * | 0.00006 * | 1.87 (1.38–2.53) * | |
| S. Saravani et al. (2017) [40] | +279 A/G (rs17576) | Σn = 120 | Σn = 120 | |||
| genotype | ||||||
| GG | 102 (85) | 84 (70) | Ref. | |||
| AG | 18 (15) | 36 (30) | 7.74 * | 0.00540* | 2.43 (1.29–4.58) * | |
| AA | 0 | 0 | 1.00000 | |||
| allele frequency | ||||||
| G | 222 (92.5) | 204 (85) | Ref. | |||
| A | 18 (7.5) | 36(15) | 6.76 * | 0.00932 * | 2.18 (1.2–3.95) * |
| Control Subjects | T2D Patients | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Study | Ethnicity | Gender (Male:Female) | Age (Years), Mean ± SD | BMI (kg/m2), Mean ± SD | Gender (Male:Female) | Age (Years), Mean ± SD | BMI (kg/m2), Mean ± SD | Diabetes Duration (Years), Mean ± SD | |
| MMP-2 | S. Sarray et al. (2021) [29] | Tunisian Arab | 141:169 | 60 ± 12.6 | 24.8 ± 2.8 | 440:351 | 60.6 ± 7.5 | 28.5 ± 5 | 13.6 ± 7.8 |
| MMP-9 | K. Singh et al. (2013) [39] | Homogeneous ethnic group of north Indian population | N/A | N/A | N/A | 215:138 | 51.41 ± 10.56 | 23.89 ± 4.52 | 6.14 ± 5.42 |
| S. Saravani et al. (2017) [40] | Iranian | 32:88 | 56.11 ± 11.075 | N/A | 29:91 | 56.57 ± 10.602 | N/A | 9.91 ± 6.90 | |
| Disease | Gelatinase | SNP | Association | |
|---|---|---|---|---|
| Retinopathy | ||||
| MMP-2 | −790 T/G (rs243864) | increased risk of the disease | S. Sarray et al. (2022) [56] | |
| −1575 G/A (rs243866) | increased risk of the disease | S. Sarray et al. (2022) [56] | ||
| −1306 C/T (rs243865) | doubled risk of the disease | J. Yang et al. (2010) [34] | ||
| marginally significant increased risk of disease in males | M. Beránek et al. (2008) [57] | |||
| MMP-9 | −1562 C/T (rs3918242) | possible risk factor for the disease | K. Singh et al. (2017) [55] | |
| Nephropathy | ||||
| MMP-2 | −1306 C/T (rs243865) | the presence of the C allele was associated with disease susceptibility and progression | S.R. Gantala et al. (2018) [60] | |
| MMP-9 | +279 A/G (rs17576) | the presence of the GG genotype was independently associated with disease | C. Albert et al. (2019) [58] | |
| −1562 C/T (rs3918242) | the T allele was a protective factor, while the C allele contributed to the disease | S. Feng et al. (2016) [59] | ||
| the T allele reduces the risk of disease | Z. Zhang et al. (2015) [61] | |||
| Diabetic foot ulcers | ||||
| MMP-9 | −1562 C/T (rs3918242) | presence of allele T leads to a higher risk of developing the disease | K. Singh et al. (2013) [39] | |
| Macroangiopathy | ||||
| MMP-9 | −1562 C/T (rs3918242) | the presence of the T allele was higher in patients with disease | Y. Wang et al. (2010) [62] | |
| Cardiovascular comorbidity | ||||
| MMP-2 | −1306 C/T (rs243865) | possession of the T allele was associated with a reduced risk of disease | M. Buraczynska et al. (2015) [63] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gajewska, B.; Śliwińska-Mossoń, M. Association of MMP-2 and MMP-9 Polymorphisms with Diabetes and Pathogenesis of Diabetic Complications. Int. J. Mol. Sci. 2022, 23, 10571. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms231810571
Gajewska B, Śliwińska-Mossoń M. Association of MMP-2 and MMP-9 Polymorphisms with Diabetes and Pathogenesis of Diabetic Complications. International Journal of Molecular Sciences. 2022; 23(18):10571. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms231810571
Chicago/Turabian StyleGajewska, Beata, and Mariola Śliwińska-Mossoń. 2022. "Association of MMP-2 and MMP-9 Polymorphisms with Diabetes and Pathogenesis of Diabetic Complications" International Journal of Molecular Sciences 23, no. 18: 10571. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms231810571