
The Plasma Oxylipin Signature Provides a Deep Phenotyping of Metabolic Syndrome Complementary to the Clinical Criteria
 , , , , , , , , , and
, , , , , , , , , and
Abstract
:1. Introduction
2. Results
2.1. Baseline Characteristics of Participants
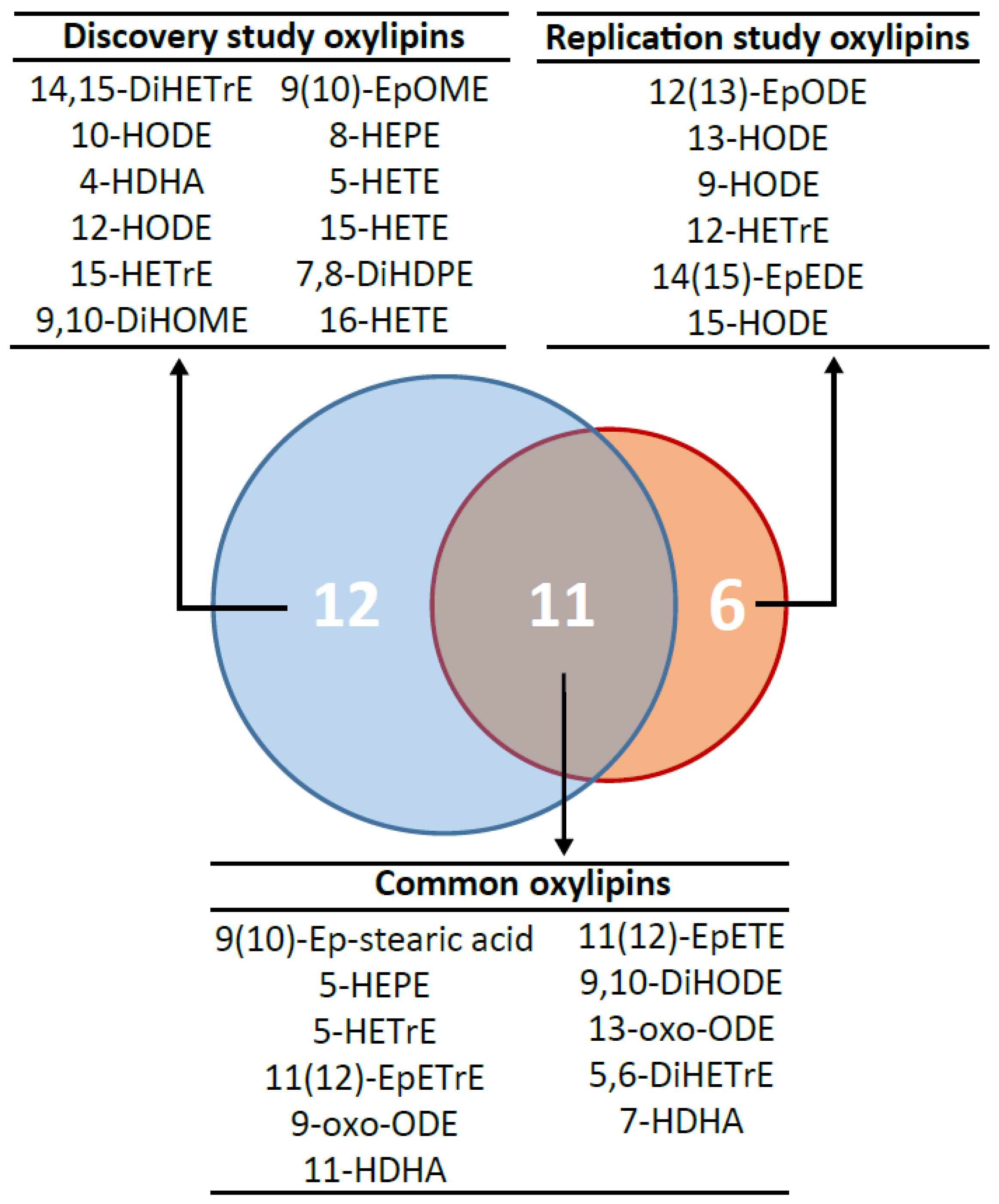
2.2. Lipidomic Profiling Identifies Consistent Oxylipin Signatures of Metabolic Syndrome between the Discovery and the Replication Studies
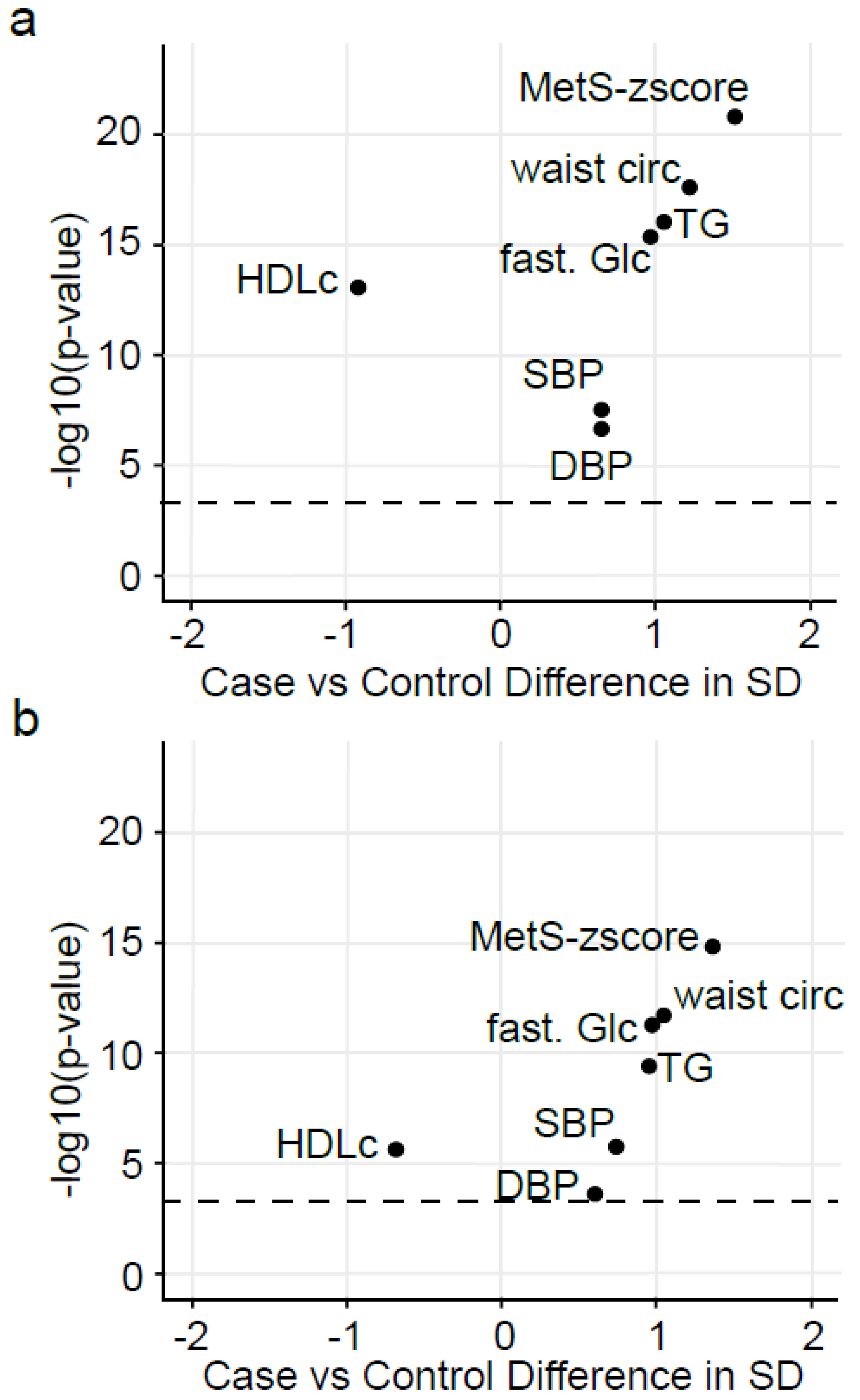
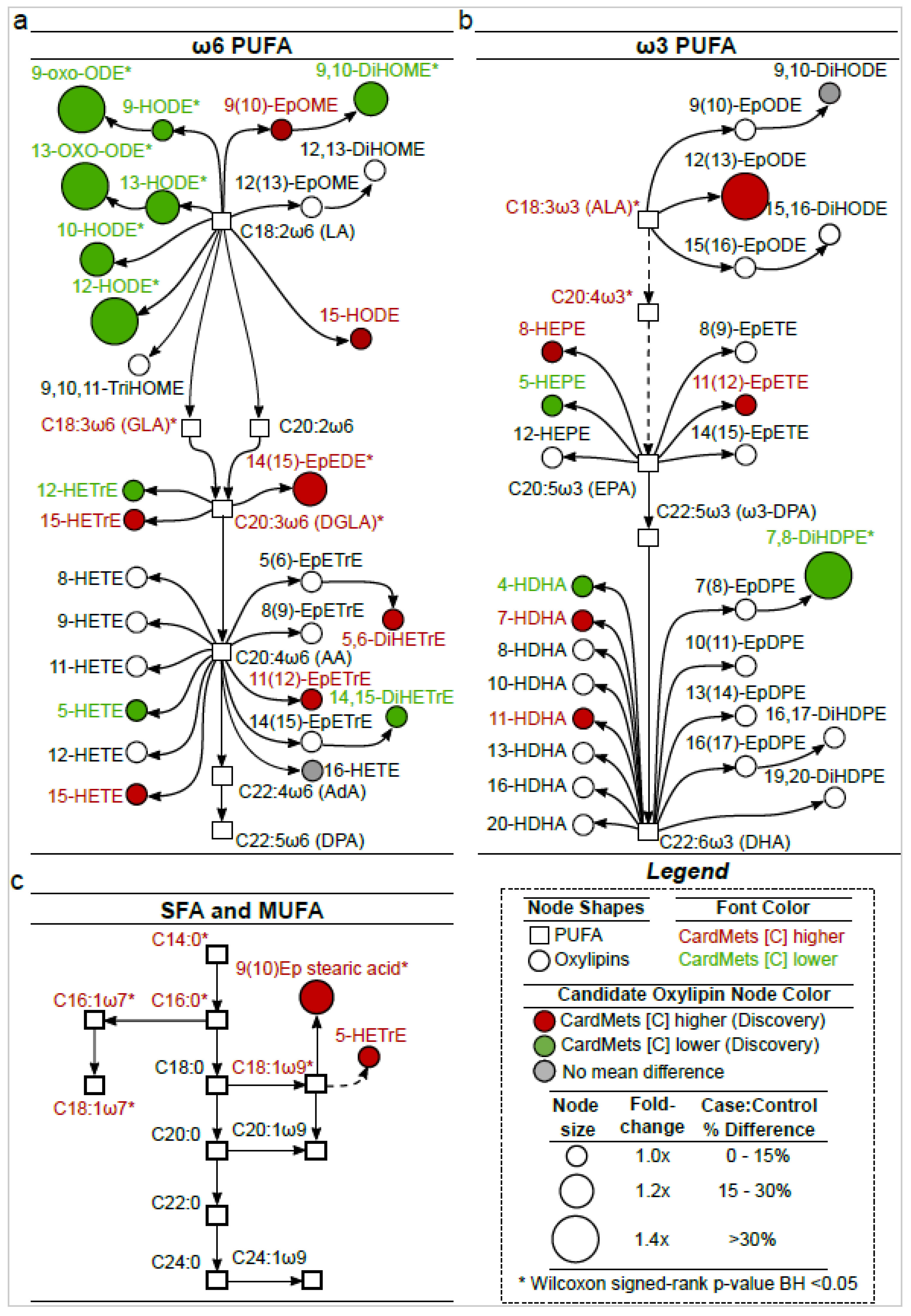
2.3. The Identified Candidate Oxylipins Provide a Subtle and Integrative Phenotype of MetS
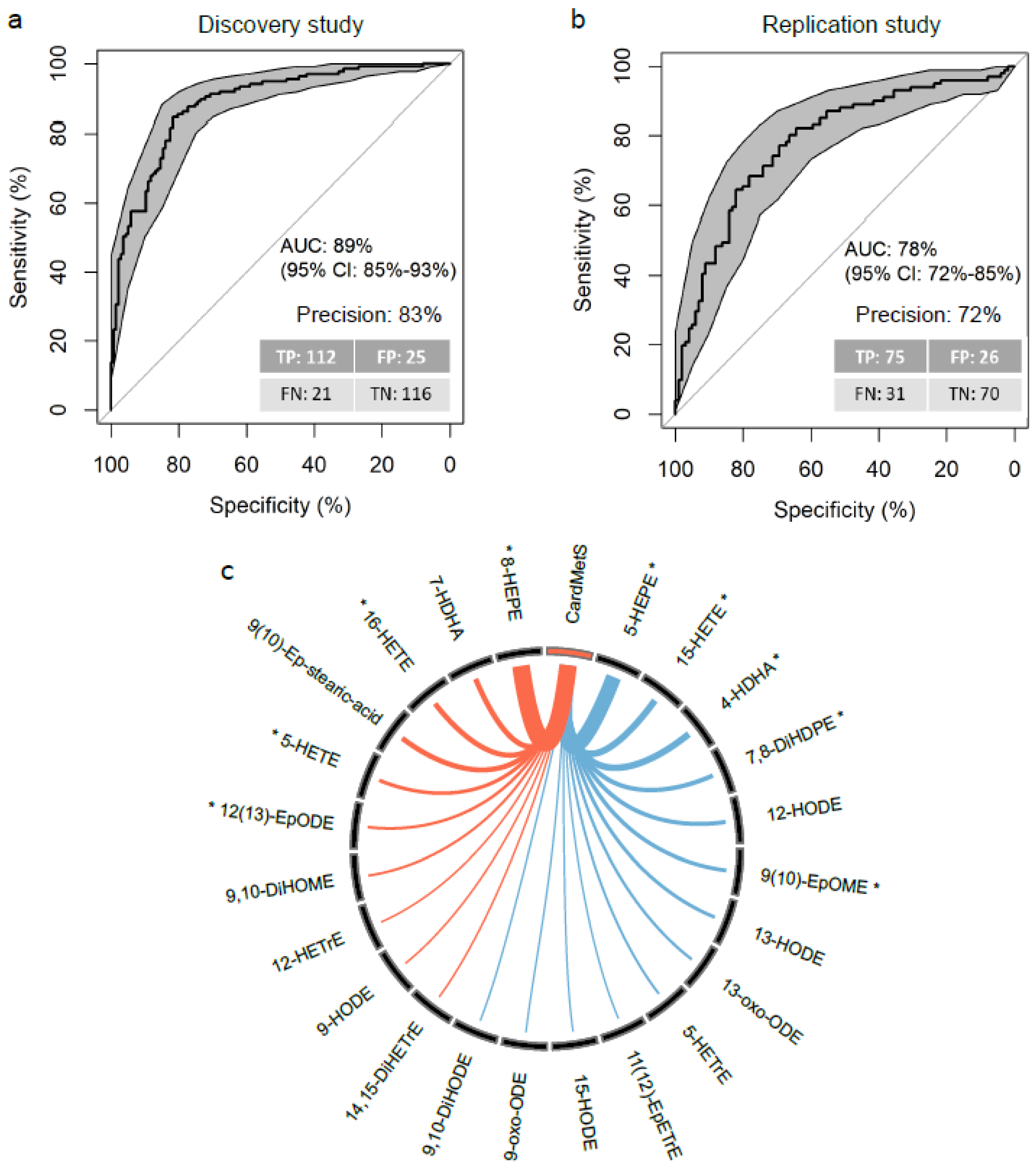
2.4. The OxyScore Includes 23 Candidate Oxylipins and Has High Performances of Classification and Replicability
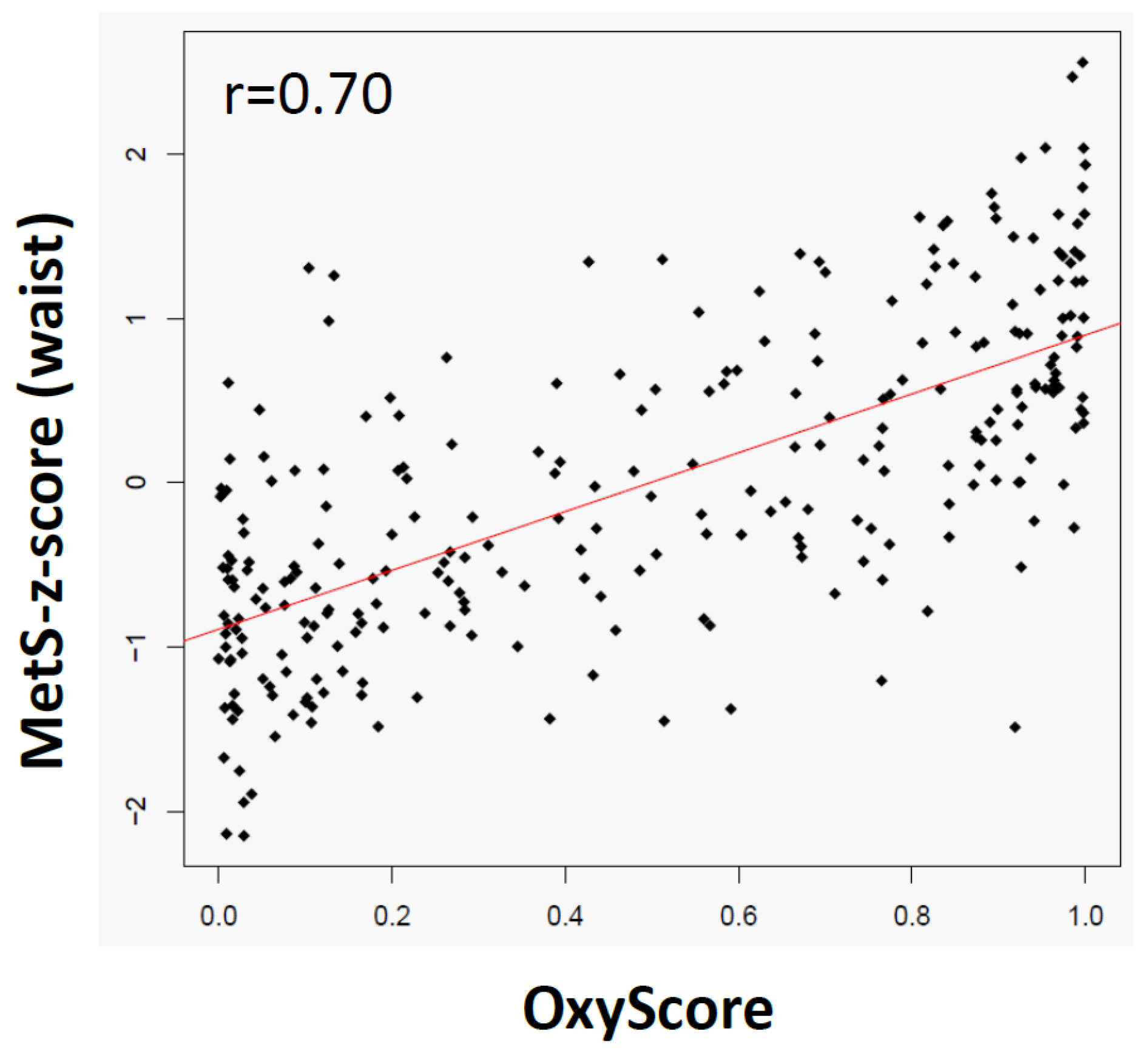
2.5. The OxyScore Is Consistent and Complementary with the MetS-z-Score and the Clinical Criteria of MetS
3. Discussion
4. Materials and Methods
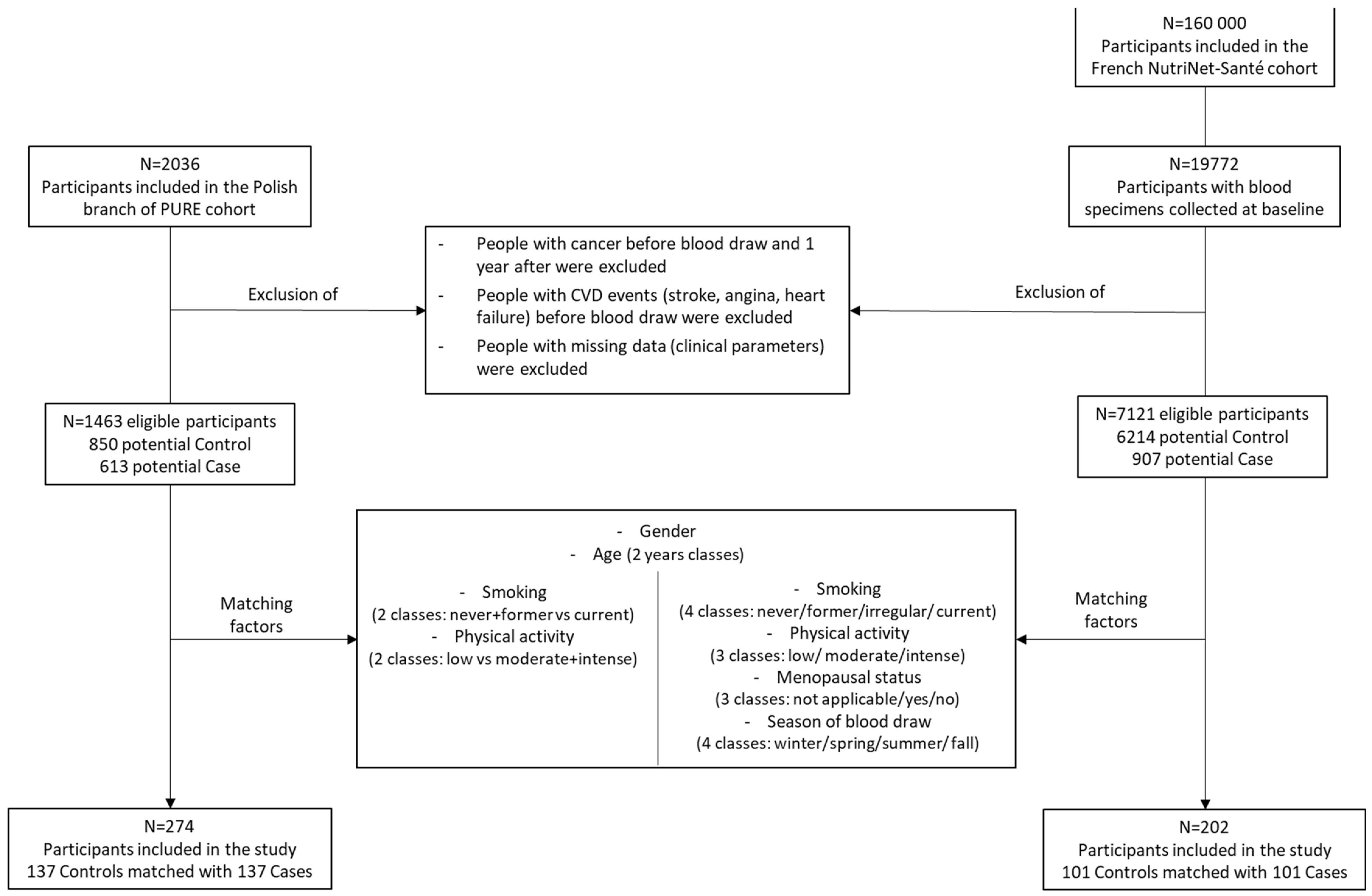
4.1. Discovery and Replication Cohorts
4.2. Study Design and Selection of Participants
4.3. Oxylipin and Fatty Acid Quantification
4.3.1. Extraction and MS Quantitative Profiling of Oxylipins
4.3.2. Fatty Acid Profiling
4.4. Statistical Analysis
4.4.1. Metadata Statistical Analysis
4.4.2. Pre-processing of MS Oxylipin Data
4.4.3. Candidate Oxylipins Selection
4.4.4. Model Construction and Validation
4.4.5. OxyScore Calculation
4.4.6. OxyScore Correlation
4.4.7. OxyScore Adjustment
4.4.8. Univariate Analysis
4.4.9. Study Approval
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saklayen, M.G. The Global Epidemic of the Metabolic Syndrome. Curr. Hypertens Rep. 2018, 20, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grundy, S.M. Metabolic syndrome update. Trends Cardiovasc. Med. 2016, 26, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Alberti, K.G.; Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z.; Cleeman, J.I.; Donato, K.A.; Fruchart, J.C.; James, W.P.; Loria, C.M.; Smith, S.C., Jr. Harmonizing the metabolic syndrome: A joint interim statement of the International Diabetes Federation Task Force on Epidemiology and Prevention; National Heart, Lung, and Blood Institute; American Heart Association; World Heart Federation; International Atherosclerosis Society; and International Association for the Study of Obesity. Circulation 2009, 120, 1640–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robberecht, H.; Hermans, N. Biomarkers of Metabolic Syndrome: Biochemical Background and Clinical Significance. Metab. Syndr. Relat. Disord. 2016, 14, 47–93. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, S.; Bohl, M.; Gregersen, S.; Hermansen, K.; O’Driscoll, L. Blood-Based Biomarkers for Metabolic Syndrome. Trends Endocrinol. Metab. 2016, 27, 363–374. [Google Scholar] [CrossRef]
- Monnerie, S.; Comte, B.; Ziegler, D.; Morais, J.A.; Pujos-Guillot, E.; Gaudreau, P. Metabolomic and Lipidomic Signatures of Metabolic Syndrome and its Physiological Components in Adults: A Systematic Review. Sci. Rep. 2020, 10, 669. [Google Scholar] [CrossRef] [Green Version]
- Comte, B.; Monnerie, S.; Brandolini-Bunlon, M.; Canlet, C.; Castelli, F.; Chu-Van, E.; Colsch, B.; Fenaille, F.; Joly, C.; Jourdan, F.; et al. Multiplatform metabolomics for an integrative exploration of metabolic syndrome in older men. EBioMedicine 2021, 69, 103440. [Google Scholar] [CrossRef]
- Yin, X.; Willinger, C.M.; Keefe, J.; Liu, J.; Fernández-Ortiz, A.; Ibáñez, B.; Peñalvo, J.; Adourian, A.; Chen, G.; Corella, D.; et al. Lipidomic profiling identifies signatures of metabolic risk. EBioMedicine 2020, 51, 102520. [Google Scholar] [CrossRef] [Green Version]
- Gallart-Ayala, H.; Teav, T.; Ivanisevic, J. Metabolomics meets lipidomics: Assessing the small molecule component of metabolism. BioEssays 2020, 42, 2000052. [Google Scholar] [CrossRef]
- Dennis, E.A.; Norris, P.C. Eicosanoid storm in infection and inflammation. Nat. Rev. Immunol. 2015, 15, 511–523. [Google Scholar] [CrossRef]
- Fernández-García, J.C.; Cardona, F.; Tinahones, F.J. Inflammation, oxidative stress and metabolic syndrome: Dietary modulation. Curr. Vasc. Pharm. 2013, 11, 906–919. [Google Scholar] [CrossRef] [PubMed]
- Tourdot, B.E.; Ahmed, I.; Holinstat, M. The emerging role of oxylipins in thrombosis and diabetes. Front. Pharm. 2014, 4, 176. [Google Scholar] [CrossRef] [PubMed]
- Shearer, G.C.; Newman, J.W. Impact of circulating esterified eicosanoids and other oxylipins on endothelial function. Curr. Atheroscler Rep. 2009, 11, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Mainka, M.; Dalle, C.; Pétéra, M.; Dalloux-Chioccioli, J.; Kampschulte, N.; Ostermann, A.I.; Rothe, M.; Bertrand-Michel, J.; Newman, J.W.; Gladine, C.; et al. Harmonized procedures lead to comparable quantification of total oxylipins across laboratories. J. Lipid Res. 2020, 61, 1424–1436. [Google Scholar] [CrossRef] [PubMed]
- Rund, K.M.; Ostermann, A.I.; Kutzner, L.; Galano, J.M.; Oger, C.; Vigor, C.; Wecklein, S.; Seiwert, N.; Durand, T.; Schebb, N.H. Development of an LC-ESI(-)-MS/MS method for the simultaneous quantification of 35 isoprostanes and isofurans derived from the major n3- and n6-PUFAs. Anal Chim. Acta 2018, 1037, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Gurka, M.J.; Lilly, C.L.; Oliver, M.N.; DeBoer, M.D. An examination of sex and racial/ethnic differences in the metabolic syndrome among adults: A confirmatory factor analysis and a resulting continuous severity score. Metabolism 2014, 63, 218–225. [Google Scholar] [CrossRef] [Green Version]
- Vangaveti, V.N.; Jansen, H.; Kennedy, R.L.; Malabu, U.H. Hydroxyoctadecadienoic acids: Oxidised derivatives of linoleic acid and their role in inflammation associated with metabolic syndrome and cancer. Eur. J. Pharm. 2016, 785, 70–76. [Google Scholar] [CrossRef]
- Yeung, J.; Holinstat, M. Who is the real 12-HETrE? Prostaglandins Other Lipid Mediat. 2017, 132, 25–30. [Google Scholar] [CrossRef]
- Ostermann, A.I.; Koch, E.; Rund, K.M.; Kutzner, L.; Mainka, M.; Schebb, N.H. Targeting esterified oxylipins by LC–MS - Effect of sample preparation on oxylipin pattern. Prostaglandins Other Lipid Mediat. 2020, 146, 106384. [Google Scholar] [CrossRef]
- Umeno, A.; Shichiri, M.; Ishida, N.; Hashimoto, Y.; Abe, K.; Kataoka, M.; Yoshino, K.; Hagihara, Y.; Aki, N.; Funaki, M.; et al. Singlet oxygen induced products of linoleates, 10- and 12-(Z,E)-hydroxyoctadecadienoic acids (HODE), can be potential biomarkers for early detection of type 2 diabetes. PLoS ONE 2013, 8, e63542. [Google Scholar] [CrossRef]
- Schebb, N.H.; Ostermann, A.I.; Yang, J.; Hammock, B.D.; Hahn, A.; Schuchardt, J.P. Comparison of the effects of long-chain omega-3 fatty acid supplementation on plasma levels of free and esterified oxylipins. Prostaglandins Other Lipid Mediat. 2014, 113–115, 21–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shearer, G.C.; Newman, J.W. Lipoprotein lipase releases esterified oxylipins from very low-density lipoproteins. Prostaglandins Leukot. Essent. Fat. Acids 2008, 79, 215–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ek-Von Mentzer, B.A.; Zhang, F.; Hamilton, J.A. Binding of 13-HODE and 15-HETE to phospholipid bilayers, albumin, and intracellular fatty acid binding proteins. Implications for transmembrane and intracellular transport and for protection from lipid peroxidation. J. Biol. Chem. 2001, 276, 15575–15580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maclouf, J.; Kindahl, H.; Granström, E.; Samuelsson, B. Interactions of prostaglandin H2 and thromboxane A2 with human serum albumin. Eur. J. Biochem. 1980, 109, 561–566. [Google Scholar] [CrossRef]
- Boilard, E. Extracellular vesicles and their content in bioactive lipid mediators: More than a sack of microRNA. J. Lipid Res. 2018, 59, 2037–2046. [Google Scholar] [CrossRef] [Green Version]
- Pizzinat, N.; Ong-Meang, V.; Bourgailh-Tortosa, F.; Blanzat, M.; Perquis, L.; Cussac, D.; Parini, A.; Poinsot, V. Extracellular vesicles of MSCs and cardiomyoblasts are vehicles for lipid mediators. Biochimie 2020, 178, 69–80. [Google Scholar] [CrossRef]
- Vangaveti, V.; Baune, B.T.; Kennedy, R.L. Hydroxyoctadecadienoic acids: Novel regulators of macrophage differentiation and atherogenesis. Adv. Endocrinol. Metab. 2010, 1, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Nagy, L.; Tontonoz, P.; Alvarez, J.G.; Chen, H.; Evans, R.M. Oxidized LDL regulates macrophage gene expression through ligand activation of PPARgamma. Cell 1998, 93, 229–240. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Luo, N.; Lopes-Virella, M.F.; Garvey, W.T. The adipocyte lipid binding protein (ALBP/aP2) gene facilitates foam cell formation in human THP-1 macrophages. Atheroscler. 2002, 165, 259–269. [Google Scholar] [CrossRef]
- Vangaveti, V.N.; Shashidhar, V.M.; Rush, C.; Malabu, U.H.; Rasalam, R.R.; Collier, F.; Baune, B.T.; Kennedy, R.L. Hydroxyoctadecadienoic acids regulate apoptosis in human THP-1 cells in a PPARγ-dependent manner. Lipids 2014, 49, 1181–1192. [Google Scholar] [CrossRef]
- Iversen, L.; Fogh, K.; Bojesen, G.; Kragballe, K. Linoleic acid and dihomogammalinolenic acid inhibit leukotriene B4 formation and stimulate the formation of their 15-lipoxygenase products by human neutrophils in vitro. Evidence of formation of antiinflammatory compounds. Agents Actions 1991, 33, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Camp, R.D.; Fincham, N.J. Inhibition of ionophore-stimulated leukotriene B4 production in human leucocytes by monohydroxy fatty acids. Br. J. Pharm. 1985, 85, 837–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warner, D.R.; Liu, H.; Miller, M.E.; Ramsden, C.E.; Gao, B.; Feldstein, A.E.; Schuster, S.; McClain, C.J.; Kirpich, I.A. Dietary Linoleic Acid and Its Oxidized Metabolites Exacerbate Liver Injury Caused by Ethanol via Induction of Hepatic Proinflammatory Response in Mice. Am. J. Pathol. 2017, 187, 2232–2245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delerive, P.; Furman, C.; Teissier, E.; Fruchart, J.; Duriez, P.; Staels, B. Oxidized phospholipids activate PPARalpha in a phospholipase A2-dependent manner. FEBS Lett. 2000, 471, 34–38. [Google Scholar] [CrossRef] [Green Version]
- Buchanan, M.R.; Bertomeu, M.C.; Haas, T.A.; Orr, F.W.; Eltringham-Smith, L.L. Localization of 13-hydroxyoctadecadienoic acid and the vitronectin receptor in human endothelial cells and endothelial cell/platelet interactions in vitro. Blood 1993, 81, 3303–3312. [Google Scholar] [CrossRef] [Green Version]
- Haas, T.A.; Bastida, E.; Nakamura, K.; Hullin, F.; Admirall, L.; Buchanan, M.R. Binding of 13-HODE and 5-, 12- and 15-HETE to endothelial cells and subsequent platelet, neutrophil and tumor cell adhesion. Biochim. Biophys. Acta 1988, 961, 153–159. [Google Scholar] [CrossRef]
- Haas, T.A.; Bertomeu, M.C.; Bastida, E.; Buchanan, M.R. Cyclic AMP regulation of endothelial cell triacylglycerol turnover, 13-hydroxyoctadecadienoic acid (13-HODE) synthesis and endothelial cell thrombogenicity. Biochim. Biophys. Acta 1990, 1051, 174–178. [Google Scholar] [CrossRef]
- Marx, N.; Bourcier, T.; Sukhova, G.K.; Libby, P.; Plutzky, J. PPARgamma activation in human endothelial cells increases plasminogen activator inhibitor type-1 expression: PPARgamma as a potential mediator in vascular disease. Arter. Thromb. Vasc. Biol. 1999, 19, 546–551. [Google Scholar] [CrossRef] [Green Version]
- Coene, M.C.; Bult, H.; Claeys, M.; Laekeman, G.M.; Herman, A.G. Inhibition of rabbit platelet activation by lipoxygenase products of arachidonic and linoleic acid. Thromb. Res. 1986, 42, 205–214. [Google Scholar] [CrossRef]
- Setty, B.N.; Berger, M.; Stuart, M.J. 13-Hydroxyoctadecadienoic acid (13-HODE) stimulates prostacyclin production by endothelial cells. Biochem. Biophys. Res. Commun. 1987, 146, 502–509. [Google Scholar] [CrossRef]
- Shearer, G.C.; Borkowski, K.; Puumala, S.L.; Harris, W.S.; Pedersen, T.L.; Newman, J.W. Abnormal lipoprotein oxylipins in metabolic syndrome and partial correction by omega-3 fatty acids. Prostaglandins Leukot. Essent. Fat. Acids 2018, 128, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Colas, R.; Pruneta-Deloche, V.; Guichardant, M.; Luquain-Costaz, C.; Cugnet-Anceau, C.; Moret, M.; Vidal, H.; Moulin, P.; Lagarde, M.; Calzada, C. Increased lipid peroxidation in LDL from type-2 diabetic patients. Lipids 2010, 45, 723–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, E.; Mainka, M.; Dalle, C.; Ostermann, A.I.; Rund, K.M.; Kutzner, L.; Froehlich, L.-F.; Bertrand-Michel, J.; Gladine, C.; Schebb, N.H. Stability of oxylipins during plasma generation and long-term storage. Talanta 2020, 217, 121074. [Google Scholar] [CrossRef]
- Dos Santos, L.R.B.; Fleming, I. Role of cytochrome P450-derived, polyunsaturated fatty acid mediators in diabetes and the metabolic syndrome. Prostaglandins Other. Lipid Mediat. 2020, 148, 106407. [Google Scholar] [CrossRef] [PubMed]
- Anzenbacher, P.; Hudecek, J. Differences in flexibility of active sites of cytochromes P450 probed by resonance Raman and UV-Vis absorption spectroscopy. J. Inorg. Biochem. 2001, 87, 209–213. [Google Scholar] [CrossRef]
- Zhu, Y.; Schieber, E.B.; McGiff, J.C.; Balazy, M. Identification of arachidonate P-450 metabolites in human platelet phospholipids. Hypertension 1995, 25, 854–859. [Google Scholar] [CrossRef]
- Falck, J.R.; Manna, S.; Moltz, J.; Chacos, N.; Capdevila, J. Epoxyeicosatrienoic acids stimulate glucagon and insulin release from isolated rat pancreatic islets. Biochem. Biophys. Res. Commun. 1983, 114, 743–749. [Google Scholar] [CrossRef]
- Huang, H.; Weng, J.; Wang, M.H. EETs/sEH in diabetes and obesity-induced cardiovascular diseases. Prostaglandins Other Lipid Mediat. 2016, 125, 80–89. [Google Scholar] [CrossRef]
- Sonnweber, T.; Pizzini, A.; Nairz, M.; Weiss, G.; Tancevski, I. Arachidonic Acid Metabolites in Cardiovascular and Metabolic Diseases. Int. J. Mol. Sci. 2018, 19, 3285. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Fu, J.L.; Miao, Y.F.; Wang, C.J.; Han, Q.F.; Li, S.; Huang, S.Z.; Du, S.N.; Qiu, Y.X.; Yang, J.C.; et al. Prostaglandin E2 receptor EP3 regulates both adipogenesis and lipolysis in mouse white adipose tissue. J. Mol. Cell Biol. 2016, 8, 518–529. [Google Scholar] [CrossRef]
- Ma, B.; Xiong, X.; Chen, C.; Li, H.; Xu, X.; Li, X.; Li, R.; Chen, G.; Dackor, R.T.; Zeldin, D.C.; et al. Cardiac-specific overexpression of CYP2J2 attenuates diabetic cardiomyopathy in male streptozotocin-induced diabetic mice. Endocrinology 2013, 154, 2843–2856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Wang, D.W. CYP epoxygenase derived EETs: From cardiovascular protection to human cancer therapy. Curr. Top. Med. Chem. 2013, 13, 1454–1469. [Google Scholar] [CrossRef] [PubMed]
- Luo, P.; Chang, H.H.; Zhou, Y.; Zhang, S.; Hwang, S.H.; Morisseau, C.; Wang, C.Y.; Inscho, E.W.; Hammock, B.D.; Wang, M.H. Inhibition or deletion of soluble epoxide hydrolase prevents hyperglycemia, promotes insulin secretion, and reduces islet apoptosis. J. Pharm. Exp. 2010, 334, 430–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aliwarga, T.; Evangelista, E.A.; Sotoodehnia, N.; Lemaitre, R.N.; Totah, R.A. Regulation of CYP2J2 and EET Levels in Cardiac Disease and Diabetes. Int. J. Mol. Sci. 2018, 19, 1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellien, J.; Iacob, M.; Remy-Jouet, I.; Lucas, D.; Monteil, C.; Gutierrez, L.; Vendeville, C.; Dreano, Y.; Mercier, A.; Thuillez, C.; et al. Epoxyeicosatrienoic acids contribute with altered nitric oxide and endothelin-1 pathways to conduit artery endothelial dysfunction in essential hypertension. Circulation 2012, 125, 1266–1275. [Google Scholar] [CrossRef]
- Imig, J.D. Epoxyeicosatrienoic Acids and 20-Hydroxyeicosatetraenoic Acid on Endothelial and Vascular Function. Adv. Pharm. 2016, 77, 105–141. [Google Scholar] [CrossRef] [Green Version]
- Imig, J.D. Epigenetic soluble epoxide hydrolase regulation causes endothelial dysfunction. Acta. Physiol. 2019, 225, e13203. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Cheriyan, J.; Gutterman, D.D.; Mayer, R.J.; Ament, Z.; Griffin, J.L.; Lazaar, A.L.; Newby, D.E.; Tal-Singer, R.; Wilkinson, I.B. Mechanisms of Vascular Dysfunction in COPD and Effects of a Novel Soluble Epoxide Hydrolase Inhibitor in Smokers. Chest 2017, 151, 555–563. [Google Scholar] [CrossRef] [Green Version]
- Hamzaoui, M.; Guerrot, D.; Djerada, Z.; Duflot, T.; Richard, V.; Bellien, J. Cardiovascular consequences of chronic kidney disease, impact of modulation of epoxyeicosatrienoic acids. Ann. Cardiol. Angeiol. 2018, 67, 141–148. [Google Scholar] [CrossRef]
- Hercule, H.C.; Schunck, W.H.; Gross, V.; Seringer, J.; Leung, F.P.; Weldon, S.M.; da Costa Goncalves, A.; Huang, Y.; Luft, F.C.; Gollasch, M. Interaction between P450 eicosanoids and nitric oxide in the control of arterial tone in mice. Arter. Thromb. Vasc. Biol. 2009, 29, 54–60. [Google Scholar] [CrossRef]
- Oltman, C.L.; Weintraub, N.L.; VanRollins, M.; Dellsperger, K.C. Epoxyeicosatrienoic acids and dihydroxyeicosatrienoic acids are potent vasodilators in the canine coronary microcirculation. Circ. Res. 1998, 83, 932–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chabowski, D.S.; Cohen, K.E.; Abu-Hatoum, O.; Gutterman, D.D.; Freed, J.K. Crossing signals: Bioactive lipids in the microvasculature. Am. J. Physiol. Heart. Circ. Physiol. 2020, 318, H1185–H1197. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.H.; Falck, J.R.; Manthati, V.L.; Campbell, W.B.; Imig, J.D. Epoxyeicosatrienoic acid analog attenuates angiotensin II hypertension and kidney injury. Front. Pharm. 2014, 5, 216. [Google Scholar] [CrossRef] [Green Version]
- Roman, R.J. P-450 metabolites of arachidonic acid in the control of cardiovascular function. Physiol. Rev. 2002, 82, 131–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romashko, M.; Schragenheim, J.; Abraham, N.G.; McClung, J.A. Epoxyeicosatrienoic Acid as Therapy for Diabetic and Ischemic Cardiomyopathy. Trends Pharm. Sci. 2016, 37, 945–962. [Google Scholar] [CrossRef] [PubMed]
- Proctor, K.G.; Falck, J.R.; Capdevila, J. Intestinal vasodilation by epoxyeicosatrienoic acids: Arachidonic acid metabolites produced by a cytochrome P450 monooxygenase. Circ. Res. 1987, 60, 50–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krötz, F.; Riexinger, T.; Buerkle, M.A.; Nithipatikom, K.; Gloe, T.; Sohn, H.Y.; Campbell, W.B.; Pohl, U. Membrane-potential-dependent inhibition of platelet adhesion to endothelial cells by epoxyeicosatrienoic acids. Arter. Thromb. Vasc. Biol. 2004, 24, 595–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitzpatrick, F.A.; Ennis, M.D.; Baze, M.E.; Wynalda, M.A.; McGee, J.E.; Liggett, W.F. Inhibition of cyclooxygenase activity and platelet aggregation by epoxyeicosatrienoic acids. Influence of stereochemistry. J. Biol. Chem. 1986, 261, 15334–15338. [Google Scholar]
- VanRollins, M. Epoxygenase metabolites of docosahexaenoic and eicosapentaenoic acids inhibit platelet aggregation at concentrations below those affecting thromboxane synthesis. J. Pharm. Exp. 1995, 274, 798–804. [Google Scholar]
- Li, R.; Xu, X.; Chen, C.; Wang, Y.; Gruzdev, A.; Zeldin, D.C.; Wang, D.W. CYP2J2 attenuates metabolic dysfunction in diabetic mice by reducing hepatic inflammation via the PPARγ. Am. J. Physiol. Endocrinol. Metab. 2015, 308, E270–E282. [Google Scholar] [CrossRef] [Green Version]
- Node, K.; Huo, Y.; Ruan, X.; Yang, B.; Spiecker, M.; Ley, K.; Zeldin, D.C.; Liao, J.K. Anti-inflammatory properties of cytochrome P450 epoxygenase-derived eicosanoids. Sci. 1999, 285, 1276–1279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Oltman, C.L.; Lu, T.; Lee, H.C.; Dellsperger, K.C.; VanRollins, M. EET homologs potently dilate coronary microvessels and activate BK(Ca) channels. Am. J. Physiol. Heart. Circ. Physiol. 2001, 280, H2430–H2440. [Google Scholar] [CrossRef] [PubMed]
- Bettaieb, A.; Nagata, N.; AbouBechara, D.; Chahed, S.; Morisseau, C.; Hammock, B.D.; Haj, F.G. Soluble epoxide hydrolase deficiency or inhibition attenuates diet-induced endoplasmic reticulum stress in liver and adipose tissue. J. Biol. Chem. 2013, 288, 14189–14199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Cheng, Y.; Li, J.; Wang, Y.; Liu, Y. Epoxy Stearic Acid, an Oxidative Product Derived from Oleic Acid, Induces Cytotoxicity, Oxidative Stress, and Apoptosis in HepG2 Cells. J. Agric. Food. Chem. 2018, 66, 5237–5246. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, J.; Liu, Y. Effects of epoxy stearic acid on lipid metabolism in HepG2 cells. J. Food Sci. 2020, 85, 3644–3652. [Google Scholar] [CrossRef]
- Edin, M.L.; Wang, Z.; Bradbury, J.A.; Graves, J.P.; Lih, F.B.; DeGraff, L.M.; Foley, J.F.; Torphy, R.; Ronnekleiv, O.K.; Tomer, K.B.; et al. Endothelial expression of human cytochrome P450 epoxygenase CYP2C8 increases susceptibility to ischemia-reperfusion injury in isolated mouse heart. FASEB J. 2011, 25, 3436–3447. [Google Scholar] [CrossRef] [Green Version]
- Kundu, S.; Roome, T.; Bhattacharjee, A.; Carnevale, K.A.; Yakubenko, V.P.; Zhang, R.; Hwang, S.H.; Hammock, B.D.; Cathcart, M.K. Metabolic products of soluble epoxide hydrolase are essential for monocyte chemotaxis to MCP-1 in vitro and in vivo. J. Lipid Res. 2013, 54, 436–447. [Google Scholar] [CrossRef] [Green Version]
- Theken, K.N.; Schuck, R.N.; Edin, M.L.; Tran, B.; Ellis, K.; Bass, A.; Lih, F.B.; Tomer, K.B.; Poloyac, S.M.; Wu, M.C.; et al. Evaluation of cytochrome P450-derived eicosanoids in humans with stable atherosclerotic cardiovascular disease. Atherosclerosis 2012, 222, 530–536. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, C.E.; Shuey, M.M.; Milne, G.L.; Gilbert, K.; Hui, N.; Yu, C.; Luther, J.M.; Brown, N.J. Arg287Gln variant of EPHX2 and epoxyeicosatrienoic acids are associated with insulin sensitivity in humans. Prostaglandins Other Lipid Mediat. 2014, 113–115, 38–44. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, S.J.; Kane, A.E.; Hilmer, S.N. Age-related changes in the hepatic pharmacology and toxicology of paracetamol. Curr. Gerontol. Geriatr. Res. 2011, 2011, 624156. [Google Scholar] [CrossRef] [Green Version]
- Nayeem, M.A. Role of oxylipins in cardiovascular diseases. Acta. Pharm. Sin. 2018, 39, 1142–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goetzl, E.J.; Pickett, W.C. The human PMN leukocyte chemotactic activity of complex hydroxy-eicosatetraenoic acids (HETEs). J. Immunol. 1980, 125, 1789–1791. [Google Scholar] [PubMed]
- Valone, F.H.; Franklin, M.; Sun, F.F.; Goetzl, E.J. Alveolar macrophage lipoxygenase products of arachidonic acid: Isolation and recognition as the predominant constituents of the neutrophil chemotactic activity elaborated by alveolar macrophages. Cell Immunol. 1980, 54, 390–401. [Google Scholar] [CrossRef]
- Stenson, W.F.; Parker, C.W. Monohydroxyeicosatetraenoic acids (HETEs) induce degranulation of human neutrophils. J. Immunol. 1980, 124, 2100–2104. [Google Scholar]
- Maayah, Z.H.; El-Kadi, A.O. The role of mid-chain hydroxyeicosatetraenoic acids in the pathogenesis of hypertension and cardiac hypertrophy. Arch. Toxicol. 2016, 90, 119–136. [Google Scholar] [CrossRef]
- Setty, B.N.; Werner, M.H.; Hannun, Y.A.; Stuart, M.J. 15-Hydroxyeicosatetraenoic acid-mediated potentiation of thrombin-induced platelet functions occurs via enhanced production of phosphoinositide-derived second messengers--sn-1,2-diacylglycerol and inositol-1,4,5-trisphosphate. Blood 1992, 80, 2765–2773. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.J.; Justen, J.M.; Nidy, E.G.; Sam, L.M.; Bleasdale, J.E. Transmembrane signaling in human polymorphonuclear neutrophils: 15(S)-hydroxy-(5Z,8Z,11Z,13E)-eicosatetraenoic acid modulates receptor agonist-triggered cell activation. Proc. Natl. Acad. Sci. USA 1993, 90, 7270–7274. [Google Scholar] [CrossRef] [Green Version]
- Petrich, K.; Ludwig, P.; Kühn, H.; Schewe, T. The suppression of 5-lipoxygenation of arachidonic acid in human polymorphonuclear leucocytes by the 15-lipoxygenase product (15S)-hydroxy-(5Z,8Z,11Z,13E)-eicosatetraenoic acid: Structure-activity relationship and mechanism of action. Biochem. J. 1996, 314, 911–916. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.M.; Thompson, H.; Vanden-Heuvel, J.P.; Sun, Y.W.; Trushin, N.; Aliaga, C.; Gowda, K.; Amin, S.; Stanley, B.; Manni, A.; et al. Lipoxygenase catalyzed metabolites derived from docosahexaenoic acid are promising antitumor agents against breast cancer. Sci. Rep. 2021, 11, 410. [Google Scholar] [CrossRef]
- González-Périz, A.; Planagumà, A.; Gronert, K.; Miquel, R.; López-Parra, M.; Titos, E.; Horrillo, R.; Ferré, N.; Deulofeu, R.; Arroyo, V.; et al. Docosahexaenoic acid (DHA) blunts liver injury by conversion to protective lipid mediators: Protectin D1 and 17S-hydroxy-DHA. Faseb J. 2006, 20, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Sapieha, P.; Stahl, A.; Chen, J.; Seaward, M.R.; Willett, K.L.; Krah, N.M.; Dennison, R.J.; Connor, K.M.; Aderman, C.M.; Liclican, E.; et al. 5-Lipoxygenase metabolite 4-HDHA is a mediator of the antiangiogenic effect of ω-3 polyunsaturated fatty acids. Sci. Transl. Med. 2011, 3, 69ra12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeung, J.; Hawley, M.; Holinstat, M. The expansive role of oxylipins on platelet biology. J. Mol. Med. 2017, 95, 575–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikei, K.N.; Yeung, J.; Apopa, P.L.; Ceja, J.; Vesci, J.; Holman, T.R.; Holinstat, M. Investigations of human platelet-type 12-lipoxygenase: Role of lipoxygenase products in platelet activation. J. Lipid Res. 2012, 53, 2546–2559. [Google Scholar] [CrossRef] [PubMed]
- Yeung, J.; Tourdot, B.E.; Adili, R.; Green, A.R.; Freedman, C.J.; Fernandez-Perez, P.; Yu, J.; Holman, T.R.; Holinstat, M. 12(S)-HETrE, a 12-Lipoxygenase Oxylipin of Dihomo-γ-Linolenic Acid, Inhibits Thrombosis via Gαs Signaling in Platelets. Arter. Thromb. Vasc. Biol. 2016, 36, 2068–2077. [Google Scholar] [CrossRef] [Green Version]
- Bednar, M.M.; Gross, C.E.; Russell, S.R.; Fuller, S.P.; Ahern, T.P.; Howard, D.B.; Falck, J.R.; Reddy, K.M.; Balazy, M. 16(R)-hydroxyeicosatetraenoic acid, a novel cytochrome P450 product of arachidonic acid, suppresses activation of human polymorphonuclear leukocyte and reduces intracranial pressure in a rabbit model of thromboembolic stroke. Neurosurgery 2000, 47, 1410–1418; discussion 1418–1419. [Google Scholar] [CrossRef]
- Caligiuri, S.P.; Aukema, H.M.; Ravandi, A.; Pierce, G.N. Elevated levels of pro-inflammatory oxylipins in older subjects are normalized by flaxseed consumption. Exp. Gerontol. 2014, 59, 51–57. [Google Scholar] [CrossRef]
- Teo, K.; Chow, C.K.; Vaz, M.; Rangarajan, S.; Yusuf, S. The Prospective Urban Rural Epidemiology (PURE) study: Examining the impact of societal influences on chronic noncommunicable diseases in low-, middle-, and high-income countries. Am. Heart J. 2009, 158, 1–7.e1. [Google Scholar] [CrossRef]
- Hercberg, S.; Castetbon, K.; Czernichow, S.; Malon, A.; Mejean, C.; Kesse, E.; Touvier, M.; Galan, P. The Nutrinet-Santé Study: A web-based prospective study on the relationship between nutrition and health and determinants of dietary patterns and nutritional status. BMC Public Health 2010, 10, 242. [Google Scholar] [CrossRef]
- Kutzner, L.; Rund, K.M.; Ostermann, A.I.; Hartung, N.M.; Galano, J.M.; Balas, L.; Durand, T.; Balzer, M.S.; David, S.; Schebb, N.H. Development of an Optimized LC-MS Method for the Detection of Specialized Pro-Resolving Mediators in Biological Samples. Front. Pharm. 2019, 10, 169. [Google Scholar] [CrossRef]
- Lillington, J.M.; Trafford, D.J.H.; Makin, H.L.J. A rapid and simple method for the esterification of fatty acids and steroid carboxylic acids prior to gas-liquid chromatography. Clin. Chim. Acta 1981, 111, 91–98. [Google Scholar] [CrossRef]
- Chiuve, S.E.; Fung, T.T.; Rimm, E.B.; Hu, F.B.; McCullough, M.L.; Wang, M.; Stampfer, M.J.; Willett, W.C. Alternative dietary indices both strongly predict risk of chronic disease. J. Nutr. 2012, 142, 1009–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, H.; Hastie, T. Regularization and variable selection via the elastic net. J. R. Stat. Soc. Ser. B (Stat. Methodol.) 2005, 67, 301–320. [Google Scholar] [CrossRef] [Green Version]
- Goeman, J.J. L1 penalized estimation in the Cox proportional hazards model. Biom. J. 2010, 52, 70–84. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.H.; Hastie, T.; Tibshirani, R. Regularization Paths for Generalized Linear Models via Coordinate Descent. J. Stat. Softw. 2010, 33, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Robin, X.; Turck, N.; Hainard, A.; Tiberti, N.; Lisacek, F.; Sanchez, J.C.; Müller, M. pROC: An open-source package for R and S+ to analyze and compare ROC curves. BMC Bioinform. 2011, 12, 77. [Google Scholar] [CrossRef] [PubMed]
- Tibshirani, R.J.; Taylor, J.; Lockhart, R.; Tibshirani, R. Exact Post-Selection Inference for Sequential Regression Procedures. J. Am. Stat. Assoc. 2016, 111, 600–620. [Google Scholar] [CrossRef] [Green Version]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [Green Version]
- Giacomoni, F.; Le Corguillé, G.; Monsoor, M.; Landi, M.; Pericard, P.; Pétéra, M.; Duperier, C.; Tremblay-Franco, M.; Martin, J.F.; Jacob, D.; et al. Workflow4Metabolomics: A collaborative research infrastructure for computational metabolomics. Bioinformatics 2015, 31, 1493–1495. [Google Scholar] [CrossRef] [Green Version]
- van der Kloet, F.M.; Bobeldijk, I.; Verheij, E.R.; Jellema, R.H. Analytical error reduction using single point calibration for accurate and precise metabolomic phenotyping. J. Proteome Res. 2009, 8, 5132–5141. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Discovery Study 1 | Replication Study 2 | |||||
|---|---|---|---|---|---|---|
| Variables | Controls (n = 137) | Cases (n = 137) | p-Value 3 | Controls (n = 101) | Cases (n = 101) | p-Value 3 |
| Men, N (%) | 53 (38.7) | 53 (38.7) | NS | 36 (35.6) | 36 (35.6) | NS |
| Age, mean (SD) | 53.8 (8.4) | 53.8 (8.3) | NS | 61.1 (8.4) | 61. 2 (8.3) | NS |
| Education level (primary/secondary/superior), N (%) | 6/71/60 (4.4/51.8/43.8) | 25/80/32 (18.2/58.4/23.4) | <0.001 | 5/35/61 (4.9/34.7/60.4) | 5/45/51 (4.9/44.6/50.5) | NS |
| Localization (rural/urban), N (%) | 30/107 (21.9/78.1) | 69/68 (50.4/49.6) | <0.001 | 28/73 (27.7/72.3) | 26/75 (25.7/74.3) | NS |
| Smoking status (never/former/current), N (%) | 76/32/29 (55.5/23.4/21.2) | 66/42/29 (48.2/30.7/21.2) | NS | 46/50/5 (45.5/49.5/5.0) | 46/50/5 (45.5/49.5/5.0) | NS |
| Physical activity (low/moderate/intense), N (%) | 0/37/100 (0/27.0/73.0) | 0/32/105 (0/23.4/76.6) | NS | 14/41/46 (13.9/40.6/45.5) | 14/41/46 (13.9/40.6/45.5) | NS |
| Season of blood draw (winter/spring/summer/fall), N (%) | 37/25/15/60 (27.0/18.2/10.9/43.8) | 24/26/25/62 (17.5/19.0/18.2/45.3) | NS | 28/34/14/25 (27.7/33.7/13.9/24.8) | 28/34/14/25 (27.7/33.7/13.9/24.8) | NS |
| Menopausal status (NA/non menop/menop), N (%) | n.d. | n.d. | n.d. | 36/5/60 (35.6/5.0/59.4) | 36/5/60 (35.6/5.0/59.4) | NS |
| AHEI 4 score, mean (SD) | 47.8 (8.9) | 44.9 (8.2) | <0.001 | 53.9 (12.1) | 47.7 (11.6) | <0.01 |
| Waist circumference (cm), mean (SD) | 81 (10) | 100 (14) | <0.001 | 82 (13) | 96 (10) | <0.001 |
| SBP (systolic blood pressure, mmHg), mean (SD) | 139 (20) | 152 (19) | <0.001 | 130 (18) | 143 (16) | <0.001 |
| DBP (diastolic blood pressure, mmHg), mean (SD) | 83 (10) | 90 (10) | <0.001 | 77 (10) | 83 (9) | <0.001 |
| Fasting glucose (mg/dL), mean (SD) | 90.0 (10.7) | 108.5 (21.1) | <0.001 | 89.3 (7.6) | 100.7 (12.5) | <0.001 |
| TG (triglycerides, mg/dL), mean (SD) | 85.3 (34.3) | 160.7 (79.0) | <0.001 | 90.6 (37.9) | 144.9 (60.8) | <0.001 |
| HDLc (high density lipoprotein cholesterol, mg/dL), mean (SD) | 66.9 (15.1) | 50.8 (15.8) | <0.001 | 65.7 (13.5) | 55.9 (13.3) | <0.001 |
| MetS-z-Score 5, mean (SD) | −0.7 (0.6) | 0.7 (0.7) | <0.001 | −0.7 (0.6) | 0.3 (0.5) | <0.001 |
| Weight (kg), mean (SD) | 67 (12) | 85 (17) | <0.001 | 65 (16) | 81 (15) | <0.001 |
| BMI (body mass index, kg/m2), mean (SD) | 25 (4) | 31 (5) | <0.001 | 24 (4) | 29 (5) | <0.001 |
| Hip circumference (cm), mean (SD) | 98 (7) | 109 (10) | <0.001 | 96 (9) | 105 (10) | <0.001 |
| Total cholesterol (mg/dL), mean (SD) | 192.9 (31.3) | 198.7 (38.9) | NS | 230.8 (39.5) | 226.5 (44.2) | NS |
| LDLc (low density lipoprotein cholesterol, mg/dL), mean (SD) | 109.5 (29.9) | 116.5 (34.2) | <0.05 | 146.9 (34.3) | 141.6 (36.1) | NS |
| Visceral fat (kg), mean (SD) | n.d. | n.d. | n.d. | 8 (4) | 11 (4) | <0.001 |
| Visceral mass on body trunk (%) | n.d. | n.d. | n.d. | 24 | 32 | <0.001 |
| Fat mass (kg), mean (SD) | n.d. | n.d. | n.d. | 17 (8) | 27 (10) | <0.001 |
| Fat mass on body trunk (kg), mean (SD) | n.d. | n.d. | n.d. | 9 (5) | 14 (5) | <0.001 |
| Waist circumference, N (%) | 32 (23.4) | 133 (97.1) | <0.001 | 34 (33.7) | 95 (94.1) | <0.001 |
| High blood pressure, N (%) | 89 (65.0) | 129 (94.2) | <0.001 | 50 (49.5) | 98 (97.0) | <0.001 |
| Hypertriglyceridemia, N (%) | 5 (3.6) | 75 (54.7) | <0.001 | 6 (5.9) | 49 (48.5) | <0.001 |
| Low HDLc, N (%) | 2 (1.5) | 50 (36.5) | <0.001 | 1 (1.0) | 27 (26.7) | <0.001 |
| Hyperglycemia, N (%) | 10 (7.3) | 103 (75.2) | <0.001 | 8 (7.9) | 63 (62.4) | <0.001 |
| Odd Ratios Associated with the Variables Selected in the LASSO Models | ||
|---|---|---|
| LASSO Model N°2 (using only the 5 MetS criteria) | LASSO Model N°3 (using both the 23 candidate oxylipins and the 5 MetS criteria) | |
| TG | 1.13 | 1.11 |
| SBP | 1.04 | 1.02 |
| DBP | 1.02 | 1.04 |
| Waist circumference | 1.18 | 1.13 |
| Fasting glucose | 1.15 | 1.10 |
| HDLc | 0.96 | 0.96 |
| 8-HEPE | – | 1.53 |
| 16-HETE | – | 1.16 |
| 12(13)-EpODE | – | 1.01 |
| 5-HETE | – | 0.11 |
| 9-HODE | – | 1.01 |
| 14,15-DiHETrE | – | 1.02 |
| 5-HETrE | – | 0.95 |
| 15-HODE | – | 0.96 |
| 13-oxo-ODE | – | 0.93 |
| 11(12)-EpETrE | – | 0.95 |
| 9-oxo-ODE | – | 1.01 |
| 4-HDHA | – | 0.92 |
| 13-HODE | – | 0.95 |
| 12-HODE | – | 0.91 |
| 7,8-DiHDPE | – | 0.96 |
| 15-HETE | – | 0.86 |
| 5-HEPE | – | 0.71 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dalle, C.; Tournayre, J.; Mainka, M.; Basiak-Rasała, A.; Pétéra, M.; Lefèvre-Arbogast, S.; Dalloux-Chioccioli, J.; Deschasaux-Tanguy, M.; Lécuyer, L.; Kesse-Guyot, E.; et al. The Plasma Oxylipin Signature Provides a Deep Phenotyping of Metabolic Syndrome Complementary to the Clinical Criteria. Int. J. Mol. Sci. 2022, 23, 11688. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms231911688
Dalle C, Tournayre J, Mainka M, Basiak-Rasała A, Pétéra M, Lefèvre-Arbogast S, Dalloux-Chioccioli J, Deschasaux-Tanguy M, Lécuyer L, Kesse-Guyot E, et al. The Plasma Oxylipin Signature Provides a Deep Phenotyping of Metabolic Syndrome Complementary to the Clinical Criteria. International Journal of Molecular Sciences. 2022; 23(19):11688. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms231911688
Chicago/Turabian StyleDalle, Céline, Jérémy Tournayre, Malwina Mainka, Alicja Basiak-Rasała, Mélanie Pétéra, Sophie Lefèvre-Arbogast, Jessica Dalloux-Chioccioli, Mélanie Deschasaux-Tanguy, Lucie Lécuyer, Emmanuelle Kesse-Guyot, and et al. 2022. "The Plasma Oxylipin Signature Provides a Deep Phenotyping of Metabolic Syndrome Complementary to the Clinical Criteria" International Journal of Molecular Sciences 23, no. 19: 11688. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms231911688