
Protective Effect of Rutin on Triethylene Glycol Dimethacrylate-Induced Toxicity through the Inhibition of Caspase Activation and Reactive Oxygen Species Generation in Macrophages
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effects of Rutin on Cytotoxicity Induced by Triethylene Glycol Dimethacrylate (TEGDMA)
2.2. Effects of Rutin on Necrosis and Apoptosis Induced by Triethylene Glycol Dimethacrylate (TEGDMA)
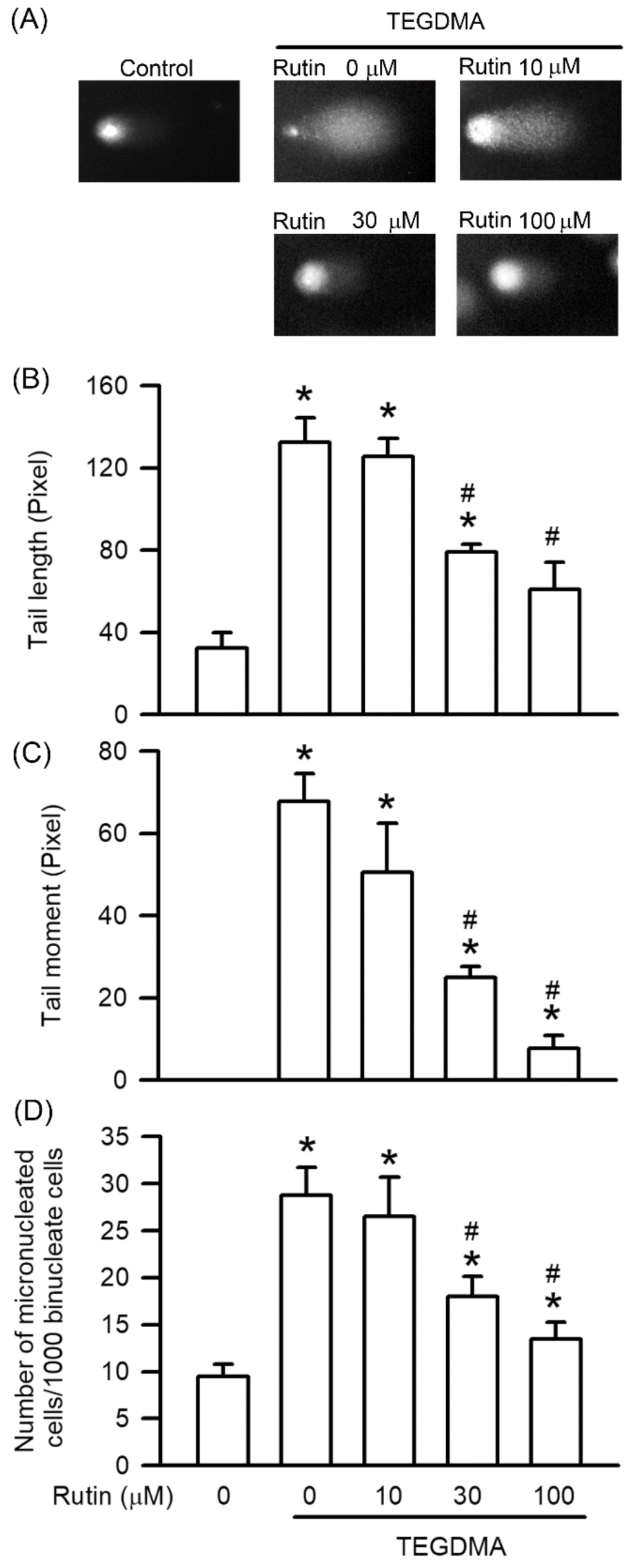
2.3. Effects of Rutin on Genotoxicity Induced by Triethylene Glycol Dimethacrylate (TEGDMA)
2.4. Effects of Rutin on Caspase Activity Induced by Triethylene Glycol Dimethacrylate (TEGDMA)
2.5. Effects of Rutin on Reactive Oxygen Species Generation Induced by Triethylene Glycol Dimethacrylate (TEGDMA)
2.6. Effects of Rutin on Antioxidant Enzyme Activity by Triethylene Glycol Dimethacrylate
2.7. Effects of Rutin on HO-1 and Nrf-2 Expression on Triethylene Glycol Dimethacrylate (TEGDMA)-Treated RAW264.7 Macrophages
2.8. Effects of Rutin on AMPK Phosphorylation Reduced by Triethylene Glycol Dimethacrylate (TEGDMA)
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Treatment
4.2. Cell Viability Assay
4.3. Evaluation of Apoptosis and Necrosis
4.4. Evaluation of Genotoxicity
4.5. Detection of Caspase Activity
4.6. Assessment of Intracellular Reactive Oxygen Species (ROS) Generation
4.7. Detection of Antioxidant Enzyme (AOE) Activity
4.8. Western Blot Analysis
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ganeshpurkar, A.; Saluja, A.K. The pharmacological potential of rutin. J. Saudi Pharm. Soc. 2017, 25, 149–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enogieru, A.B.; Haylett, W.; Hiss, D.C.; Bardien, S.; Ekpo, O.E. Rutin as a Potent Antioxidant: Implications for Neurodegen-erative Disorders. Oxid. Med. Cell. Longev. 2018, 2018, 6241017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandemir, F.M.; Ileriturk, M.; Gur, C. Rutin protects rat liver and kidney from sodium valproate-induce damage by attenuating oxidative stress, ER stress, inflammation, apoptosis and autophagy. Mol. Biol. Rep. 2022, 49, 6063–6074. [Google Scholar] [CrossRef] [PubMed]
- Tung, Y.T.; Zeng, J.L.; Ho, S.T.; Xu, J.W.; Lin, I.H.; Wu, J.H. Djulis Hull Improves Insulin Resistance and Modulates the Gut Microbiota in High-Fat Diet (HFD)-Induced Hyperglycaemia. Antioxidants 2021, 11, 45. [Google Scholar] [CrossRef]
- Surendra, V.; Palei, N.N. Formulation and Characterization of Rutin Loaded Chitosan-alginate Nanoparticles: Antidiabetic and Cytotoxicity Studies. Curr. Drug Deliv. 2022, 19, 379–394. [Google Scholar] [CrossRef]
- Ghanbari-Movahed, M.; Mondal, A.; Farzaei, M.H.; Bishayee, A. Quercetin- and rutin-based nano-formulations for cancer treatment: A systematic review of improved efficacy and molecular mechanisms. Phytomedicine 2022, 97, 153909. [Google Scholar] [CrossRef]
- Shahid, A.; Ali, R.; Ali, N.; Hasan, S.K.; Rashid, S.; Majed, F.; Sultana, S. Attenuation of genotoxicity, oxidative stress, apoptosis and inflammation by rutin in benzo(a)pyrene exposed lungs of mice: Plausible role of NF-κB, TNF-α and Bcl-2. J. Complement. Integr. Med. 2016, 13, 17–29. [Google Scholar] [CrossRef]
- Huang, F.M.; Chang, Y.C.; Su, C.H.; Wu, S.W.; Lee, S.S.; Lee, M.W.; Yeh, K.L.; Chiang, C.Y.; Tu, D.G.; Lu, Y.C.; et al. Rutin-protected BisGMA-induced cytotoxicity, genotoxicity, and apoptosis in macrophages through the reduction of the mitochondrial apoptotic pathway and induction of antioxidant enzymes. Environ. Toxicol. 2020, 36, 45–54. [Google Scholar] [CrossRef]
- Park, M.N.; Um, E.S.; Rahman, M.A.; Kim, J.W.; Park, S.S.; Cho, Y.; Song, H.; Son, S.R.; Jang, D.S.; Kim, W.; et al. Leonurus japonicus Houttuyn induces reactive oxygen species-mediated apoptosis via regulation of miR-19a-3p/PTEN/PI3K/AKT in U937 and THP-1 cells. J. Ethnopharmacol. 2022, 291, 115129. [Google Scholar] [CrossRef]
- Kux, B.J.; Bacigalupo, L.M.; Scriba, A.; Emmrich, M.; Jost-Brinkmann, P.G. Elution study of acrylic monomers from orthodontic materials using high performance liquid chromatography (HPLC). J. Orofac. Orthop. 2022, 83, 34–47. [Google Scholar] [CrossRef]
- Alizadehgharib, S.; Östberg, A.K.; Dahlstrand Rudin, A.; Dahlgren, U.; Christenson, K. The effects of the dental methacrylates TEGDMA, Bis-GMA, and UDMA on neutrophils in vitro. Clin. Exp. Dent. Res. 2020, 6, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Gerzina, T.; Hume, W. Diffusion of monomers from bonding resin-resin composite combinations through dentine in vitro. J. Dent. 1996, 24, 125–128. [Google Scholar] [CrossRef]
- Barreby, E.; Chen, P.; Aouadi, M. Macrophage functional diversity in NAFLD-more than inflammation. Nat. Rev. Endocrinol. 2022, 18, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Herb, M.; Schramm, M. Functions of ROS in Macrophages and Antimicrobial Immunity. Antioxidants 2021, 10, 313. [Google Scholar] [CrossRef]
- Canton, M.; Sánchez-Rodríguez, R.; Spera, I.; Venegas, F.C.; Favia, M.; Viola, A.; Castegna, A. Reactive Oxygen Species in Macrophages: Sources and Targets. Front. Immunol. 2021, 12, 734229. [Google Scholar] [CrossRef]
- Rafiee, A.; Ospina, M.B.; Pitt, T.M.; Quémerais, B. Oxidative stress and DNA damage resulting from welding fumes exposure among professional welders: A systematic review and meta-analysis. Environ. Res. 2022, 214, 114152. [Google Scholar] [CrossRef]
- Murakami, Y.; Kawata, A.; Suzuki, S.; Fujisawa, S. Cytotoxicity and Pro-inflammatory Properties of Aliphatic Alpha, Beta-unsaturated Acid and Ester Monomers in RAW264.7 Cells and Their Chemical Reactivity. In Vivo 2019, 33, 313–323. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.H.; Kim, N.R.; Lim, B.S.; Lee, Y.K.; Yang, H.C. Effects of TEGDMA and HEMA on the expression of COX-2 and iNOS in cultured murine macrophage cells. Dent. Mater. 2009, 25, 240–246. [Google Scholar] [CrossRef]
- Huang, F.M.; Kuan, Y.H.; Lee, S.S.; Chang, Y.C. Cytotoxicity and genotoxicity of triethyleneglycol-dimethacrylate in macrophages involved in DNA damage and caspases activation. Environ. Toxicol. 2015, 30, 581–588. [Google Scholar] [CrossRef]
- Krifka, S.; Hiller, K.A.; Bolay, C.; Petzel, C.; Spagnuolo, G.; Reichl, F.X.; Schmalz, G.; Schweikl, H. Function of MAPK and downstream transcription factors in monomer-induced apoptosis. Biomaterials 2012, 33, 740–750. [Google Scholar] [CrossRef]
- Krifka, S.; Petzel, C.; Bolay, C.; Hiller, K.A.; Spagnuolo, G.; Schmalz, G.; Schweikl, H. Activation of stress-regulated transcription factors by triethylene glycol dimethacrylate monomer. Biomaterials 2011, 32, 1787–1795. [Google Scholar] [CrossRef] [PubMed]
- Gęgotek, A.; Bielawska, K.; Biernacki, M.; Dobrzyńska, I.; Skrzydlewska, E. Time-dependent effect of rutin on skin fibroblasts membrane disruption following UV radiation. Redox. Biol. 2017, 12, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Gęgotek, A.; Rybałtowska-Kawałko, P.; Skrzydlewska, E. Rutin as a Mediator of Lipid Metabolism and Cellular Signaling Pathways Interactions in Fibroblasts Altered by UVA and UVB Radiation. Oxid. Med. Cell. Longev. 2017, 2017, 4721352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, C.H.; Ho, Y.C.; Lee, M.W.; Tseng, C.C.; Lee, S.S.; Hsieh, M.K.; Chen, H.H.; Lee, C.Y.; Wu, S.W.; Kuan, Y.H. 1-Nitropyrene Induced Reactive Oxygen Species-Mediated Apoptosis in Macrophages through AIF Nuclear Translocation and AMPK/Nrf-2/HO-1 Pathway Activation. Oxid. Med. Cell. Longev. 2021, 2021, 9314342. [Google Scholar] [CrossRef] [PubMed]
- Su, C.H.; Chen, S.P.; Chen, L.Y.; Yang, J.J.; Lee, Y.C.; Lee, S.S.; Chen, H.H.; Ng, Y.Y.; Kuan, Y.H. 3-Bromofluoranthene-induced cardiotoxicity of zebrafish and apoptosis in the vascular endothelial cells via intrinsic and extrinsic caspase-dependent pathways. Ecotoxicol. Environ. Saf. 2021, 228, 112962. [Google Scholar] [CrossRef] [PubMed]
- Chiang, Y.W.; Su, C.H.; Sun, H.Y.; Chen, S.P.; Chen, C.J.; Chen, W.Y.; Chang, C.C.; Chen, C.M.; Kuan, Y.H. Bisphenol A induced apoptosis via oxidative stress generation involved Nrf2/HO-1 pathway and mitochondrial dependent pathways in human retinal pigment epithelium (ARPE-19) cells. Environ. Toxicol. 2022, 37, 131–141. [Google Scholar] [CrossRef]
- Wu, S.W.; Su, C.H.; Ho, Y.C.; Huang-Liu, R.; Tseng, C.C.; Chiang, Y.W.; Yeh, K.L.; Lee, S.S.; Chen, W.Y.; Chen, C.J.; et al. Genotoxic effects of 1-nitropyrene in macrophages are mediated through a p53-dependent pathway involving cytochrome c release, caspase activation, and PARP-1 cleavage. Ecotoxicol. Environ. Saf. 2021, 213, 112062. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, X.; Gong, Q.; Li, J.; Huang, H.; Xue, F.; Dai, J.; Tang, F. Extraintestinal Pathogenic Escherichia coli Utilizes Surface-Located Elongation Factor G to Acquire Iron from Holo-Transferrin. Microbiol. Spectr. 2022, 10, e0166221. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, M.; Ericsson, A.C. Function of Macrophages in Disease: Current Understanding on Molecular Mechanisms. Front. Immunol. 2021, 12, 620510. [Google Scholar] [CrossRef]
- Ardura, J.A.; Rackov, G.; Izquierdo, E.; Alonso, V.; Gortazar, A.R.; Escribese, M.M. Targeting Macrophages: Friends or Foes in Disease? Front. Pharmacol. 2019, 10, 1255. [Google Scholar] [CrossRef]
- Al-Hiyasat, A.S.; Darmani, H.; Milhem, M.M. Cytotoxicity evaluation of dental resin composites and their flowable derivatives. Clin. Oral Investig. 2005, 9, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Lovász, B.V.; Berta, G.; Lempel, E.; Sétáló, G., Jr.; Vecsernyés, M.; Szalma, J. TEGDMA (Triethylene Glycol Dimethacrylate) Induces Both Caspase-Dependent and Caspase-Independent Apoptotic Pathways in Pulp Cells. Polymers 2021, 13, 699. [Google Scholar] [CrossRef] [PubMed]
- Baldion, P.A.; Velandia-Romero, M.L.; Castellanos, J.E. Dental resin monomers induce early and potent oxidative damage on human odontoblast-like cells. Chem. Biol. Interact. 2021, 333, 109336. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Yu, Q.; Wu, D.; Liu, R.; Ren, X.; Fu, H.; Zhang, X.; Pan, Y.; Huang, S. JNK-mediated blockage of autophagic flux exacerbates the triethylene glycol dimethacrylate-induced mitochondrial oxidative damage and apoptosis in preodontoblast. Chem. Biol. Interact. 2021, 339, 109432. [Google Scholar] [CrossRef]
- Wang, J.; Wang, R.; Li, J.; Yao, Z. Rutin alleviates cardiomyocyte injury induced by high glucose through inhibiting apoptosis and endoplasmic reticulum stress. Exp. Ther. Med. 2021, 22, 944. [Google Scholar] [CrossRef]
- Meng, X.L.; Yu, M.M.; Liu, Y.C.; Gao, Y.L.; Chen, X.S.; Shou, S.T.; Chai, Y.F. Rutin Inhibits Cardiac Apoptosis and Prevents Sepsis-Induced Cardiomyopathy. Front. Physiol. 2022, 13, 834077. [Google Scholar] [CrossRef]
- Küçükler, S.; Kandemir, F.M.; Özdemir, S.; Çomaklı, S.; Caglayan, C. Protective effects of rutin against deltamethrin-induced hepatotoxicity and nephrotoxicity in rats via regulation of oxidative stress, inflammation, and apoptosis. Environ. Sci. Pollut. Res. Int. 2021, 28, 62975–62990. [Google Scholar] [CrossRef]
- Li, R.; Liu, B.; Xu, W.; Yu, L.; Zhang, C.; Cheng, J.; Tao, L.; Li, Z.; Zhang, Y. DNA damage and cell apoptosis induced by fungicide difenoconazole in mouse mononuclear macrophage RAW264.7. Environ. Toxicol. 2022, 37, 650–659. [Google Scholar] [CrossRef]
- Schaffert, A.; Arnold, J.; Karkossa, I.; Blüher, M.; von Bergen, M.; Schubert, K. The Emerging Plasticizer Alternative DINCH and Its Metabolite MINCH Induce Oxidative Stress and Enhance Inflammatory Responses in Human THP-1 Macrophages. Cells 2021, 10, 2367. [Google Scholar] [CrossRef]
- Styllou, P.; Styllou, M.; Hickel, R.; Högg, C.; Reichl, F.X.; Scherthan, H. NAC ameliorates dental composite-induced DNA double-strand breaks and chromatin condensation. Dent. Mater. J. 2017, 36, 638–646. [Google Scholar] [CrossRef]
- Ginzkey, C.; Zinnitsch, S.; Steussloff, G.; Koehler, C.; Hackenberg, S.; Hagen, R.; Kleinsasser, N.H.; Froelich, K. Assessment of HEMA and TEGDMA induced DNA damage by multiple genotoxicological endpoints in human lymphocytes. Dent. Mater. 2015, 31, 865–876. [Google Scholar] [CrossRef] [PubMed]
- Volk, J.; Leyhausen, G.; Geurtsen, W. Glutathione level and genotoxicity in human oral keratinocytes exposed to TEGDMA. J. Biomed. Mater Res. B Appl. Biomater. 2012, 100, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, A.; Arthur, D.E.; Babangida, S.; Erukainure, O.L.; Malami, I.; Sani, H.; Abdulhamid, A.W.; Ajiboye, I.O.; Saka, A.A.; Hamza, N.M.; et al. Modulatory role of rutin on 2,5-hexanedione-induced chromosomal and DNA damage in rats: Validation of computational predictions. Drug Chem. Toxicol. 2020, 43, 113–126. [Google Scholar] [CrossRef]
- Nassiri-Asl, M.; Ghorbani, A.; Salehisar, S.; Asadpour, E.; Sadeghnia, H.R. Effect of rutin on oxidative DNA damage in PC12 neurons cultured in nutrients deprivation condition. Iran J. Basic Med. Sci. 2020, 23, 390–395. [Google Scholar] [PubMed]
- Shalini, S.; Dorstyn, L.; Dawar, S.; Kumar, S. Old, new and emerging functions of caspases. Cell. Death Differ. 2015, 22, 526–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dogra, A.; Kour, D.; Gour, A.; Bhardwaj, M.; Bag, S.; Dhiman, S.K.; Kumar, A.; Singh, G.; Nandi, U. Ameliorating effect of rutin against diclofenac-induced cardiac injury in rats with underlying function of FABP3, MYL3, and ANP. Drug Chem. Toxicol. 2022, 4, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Liu, C.; Tang, D.; Zheng, Z.; Yan, R.; Wu, C.; Zuo, N.; Ma, J.; He, Y.; Liu, S. The protective effect of rutin against the cisplatin-induced cochlear damage in vitro. Neurotoxicology 2022, 90, 102–111. [Google Scholar] [CrossRef]
- Diessenbacher, P.; Hupe, M.; Sprick, M.R.; Kerstan, A.; Geserick, P.; Haas, T.L.; Wachter, T.; Neumann, M.; Walczak, H.; Silke, J.; et al. NF-kappaB inhibition reveals differential mechanisms of TNF versus TRAIL-induced apoptosis upstream or at the level of caspase-8 activation independent of cIAP2. J. Investig. Dermatol. 2008, 128, 1134–1147. [Google Scholar] [CrossRef] [Green Version]
- Kim, R.; Emi, M.; Tanabe, K. Caspase-dependent and -independent cell death pathways after DNA damage (Review). Oncol. Rep. 2005, 14, 595–599. [Google Scholar] [CrossRef]
- Roux, C.; Jafari, S.M.; Shinde, R.; Duncan, G.; Cescon, D.W.; Silvester, J.; Chu, M.F.; Hodgson, K.; Berger, T.; Wakeham, A.; et al. Reactive oxygen species modulate macrophage immunosuppressive phenotype through the up-regulation of PD-L1. Proc. Natl. Acad. Sci. USA 2019, 116, 4326–4335. [Google Scholar] [CrossRef]
- Liu, B.; Gan, X.; Zhao, Y.; Chen, J.; Yu, H.; Gao, J.; Yu, H. TEGDMA releasing in resin composites with different filler contents and its correlation with mitochondrial mediated cytotoxicity in human gingival fibroblasts. J. Biomed. Mater. Res. A 2019, 107, 1132–1142. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.C.; Chang, J.Z.; Yang, W.H.; Chang, H.H.; Lai, E.H.; Kuo, M.Y. NADPH oxidase 4 is involved in the triethylene glycol dimethacrylate-induced reactive oxygen species and apoptosis in human embryonic palatal mesenchymal and dental pulp cells. Clin. Oral Investig. 2015, 19, 1463–1471. [Google Scholar] [CrossRef] [PubMed]
- Zandi, P.; Schnug, E. Reactive Oxygen Species, Antioxidant Responses and Implications from a Microbial Modulation Perspective. Biology 2022, 11, 155. [Google Scholar] [CrossRef] [PubMed]
- Vijayan, V.; Wagener, F.A.D.T.G.; Immenschuh, S. The macrophage heme-heme oxygenase-1 system and its role in inflammation. Biochem. Pharmacol. 2018, 153, 159–167. [Google Scholar] [CrossRef]
- Loboda, A.; Damulewicz, M.; Pyza, E.; Jozkowicz, A.; Dulak, J. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: An evolutionarily conserved mechanism. Cellular Mol. Life Sci.: CMLS 2016, 73, 3221–3247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matzinger, M.; Fischhuber, K.; Pölöske, D.; Mechtler, K.; Heiss, E.H. AMPK leads to phosphorylation of the transcription factor Nrf2, tuning transactivation of selected target genes. Redox Biol. 2020, 29, 101393. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhao, L.; Liu, Y.; Yu, X.; Qiao, X. Rutin Ameliorates Cadmium-Induced Necroptosis in the Chicken Liver via Inhibiting Oxidative Stress and MAPK/NF-κB Pathway. Biol. Trace Elem. Res. 2022, 200, 1799–1810. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.-C.; Chang, Y.-C.; Yeh, K.-L.; Huang, F.-M.; Su, N.-Y.; Kuan, Y.-H. Protective Effect of Rutin on Triethylene Glycol Dimethacrylate-Induced Toxicity through the Inhibition of Caspase Activation and Reactive Oxygen Species Generation in Macrophages. Int. J. Mol. Sci. 2022, 23, 11773. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms231911773
Yang L-C, Chang Y-C, Yeh K-L, Huang F-M, Su N-Y, Kuan Y-H. Protective Effect of Rutin on Triethylene Glycol Dimethacrylate-Induced Toxicity through the Inhibition of Caspase Activation and Reactive Oxygen Species Generation in Macrophages. International Journal of Molecular Sciences. 2022; 23(19):11773. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms231911773
Chicago/Turabian StyleYang, Li-Chiu, Yu-Chao Chang, Kun-Lin Yeh, Fu-Mei Huang, Ni-Yu Su, and Yu-Hsiang Kuan. 2022. "Protective Effect of Rutin on Triethylene Glycol Dimethacrylate-Induced Toxicity through the Inhibition of Caspase Activation and Reactive Oxygen Species Generation in Macrophages" International Journal of Molecular Sciences 23, no. 19: 11773. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms231911773