
Hypoxia-Regulated Tumor-Derived Exosomes and Tumor Progression: A Focus on Immune Evasion

Abstract
:1. Background
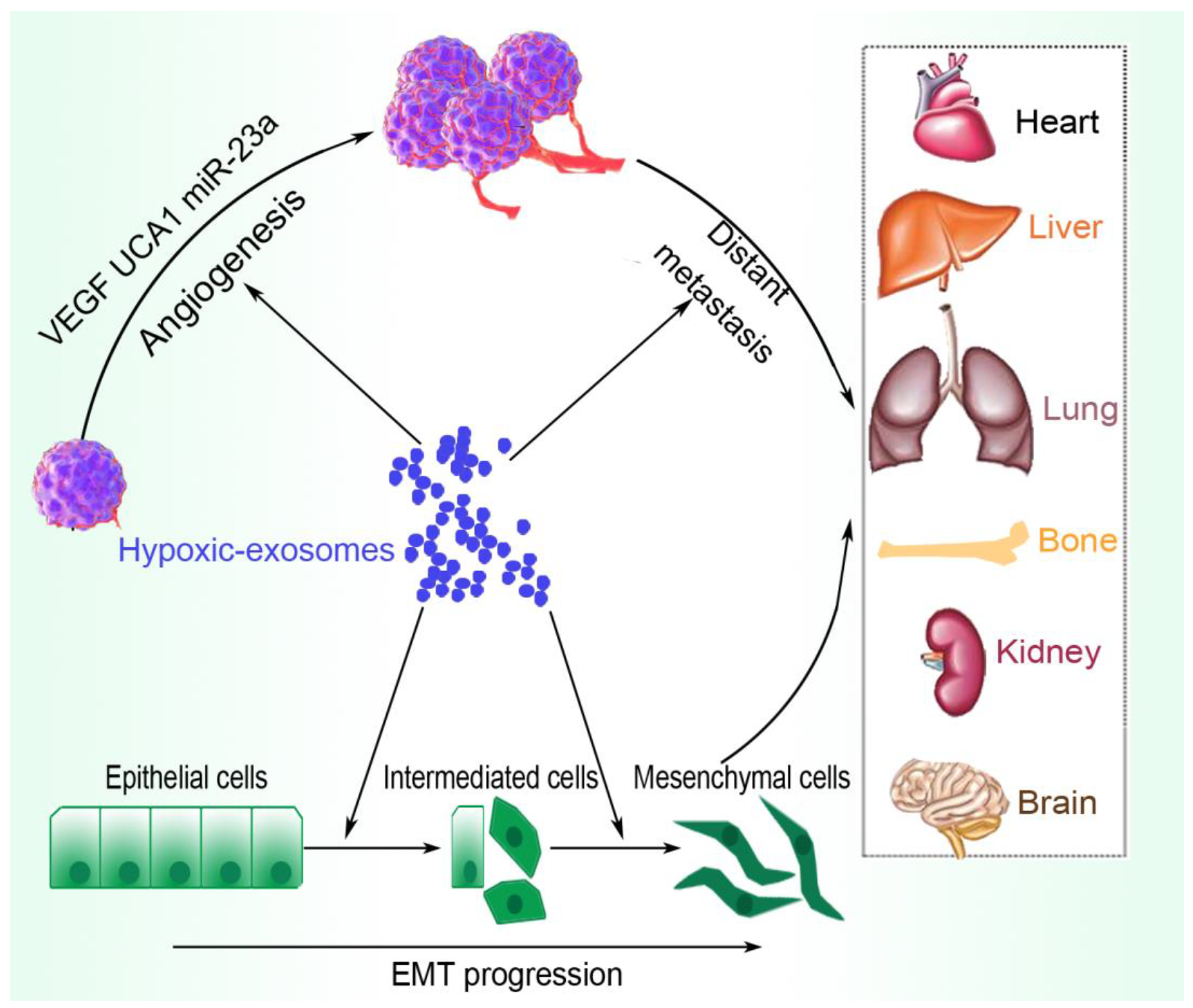
2. Overview of Hypoxic and Tumor-Derived Exosomal Effects on Tumor Progression
2.1. Angiogenesis
2.2. Metastasis and Pre-Metastasis
2.3. EMT
2.4. Autophagy
2.5. Cell Survival and Proliferation
2.6. Therapy Resistance
2.7. Immune Evasion
3. Hypoxia-Inducible Factor (HIF)
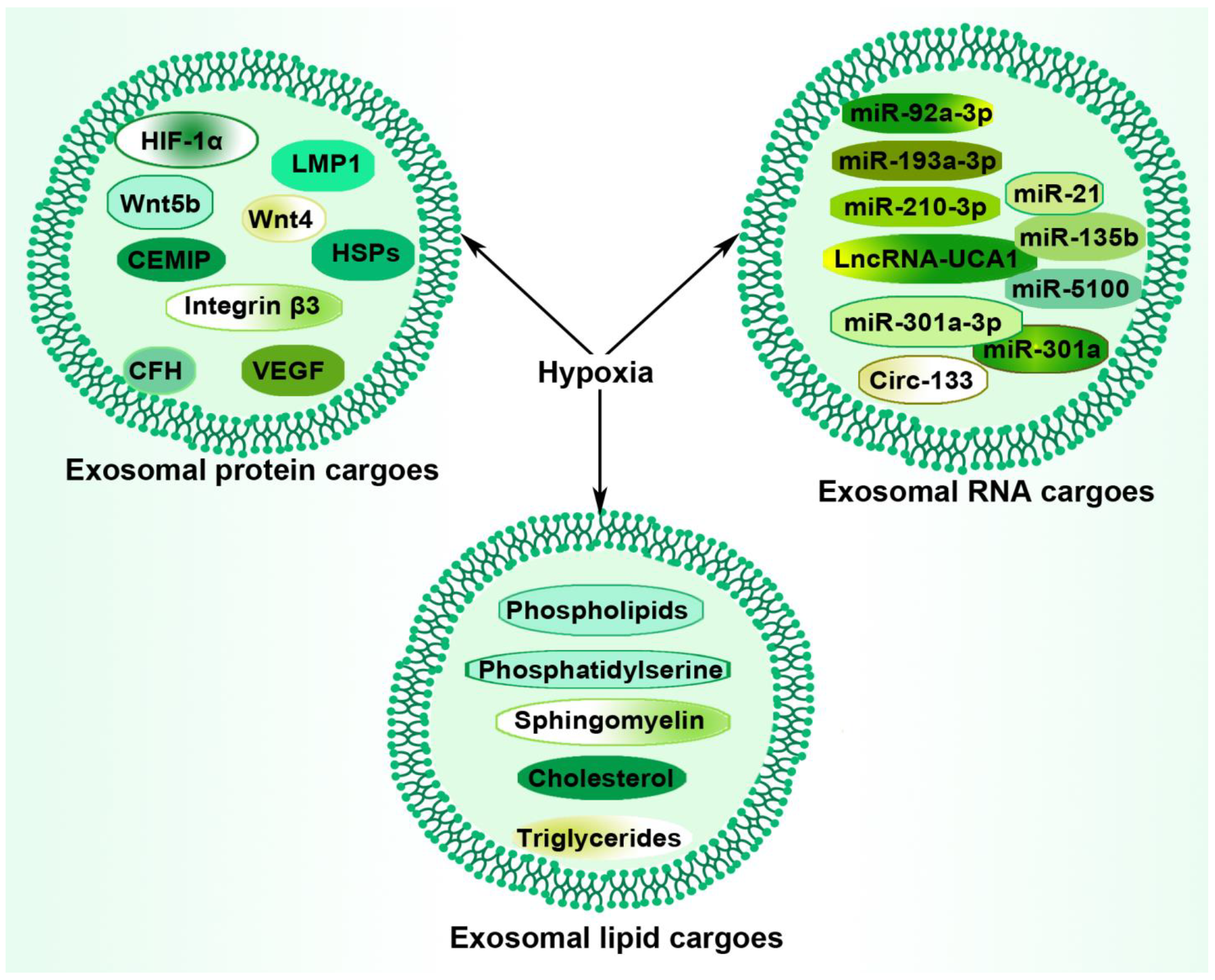
4. Hypoxia and Tumor-Derived Exosomal Components
4.1. Hypoxia and Tumor-Derived Exosomal Protein
4.2. Hypoxia and Tumor-Derived Exosomal RNAs
4.3. Hypoxia and Tumor-Derived Exosomal Lipids
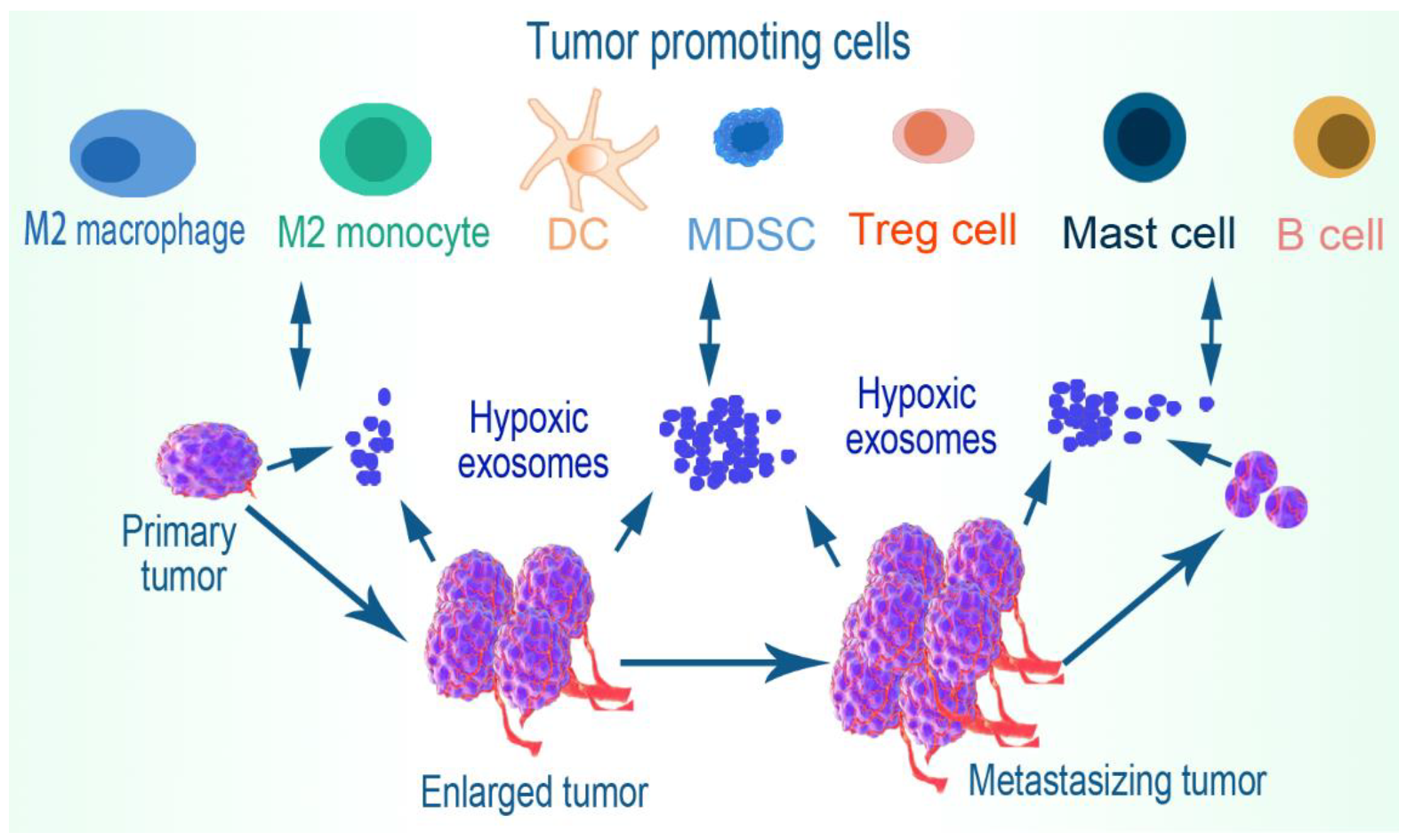
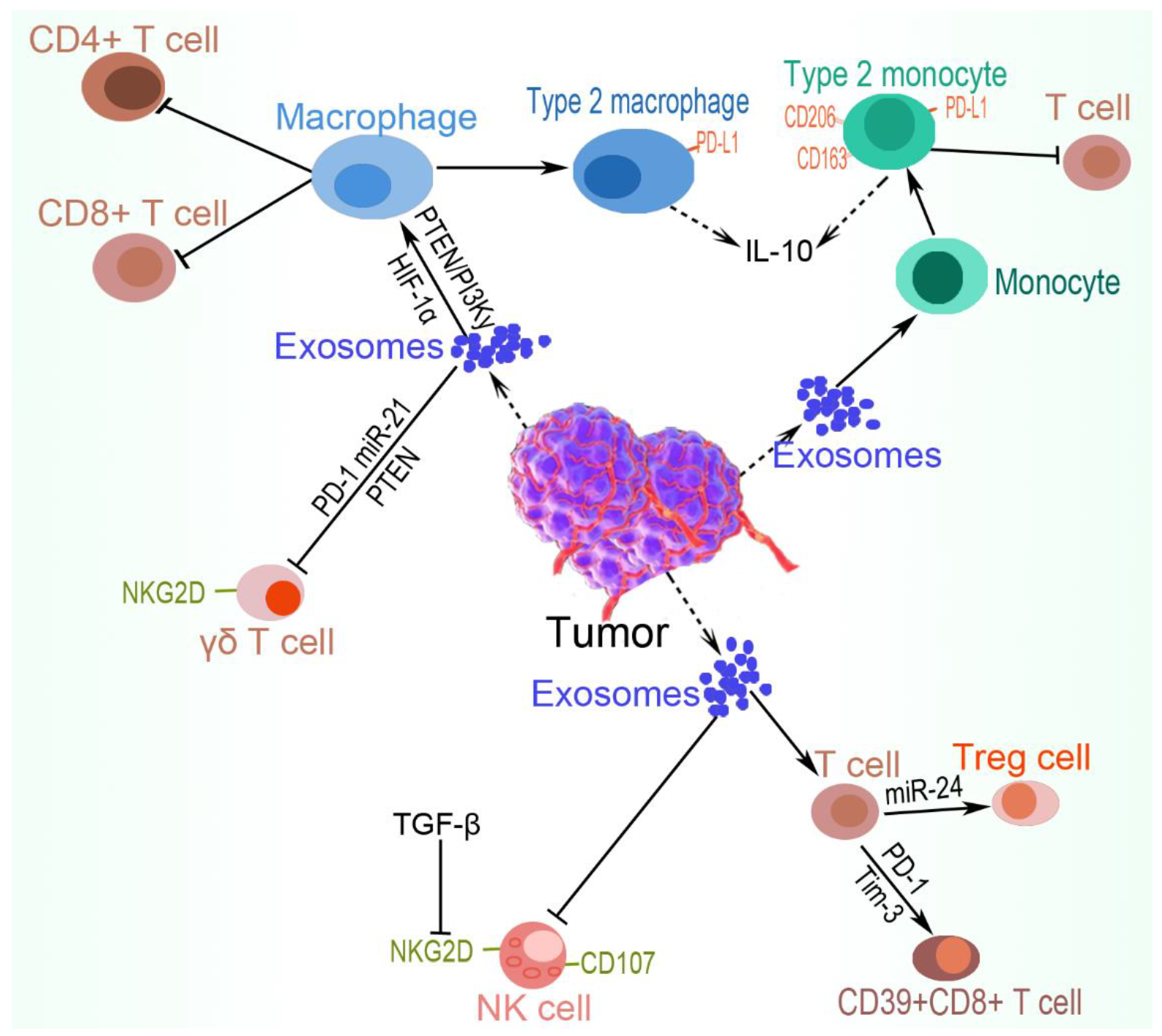
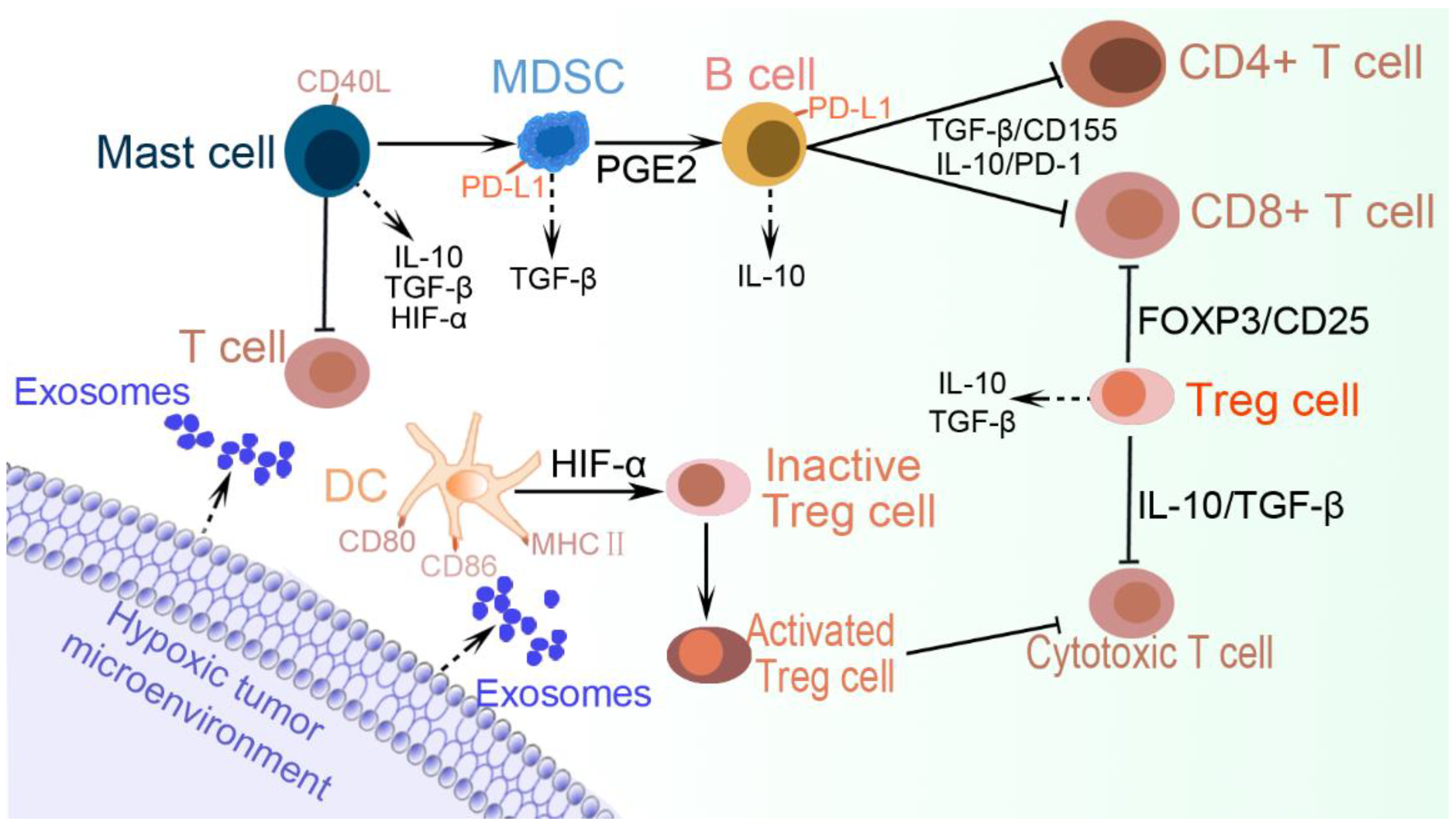
5. Immune Cell Evasion
5.1. Macrophages
5.2. Monocytes
5.3. Natural Killer (NK) Cells
5.4. T Cells
5.5. γδ. T lymphocytes
5.6. Myeloid-Derived Suppressor Cells (MDSCs)
5.7. Dendritic Cells
5.8. B Cells
5.9. Mast Cells
5.10. Regulatory T Cells
6. Escape of Immunogenic Cell Death via DAMPs
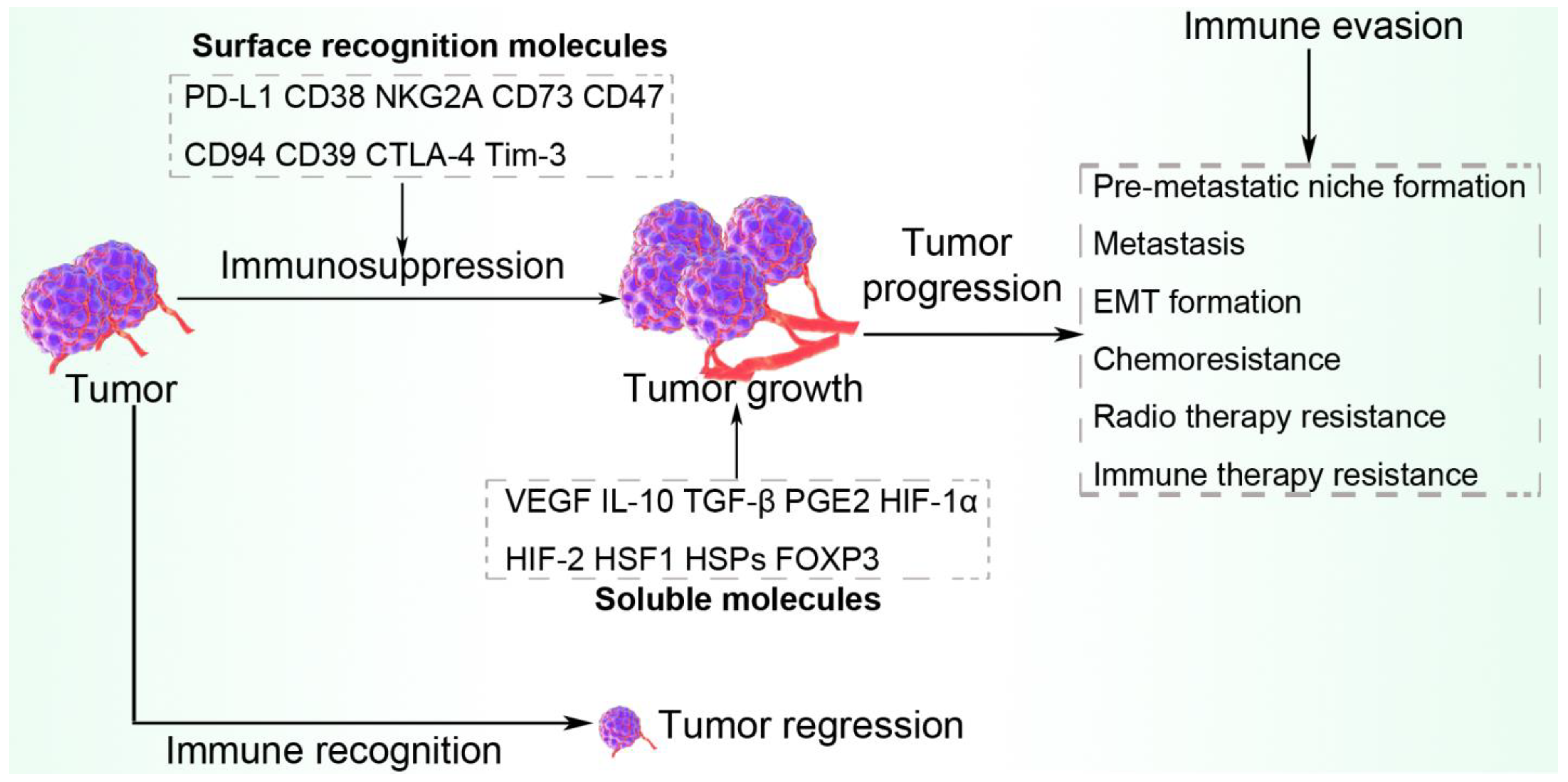
7. Escape from Immune Surveillance via Surface Recognition Molecules
7.1. PD-1/PD-L1
7.2. CD73/CD39
7.3. CD38
7.4. CD47
7.5. Other Surface Recognition Molecules
8. Escape from Immune Surveillance via Antitumor-Suppressive Molecules
8.1. TGF-β
8.2. IL-10
8.3. PGE2
8.4. VEGF
8.5. Others
9. Conclusions and Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Meng, W.; Hao, Y.; He, C.; Li, L.; Zhu, G. Exosome-Orchestrated Hypoxic Tumor Microenvironment. Mol. Cancer 2019, 18, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, X.; Qu, X.; Tian, R.; Xu, L.; Jin, X.; Yu, S.; Zhao, Y.; Ma, J.; Liu, Y.; Sun, L.; et al. Hypoxia-Induced NAD+ Interventions Promote Tumor Survival and Metastasis by Regulating Mitochondrial Dynamics. Life Sci. 2020, 259, 118171. [Google Scholar] [CrossRef] [PubMed]
- Joseph, J.P.; Harishankar, M.K.; Pillai, A.A.; Devi, A. Hypoxia Induced EMT: A Review on the Mechanism of Tumor Progression and Metastasis in OSCC. Oral Oncol. 2018, 80, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Jing, X.; Jiang, T.; Dai, L.; Wang, X.; Jia, L.; Wang, H.; An, L.; Liu, M.; Zhang, R.; Cheng, Z. Hypoxia-Induced Autophagy Activation through NF-ΚB Pathway Regulates Cell Proliferation and Migration to Induce Pulmonary Vascular Remodeling. Exp. Cell Res. 2018, 368, 174–183. [Google Scholar] [CrossRef]
- Liu, L.-L.; Li, D.; He, Y.-L.; Zhou, Y.-Z.; Gong, S.-H.; Wu, L.-Y.; Zhao, Y.-Q.; Huang, X.; Zhao, T.; Xu, L.; et al. MiR-210 Protects Renal Cell Against Hypoxia-Induced Apoptosis by Targeting HIF-1 Alpha. Mol. Med. 2017, 23, 258–271. [Google Scholar] [CrossRef] [Green Version]
- Saggar, J.K.; Yu, M.; Tan, Q.; Tannock, I.F. The Tumor Microenvironment and Strategies to Improve Drug Distribution. Front. Oncol. 2013, 3, 154. [Google Scholar] [CrossRef] [Green Version]
- Vaupel, P.; Multhoff, G. Fatal Alliance of Hypoxia-/HIF-1α-Driven Microenvironmental Traits Promoting Cancer Progression. In Oxygen Transport to Tissue XLI; Advances in Experimental Medicine and Biology; Springer: Berlin, Germany, 2020; Volume 1232, pp. 169–176. [Google Scholar] [CrossRef]
- Multhoff, G.; Vaupel, P. Hypoxia Compromises Anti-Cancer Immune Responses. In Oxygen Transport to Tissue XLI; Advances in Experimental Medicine and Biology; Springer: Berlin, Germany, 2020; Volume 1232, pp. 131–143. [Google Scholar] [CrossRef]
- Li, Y.; Patel, S.P.; Roszik, J.; Qin, Y. Hypoxia-Driven Immunosuppressive Metabolites in the Tumor Microenvironment: New Approaches for Combinational Immunotherapy. Front. Immunol. 2018, 9, 1591. [Google Scholar] [CrossRef] [Green Version]
- Morand, G.B.; Broglie, M.A.; Schumann, P.; Huellner, M.W.; Rupp, N.J. Histometabolic Tumor Imaging of Hypoxia in Oral Cancer: Clinicopathological Correlation for Prediction of an Aggressive Phenotype. Front. Oncol. 2020, 10, 1670. [Google Scholar] [CrossRef]
- Wigerup, C.; Påhlman, S.; Bexell, D. Therapeutic Targeting of Hypoxia and Hypoxia-Inducible Factors in Cancer. Pharmacol. Ther. 2016, 164, 152–169. [Google Scholar] [CrossRef] [Green Version]
- Vito, A.; El-Sayes, N.; Mossman, K. Hypoxia-Driven Immune Escape in the Tumor Microenvironment. Cells 2020, 9, 992. [Google Scholar] [CrossRef]
- Zhao, S.; Ren, S.; Jiang, T.; Zhu, B.; Li, X.; Zhao, C.; Jia, Y.; Shi, J.; Zhang, L.; Liu, X.; et al. Low-Dose Apatinib Optimizes Tumor Microenvironment and Potentiates Antitumor Effect of PD-1/PD-L1 Blockade in Lung Cancer. Cancer Immunol. Res. 2019, 7, 630–643. [Google Scholar] [CrossRef] [PubMed]
- Marar, C.; Starich, B.; Wirtz, D. Extracellular Vesicles in Immunomodulation and Tumor Progression. Nat. Immunol. 2021, 22, 560–570. [Google Scholar] [CrossRef] [PubMed]
- Ocansey, D.K.W.; Xu, X.; Zhang, L.; Mao, F. Mesenchymal Stem Cell-Derived Exosome: The Likely Game-Changer in Stem Cell Research. BIOCELL 2022, 46, 1169–1172. [Google Scholar] [CrossRef]
- Saber, S.H.; Ali, H.E.A.; Gaballa, R.; Gaballah, M.; Ali, H.I.; Zerfaoui, M.; Abd Elmageed, Z.Y. Exosomes Are the Driving Force in Preparing the Soil for the Metastatic Seeds: Lessons from the Prostate Cancer. Cells 2020, 9, 564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Sai, B.; Wang, F.; Wang, L.; Wang, Y.; Zheng, L.; Li, G.; Tang, J.; Xiang, J. Hypoxic BMSC-Derived Exosomal MiRNAs Promote Metastasis of Lung Cancer Cells via STAT3-Induced EMT. Mol. Cancer 2019, 18, 40. [Google Scholar] [CrossRef] [Green Version]
- Xue, M.; Chen, W.; Xiang, A.; Wang, R.; Chen, H.; Pan, J.; Pang, H.; An, H.; Wang, X.; Hou, H.; et al. Hypoxic Exosomes Facilitate Bladder Tumor Growth and Development through Transferring Long Non-Coding RNA-UCA1. Mol. Cancer 2017, 16, 143. [Google Scholar] [CrossRef] [Green Version]
- Baginska, J.; Viry, E.; Paggetti, J.; Medves, S.; Berchem, G.; Moussay, E.; Janji, B. The Critical Role of the Tumor Microenvironment in Shaping Natural Killer Cell-Mediated Anti-Tumor Immunity. Front. Immunol. 2013, 4, 490. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Deep, G. Hypoxia in Tumor Microenvironment Regulates Exosome Biogenesis: Molecular Mechanisms and Translational Opportunities. Cancer Lett. 2020, 479, 23–30. [Google Scholar] [CrossRef]
- Shao, C.; Yang, F.; Miao, S.; Liu, W.; Wang, C.; Shu, Y.; Shen, H. Role of Hypoxia-Induced Exosomes in Tumor Biology. Mol. Cancer 2018, 17, 120. [Google Scholar] [CrossRef]
- You, L.; Wu, W.; Wang, X.; Fang, L.; Adam, V.; Nepovimova, E.; Wu, Q.; Kuca, K. The Role of Hypoxia-inducible Factor 1 in Tumor Immune Evasion. Med. Res. Rev. 2020, 41, 1622–1643. [Google Scholar] [CrossRef]
- Vaupel, P.; Multhoff, G. Hypoxia-/HIF-1α-Driven Factors of the Tumor Microenvironment Impeding Antitumor Immune Responses and Promoting Malignant Progression. Adv. Exp. Med. Biol. 2018, 1072, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Park, J.E.; Dutta, B.; Tse, S.W.; Gupta, N.; Tan, C.F.; Low, J.K.; Yeoh, K.W.; Kon, O.L.; Tam, J.P.; Sze, S.K. Hypoxia-Induced Tumor Exosomes Promote M2-like Macrophage Polarization of Infiltrating Myeloid Cells and MicroRNA-Mediated Metabolic Shift. Oncogene 2019, 38, 5158–5173. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Wang, S.; Ni, B.; Xing, S.; Cao, H.; Zhang, Z.; Yu, F.; Zhao, E.; Zhao, G. Hypoxic Gastric Cancer-Derived Exosomes Promote Progression and Metastasis via MiR-301a-3p/PHD3/HIF-1α Positive Feedback Loop. Oncogene 2020, 39, 6231–6244. [Google Scholar] [CrossRef]
- Li, L.; Li, C.; Wang, S.; Wang, Z.; Jiang, J.; Wang, W.; Li, X.; Chen, J.; Liu, K.; Li, C.; et al. Exosomes Derived from Hypoxic Oral Squamous Cell Carcinoma Cells Deliver MiR-21 to Normoxic Cells to Elicit a Prometastatic Phenotype. Cancer Res. 2016, 76, 1770–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, C.; Chen, M.; Jiang, R.; Guo, Y.; Wu, M.; Zhang, X. Exosome-Related Tumor Microenvironment. J. Cancer 2018, 9, 3084–3092. [Google Scholar] [CrossRef] [Green Version]
- Olejarz, W.; Kubiak-Tomaszewska, G.; Chrzanowska, A.; Lorenc, T. Exosomes in Angiogenesis and Anti-Angiogenic Therapy in Cancers. Int. J. Mol. Sci. 2020, 21, 5840. [Google Scholar] [CrossRef] [PubMed]
- Ohyashiki, J.H.; Umezu, T.; Ohyashiki, K. Exosomes Promote Bone Marrow Angiogenesis in Hematologic Neoplasia. Curr. Opin. Hematol. 2016, 23, 268–273. [Google Scholar] [CrossRef]
- Guo, Z.; Wang, X.; Yang, Y.; Chen, W.; Zhang, K.; Teng, B.; Huang, C.; Zhao, Q.; Qiu, Z. Hypoxic Tumor-Derived Exosomal Long Noncoding RNA UCA1 Promotes Angiogenesis via MiR-96-5p/AMOTL2 in Pancreatic Cancer. Mol. Ther.—Nucleic Acids 2020, 22, 179–195. [Google Scholar] [CrossRef]
- Mao, Y.; Wang, Y.; Dong, L.; Zhang, Y.; Zhang, Y.; Wang, C.; Zhang, Q.; Yang, S.; Cao, L.; Zhang, X.; et al. Hypoxic Exosomes Facilitate Angiogenesis and Metastasis in Esophageal Squamous Cell Carcinoma through Altering the Phenotype and Transcriptome of Endothelial Cells. J. Exp. Clin. Cancer Res. 2019, 38, 389. [Google Scholar] [CrossRef] [Green Version]
- Hsu, Y.-L.; Hung, J.-Y.; Chang, W.-A.; Lin, Y.-S.; Pan, Y.-C.; Tsai, P.-H.; Wu, C.-Y.; Kuo, P.-L. Hypoxic Lung Cancer-Secreted Exosomal MiR-23a Increased Angiogenesis and Vascular Permeability by Targeting Prolyl Hydroxylase and Tight Junction Protein ZO-1. Oncogene 2017, 36, 4929–4942. [Google Scholar] [CrossRef]
- Guan, X. Cancer Metastases: Challenges and Opportunities. Acta Pharm. Sin. B 2015, 5, 402–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshino, A.; Costa-Silva, B.; Shen, T.-L.; Rodrigues, G.; Hashimoto, A.; Tesic Mark, M.; Molina, H.; Kohsaka, S.; Di Giannatale, A.; Ceder, S.; et al. Tumour Exosome Integrins Determine Organotropic Metastasis. Nature 2015, 527, 329–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Y.; Song, X.; Li, Y.; Chen, B.; Zhao, W.; Wang, L.; Zhang, H.; Liu, Y.; Han, D.; Zhang, N.; et al. LncRNA BCRT1 Promotes Breast Cancer Progression by Targeting MiR-1303/PTBP3 Axis. Mol. Cancer 2020, 19, 85. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zhang, H.; Yang, Y.; Wang, X.; Deng, T.; Liu, R.; Ning, T.; Bai, M.; Li, H.; Zhu, K.; et al. Hypoxia Induced Exosomal CircRNA Promotes Metastasis of Colorectal Cancer via Targeting GEF-H1/RhoA Axis. Theranostics 2020, 10, 8211–8226. [Google Scholar] [CrossRef] [PubMed]
- Bao, B.; Azmi, A.S.; Ali, S.; Ahmad, A.; Li, Y.; Banerjee, S.; Kong, D.; Sarkar, F.H. The Biological Kinship of Hypoxia with CSC and EMT and Their Relationship with Deregulated Expression of MiRNAs and Tumor Aggressiveness. Biochim. Biophys. Acta—Rev. Cancer 2012, 1826, 272–296. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Liao, W.-L.; Lu, Q.-J.; Zhang, P.; Zhu, J.; Jiang, G.-N. Hypoxic Tumor-Derived Exosomal Circular RNA SETDB1 Promotes Invasive Growth and EMT via the MiR-7/Sp1 Axis in Lung Adenocarcinoma. Mol. Ther.—Nucleic Acids 2021, 23, 1078–1092. [Google Scholar] [CrossRef]
- Ahuja, N.; Ashok, C.; Natua, S.; Pant, D.; Cherian, A.; Pandkar, M.R.; Yadav, P.; Vishnu, N.S.S.; Mishra, J.; Samaiya, A.; et al. Hypoxia-Induced TGF-β–RBFOX2–ESRP1 Axis Regulates Human MENA Alternative Splicing and Promotes EMT in Breast Cancer. NAR Cancer 2020, 2, zcaa021. [Google Scholar] [CrossRef]
- Santana-Codina, N.; Mancias, J.D.; Kimmelman, A.C. The Role of Autophagy in Cancer. Annu. Rev. Cancer Biol. 2017, 1, 19–39. [Google Scholar] [CrossRef]
- Choi, K.S. Autophagy and Cancer. Exp. Mol. Med. 2012, 44, 109–120. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Ni, Y.; Zhang, L.; Jiang, R.; Xu, J.; Yang, H.; Hu, Y.; Qiu, J.; Pu, L.; Tang, J.; et al. HIF-1α-Induced Expression of M6A Reader YTHDF1 Drives Hypoxia-Induced Autophagy and Malignancy of Hepatocellular Carcinoma by Promoting ATG2A and ATG14 Translation. Signal Transduct. Target. Ther. 2021, 6, 76. [Google Scholar] [CrossRef]
- Abdel Karim, N.; Gaber, O.; Eldessouki, I.; Bahassi, E.M.; Morris, J. Exosomes as a Surrogate Marker for Autophagy in Peripheral Blood, Correlative Data from Phase I Study of Chloroquine in Combination with Carboplatin/Gemcitabine in Advanced Solid Tumors. Asian Pac. J. Cancer Prev. 2019, 20, 3789–3796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Chu, L.; Li, J.; Shi, Y.; Xu, B.; Gu, J.; Yao, X.; Tian, M.; Yang, X.; Sun, X. Hypoxia Induced Changes in MiRNAs and Their Target MRNAs in Extracellular Vesicles of Esophageal Squamous Cancer Cells. Thorac. Cancer 2020, 11, 570–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Min, Z.; Zhou, Z.; Linhong, M.; Tao, R.; Yan, L.; Song, H. Hypoxia-Induced Exosomes Promote Hepatocellular Carcinoma Proliferation and Metastasis via MiR-1273f Transfer. Exp. Cell Res. 2019, 385, 111649. [Google Scholar] [CrossRef] [PubMed]
- Kucharzewska, P.; Christianson, H.C.; Welch, J.E.; Svensson, K.J.; Fredlund, E.; Ringner, M.; Morgelin, M.; Bourseau-Guilmain, E.; Bengzon, J.; Belting, M. Exosomes Reflect the Hypoxic Status of Glioma Cells and Mediate Hypoxia-Dependent Activation of Vascular Cells during Tumor Development. Proc. Natl. Acad. Sci. USA 2013, 110, 7312–7317. [Google Scholar] [CrossRef] [Green Version]
- Richards, K.E.; Zeleniak, A.E.; Fishel, M.L.; Wu, J.; Littlepage, L.E.; Hill, R. Cancer-Associated Fibroblast Exosomes Regulate Survival and Proliferation of Pancreatic Cancer Cells. Oncogene 2017, 36, 1770–1778. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Zhou, L.; Tey, S.K.; Ma, A.P.Y.; Yeung, C.L.S.; Ng, T.H.; Wong, S.W.K.; Liu, B.H.M.; Fung, Y.M.E.; Patz, E.F.; et al. Tumour Extracellular Vesicle-derived Complement Factor H Promotes Tumorigenesis and Metastasis by Inhibiting Complement-dependent Cytotoxicity of Tumour Cells. J. Extracell. Vesicles 2020, 10, e12031. [Google Scholar] [CrossRef]
- Mashouri, L.; Yousefi, H.; Aref, A.R.; Ahadi, A.M.; Molaei, F.; Alahari, S.K. Exosomes: Composition, Biogenesis, and Mechanisms in Cancer Metastasis and Drug Resistance. Mol. Cancer 2019, 18, 75. [Google Scholar] [CrossRef] [Green Version]
- Olejarz, W.; Dominiak, A.; Żołnierzak, A.; Kubiak-Tomaszewska, G.; Lorenc, T. Tumor-Derived Exosomes in Immunosuppression and Immunotherapy. J. Immunol. Res. 2020, 2020, 6272498. [Google Scholar] [CrossRef]
- Xie, F.; Xu, M.; Lu, J.; Mao, L.; Wang, S. The Role of Exosomal PD-L1 in Tumor Progression and Immunotherapy. Mol. Cancer 2019, 18, 146. [Google Scholar] [CrossRef] [Green Version]
- Yue, X.; Lan, F.; Xia, T. Hypoxic Glioma Cell-Secreted Exosomal MiR-301a Activates Wnt/β-Catenin Signaling and Promotes Radiation Resistance by Targeting TCEAL7. Mol. Ther. 2019, 27, 1939–1949. [Google Scholar] [CrossRef]
- Narayanan, D.; Ma, S.; Özcelik, D. Targeting the Redox Landscape in Cancer Therapy. Cancers 2020, 12, 1706. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.; Kim, S.; Hong, B.-J.; Lee, C.-J.; Kim, Y.-E.; Bok, S.; Oh, J.-M.; Gwak, S.-H.; Yoo, M.Y.; Lee, M.S.; et al. Tumor-Associated Macrophages Enhance Tumor Hypoxia and Aerobic Glycolysis. Cancer Res. 2019, 79, 795–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loeuillard, E.; Yang, J.; Buckarma, E.; Wang, J.; Liu, Y.; Conboy, C.; Pavelko, K.D.; Li, Y.; O’Brien, D.; Wang, C.; et al. Targeting Tumor-Associated Macrophages and Granulocytic Myeloid-Derived Suppressor Cells Augments PD-1 Blockade in Cholangiocarcinoma. J. Clin. Investig. 2020, 130, 5380–5396. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Qiu, W.; Liu, Q.; Qian, M.; Wang, S.; Zhang, Z.; Gao, X.; Chen, Z.; Xue, H.; Li, G. Immunosuppressive Effects of Hypoxia-Induced Glioma Exosomes through Myeloid-Derived Suppressor Cells via the MiR-10a/Rora and MiR-21/Pten Pathways. Oncogene 2018, 37, 4239–4259. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.-B.; Zhang, H.; Cai, T.-T.; Liu, Y.-N.; Ni, J.-J.; He, J.; Peng, J.-Y.; Chen, Q.-Y.; Mo, H.-Y.; Jun-Cui; et al. Exosomal MiR-24-3p Impedes T-Cell Function by Targeting FGF11 and Serves as a Potential Prognostic Biomarker for Nasopharyngeal Carcinoma. J. Pathol. 2016, 240, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Sormendi, S.; Wielockx, B. Hypoxia Pathway Proteins As Central Mediators of Metabolism in the Tumor Cells and Their Microenvironment. Front. Immunol. 2018, 9, 40. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Lin, D.; Taniguchi, C.M. Hypoxia Inducible Factor (HIF) in the Tumor Microenvironment: Friend or Foe? Sci. China Life Sci. 2017, 60, 1114–1124. [Google Scholar] [CrossRef]
- Luo, W.; Wang, Y. Hypoxia Mediates Tumor Malignancy and Therapy Resistance. In Hypoxia and Cancer Metastasis; Advances in Experimental Medicine and Biology; Springer: Berlin, Germany, 2019; Volume 1136, pp. 1–18. [Google Scholar] [CrossRef]
- Zhou, X.; Liu, H.; Zheng, Y.; Han, Y.; Wang, T.; Zhang, H.; Sun, Q.; Li, Z. Overcoming Radioresistance in Tumor Therapy by Alleviating Hypoxia and Using the HIF-1 Inhibitor. ACS Appl. Mater. Interfaces 2020, 12, 4231–4240. [Google Scholar] [CrossRef]
- Zhang, H.; Lu, H.; Xiang, L.; Bullen, J.W.; Zhang, C.; Samanta, D.; Gilkes, D.M.; He, J.; Semenza, G.L. HIF-1 Regulates CD47 Expression in Breast Cancer Cells to Promote Evasion of Phagocytosis and Maintenance of Cancer Stem Cells. Proc. Natl. Acad. Sci. USA 2015, 112, E6215–E6223. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Sun, C.; Qin, Z. Metabolic Reprogramming of Cancer-Associated Fibroblasts and Its Effect on Cancer Cell Reprogramming. Theranostics 2021, 11, 8322–8336. [Google Scholar] [CrossRef]
- Semenza, G.L. HIF-1: Upstream and Downstream of Cancer Metabolism. Curr. Opin. Genet. Dev. 2010, 20, 51–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kodama, M.; Oshikawa, K.; Shimizu, H.; Yoshioka, S.; Takahashi, M.; Izumi, Y.; Bamba, T.; Tateishi, C.; Tomonaga, T.; Matsumoto, M.; et al. A Shift in Glutamine Nitrogen Metabolism Contributes to the Malignant Progression of Cancer. Nat. Commun. 2020, 11, 1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peiris-Pagès, M.; Martinez-Outschoorn, U.E.; Pestell, R.G.; Sotgia, F.; Lisanti, M.P. Cancer Stem Cell Metabolism. Breast Cancer Res. 2016, 18, 55. [Google Scholar] [CrossRef]
- Ocansey, D.K.W.; Zhang, L.; Wang, Y.; Yan, Y.; Qian, H.; Zhang, X.; Xu, W.; Mao, F. Exosome-mediated Effects and Applications in Inflammatory Bowel Disease. Biol. Rev. 2020, 95, 1287–1307. [Google Scholar] [CrossRef] [PubMed]
- Milane, L.; Singh, A.; Mattheolabakis, G.; Suresh, M.; Amiji, M.M. Exosome Mediated Communication within the Tumor Microenvironment. J. Control. Release 2015, 219, 278–294. [Google Scholar] [CrossRef] [PubMed]
- Panigrahi, G.K.; Praharaj, P.P.; Peak, T.C.; Long, J.; Singh, R.; Rhim, J.S.; Abd Elmageed, Z.Y.; Deep, G. Hypoxia-Induced Exosome Secretion Promotes Survival of African-American and Caucasian Prostate Cancer Cells. Sci. Rep. 2018, 8, 3853. [Google Scholar] [CrossRef]
- Zhang, W.; Zhou, X.; Yao, Q.; Liu, Y.; Zhang, H.; Dong, Z. HIF-1-Mediated Production of Exosomes during Hypoxia Is Protective in Renal Tubular Cells. Am. J. Physiol. Physiol. 2017, 313, F906–F913. [Google Scholar] [CrossRef]
- Aga, M.; Bentz, G.L.; Raffa, S.; Torrisi, M.R.; Kondo, S.; Wakisaka, N.; Yoshizaki, T.; Pagano, J.S.; Shackelford, J. Exosomal HIF1α Supports Invasive Potential of Nasopharyngeal Carcinoma-Associated LMP1-Positive Exosomes. Oncogene 2014, 33, 4613–4622. [Google Scholar] [CrossRef] [Green Version]
- Kore, R.A.; Edmondson, J.L.; Jenkins, S.V.; Jamshidi-Parsian, A.; Dings, R.P.M.; Reyna, N.S.; Griffin, R.J. Hypoxia-Derived Exosomes Induce Putative Altered Pathways in Biosynthesis and Ion Regulatory Channels in Glioblastoma Cells. Biochem. Biophys. Rep. 2018, 14, 104–113. [Google Scholar] [CrossRef]
- Huang, Z.; Yang, M.; Li, Y.; Yang, F.; Feng, Y. Exosomes Derived from Hypoxic Colorectal Cancer Cells Transfer Wnt4 to Normoxic Cells to Elicit a Prometastatic Phenotype. Int. J. Biol. Sci. 2018, 14, 2094–2102. [Google Scholar] [CrossRef]
- Taha, E.A.; Ono, K.; Eguchi, T. Roles of Extracellular HSPs as Biomarkers in Immune Surveillance and Immune Evasion. Int. J. Mol. Sci. 2019, 20, 4588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.-Y.; Cheng, J.C.-H.; Chen, Y.-F.; Yang, J.C.-H.; Hsu, F.-M. Circulating Exosomal Integrin Β3 Is Associated with Intracranial Failure and Survival in Lung Cancer Patients Receiving Cranial Irradiation for Brain Metastases: A Prospective Observational Study. Cancers 2021, 13, 380. [Google Scholar] [CrossRef] [PubMed]
- Harada, T.; Yamamoto, H.; Kishida, S.; Kishida, M.; Awada, C.; Takao, T.; Kikuchi, A. Wnt5b-Associated Exosomes Promote Cancer Cell Migration and Proliferation. Cancer Sci. 2017, 108, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Pocock, R. Invited Review: Decoding the MicroRNA Response to Hypoxia. Pflügers Arch.—Eur. J. Physiol. 2011, 461, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Luo, G.; Zhang, K.; Cao, J.; Huang, C.; Jiang, T.; Liu, B.; Su, L.; Qiu, Z. Hypoxic Tumor-Derived Exosomal MiR-301a Mediates M2 Macrophage Polarization via PTEN/PI3Kγ to Promote Pancreatic Cancer Metastasis. Cancer Res. 2018, 78, 4586–4598. [Google Scholar] [CrossRef] [Green Version]
- Kang, W.K.; Lee, J.K.; Oh, S.T.; Lee, S.H.; Jung, C.K. Stromal Expression of MiR-21 in T3-4a Colorectal Cancer Is an Independent Predictor of Early Tumor Relapse. BMC Gastroenterol. 2015, 15, 2. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Feng, X.; Liu, H.; Tong, R.; Wu, J.; Li, C.; Yu, H.; Chen, Y.; Cheng, Q.; Chen, J.; et al. High-Metastatic Cancer Cells Derived Exosomal MiR92a-3p Promotes Epithelial-Mesenchymal Transition and Metastasis of Low-Metastatic Cancer Cells by Regulating PTEN/Akt Pathway in Hepatocellular Carcinoma. Oncogene 2020, 39, 6529–6543. [Google Scholar] [CrossRef]
- Wei, X.; Liu, C.; Wang, H.; Wang, L.; Xiao, F.; Guo, Z.; Zhang, H. Surface Phosphatidylserine Is Responsible for the Internalization on Microvesicles Derived from Hypoxia-Induced Human Bone Marrow Mesenchymal Stem Cells into Human Endothelial Cells. PLoS ONE 2016, 11, e0147360. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.; Zeng, W.; Wu, B.; Wang, L.; Wang, Z.; Tian, H.; Wang, L.; Jiang, Y.; Clay, R.; Wei, X.; et al. PPARα Inhibition Overcomes Tumor-Derived Exosomal Lipid-Induced Dendritic Cell Dysfunction. Cell Rep. 2020, 33, 108278. [Google Scholar] [CrossRef]
- Schlaepfer, I.R.; Nambiar, D.K.; Ramteke, A.; Kumar, R.; Dhar, D.; Agarwal, C.; Bergman, B.; Graner, M.; Maroni, P.; Singh, R.P.; et al. Hypoxia Induces Triglycerides Accumulation in Prostate Cancer Cells and Extracellular Vesicles Supporting Growth and Invasiveness Following Reoxygenation. Oncotarget 2015, 6, 22836–22856. [Google Scholar] [CrossRef] [Green Version]
- Umezu, T.; Tadokoro, H.; Azuma, K.; Yoshizawa, S.; Ohyashiki, K.; Ohyashiki, J.H. Exosomal MiR-135b Shed from Hypoxic Multiple Myeloma Cells Enhances Angiogenesis by Targeting Factor-Inhibiting HIF-1. Blood 2014, 124, 3748–3757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, G.; Hoshino, A.; Kenific, C.M.; Matei, I.R.; Steiner, L.; Freitas, D.; Kim, H.S.; Oxley, P.R.; Scandariato, I.; Casanova-Salas, I.; et al. Tumour Exosomal CEMIP Protein Promotes Cancer Cell Colonization in Brain Metastasis. Nat. Cell Biol. 2019, 21, 1403–1412. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Wang, L.; Dai, T.; Jin, K.; Zhang, Z.; Wang, S.; Xie, F.; Fang, P.; Yang, B.; Huang, H.; et al. Tumor-Derived Exosomes Antagonize Innate Antiviral Immunity. Nat. Immunol. 2018, 19, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, S.; Floros, T.; Theodoraki, M.-N.; Hong, C.-S.; Jackson, E.K.; Lang, S.; Whiteside, T.L. Suppression of Lymphocyte Functions by Plasma Exosomes Correlates with Disease Activity in Patients with Head and Neck Cancer. Clin. Cancer Res. 2017, 23, 4843–4854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Y.; Zheng, X.; Gao, W.; Wang, B.; Wu, Y. Tumor Microenvironment and Immune-Related Therapies of Head and Neck Squamous Cell Carcinoma. Mol. Ther.—Oncolytics 2021, 20, 342–351. [Google Scholar] [CrossRef]
- Kumar, A.; Deep, G. Exosomes in Hypoxia-Induced Remodeling of the Tumor Microenvironment. Cancer Lett. 2020, 488, 1–8. [Google Scholar] [CrossRef]
- Droste, M.; Thakur, B.K.; Eliceiri, B.P. Tumor-Derived Extracellular Vesicles and the Immune System—Lessons From Immune-Competent Mouse-Tumor Models. Front. Immunol. 2020, 11, 606859. [Google Scholar] [CrossRef]
- Qian, M.; Wang, S.; Guo, X.; Wang, J.; Zhang, Z.; Qiu, W.; Gao, X.; Chen, Z.; Xu, J.; Zhao, R.; et al. Hypoxic Glioma-Derived Exosomes Deliver MicroRNA-1246 to Induce M2 Macrophage Polarization by Targeting TERF2IP via the STAT3 and NF-ΚB Pathways. Oncogene 2020, 39, 428–442. [Google Scholar] [CrossRef]
- Yin, Z.; Ma, T.; Huang, B.; Lin, L.; Zhou, Y.; Yan, J.; Zou, Y.; Chen, S. Macrophage-Derived Exosomal MicroRNA-501-3p Promotes Progression of Pancreatic Ductal Adenocarcinoma through the TGFBR3-Mediated TGF-β Signaling Pathway. J. Exp. Clin. Cancer Res. 2019, 38, 310. [Google Scholar] [CrossRef] [Green Version]
- Wen, Z.-F.; Liu, H.; Gao, R.; Zhou, M.; Ma, J.; Zhang, Y.; Zhao, J.; Chen, Y.; Zhang, T.; Huang, F.; et al. Tumor Cell-Released Autophagosomes (TRAPs) Promote Immunosuppression through Induction of M2-like Macrophages with Increased Expression of PD-L1. J. Immunother. Cancer 2018, 6, 151. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Ying, X.; Wang, X.; Wu, X.; Zhu, Q.; Wang, X. Exosomes Derived from Hypoxic Epithelial Ovarian Cancer Deliver MicroRNA-940 to Induce Macrophage M2 Polarization. Oncol. Rep. 2017, 38, 522–528. [Google Scholar] [CrossRef] [Green Version]
- Kuang, D.-M.; Zhao, Q.; Peng, C.; Xu, J.; Zhang, J.-P.; Wu, C.; Zheng, L. Activated Monocytes in Peritumoral Stroma of Hepatocellular Carcinoma Foster Immune Privilege and Disease Progression through PD-L1. J. Exp. Med. 2009, 206, 1327–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gärtner, K.; Battke, C.; Dünzkofer, J.; Hüls, C.; von Neubeck, B.; Kellner, M.-K.; Fiestas, E.; Fackler, S.; Lang, S.; Zeidler, R. Tumor-Derived Extracellular Vesicles Activate Primary Monocytes. Cancer Med. 2018, 7, 2013–2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popēna, I.; Ābols, A.; Saulīte, L.; Pleiko, K.; Zandberga, E.; Jēkabsons, K.; Endzeliņš, E.; Llorente, A.; Linē, A.; Riekstiņa, U. Effect of Colorectal Cancer-Derived Extracellular Vesicles on the Immunophenotype and Cytokine Secretion Profile of Monocytes and Macrophages. Cell Commun. Signal. 2018, 16, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, L.; He, Y.; Peng, F.; Yang, J.; Yuan, C. Endometrial Cancer Cells Promote M2-Like Macrophage Polarization by Delivering Exosomal MiRNA-21 under Hypoxia Condition. J. Immunol. Res. 2020, 2020, 9731049. [Google Scholar] [CrossRef] [PubMed]
- Gaggero, S.; Witt, K.; Carlsten, M.; Mitra, S. Cytokines Orchestrating the Natural Killer-Myeloid Cell Crosstalk in the Tumor Microenvironment: Implications for Natural Killer Cell-Based Cancer Immunotherapy. Front. Immunol. 2021, 11, 621225. [Google Scholar] [CrossRef] [PubMed]
- Berchem, G.; Noman, M.Z.; Bosseler, M.; Paggetti, J.; Baconnais, S.; Le cam, E.; Nanbakhsh, A.; Moussay, E.; Mami-Chouaib, F.; Janji, B.; et al. Hypoxic Tumor-Derived Microvesicles Negatively Regulate NK Cell Function by a Mechanism Involving TGF-β and MiR23a Transfer. Oncoimmunology 2016, 5, e1062968. [Google Scholar] [CrossRef]
- Melaiu, O.; Lucarini, V.; Cifaldi, L.; Fruci, D. Influence of the Tumor Microenvironment on NK Cell Function in Solid Tumors. Front. Immunol. 2020, 10, 3038. [Google Scholar] [CrossRef]
- Romero, A.I.; Thorén, F.B.; Brune, M.; Hellstrand, K. NKp46 and NKG2D Receptor Expression in NK Cells with CD56dim and CD56bright Phenotype: Regulation by Histamine and Reactive Oxygen Species. Br. J. Haematol. 2006, 132, 91–98. [Google Scholar] [CrossRef]
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, Inflammation, and Cancer. Cell 2010, 140, 883–899. [Google Scholar] [CrossRef] [Green Version]
- Gornostaeva, A.N.; Bobyleva, P.I.; Andreeva, E.R.; Yakubets, D.A.; Buravkova, L.B. Adipose-Derived Stromal Cell Immunosuppression of T Cells Is Enhanced under “Physiological” Hypoxia. Tissue Cell 2020, 63, 101320. [Google Scholar] [CrossRef]
- Jayaprakash, P.; Ai, M.; Liu, A.; Budhani, P.; Bartkowiak, T.; Sheng, J.; Ager, C.; Nicholas, C.; Jaiswal, A.R.; Sun, Y.; et al. Targeted Hypoxia Reduction Restores T Cell Infiltration and Sensitizes Prostate Cancer to Immunotherapy. J. Clin. Investig. 2018, 128, 5137–5149. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Xia, Y.; Lin, Z.; Qu, Y.; Qi, Y.; Chen, Y.; Zhou, Q.; Zeng, H.; Wang, J.; Chang, Y.; et al. Tumor-Infiltrating CD39+CD8+ T Cells Determine Poor Prognosis and Immune Evasion in Clear Cell Renal Cell Carcinoma Patients. Cancer Immunol. Immunother. 2020, 69, 1565–1576. [Google Scholar] [CrossRef] [PubMed]
- Lo Presti, E.; Pizzolato, G.; Corsale, A.M.; Caccamo, N.; Sireci, G.; Dieli, F.; Meraviglia, S. Γδ T Cells and Tumor Microenvironment: From Immunosurveillance to Tumor Evasion. Front. Immunol. 2018, 9, 1395. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Cao, B.; Liang, X.; Lu, S.; Luo, H.; Wang, Z.; Wang, S.; Jiang, J.; Lang, J.; Zhu, G. Microenvironmental Oxygen Pressure Orchestrates an Anti- and pro-Tumoral Γδ T Cell Equilibrium via Tumor-Derived Exosomes. Oncogene 2019, 38, 2830–2843. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Gu, Y.; Cao, X. The Exosomes in Tumor Immunity. Oncoimmunology 2015, 4, e1027472. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Wang, J.; Shao, C.; Liu, S.; Yu, Y.; Wang, Q.; Cao, X. Efficient Induction of Antitumor T Cell Immunity by Exosomes Derived from Heat-Shocked Lymphoma Cells. Eur. J. Immunol. 2006, 36, 1598–1607. [Google Scholar] [CrossRef]
- Wu, P.; Wu, D.; Ni, C.; Ye, J.; Chen, W.; Hu, G.; Wang, Z.; Wang, C.; Zhang, Z.; Xia, W.; et al. ΓδT17 Cells Promote the Accumulation and Expansion of Myeloid-Derived Suppressor Cells in Human Colorectal Cancer. Immunity 2014, 40, 785–800. [Google Scholar] [CrossRef] [Green Version]
- Daley, D.; Zambirinis, C.P.; Seifert, L.; Akkad, N.; Mohan, N.; Werba, G.; Barilla, R.; Torres-Hernandez, A.; Hundeyin, M.; Mani, V.R.K.; et al. Γδ T Cells Support Pancreatic Oncogenesis by Restraining Aβ T Cell Activation. Cell 2016, 166, 1485–1499.e15. [Google Scholar] [CrossRef] [Green Version]
- Park, J.H.; Kim, H.-J.; Kim, C.W.; Kim, H.C.; Jung, Y.; Lee, H.-S.; Lee, Y.; Ju, Y.S.; Oh, J.E.; Park, S.-H.; et al. Tumor Hypoxia Represses Γδ T Cell-Mediated Antitumor Immunity against Brain Tumors. Nat. Immunol. 2021, 22, 336–346. [Google Scholar] [CrossRef]
- Sureshbabu, S.K.; Chaukar, D.; Chiplunkar, S.V. Hypoxia Regulates the Differentiation and Anti-tumor Effector Functions of ΓδT Cells in Oral Cancer. Clin. Exp. Immunol. 2020, 201, 40–57. [Google Scholar] [CrossRef] [PubMed]
- Groth, C.; Hu, X.; Weber, R.; Fleming, V.; Altevogt, P.; Utikal, J.; Umansky, V. Immunosuppression Mediated by Myeloid-Derived Suppressor Cells (MDSCs) during Tumour Progression. Br. J. Cancer 2019, 120, 16–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, V.; Patel, S.; Tcyganov, E.; Gabrilovich, D.I. The Nature of Myeloid-Derived Suppressor Cells in the Tumor Microenvironment. Trends Immunol. 2016, 37, 208–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabrilovich, D.I. Myeloid-Derived Suppressor Cells. Cancer Immunol. Res. 2017, 5, 3–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Qiu, W.; Wang, J.; Liu, Q.; Qian, M.; Wang, S.; Zhang, Z.; Gao, X.; Chen, Z.; Guo, Q.; et al. Glioma Exosomes Mediate the Expansion and Function of Myeloid-derived Suppressor Cells through MicroRNA-29a/ Hbp1 and MicroRNA-92a/ Prkar1a Pathways. Int. J. Cancer 2019, 144, 3111–3126. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yin, K.; Tian, J.; Xia, X.; Ma, J.; Tang, X.; Xu, H.; Wang, S. Granulocytic Myeloid-Derived Suppressor Cells Promote the Stemness of Colorectal Cancer Cells through Exosomal S100A9. Adv. Sci. 2019, 6, 1901278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, M.; Wu, R.; Chen, L.; Peng, Q.; Li, S.; Zhang, Y.; Zhou, L.; Duan, L. S100A9 Regulates MDSCs-Mediated Immune Suppression via the RAGE and TLR4 Signaling Pathways in Colorectal Carcinoma. Front. Immunol. 2019, 10, 2243. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Li, X.; Chen, H.; Cao, Y.; Xiao, Q.; He, Y.; Wei, J.; Zhou, J. IRF7 Regulates the Development of Granulocytic Myeloid-Derived Suppressor Cells through S100A9 Transrepression in Cancer. Oncogene 2017, 36, 2969–2980. [Google Scholar] [CrossRef]
- Wu, X.; Zhu, D.; Tian, J.; Tang, X.; Guo, H.; Ma, J.; Xu, H.; Wang, S. Granulocytic Myeloid-Derived Suppressor Cell Exosomal Prostaglandin E2 Ameliorates Collagen-Induced Arthritis by Enhancing IL-10+ B Cells. Front. Immunol. 2020, 11, 588500. [Google Scholar] [CrossRef]
- Dai, H.; Thomson, A.W.; Rogers, N.M. Dendritic Cells as Sensors, Mediators, and Regulators of Ischemic Injury. Front. Immunol. 2019, 10, 2418. [Google Scholar] [CrossRef] [Green Version]
- Winning, S.; Fandrey, J. Dendritic Cells under Hypoxia: How Oxygen Shortage Affects the Linkage between Innate and Adaptive Immunity. J. Immunol. Res. 2016, 2016, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruez, R.; Dubrot, J.; Zoso, A.; Bacchetta, M.; Molica, F.; Hugues, S.; Kwak, B.R.; Chanson, M. Dendritic Cell Migration Toward CCL21 Gradient Requires Functional Cx43. Front. Physiol. 2018, 9, 288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Y.-R.; Zhang, T.; Jia, P.; Xu, X.-L.; Fang, Y.; Ding, X.-Q. Interaction between Bone Marrow-Derived Dendritic Cells and MiR-21 of Tubular Renal Epithelial Cells under Hypoxia. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 1641–1651. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Liu, Y.; Zhou, X. Immune Modulation by Silencing CD80 and CD86 Production in Dendritic Cells Using Small Hairpin RNA to Reduce Heart Transplant Rejection. Transpl. Immunol. 2018, 49, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, B.; Jia, X.; Ma, Y.; Gu, Y.; Zhang, P.; Wei, Q.; Cai, J.; Cui, J.; Gao, F.; et al. Radiation-Induced Decrease of CD8 + Dendritic Cells Contributes to Th1/Th2 Shift. Int. Immunopharmacol. 2017, 46, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Jeong, Y.; Ashraf, M.U.; Bae, Y.-S. Dendritic Cell-Mediated Th2 Immunity and Immune Disorders. Int. J. Mol. Sci. 2019, 20, 2159. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.; Rao, Q.; Zhang, C.; Zhang, X.; Qin, Y.; Niu, Z. Dendritic Cells Pulsed with Exosomes in Combination with PD-1 Antibody Increase the Efficacy of Sorafenib in Hepatocellular Carcinoma Model. Transl. Oncol. 2018, 11, 250–258. [Google Scholar] [CrossRef]
- Weigert, A.; Weichand, B.; Sekar, D.; Sha, W.; Hahn, C.; Mora, J.; Ley, S.; Essler, S.; Dehne, N.; Brüne, B. HIF-1α Is a Negative Regulator of Plasmacytoid DC Development in Vitro and in Vivo. Blood 2012, 120, 3001–3006. [Google Scholar] [CrossRef] [Green Version]
- Benson, M.J.; Pino-Lagos, K.; Rosemblatt, M.; Noelle, R.J. All-Trans Retinoic Acid Mediates Enhanced T Reg Cell Growth, Differentiation, and Gut Homing in the Face of High Levels of Co-Stimulation. J. Exp. Med. 2007, 204, 1765–1774. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Wen, Z.; Cheng, F.; Ma, J.; Li, W.; Ren, H.; Sheng, Y.; Dong, H.; Lu, L.; Hu, H.-M.; et al. Tumor-Released Autophagosomes Induce IL-10-Producing B Cells with Suppressive Activity on T Lymphocytes via TLR2-MyD88-NF-ΚB Signal Pathway. Oncoimmunology 2016, 5, e1180485. [Google Scholar] [CrossRef]
- Zhang, Y.; Pan, N.; Sheng, Y.; Zhou, M.; Wen, Z.; Chen, Y.; Huang, F.; Wang, L.-X. Hypoxia Enhances IL-10-Producing B Cell Generation through Upregulating High-Mobility Group B1 on Tumor Cell-Released Autophagosomes. Immunol. Lett. 2019, 216, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Lee-Chang, C.; Rashidi, A.; Miska, J.; Zhang, P.; Pituch, K.C.; Hou, D.; Xiao, T.; Fischietti, M.; Kang, S.J.; Appin, C.L.; et al. Myeloid-Derived Suppressive Cells Promote B Cell–Mediated Immunosuppression via Transfer of PD-L1 in Glioblastoma. Cancer Immunol. Res. 2019, 7, 1928–1943. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-Q.; Li, P.-C.; Pan, N.; Gao, R.; Wen, Z.-F.; Zhang, T.-Y.; Huang, F.; Wu, F.-Y.; Ou, X.-L.; Zhang, J.-P.; et al. Tumor-Released Autophagosomes Induces CD4+ T Cell-Mediated Immunosuppression via a TLR2–IL-6 Cascade. J. Immunother. Cancer 2019, 7, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jachetti, E.; Cancila, V.; Rigoni, A.; Bongiovanni, L.; Cappetti, B.; Belmonte, B.; Enriquez, C.; Casalini, P.; Ostano, P.; Frossi, B.; et al. Cross-Talk between Myeloid-Derived Suppressor Cells and Mast Cells Mediates Tumor-Specific Immunosuppression in Prostate Cancer. Cancer Immunol. Res. 2018, 6, 552–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Y.; Liu, L.; Xia, Y.; Qi, Y.; Chen, Y.; Chen, L.; Zhang, P.; Kong, Y.; Qu, Y.; Wang, Z.; et al. Tumor Infiltrating Mast Cells Determine Oncogenic HIF-2α-Conferred Immune Evasion in Clear Cell Renal Cell Carcinoma. Cancer Immunol. Immunother. 2019, 68, 731–741. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.-J.; Oh, H.-A.; Nam, S.-Y.; Han, N.-R.; Kim, Y.-S.; Kim, J.-H.; Lee, S.-J.; Kim, M.-H.; Moon, P.-D.; Kim, H.-M.; et al. The Critical Role of Mast Cell-Derived Hypoxia-Inducible Factor-1α in Human and Mice Melanoma Growth. Int. J. Cancer 2013, 132, 2492–2501. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Moreno, I.G.; Ibarra-Sánchez, A.; Castillo-Arellano, J.I.; Blank, U.; González-Espinosa, C. Mast Cells Localize in Hypoxic Zones of Tumors and Secrete CCL-2 under Hypoxia through Activation of L-Type Calcium Channels. J. Immunol. 2020, 204, 1056–1068. [Google Scholar] [CrossRef]
- Liu, T.; Xia, Q.; Zhang, H.; Wang, Z.; Yang, W.; Gu, X.; Hou, T.; Chen, Y.; Pei, X.; Zhu, G.; et al. CCL5-Dependent Mast Cell Infiltration into the Tumor Microenvironment in Clear Cell Renal Cell Carcinoma Patients. Aging 2020, 12, 21809–21836. [Google Scholar] [CrossRef]
- Ammendola, M.; Currò, G.; Laface, C.; Zuccalà, V.; Memeo, R.; Luposella, F.; Laforgia, M.; Zizzo, N.; Zito, A.; Loisi, D.; et al. Mast Cells Positive for C-Kit Receptor and Tryptase Correlate with Angiogenesis in Cancerous and Adjacent Normal Pancreatic Tissue. Cells 2021, 10, 444. [Google Scholar] [CrossRef]
- Mrizak, D.; Martin, N.; Barjon, C.; Jimenez-Pailhes, A.-S.; Mustapha, R.; Niki, T.; Guigay, J.; Pancré, V.; de Launoit, Y.; Busson, P.; et al. Effect of Nasopharyngeal Carcinoma-Derived Exosomes on Human Regulatory T Cells. JNCI J. Natl. Cancer Inst. 2015, 107, 363. [Google Scholar] [CrossRef] [Green Version]
- Fu, Q.; Xu, L.; Wang, Y.; Jiang, Q.; Liu, Z.; Zhang, J.; Zhou, Q.; Zeng, H.; Tong, S.; Wang, T.; et al. Tumor-Associated Macrophage-Derived Interleukin-23 Interlinks Kidney Cancer Glutamine Addiction with Immune Evasion. Eur. Urol. 2019, 75, 752–763. [Google Scholar] [CrossRef] [PubMed]
- Huo, S.; Luo, Y.; Deng, R.; Liu, X.; Wang, J.; Wang, L.; Zhang, B.; Wang, F.; Lu, J.; Li, X. EBV-EBNA1 Constructs an Immunosuppressive Microenvironment for Nasopharyngeal Carcinoma by Promoting the Chemoattraction of Treg Cells. J. Immunother. Cancer 2020, 8, e001588. [Google Scholar] [CrossRef] [PubMed]
- Doedens, A.L.; Stockmann, C.; Rubinstein, M.P.; Liao, D.; Zhang, N.; DeNardo, D.G.; Coussens, L.M.; Karin, M.; Goldrath, A.W.; Johnson, R.S. Macrophage Expression of Hypoxia-Inducible Factor-1α Suppresses T-Cell Function and Promotes Tumor Progression. Cancer Res. 2010, 70, 7465–7475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Zhou, J.; Li, X.; Wang, X.; Lin, Y.; Wang, X. Exosomes Derived from Hypoxic Epithelial Ovarian Cancer Cells Deliver MicroRNAs to Macrophages and Elicit a Tumor-Promoted Phenotype. Cancer Lett. 2018, 435, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Boone, B.A.; Lotze, M.T. Targeting Damage-Associated Molecular Pattern Molecules (DAMPs) and DAMP Receptors in Melanoma. In Molecular Diagnostics for Melanoma; Methods in Molecular Biology; Springer: Berlin, Germany, 2014; Volume 1102, pp. 537–552. [Google Scholar] [CrossRef]
- Álvarez, K.; Vasquez, G. Damage-Associated Molecular Patterns and Their Role as Initiators of Inflammatory and Auto-Immune Signals in Systemic Lupus Erythematosus. Int. Rev. Immunol. 2017, 36, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yan, W.; Tohme, S.; Chen, M.; Fu, Y.; Tian, D.; Lotze, M.; Tang, D.; Tsung, A. Hypoxia Induced HMGB1 and Mitochondrial DNA Interactions Mediate Tumor Growth in Hepatocellular Carcinoma through Toll-like Receptor 9. J. Hepatol. 2015, 63, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Huang, Y.; Huang, Y.; Fu, Y.; Tang, D.; Kang, R.; Zhou, R.; Fan, X.-G. The Long Non-Coding RNA TP73-AS1 Modulates HCC Cell Proliferation through MiR-200a-Dependent HMGB1/RAGE Regulation. J. Exp. Clin. Cancer Res. 2017, 36, 51. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Huang, W.; Luo, R. The MicroRNA-325 Inhibits Hepatocellular Carcinoma Progression by Targeting High Mobility Group Box 1. Diagn. Pathol. 2015, 10, 117. [Google Scholar] [CrossRef] [Green Version]
- Liu, A.-H.; Wu, Y.-T.; Wang, Y.-P. MicroRNA-129-5p Inhibits the Development of Autoimmune Encephalomyelitis-Related Epilepsy by Targeting HMGB1 through the TLR4/NF-KB Signaling Pathway. Brain Res. Bull. 2017, 132, 139–149. [Google Scholar] [CrossRef]
- Zhong, B.; Qin, Z.; Zhou, H.; Yang, F.; Wei, K.; Jiang, X.; Jia, R. MicroRNA-505 Negatively Regulates HMGB1 to Suppress Cell Proliferation in Renal Cell Carcinoma. J. Cell. Physiol. 2019, 234, 15025–15034. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.; Wang, B.; Zhang, C. MicroRNA-505-3p Inhibits Development of Glioma by Targeting HMGB1 and Regulating AKT Expression. Oncol. Lett. 2020, 20, 1663–1670. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Ying, S.; Cai, X. MicroRNA-Mediated Regulation of HMGB1 in Human Hepatocellular Carcinoma. Biomed Res. Int. 2018, 2018, 2754941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pistoia, V.; Pezzolo, A. Involvement of HMGB1 in Resistance to Tumor Vessel-Targeted, Monoclonal Antibody-Based Immunotherapy. J. Immunol. Res. 2016, 2016, 3142365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Yang, M.; Kang, R.; Wang, Z.; Zhao, Y.; Yu, Y.; Xie, M.; Yin, X.; Livesey, K.M.; Lotze, M.T.; et al. HMGB1-Induced Autophagy Promotes Chemotherapy Resistance in Leukemia Cells. Leukemia 2011, 25, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Vanichapol, T.; Chiangjong, W.; Panachan, J.; Anurathapan, U.; Chutipongtanate, S.; Hongeng, S. Secretory High-Mobility Group Box 1 Protein Affects Regulatory T Cell Differentiation in Neuroblastoma Microenvironment In Vitro. J. Oncol. 2018, 2018, 7946021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, J.; Ni, A. Landscape of PD-1/PD-L1 Regulation and Targeted Immunotherapy. Chin. Med. Sci. J. 2018, 33, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Kuai, W.; Xu, X.; Yan, J.; Zhao, W.; Li, Y.; Wang, B.; Yuan, N.; Li, Z.; Jia, Y. Prognostic Impact of PD-1 and Tim-3 Expression in Tumor Tissue in Stage I-III Colorectal Cancer. Biomed Res. Int. 2020, 2020, 5294043. [Google Scholar] [CrossRef] [PubMed]
- Ayala-Mar, S.; Donoso-Quezada, J.; González-Valdez, J. Clinical Implications of Exosomal PD-L1 in Cancer Immunotherapy. J. Immunol. Res. 2021, 2021, 8839978. [Google Scholar] [CrossRef]
- Poggio, M.; Hu, T.; Pai, C.-C.; Chu, B.; Belair, C.D.; Chang, A.; Montabana, E.; Lang, U.E.; Fu, Q.; Fong, L.; et al. Suppression of Exosomal PD-L1 Induces Systemic Anti-Tumor Immunity and Memory. Cell 2019, 177, 414–427.e13. [Google Scholar] [CrossRef] [Green Version]
- Yun, S.; Yun, C.W.; Lee, J.H.; Kim, S.; Lee, S.H. Cripto Enhances Proliferation and Survival of Mesenchymal Stem Cells by Up-Regulating JAK2/STAT3 Pathway in a GRP78-Dependent Manner. Biomol. Ther. 2018, 26, 464–473. [Google Scholar] [CrossRef]
- Gordon, S.R.; Maute, R.L.; Dulken, B.W.; Hutter, G.; George, B.M.; McCracken, M.N.; Gupta, R.; Tsai, J.M.; Sinha, R.; Corey, D.; et al. PD-1 Expression by Tumour-Associated Macrophages Inhibits Phagocytosis and Tumour Immunity. Nature 2017, 545, 495–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodman, A.; Patel, S.P.; Kurzrock, R. PD-1–PD-L1 Immune-Checkpoint Blockade in B-Cell Lymphomas. Nat. Rev. Clin. Oncol. 2017, 14, 203–220. [Google Scholar] [CrossRef] [PubMed]
- Pollari, M.; Brück, O.; Pellinen, T.; Vähämurto, P.; Karjalainen-Lindsberg, M.-L.; Mannisto, S.; Kallioniemi, O.; Kellokumpu-Lehtinen, P.-L.; Mustjoki, S.; Leivonen, S.-K.; et al. PD-L1+ Tumor-Associated Macrophages and PD-1+ Tumor-Infiltrating Lymphocytes Predict Survival in Primary Testicular Lymphoma. Haematologica 2018, 103, 1908–1914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, M.; Wang, J.; Yu, W.; Zhang, C.; Liu, M.; Wang, K.; Yang, L.; Wei, F.; Wang, S.E.; Sun, Q.; et al. A Novel MDSC-Induced PD-1−PD-L1+ B-Cell Subset in Breast Tumor Microenvironment Possesses Immuno-Suppressive Properties. Oncoimmunology 2018, 7, e1413520. [Google Scholar] [CrossRef] [Green Version]
- Rühle, A.; Grosu, A.-L.; Wiedenmann, N.; Mix, M.; Stoian, R.; Niedermann, G.; Baltas, D.; Werner, M.; Weber, W.A.; Kayser, G.; et al. Hypoxia Dynamics on FMISO-PET in Combination with PD-1/PD-L1 Expression Has an Impact on the Clinical Outcome of Patients with Head-and-Neck Squamous Cell Carcinoma Undergoing Chemoradiation. Theranostics 2020, 10, 9395–9406. [Google Scholar] [CrossRef]
- Li, J.; Wang, L.; Chen, X.; Li, L.; Li, Y.; Ping, Y.; Huang, L.; Yue, D.; Zhang, Z.; Wang, F.; et al. CD39/CD73 Upregulation on Myeloid-Derived Suppressor Cells via TGF-β-MTOR-HIF-1 Signaling in Patients with Non-Small Cell Lung Cancer. Oncoimmunology 2017, 6, e1320011. [Google Scholar] [CrossRef]
- Giatromanolaki, A.; Kouroupi, M.; Pouliliou, S.; Mitrakas, A.; Hasan, F.; Pappa, A.; Koukourakis, M.I. Ectonucleotidase CD73 and CD39 Expression in Non-Small Cell Lung Cancer Relates to Hypoxia and Immunosuppressive Pathways. Life Sci. 2020, 259, 118389. [Google Scholar] [CrossRef]
- Häusler, S.F.M.; Montalbán del Barrio, I.; Strohschein, J.; Anoop Chandran, P.; Engel, J.B.; Hönig, A.; Ossadnik, M.; Horn, E.; Fischer, B.; Krockenberger, M.; et al. Ectonucleotidases CD39 and CD73 on OvCA Cells Are Potent Adenosine-Generating Enzymes Responsible for Adenosine Receptor 2A-Dependent Suppression of T Cell Function and NK Cell Cytotoxicity. Cancer Immunol. Immunother. 2011, 60, 1405–1418. [Google Scholar] [CrossRef]
- Schuler, P.J.; Saze, Z.; Hong, C.-S.; Muller, L.; Gillespie, D.G.; Cheng, D.; Harasymczuk, M.; Mandapathil, M.; Lang, S.; Jackson, E.K.; et al. Human CD4+CD39+ Regulatory T Cells Produce Adenosine upon Co-Expression of Surface CD73 or Contact with CD73+ Exosomes or CD73+ Cells. Clin. Exp. Immunol. 2014, 177, 531–543. [Google Scholar] [CrossRef]
- Zhang, F.; Li, R.; Yang, Y.; Shi, C.; Shen, Y.; Lu, C.; Chen, Y.; Zhou, W.; Lin, A.; Yu, L.; et al. Specific Decrease in B-Cell-Derived Extracellular Vesicles Enhances Post-Chemotherapeutic CD8+ T Cell Responses. Immunity 2019, 50, 738–750.e7. [Google Scholar] [CrossRef] [Green Version]
- Piedra-Quintero, Z.L.; Wilson, Z.; Nava, P.; Guerau-de-Arellano, M. CD38: An Immunomodulatory Molecule in Inflammation and Autoimmunity. Front. Immunol. 2020, 11, 597959. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Long, Y.; Xiao, S.; Liang, L.; He, Z.; Yue, C.; Wei, X.; Zhou, Y. CD38 Affects the Biological Behavior and Energy Metabolism of Nasopharyngeal Carcinoma Cells. Int. J. Oncol. 2018, 54, 585–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zumaquero, E.; Muñoz, P.; Cobo, M.; Lucena, G.; Pavón, E.J.; Martín, A.; Navarro, P.; García-Pérez, A.; Ariza-Veguillas, A.; Malavasi, F.; et al. Exosomes from Human Lymphoblastoid B Cells Express Enzymatically Active CD38 That Is Associated with Signaling Complexes Containing CD81, Hsc-70 and Lyn. Exp. Cell Res. 2010, 316, 2692–2706. [Google Scholar] [CrossRef] [PubMed]
- Angelicola, S.; Ruzzi, F.; Landuzzi, L.; Scalambra, L.; Gelsomino, F.; Ardizzoni, A.; Nanni, P.; Lollini, P.-L.; Palladini, A. IFN-γ and CD38 in Hyperprogressive Cancer Development. Cancers 2021, 13, 309. [Google Scholar] [CrossRef]
- Wo, Y.J.; Gan, A.S.P.; Lim, X.; Tay, I.S.Y.; Lim, S.; Lim, J.C.T.; Yeong, J.P.S. The Roles of CD38 and CD157 in the Solid Tumor Microenvironment and Cancer Immunotherapy. Cells 2019, 9, 26. [Google Scholar] [CrossRef] [Green Version]
- Matlung, H.L.; Szilagyi, K.; Barclay, N.A.; van den Berg, T.K. The CD47-SIRPα Signaling Axis as an Innate Immune Checkpoint in Cancer. Immunol. Rev. 2017, 276, 145–164. [Google Scholar] [CrossRef]
- Samanta, D.; Park, Y.; Ni, X.; Li, H.; Zahnow, C.A.; Gabrielson, E.; Pan, F.; Semenza, G.L. Chemotherapy Induces Enrichment of CD47+/CD73+/PDL1+ Immune Evasive Triple-Negative Breast Cancer Cells. Proc. Natl. Acad. Sci. USA 2018, 115, E1239–E1248. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.-T.; Bu, L.-L.; Huang, C.-F.; Zhang, W.-F.; Chen, W.-J.; Gutkind, J.S.; Kulkarni, A.B.; Sun, Z.-J. PD-1 Blockade Attenuates Immunosuppressive Myeloid Cells Due to Inhibition of CD47/SIRPα Axis in HPV Negative Head and Neck Squamous Cell Carcinoma. Oncotarget 2015, 6, 42067–42080. [Google Scholar] [CrossRef] [Green Version]
- Kaur, S.; Elkahloun, A.G.; Singh, S.P.; Chen, Q.-R.; Meerzaman, D.M.; Song, T.; Manu, N.; Wu, W.; Mannan, P.; Garfield, S.H.; et al. A Function-Blocking CD47 Antibody Suppresses Stem Cell and EGF Signaling in Triple-Negative Breast Cancer. Oncotarget 2016, 7, 10133–10152. [Google Scholar] [CrossRef] [Green Version]
- Ye, L.; Zhang, Q.; Cheng, Y.; Chen, X.; Wang, G.; Shi, M.; Zhang, T.; Cao, Y.; Pan, H.; Zhang, L.; et al. Tumor-Derived Exosomal HMGB1 Fosters Hepatocellular Carcinoma Immune Evasion by Promoting TIM-1+ Regulatory B Cell Expansion. J. Immunother. Cancer 2018, 6, 145. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Bi, J.; Zheng, X.; Chen, Y.; Wang, H.; Wu, W.; Wang, Z.; Wu, Q.; Peng, H.; Wei, H.; et al. Blockade of the Checkpoint Receptor TIGIT Prevents NK Cell Exhaustion and Elicits Potent Anti-Tumor Immunity. Nat. Immunol. 2018, 19, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, T.; Seow, S.V.; Wong, D.; Robinson, M.; Campana, D. Blocking Expression of Inhibitory Receptor NKG2A Overcomes Tumor Resistance to NK Cells. J. Clin. Investig. 2019, 129, 2094–2106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- André, P.; Denis, C.; Soulas, C.; Bourbon-Caillet, C.; Lopez, J.; Arnoux, T.; Bléry, M.; Bonnafous, C.; Gauthier, L.; Morel, A.; et al. Anti-NKG2A MAb Is a Checkpoint Inhibitor That Promotes Anti-Tumor Immunity by Unleashing Both T and NK Cells. Cell 2018, 175, 1731–1743.e13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bojović, K.; Ignjatović, Ð.; Soković Bajić, S.; Vojnović Milutinović, D.; Tomić, M.; Golić, N.; Tolinački, M. Gut Microbiota Dysbiosis Associated With Altered Production of Short Chain Fatty Acids in Children With Neurodevelopmental Disorders. Front. Cell. Infect. Microbiol. 2020, 10, 223. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Pang, Y.; Moses, H.L. TGF-β and Immune Cells: An Important Regulatory Axis in the Tumor Microenvironment and Progression. Trends Immunol. 2010, 31, 220–227. [Google Scholar] [CrossRef] [Green Version]
- van den Bulk, J.; de Miranda, N.F.C.C.; ten Dijke, P. Therapeutic Targeting of TGF-β in Cancer: Hacking a Master Switch of Immune Suppression. Clin. Sci. 2021, 135, 35–52. [Google Scholar] [CrossRef]
- Armitage, J.D.; Newnes, H.V.; McDonnell, A.; Bosco, A.; Waithman, J. Fine-Tuning the Tumour Microenvironment: Current Perspectives on the Mechanisms of Tumour Immunosuppression. Cells 2021, 10, 56. [Google Scholar] [CrossRef]
- Ma, L.; Shan, W.; Ding, X.; Yang, P.; Rozjan, A.; Yao, Q. Intermittent Hypoxia Induces Tumor Immune Escape in Murine S180 Solid Tumors via the Upregulation of TGF-Β1 in Mice. Sleep Breath. 2021, 25, 719–726. [Google Scholar] [CrossRef]
- Liu, M.; Kuo, F.; Capistrano, K.J.; Kang, D.; Nixon, B.G.; Shi, W.; Chou, C.; Do, M.H.; Stamatiades, E.G.; Gao, S.; et al. TGF-β Suppresses Type 2 Immunity to Cancer. Nature 2020, 587, 115–120. [Google Scholar] [CrossRef]
- Syn, N.; Wang, L.; Sethi, G.; Thiery, J.-P.; Goh, B.-C. Exosome-Mediated Metastasis: From Epithelial–Mesenchymal Transition to Escape from Immunosurveillance. Trends Pharmacol. Sci. 2016, 37, 606–617. [Google Scholar] [CrossRef]
- Lu, Q.; Wang, X.; Zhu, J.; Fei, X.; Chen, H.; Li, C. Hypoxic Tumor-Derived Exosomal Circ0048117 Facilitates M2 Macrophage Polarization Acting as MiR-140 Sponge in Esophageal Squamous Cell Carcinoma. OncoTargets Ther. 2020, 13, 11883–11897. [Google Scholar] [CrossRef] [PubMed]
- Noman, M.Z.; Desantis, G.; Janji, B.; Hasmim, M.; Karray, S.; Dessen, P.; Bronte, V.; Chouaib, S. PD-L1 Is a Novel Direct Target of HIF-1α, and Its Blockade under Hypoxia Enhanced MDSC-Mediated T Cell Activation. J. Exp. Med. 2014, 211, 781–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, J.; Sarkar, D.; Pramanik, A.; Mahanti, K.; Adhikary, A.; Bhattacharyya, S. PGE2-HIF1α Reciprocal Induction Regulates Migration, Phenotypic Alteration and Immunosuppressive Capacity of Macrophages in Tumor Microenvironment. Life Sci. 2020, 253, 117731. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.-F.; Liu, T.-Q.; Zhi, X.-T.; Zou, J.; Zhong, J.-T.; Li, T.; Mo, X.-L.; Zhou, W.; Guo, W.-W.; Liu, X.; et al. COX-2/PGE2 Axis Regulates HIF2α Activity to Promote Hepatocellular Carcinoma Hypoxic Response and Reduce the Sensitivity of Sorafenib Treatment. Clin. Cancer Res. 2018, 24, 3204–3216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linton, S.S.; Abraham, T.; Liao, J.; Clawson, G.A.; Butler, P.J.; Fox, T.; Kester, M.; Matters, G.L. Tumor-Promoting Effects of Pancreatic Cancer Cell Exosomes on THP-1-Derived Macrophages. PLoS ONE 2018, 13, e0206759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choueiri, T.K.; Kaelin, W.G. Targeting the HIF2–VEGF Axis in Renal Cell Carcinoma. Nat. Med. 2020, 26, 1519–1530. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D. Tumor Refractoriness to Anti-VEGF Therapy. Oncotarget 2016, 7, 46668–46677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Almeida, P.E.; Mak, J.; Hernandez, G.; Jesudason, R.; Herault, A.; Javinal, V.; Borneo, J.; Kim, J.M.; Walsh, K.B. Anti-VEGF Treatment Enhances CD8+ T-Cell Antitumor Activity by Amplifying Hypoxia. Cancer Immunol. Res. 2020, 8, 806–818. [Google Scholar] [CrossRef] [Green Version]
- Eguchi, T.; Sogawa, C.; Ono, K.; Matsumoto, M.; Tran, M.T.; Okusha, Y.; Lang, B.J.; Okamoto, K.; Calderwood, S.K. Cell Stress Induced Stressome Release Including Damaged Membrane Vesicles and Extracellular HSP90 by Prostate Cancer Cells. Cells 2020, 9, 755. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.-G.; Wang, C.; Liu, D.-Y.; Zhang, X.; Wang, L.; Yan, M.; Zhang, W.; Zhu, J.; Li, Z.-C.; Mi, C.; et al. Hypoxia Upregulates HIG2 Expression and Contributes to Bevacizumab Resistance in Glioblastoma. Oncotarget 2016, 7, 47808–47820. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exosomal Component | Tumor Type/Model | Effects | Reference |
|---|---|---|---|
| Exosomal RNAs | |||
| lncRNA-UCA1 | Bladder cancer | Promotes tumor growth and progression through EMT in vitro and in vivo lncRNA-UCA1 in the human serum serves as a possible biomarker | [18] |
| circ-133 | Colorectal cancer | Promotes cancer metastasis by acting on the miR-133a/GEF-H1/RhoA axis | [36] |
| miR-128-3p, miR-140-3p, miR-340-5p, miR-452-5p, miR-769-5p and miR-1304-p5, miR-340-5p | Esophageal squamous carcinoma | Upregulated expression of these exosomal miRNAs in hypoxic esophageal squamous carcinoma cells | [44] |
| lncRNA-UCA1 | Ladder cancer | Promotes cell proliferation, migration, and invasion | [18] |
| miR-301a | Glioma | Activates Wnt/β-catenin signaling and promotes radiation resistance by targeting TCEAL7 | [52] |
| miR-135b | Multiple myeloma | Enhances angiogenesis by targeting factor-inhibiting HIF-1 | [84] |
| Exosomal proteins | |||
| Cell migration-inducing and hyaluronan-binding protein (CEMIP) | Brain cancer | Increases pro-inflammatory cytokines Ptgs2, Tnf, and CCL/CXCL, which promote brain vascular remodeling and metastasis | [85] |
| Integrin β3 | Lung Cancer | Mediates a brain-tropic metastasis pattern and may serve as a novel prognostic biomarker for brain metastasis | [75] |
| Complement factor H (CFH) | Hepatocellular carcinoma | Promotes tumor cell growth, migration, invasiveness, and liver tumor formation in mice | [48] |
| VEGF | TME | Its overexpression along with the activation of VEGFR induces immune-suppressive | [23] |
| Wnt5b | Pancreatic cancer | Promotes cancer cell migration and proliferation | [76] |
| Exosomal lipids | |||
| Triglycerides | Prostate cancer | The activation of lipogenesis-related enzymes and signaling molecules causes increased accumulation of triglycerides in exosomes | [83] |
| Phosphatidylserine | TME | Externalization of phosphatidylserine from the inner to the outer membrane leaflet of cells and exosomes provides strong immune-suppressive signals | [23] |
| Phosphatidylserine | - | A critical molecule in the exosomal uptake by HUVECs. | [81] |
| Immune Cell | Mechanism Involved | Effects Observed | Reference |
|---|---|---|---|
| Macrophage | The transfer of let-7a miRNA resulted in the suppression of the insulin-Akt-mTOR signaling pathway | Improved macrophage recruitment and M2-like polarization in vitro and in vivo Increased expression of immunomodulators, such as CSF-1, CCL2, FTH, FTL, and TGFβ | [24] |
| Macrophage | miR-301a-3p activates the PTEN/PI3Kγ signaling pathway | Hypoxic exosomal miR-301a-3p induces M2 polarization of macrophages Hypoxic exosomes enhance malignant behaviors of pancreatic cancer cells | [78] |
| Macrophage | Macrophage expression of HIF-1α | Tumor-associated macrophages suppress tumor-infiltrating T cells | [146] |
| Macrophage | Exosomal miRNAs are induced by hypoxia vian HIFs | Tumor-associated macrophages educated by hypoxic exosomes derived from cancer cells promote tumor proliferation and migration in a feedback loop. | [147] |
| Macrophages Monocytes | MyD88-p38-STAT3 signaling The tumor cell-released TLR4-mediated autophagosomes-PD-L1 axis | Tumor cell-released autophagosomes-induced macrophage polarization into M2-like phenotype characterized by the expression of PD-L1 and IL-10 M2-like phenotype with increased expression of PD-L1, CD163, and IL-10, but decreased HLA-DR with T cell-suppressive function | [93] |
| Monocyte | Exosomal delivering of miRNA-21 | Monocyte transformation to M2-like macrophages via miRNA-21, with increased expression of IL-10 and CD206 | [98] |
| T-cells | miR-24-3p targets FGF11 to inhibit T-cell function | Hypoxia increases cellular and exosomal miR-24-3p levels and enhances the inhibitory effect on T-cell proliferation and differentiation | [57] |
| T cells | Targeted depletion or elimination of hypoxia in tumors | Increased T cells infiltration into hypoxic zones and downregulation of MDSCs | [105] |
| Γδ T-cell (lymphocyte) | Hypoxic exosomes regulate MDSC function in a miR-21/PTEN/ PD-L1-axis-dependent manner | There is an enhanced suppressive effect of MDSCs on γδ T cells | [108] |
| γδ T cells | Reduced calcium efflux and the expression of CD107a in γδT cells | Decreased antitumor cytotoxicity of γδT cells observed under hypoxia | [114] |
| MDSC | Increased level of exosomal S100A9 vian HIF-1α-dependent mechanism | MDSCs enhance colorectal cancer cell stemness and growth | [119] |
| MDSCs Regulatory B cells | Microvesicles transport membrane-bound PD-L1 from MDSCs to B cells | Suppressed CD8+ T-cell activation, and increased CD155, TGFβ, and IL10 | [135] |
| DC | PD-1/PD-L1 pathway | DCs treated with tumor cells exosomes significantly increase PD-1+CD8+T cells | [130] |
| DC | Hypoxia induces upregulation of microRNA 21 in DCs | Decreased expressions of CD80, CD86, and MHCII on DCs | [126] |
| B cells | Hypoxia-induced IL-10 secretion via HMGB1 | Hypoxia significantly enhances the level of HMGB1 on tumor cell-released autophagosomes leading to the induction of IL-10-producing B cells that suppress CD4+ and CD8+ T cells | [134] |
| B cells | IL-10-dependent manner Activation of the TLR2-MyD88-NF-κB signal pathway in B cells | B cells differentiate into IL-10-producing regulatory B cells with a distinct phenotype of CD1d(+) CD5(+), which could potently inhibit CD8(+) and CD4(+) T cell responses | [133] |
| NK cells | Hypoxic tumor-derived microvesicles miR-23a and TGF-β1 | Transfer of TGF-β1 and miR-23a to NK cells, decreases NKG2D, thereby inhibiting NK cell function. | [100] |
| Mast cells | CD40L-CD40 interaction | Promote PMN-MDSCs activity and T-cell inactivity to favor the suppression of antitumor activities and encourage tumor onset | [137] |
| Mast cells | Increased expressions of HIF-1α, VEGF, and H1 | Increased tumor growth and angiogenesis. And decreased survival rate of the mice | [139] |
| Mast cells | CCL5 dependent | Increased suppression of antitumor function and enhanced tumor progression | [141] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shao, X.; Hua, S.; Feng, T.; Ocansey, D.K.W.; Yin, L. Hypoxia-Regulated Tumor-Derived Exosomes and Tumor Progression: A Focus on Immune Evasion. Int. J. Mol. Sci. 2022, 23, 11789. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms231911789
Shao X, Hua S, Feng T, Ocansey DKW, Yin L. Hypoxia-Regulated Tumor-Derived Exosomes and Tumor Progression: A Focus on Immune Evasion. International Journal of Molecular Sciences. 2022; 23(19):11789. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms231911789
Chicago/Turabian StyleShao, Xuejun, Shenghao Hua, Tao Feng, Dickson Kofi Wiredu Ocansey, and Lei Yin. 2022. "Hypoxia-Regulated Tumor-Derived Exosomes and Tumor Progression: A Focus on Immune Evasion" International Journal of Molecular Sciences 23, no. 19: 11789. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms231911789