
Nestin+ Mesenchymal Precursors Generate Distinct Spleen Stromal Cell Subsets and Have Immunomodulatory Function
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
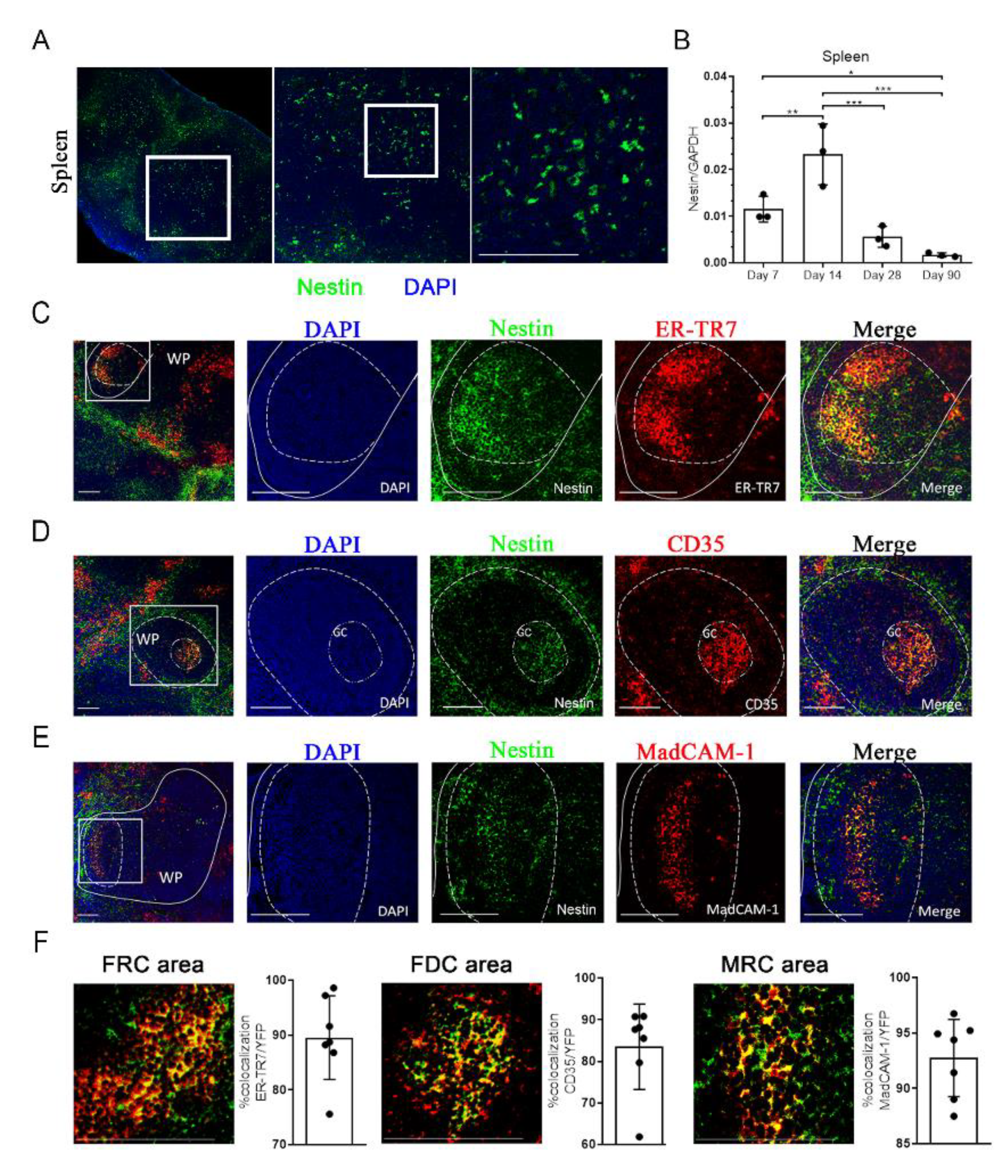
2.1. Characterization of Nestin Expression in the Spleen
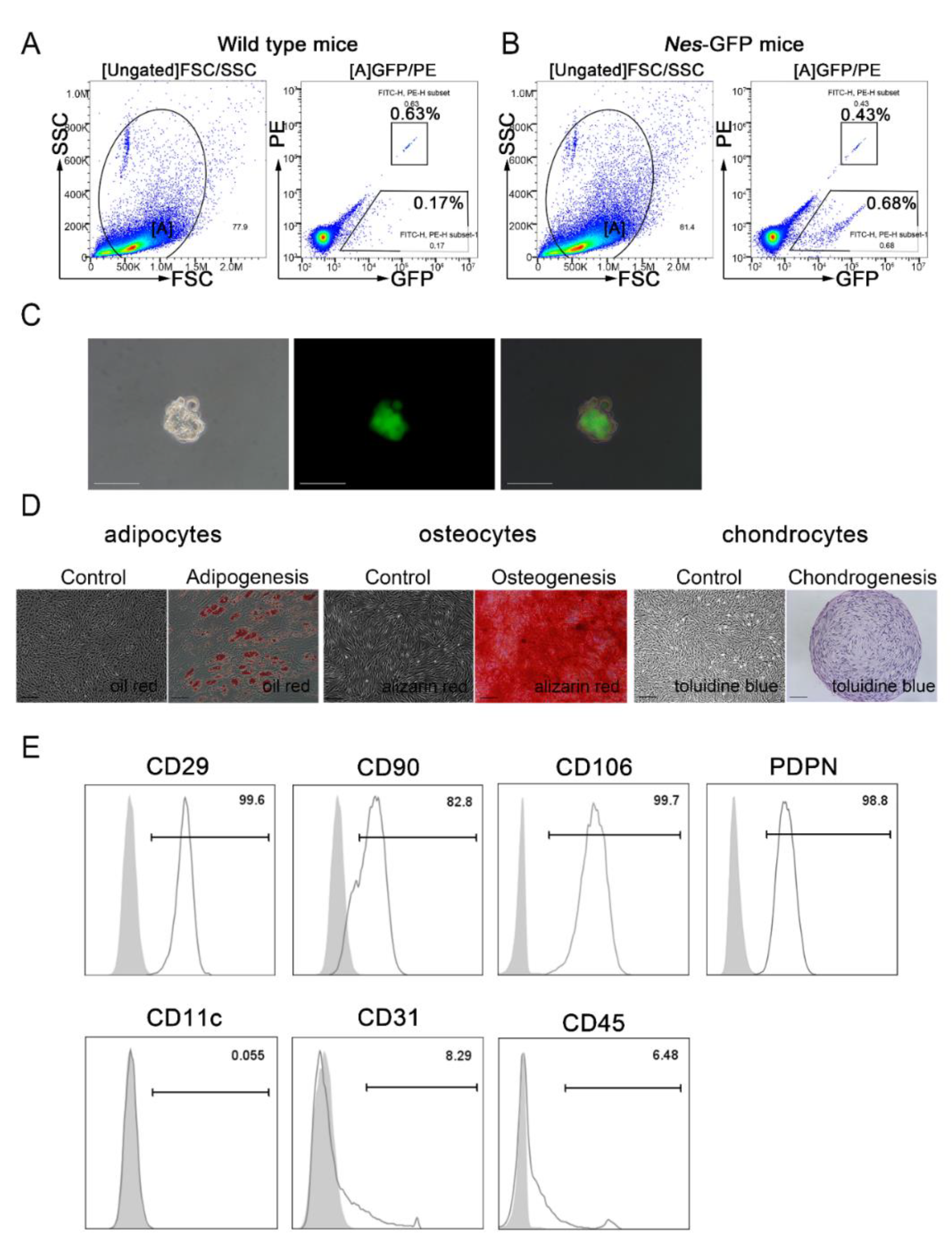
2.2. Isolation and Characterization of Nes-GFP+ Cells from the Spleen
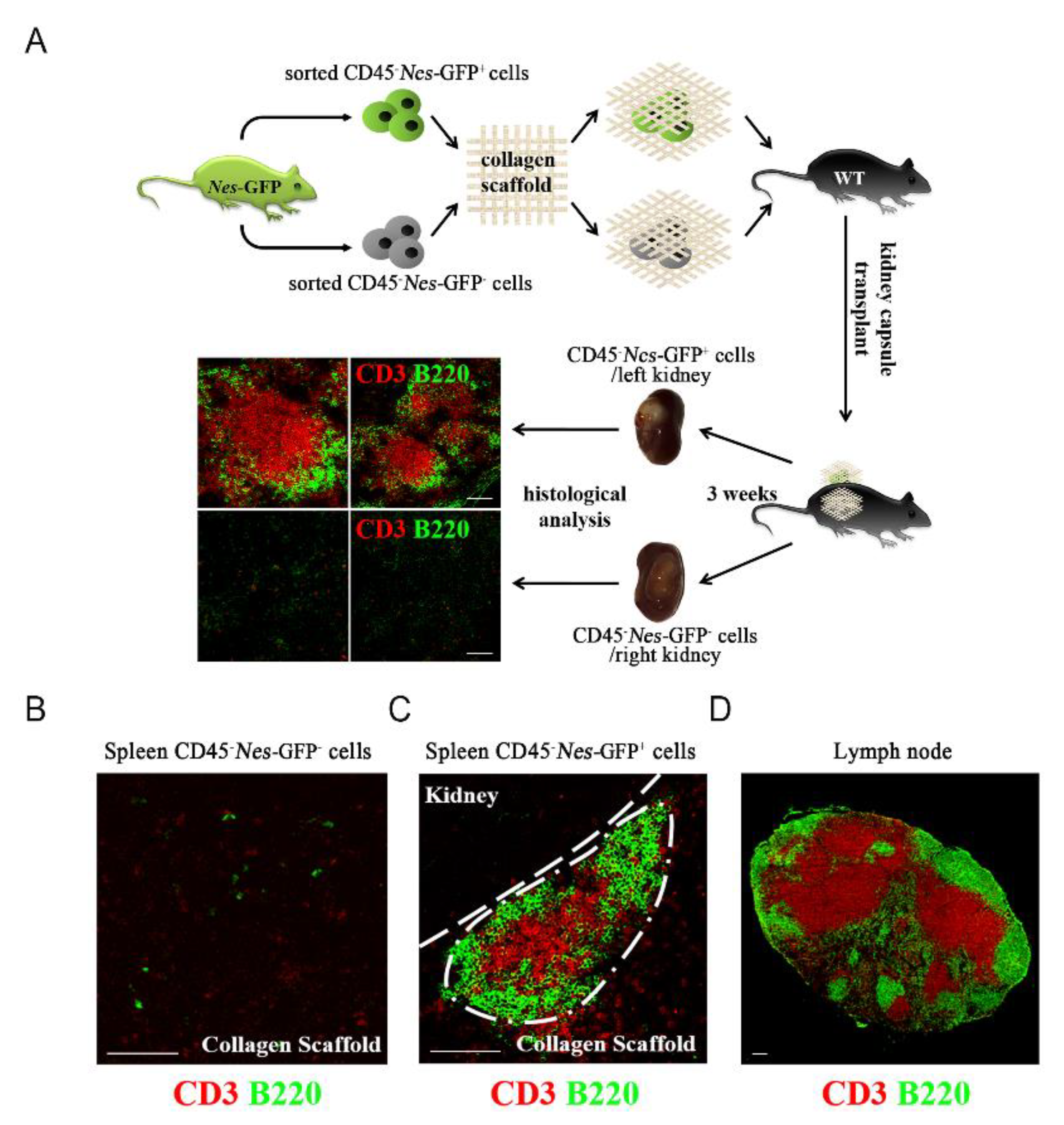
2.3. Spleen-Derived Nes-GFP+ Cells Could Form Secondary Lymphoid Organs
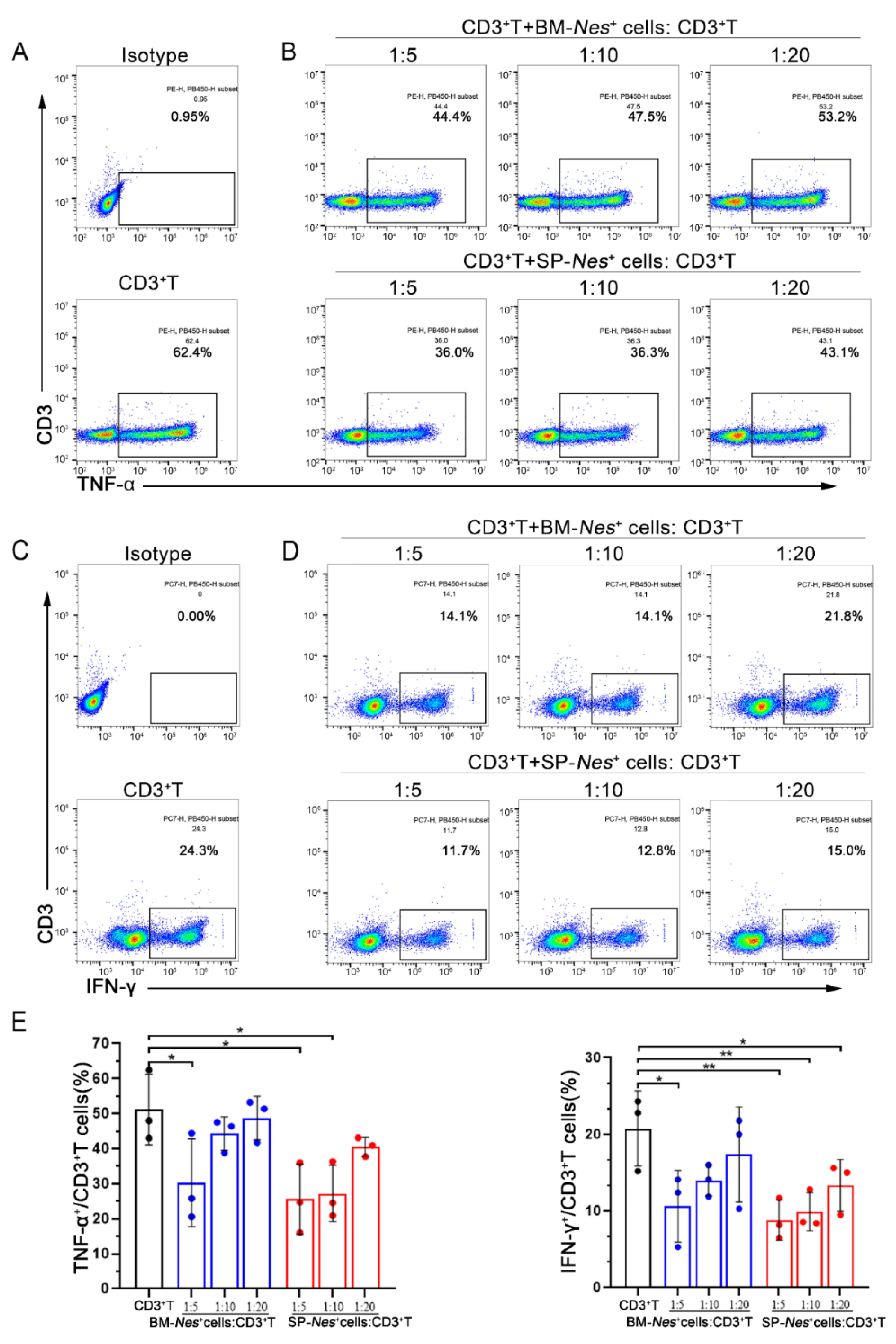
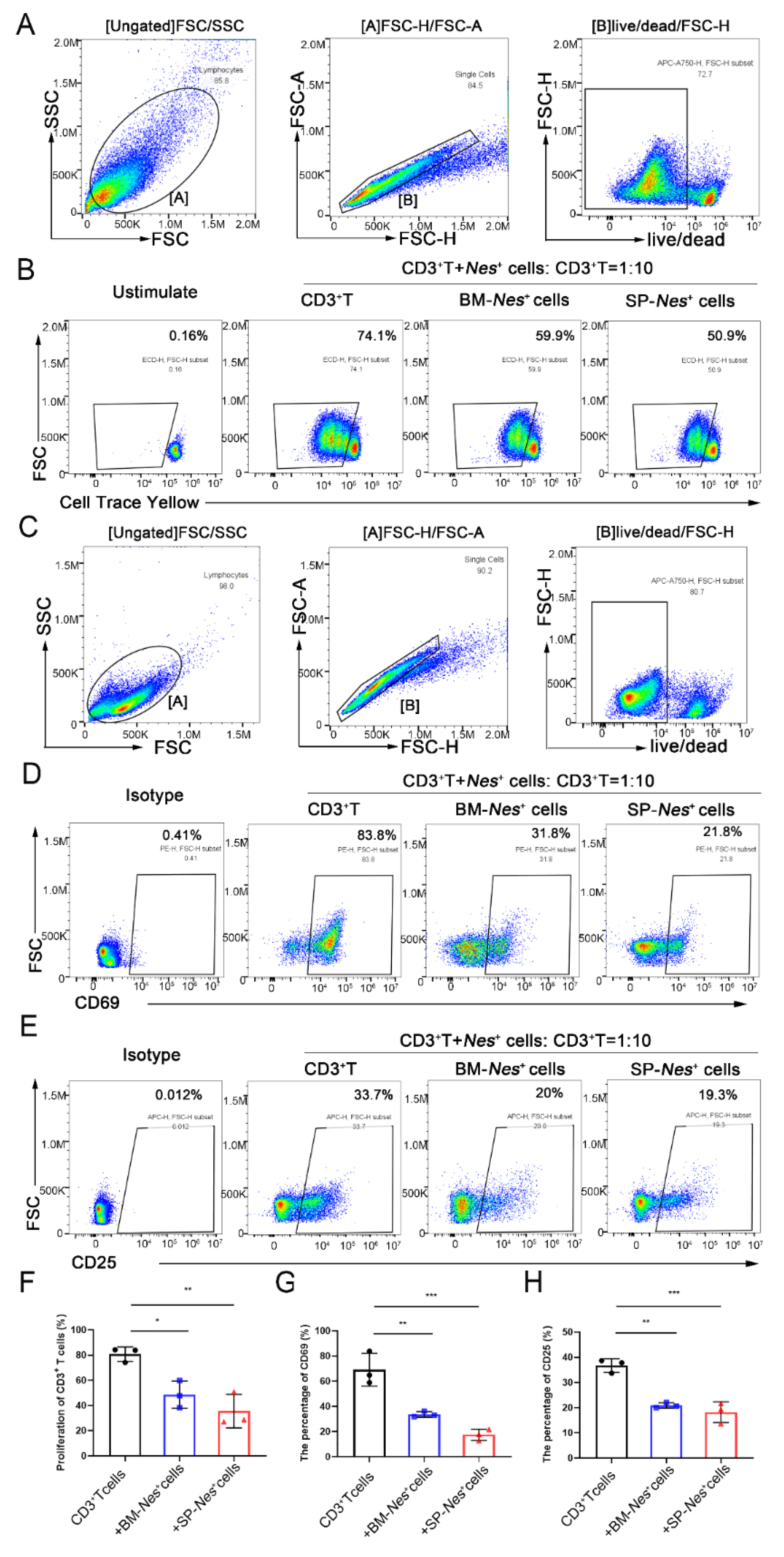
2.4. Immunosuppressive Properties of Nes-GFP+ Cells Derived from the Spleen
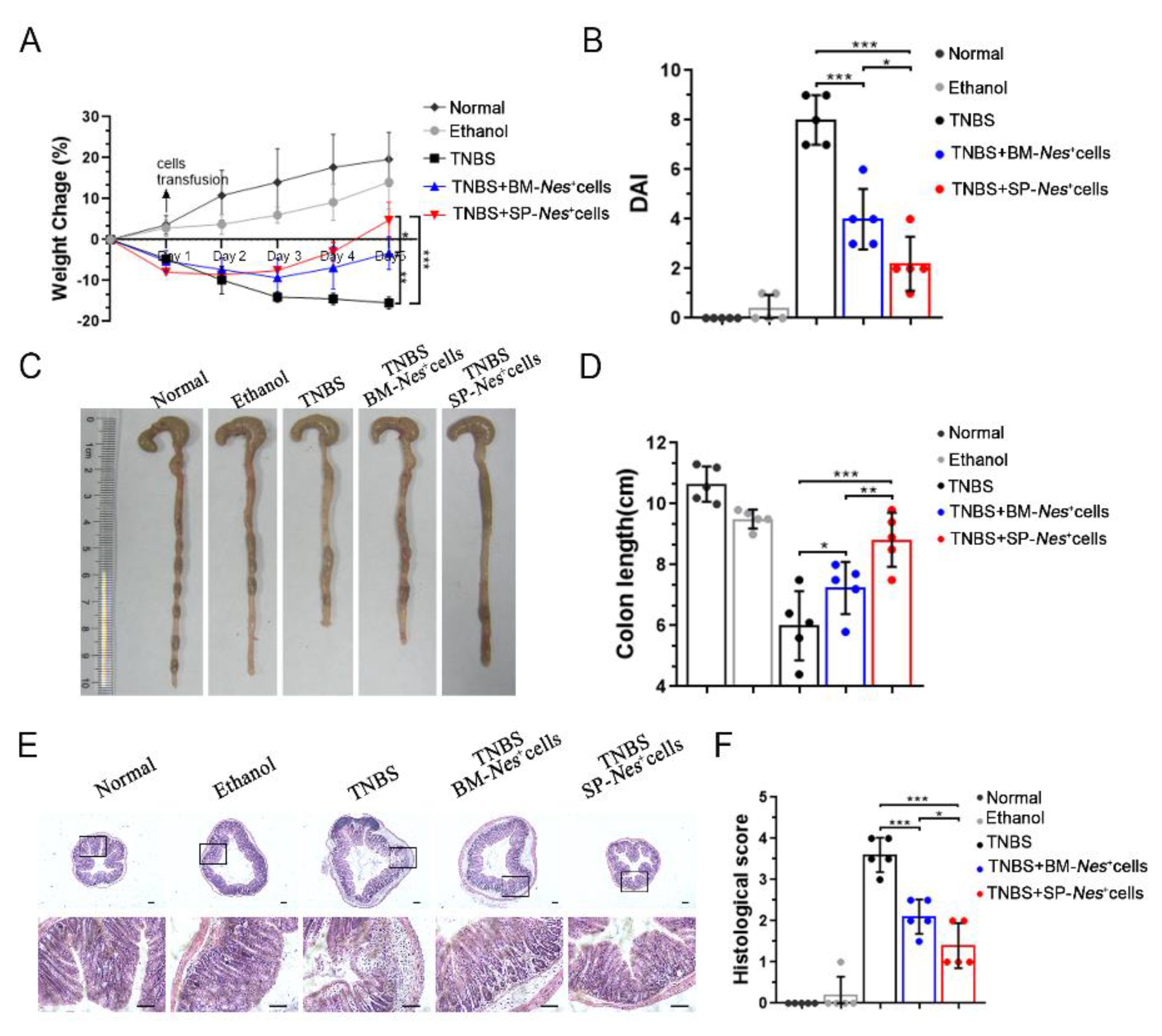
2.5. Spleen-Derived Nes-GFP+ Cells as a Potential Source for the Treatment of Inflammatory Diseases
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Isolation and Culture of Nes-GFP+ Cells from Nes-GFP Transgenic Mice
4.3. RNA Isolation and Quantitative Real-Time and Reverse Transcription PCR
4.4. Clonal Sphere Formation Assay
4.5. Cell Differentiation Ability In Vitro
4.6. Flow Cytometric Analysis
4.7. Immunofluorescence Staining
4.8. Artificial Lymphoid Formation
4.9. Cytokine Assays
4.10. Proliferation Assay
4.11. T cells Activation
4.12. Inflammatory Bowel Disease (IBD) Model
4.13. Contact Hypersensitivity Model
4.14. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sacchetti, B.F.; Michienzi, S.; di Cesare, S.; Piersanti, S.; Saggio, I.; Tagliafico, E.; Ferrari, S.; Robey, P.G.; Riminucci, M.; Bianco, P. Self-Renewing Osteoprogenitors in Bone Marrow Sinusoids Can Organize a Hematopoietic Microenvironment. Cell 2007, 131, 324–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marrazzo, P.; Pizzuti, V.; Zia, S.; Sargenti, A.; Gazzola, D.; Roda, B.; Bonsi, L.; Alviano, F. Microfluidic Tools for Enhanced Characterization of Therapeutic Stem Cells and Prediction of Their Potential Antimicrobial Secretome. Antibiotics 2021, 10, 750. [Google Scholar] [CrossRef] [PubMed]
- Le Blanc, K.; Frassoni, F.; Ball, L.; Locatelli, F.; Roelofs, H.; Lewis, I.; Lanino, E.; Sundberg, B.; Bernardo, M.E.; Remberger, M.; et al. Mesenchymal stem cells for treatment of steroid-resistant, severe, acute graft-versus-host disease: A phase II study. Lancet 2008, 371, 1579–1586. [Google Scholar] [CrossRef]
- Zhang, S.; Chuah, S.J.; Lai, R.C.; Hui, J.H.P.; Lim, S.K.; Toh, W.S. MSC exosomes mediate cartilage repair by enhancing proliferation, attenuating apoptosis and modulating immune reactivity. Biomaterials 2018, 156, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhou, T.; Zhang, Z.; Liu, H.; Zheng, Z.; Xie, H. Current therapeutic strategies for respiratory diseases using mesenchymal stem cells. MedComm 2021, 2, 351–380. [Google Scholar] [CrossRef] [PubMed]
- Shin, T.-H.; Kim, H.-S.; Kang, T.-W.; Lee, B.-C.; Lee, H.-Y.; Kim, Y.-J.; Shin, J.-H.; Seo, Y.; Choi, S.W.; Lee, S.; et al. Human umbilical cord blood-stem cells direct macrophage polarization and block inflammasome activation to alleviate rheumatoid arthritis. Cell Death Dis. 2016, 7, e2524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Zhang, H.; Liang, J.; Li, X.; Feng, X.; Wang, H.; Hua, B.; Liu, B.; Lu, L.; Gilkeson, G.S.; et al. Allogeneic mesenchymal stem cell transplantation in severe and refractory systemic lupus erythematosus: 4 years of experience. Cell Transpl. 2013, 22, 2267–2277. [Google Scholar] [CrossRef]
- Duran, J.M.; Makarewich, C.A.; Sharp, T.E.; Starosta, T.; Zhu, F.; Hoffman, N.E.; Chiba, Y.; Madesh, M.; Berretta, R.M.; Kubo, H.; et al. Bone-derived stem cells repair the heart after myocardial infarction through transdifferentiation and paracrine signaling mechanisms. Circ. Res. 2013, 113, 539–552. [Google Scholar] [CrossRef] [Green Version]
- Bang, O.Y.; Lee, J.S.; Lee, P.H.; Lee, G. Autologous mesenchymal stem cell transplantation in stroke patients. Ann. Neurol. 2005, 57, 874–882. [Google Scholar] [CrossRef]
- Panés, J.; García-Olmo, D.; Van Assche, G.; Colombel, J.F.; Reinisch, W.; Baumgart, D.C.; Dignass, A.; Nachury, M.; Ferrante, M.; Kazemi-Shirazi, L.; et al. Expanded allogeneic adipose-derived mesenchymal stem cells (Cx601) for complex perianal fistulas in Crohn’s disease: A phase 3 randomised, double-blind controlled trial. Lancet 2016, 388, 1281–1290. [Google Scholar] [CrossRef]
- Shi, Y.; Wang, Y.; Li, Q.; Liu, K.; Hou, J.; Shao, C.; Wang, Y. Immunoregulatory mechanisms of mesenchymal stem and stromal cells in inflammatory diseases. Nat. Rev. Nephrol. 2018, 14, 493–507. [Google Scholar] [CrossRef] [PubMed]
- Steven, M.L.; Williams, A.; Eisenbarth, S.C. Structure and function of the immune system in the spleen. Sci. Immunol. 2019, 4, 6085. [Google Scholar]
- den Haan, J.M.; Mebius, R.E.; Kraal, G. Stromal cells of the mouse spleen. Front. Immunol. 2012, 3, 201. [Google Scholar] [PubMed] [Green Version]
- Cheng, H.-W.; Onder, L.; Novkovic, M.; Soneson, C.; Lütge, M.; Pikor, N.; Scandella, E.; Robinson, M.D.; Miyazaki, J.-I.; Tersteegen, A.; et al. Origin and differentiation trajectories of fibroblastic reticular cells in the splenic white pulp. Nat. Commun. 2019, 10, 1739. [Google Scholar] [CrossRef] [Green Version]
- Amé-Thomas, P.; Hajjami, H.M.-E.; Monvoisin, C.; Jean, R.; Monnier, D.; Caulet-Maugendre, S.; Guillaudeux, T.; Lamy, T.; Fest, T.; Tarte, K. Human mesenchymal stem cells isolated from bone marrow and lymphoid organs support tumor B-cell growth: Role of stromal cells in follicular lymphoma pathogenesis. Blood 2007, 109, 693–702. [Google Scholar] [CrossRef] [Green Version]
- Krishnamurty, A.T.; Turley, S.J. Lymph node stromal cells: Cartographers of the immune system. Nat. Immunol. 2020, 21, 369–380. [Google Scholar] [CrossRef]
- Fletcher, A.L.; Astarita, J.; Murray, R.; Saeidi, N.; D’Rozario, J.; Knoblich, K.; Brown, F.D.; Schildberg, F.A.; Nieves, J.M.; Heng, T.S.; et al. Lymph node fibroblastic reticular cell transplants show robust therapeutic efficacy in high-mortality murine sepsis. Sci. Transl. Med. 2014, 13, 249ra109. [Google Scholar] [CrossRef] [Green Version]
- Lendahl, U.; McKay, R.D. CNS Stem Cells Express a New Class of Intermediate Filament Protein. Cell 1990, 60, 585–595. [Google Scholar] [CrossRef]
- Zulewski, H.; Abraham, E.J.; Gerlach, M.J.; Daniel, P.B.; Moritz, W.; Müller, B.; Vallejo, M.; Thomas, M.K.; Habener, J.F. Multipotential Nestin-Positive Stem Cells Isolated From Adult Pancreatic Islets Differentiate Ex Vivo Into Pancreatic Endocrine, Exocrine, and Hepatic Phenotypes. Diabetes 2001, 50, 521–533. [Google Scholar] [CrossRef]
- Jiang, M.H.; Cai, B.; Tuo, Y.; Wang, J.; Zang, Z.J.; Tu, X.; Gao, Y.; Su, Z.; Li, W.; Li, G.; et al. Characterization of Nestin-positive stem Leydig cells as a potential source for the treatment of testicular Leydig cell dysfunction. Cell Res. 2014, 24, 1466–1485. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Li, G.; Zhang, X.; Huang, W.; Xie, D.; Dai, G.; Zhu, S.; Lu, D.; Zhang, Z.; Lin, J.; et al. Cardiac Nestin(+) Mesenchymal Stromal Cells Enhance Healing of Ischemic Heart through Periostin-Mediated M2 Macrophage Polarization. Mol. Ther. 2020, 28, 855–873. [Google Scholar] [CrossRef] [PubMed]
- Amoh, Y.; Li, L.; Katsuoka, K.; Penman, S.; Hoffman, R.M. Multipotent nestin-positive, keratin-negative hair-follicle bulge stem cells can form neurons. Proc. Natl. Acad. Sci. USA 2005, 102, 5530–5534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, M.H.; Li, G.; Liu, J.; Liu, L.; Wu, B.; Huang, W.; He, W.; Deng, C.; Wang, D.; Li, C.; et al. Nestin(+) kidney resident mesenchymal stem cells for the treatment of acute kidney ischemia injury. Biomaterials 2015, 50, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Katakai, T.; Hara, T.; Lee, J.-H.; Gonda, H.; Sugai, M.; Shimizu, A. A novel reticular stromal structure in lymph node cortex: An immuno-platform for interactions among dendritic cells, T cells and B cells. Int. Immunol. 2004, 16, 1133–1142. [Google Scholar] [CrossRef] [Green Version]
- Fischer, M.B.; Goerg, S.; Shen, L.; Prodeus, A.P.; Goodnow, C.C.; Kelsoe, G.; Carroll, M.C. Dependence of Germinal Center B Cells on Expression of CD21/CD35 for Survival. Science 1998, 280, 582–585. [Google Scholar] [CrossRef]
- Katakai, T.; Suto, H.; Sugai, M.; Gonda, H.; Togawa, A.; Suematsu, S.; Ebisuno, Y.; Katagiri, K.; Kinashi, T.; Shimizu, A. Organizer-like reticular stromal cell layer common to adult secondary lymphoid organs. J. Immunol. 2008, 181, 6189–6200. [Google Scholar] [CrossRef] [Green Version]
- Honda, K.; Nakano, H.; Yoshida, H.; Nishikawa, S.; Rennert, P.; Ikuta, K.; Tamechika, M.; Yamaguchi, K.; Fukumoto, T.; Chiba, T.; et al. Molecular basis for hematopoietic/mesenchymal interaction during initiation of Peyer’s patch organogenesis. J. Exp. Med. 2001, 193, 621–630. [Google Scholar] [CrossRef]
- Yin, Z.; Hu, J.; Yang, L.; Zheng, Z.; An, C.; Wu, B.; Zhang, C.; Shen, W.; Liu, H.; Chen, J.; et al. Single-cell analysis reveals a nestin(+) tendon stem/progenitor cell population with strong tenogenic potentiality. Sci. Adv. 2016, 2, e1600874. [Google Scholar] [CrossRef] [Green Version]
- Suematsu, S.; Watanabe, T. Generation of a synthetic lymphoid tissue-like organoid in mice. Nat. Biotechnol. 2004, 22, 1539–1545. [Google Scholar] [CrossRef]
- Li, W.Q.; Huang, L.; Lin, W.; Ke, Q.; Chen, R.; Lai, X.; Wang, X.; Zhang, J.; Jiang, M.; Huang, W.; et al. Engraftable neural crest stem cells derived from cynomolgus monkey embryonic stem cells. Biomaterials 2015, 39, 75–84. [Google Scholar] [CrossRef]
- Castagnaro, L.; Lenti, E.; Maruzzelli, S.; Spinardi, L.; Migliori, E.; Farinello, D.; Sitia, G.; Harrelson, Z.; Evans, S.M.; Guidotti, L.G.; et al. Nkx2-5(+)islet1(+) mesenchymal precursors generate distinct spleen stromal cell subsets and participate in restoring stromal network integrity. Immunity 2013, 38, 782–791. [Google Scholar] [CrossRef] [PubMed]
- Krautler, N.J.; Kana, V.; Kranich, J.; Tian, Y.; Perera, D.; Lemm, D.; Schwarz, P.; Armulik, A.; Browning, J.L.; Tallquist, M.; et al. Follicular dendritic cells emerge from ubiquitous perivascular precursors. Cell 2012, 150, 194–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajenoff, M.; Glaichenhaus, N.; Germain, R.N. Fibroblastic reticular cells guide T lymphocyte entry into and migration within the splenic T cell zone. J. Immunol. 2008, 181, 3947–3954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haberman, A.M.; Shlomchik, M.J. Reassessing the function of immune-complex retention by follicular dendritic cells. Nat. Rev. Immunol. 2003, 3, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.W.; Chen, X.; Liu, Q.; Zhang, X.; Huang, K.; Liu, L.; Li, H.; Zhou, M.; Huang, F.; Fan, Z.; et al. Mesencymal stromal cells infusions improve refractory chronic graft versus host disease through an increase of CD5+ regulatory B cells producing interleukin 10. Leukemia 2015, 29, 636–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.Y.; Cai, C.; Xu, D.; Liu, Q.; Zheng, S.; Liu, L.; Li, G.; Zhang, X.; Li, X.; Ma, Y.; et al. Human Mesenchymal Stem Cell-Treated Regulatory CD23+CD43+ B Cells Alleviate Intestinal Inflammation. Theranostics 2019, 9, 4633–4647. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Saito, H.; Suzuki, M.; Mori, K. Visualization of neurogenesis in the central nervous system using nestin promoter-GFP transgenic mice. NeuroReport 2000, 11, 1991–1996. [Google Scholar] [CrossRef]
- Nagano, T.; Katase, M.; Tsumura, K.; Saito, M.; Matsuda, T. Inhibitory effects of dietary soyasaponin on 2,4-dinitrofluorobenzene-induced contact hypersensitivity in mice. Exp. Dermatol. 2017, 26, 249–254. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, J.; Deng, R.; Li, W.; Jiang, M.; Xiang, A.P.; Zhang, X. Nestin+ Mesenchymal Precursors Generate Distinct Spleen Stromal Cell Subsets and Have Immunomodulatory Function. Int. J. Mol. Sci. 2022, 23, 11819. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms231911819
Huang J, Deng R, Li W, Jiang M, Xiang AP, Zhang X. Nestin+ Mesenchymal Precursors Generate Distinct Spleen Stromal Cell Subsets and Have Immunomodulatory Function. International Journal of Molecular Sciences. 2022; 23(19):11819. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms231911819
Chicago/Turabian StyleHuang, Jing, Ronghai Deng, Weiqiang Li, Meihua Jiang, Andy Peng Xiang, and Xiaoran Zhang. 2022. "Nestin+ Mesenchymal Precursors Generate Distinct Spleen Stromal Cell Subsets and Have Immunomodulatory Function" International Journal of Molecular Sciences 23, no. 19: 11819. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms231911819