
Multifunctional Interleukin-24 Resolves Neuroretina Autoimmunity via Diverse Mechanisms


Abstract
:1. Introduction
2. Results
2.1. IL-24 Inhibits Differentiation of Th1 and Th17 Cells and Their Production of Proinflammatory Cytokines
2.2. IL-24 Does Not Induce the Expression of Immunoregulatory IL-10 or the Differentiation of Regulatory T Cells
2.3. Intravitreal Injection of IL-24 Resolves Ocular Inflammation and Suppresses Proinflammatory Cytokine Production from Ocular-Infiltrating CD4+ T Cells

2.4. IL-24 Inhibits Peripheral Antigen-Specific Response
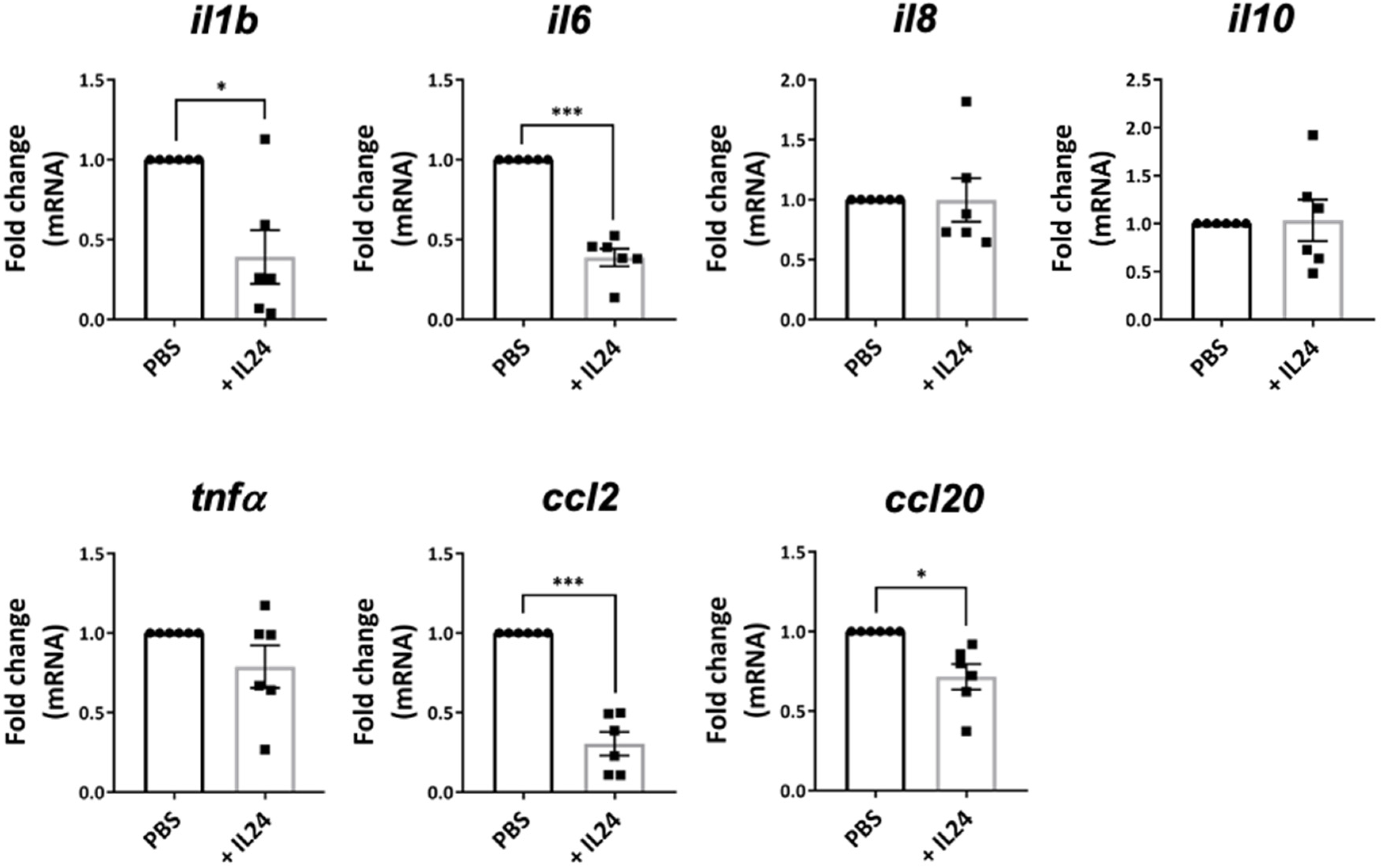
2.5. IL-24 Reduces the Expression of Proinflammatory Molecules by the Retina in EAU Mice
2.6. IL-24 Inhibits the Production of Proinflammatory Molecules by Human RPE Cells
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. ARPE-19 Cell Cultures
4.3. Induction, Treatment, and Evaluation of EAU
4.4. Isolation of Infiltrating Cells from Eyes
4.5. Intracellular Cytokine Staining for FCM Analysis
4.6. T-cell Activation and Differentiation
4.7. Real-Time PCR
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Caspi, R.R. A look at autoimmunity and inflammation in the eye. J. Clin. Investig. 2010, 120, 3073–3083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chong, W.P.; Horai, R.; Mattapallil, M.J.; Silver, P.B.; Chen, J.; Zhou, R.; Sergeev, Y.; Villasmil, R.; Chan, C.C.; Caspi, R.R. IL-27p28 inhibits central nervous system autoimmunity by concurrently antagonizing Th1 and Th17 responses. J. Autoimmun. 2014, 50, 12–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luger, D.; Silver, P.B.; Tang, J.; Cua, D.; Chen, Z.; Iwakura, Y.; Bowman, E.P.; Sgambellone, N.M.; Chan, C.C.; Caspi, R.R. Either a Th17 or a Th1 effector response can drive autoimmunity: Conditions of disease induction affect dominant effector category. J. Exp. Med. 2008, 205, 799–810. [Google Scholar] [CrossRef] [PubMed]
- Horai, R.; Silver, P.B.; Chen, J.; Agarwal, R.K.; Chong, W.P.; Jittayasothorn, Y.; Mattapallil, M.J.; Nguyen, S.; Natarajan, K.; Villasmil, R.; et al. Breakdown of immune privilege and spontaneous autoimmunity in mice expressing a transgenic T cell receptor specific for a retinal autoantigen. J. Autoimmun. 2013, 44, 21–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horai, R.; Zarate-Blades, C.R.; Dillenburg-Pilla, P.; Chen, J.; Kielczewski, J.L.; Silver, P.B.; Jittayasothorn, Y.; Chan, C.C.; Yamane, H.; Honda, K.; et al. Microbiota-Dependent Activation of an Autoreactive T Cell Receptor Provokes Autoimmunity in an Immunologically Privileged Site. Immunity 2015, 43, 343–353. [Google Scholar] [CrossRef] [Green Version]
- Taylor, A.W.; Hsu, S.; Ng, T.F. The Role of Retinal Pigment Epithelial Cells in Regulation of Macrophages/Microglial Cells in Retinal Immunobiology. Front. Immunol. 2021, 12, 724601. [Google Scholar] [CrossRef]
- Detrick, B.; Hooks, J.J. Immune regulation in the retina. Immunol. Res. 2010, 47, 153–161. [Google Scholar] [CrossRef]
- Chen, J.; Caspi, R.R.; Chong, W.P. IL-20 receptor cytokines in autoimmune diseases. J. Leukoc. Biol. 2018, 104, 953–959. [Google Scholar] [CrossRef]
- Zhong, Y.; Zhang, X.; Chong, W. Interleukin-24 Immunobiology and Its Roles in Inflammatory Diseases. Int. J. Mol. Sci. 2022, 23, 627. [Google Scholar] [CrossRef]
- Bech, R.; Otkjaer, K.; Birkelund, S.; Vorup-Jensen, T.; Agger, R.; Johansen, C.; Iversen, L.; Kragballe, K.; Romer, J. Interleukin 20 protein locates to distinct mononuclear cells in psoriatic skin. Exp. Dermatol. 2014, 23, 349–352. [Google Scholar] [CrossRef]
- Kragstrup, T.W.; Otkjaer, K.; Holm, C.; Jorgensen, A.; Hokland, M.; Iversen, L.; Deleuran, B. The expression of IL-20 and IL-24 and their shared receptors are increased in rheumatoid arthritis and spondyloarthropathy. Cytokine 2008, 41, 16–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonseca-Camarillo, G.; Furuzawa-Carballeda, J.; Granados, J.; Yamamoto-Furusho, J.K. Expression of interleukin (IL)-19 and IL-24 in inflammatory bowel disease patients: A cross-sectional study. Clin. Exp. Immunol. 2014, 177, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Muls, N.; Nasr, Z.; Dang, H.A.; Sindic, C.; van Pesch, V. IL-22, GM-CSF and IL-17 in peripheral CD4+ T cell subpopulations during multiple sclerosis relapses and remission. Impact of corticosteroid therapy. PLoS ONE 2017, 12, e0173780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaefer, G.; Venkataraman, C.; Schindler, U. Cutting edge: FISP (IL-4-induced secreted protein), a novel cytokine-like molecule secreted by Th2 cells. J. Immunol. 2001, 166, 5859–5863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anuradha, R.; George, P.J.; Hanna, L.E.; Kumaran, P.; Chandrasekaran, V.; Nutman, T.B.; Babu, S. Expansion of parasite-specific CD4+ and CD8+ T cells expressing IL-10 superfamily cytokine members and their regulation in human lymphatic filariasis. PLoS Negl. Trop. Dis. 2014, 8, e2762. [Google Scholar] [CrossRef]
- Kumar, N.P.; Moideen, K.; Banurekha, V.V.; Nair, D.; Babu, S. Modulation of Th1/Tc1 and Th17/Tc17 responses in pulmonary tuberculosis by IL-20 subfamily of cytokines. Cytokine 2018, 108, 190–196. [Google Scholar] [CrossRef]
- Chong, W.P.; Mattapallil, M.J.; Raychaudhuri, K.; Bing, S.J.; Wu, S.; Zhong, Y.; Wang, W.; Chen, Z.; Silver, P.B.; Jittayasothorn, Y.; et al. The Cytokine IL-17A Limits Th17 Pathogenicity via a Negative Feedback Loop Driven by Autocrine Induction of IL-24. Immunity 2020, 53, 384–397 e5. [Google Scholar] [CrossRef]
- Kumari, S.; Bonnet, M.C.; Ulvmar, M.H.; Wolk, K.; Karagianni, N.; Witte, E.; Uthoff-Hachenberg, C.; Renauld, J.C.; Kollias, G.; Toftgard, R.; et al. Tumor necrosis factor receptor signaling in keratinocytes triggers interleukin-24-dependent psoriasis-like skin inflammation in mice. Immunity 2013, 39, 899–911. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zhou, H.; Huang, X.; Cui, J.; Long, T.; Xu, Y.; Liu, H.; Yu, R.; Zhao, R.; Luo, G.; et al. A Broad Blockade of Signaling from the IL-20 Family of Cytokines Potently Attenuates Collagen-Induced Arthritis. J. Immunol. 2016, 197, 3029–3037. [Google Scholar] [CrossRef] [Green Version]
- Schoenborn, J.R.; Wilson, C.B. Regulation of interferon-gamma during innate and adaptive immune responses. Adv. Immunol. 2007, 96, 41–101. [Google Scholar]
- Singh, R.P.; Hasan, S.; Sharma, S.; Nagra, S.; Yamaguchi, D.T.; Wong, D.T.; Hahn, B.H.; Hossain, A. Th17 cells in inflammation and autoimmunity. Autoimmun. Rev. 2014, 13, 1174–1181. [Google Scholar] [CrossRef] [PubMed]
- Wahl, C.; Muller, W.; Leithauser, F.; Adler, G.; Oswald, F.; Reimann, J.; Schirmbeck, R.; Seier, A.; Weiss, J.M.; Prochnow, B.; et al. IL-20 receptor 2 signaling down-regulates antigen-specific T cell responses. J. Immunol. 2009, 182, 802–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattapallil, M.J.; Kielczewski, J.L.; Zarate-Blades, C.R.; St Leger, A.J.; Raychaudhuri, K.; Silver, P.B.; Jittayasothorn, Y.; Chan, C.C.; Caspi, R.R. Interleukin 22 ameliorates neuropathology and protects from central nervous system autoimmunity. J. Autoimmun. 2019, 102, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Mehta, N.S.; Emami-Naeini, P. A Review of Systemic Biologics and Local Immunosuppressive Medications in Uveitis. J. Ophthalmic Vis. Res. 2022, 17, 276–289. [Google Scholar] [CrossRef]
- Barisani-Asenbauer, T.; Maca, S.M.; Mejdoubi, L.; Emminger, W.; Machold, K.; Auer, H. Uveitis- a rare disease often associated with systemic diseases and infections- a systematic review of 2619 patients. Orphanet J. Rare Dis. 2012, 7, 57. [Google Scholar] [CrossRef] [Green Version]
- Oral, H.B.; Kotenko, S.V.; Yilmaz, M.; Mani, O.; Zumkehr, J.; Blaser, K.; Akdis, C.A.; Akdis, M. Regulation of T cells and cytokines by the interleukin-10 (IL-10)-family cytokines IL-19, IL-20, IL-22, IL-24 and IL-26. Eur. J. Immunol. 2006, 36, 380–388. [Google Scholar] [CrossRef]
- Afkarian, M.; Sedy, J.R.; Yang, J.; Jacobson, N.G.; Cereb, N.; Yang, S.Y.; Murphy, T.L.; Murphy, K.M. T-bet is a STAT1-induced regulator of IL-12R expression in naive CD4+ T cells. Nat. Immunol. 2002, 3, 549–557. [Google Scholar] [CrossRef]
- Mullen, A.C.; High, F.A.; Hutchins, A.S.; Lee, H.W.; Villarino, A.V.; Livingston, D.M.; Kung, A.L.; Cereb, N.; Yao, T.P.; Yang, S.Y.; et al. Role of T-bet in commitment of TH1 cells before IL-12-dependent selection. Science 2001, 292, 1907–1910. [Google Scholar] [CrossRef]
- Ivanov, I.I.; Zhou, L.; Littman, D.R. Transcriptional regulation of Th17 cell differentiation. Semin. Immunol. 2007, 19, 409–417. [Google Scholar] [CrossRef] [Green Version]
- Sauane, M.; Gopalkrishnan, R.V.; Lebedeva, I.; Mei, M.X.; Sarkar, D.; Su, Z.Z.; Kang, D.C.; Dent, P.; Pestka, S.; Fisher, P.B. Mda-7/IL-24 induces apoptosis of diverse cancer cell lines through JAK/STAT-independent pathways. J. Cell. Physiol. 2003, 196, 334–345. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, T.W.; Park, Y.S.; Jeong, E.M.; Lee, D.S.; Kim, I.G.; Chung, H.; Hwang, Y.I.; Lee, W.J.; Yu, H.G.; et al. The Role of Interleukin-22 and Its Receptor in the Development and Pathogenesis of Experimental Autoimmune Uveitis. PLoS ONE 2016, 11, e0154904. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, B.; Maminishkis, A.; Mahesh, S.P.; Yeh, S.; Lew, J.; Lim, W.K.; Sen, H.N.; Clarke, G.; Buggage, R.; et al. Gene expression profiling in autoimmune noninfectious uveitis disease. J. Immunol. 2008, 181, 5147–5157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meitei, H.T.; Jadhav, N.; Lal, G. CCR6-CCL20 axis as a therapeutic target for autoimmune diseases. Autoimmun. Rev. 2021, 20, 102846. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.R.; Mahdi, R.R.; Oh, H.M.; Amadi-Obi, A.; Levy-Clarke, G.; Burton, J.; Eseonu, A.; Lee, Y.; Chan, C.C.; Egwuagu, C.E. Suppressor of cytokine signaling-1 (SOCS1) inhibits lymphocyte recruitment into the retina and protects SOCS1 transgenic rats and mice from ocular inflammation. Investig. Ophthalmol. Vis. Sci. 2011, 52, 6978–6986. [Google Scholar] [CrossRef]
- Sonoda, K.H.; Sakamoto, T.; Qiao, H.; Hisatomi, T.; Oshima, T.; Tsutsumi-Miyahara, C.; Exley, M.; Balk, S.P.; Taniguchi, M.; Ishibashi, T. The analysis of systemic tolerance elicited by antigen inoculation into the vitreous cavity: Vitreous cavity-associated immune deviation. Immunology 2005, 116, 390–399. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Z.; Chong, W.P.; Wu, S.; Wang, W.; Zhou, H.; Chen, J. Comparative Analysis of the Interphotoreceptor Retinoid Binding ProteinInduced Models of Experimental Autoimmune Uveitis in B10.RIII versus C57BL/6 Mice. Curr. Mol. Med. 2018, 18, 602–611. [Google Scholar] [CrossRef]
- Wu, S.; Ma, R.; Zhong, Y.; Chen, Z.; Zhou, H.; Zhou, M.; Chong, W.; Chen, J. Deficiency of IL-27 Signaling Exacerbates Experimental Autoimmune Uveitis with Elevated Uveitogenic Th1 and Th17 Responses. Int. J. Mol. Sci. 2021, 22, 7517. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Human Genes | Primer (5′ to 3′) | |
| gapdh | Forward | ACAACAGCCTCAAGATCATCAG |
| Reverse | TCTTCTGGGTGGCAGTGATG | |
| il6 | Forward | TCTGCGCAGCTTTAAGGAGT |
| Reverse | CCCAGTGGACAGGTTTCTGA | |
| tnfa | Forward | GAGGCCAAGCCCTGGTATG |
| Reverse | CGGGCCGATTGATCTCAGC | |
| ccl2 | Forward | GCTCATAGCAGCCACCTTCATTC |
| Reverse | GTCTTCGGAGTTTGGGTTTGC | |
| il8 | Forward | ACTGAGAGTGATTGAGAGTGGAC |
| Reverse | AACCCTCTGCACCCAGTTTTC | |
| il1b | Forward | TTCGACACATGGGATAACGAGG |
| Reverse | TTTTTGCTGTGAGTCCCGGAG | |
| il10 | Forward | GACTTTAAGGGTTACCTGGGTTG |
| Reverse | TCACATGCGCCTTGATGTCTG | |
| ccl20 | Forward | TGCTGTACCAAGAGTTTGCTC |
| Reverse | CGCACACAGACAACTTTTTCTTT | |
| Mouse Genes | Primer (5′ to 3′) | |
| gapdh | Forward | AGGTCGGTGTGAACGGATTTG |
| Reverse | TGTAGACCATGTAGTTGAGGTCA | |
| ifng | Forward | TGCTGTACCAAGAGTTTGCTC |
| Reverse | CGCACACAGACAACTTTTTCTTT | |
| Il17a | Forward | TTTAACTCCCTTGGCGCAAAA |
| Reverse | CTTTCCCTCCGCATTGACAC | |
| Il17f | Forward | TGCTACTGTTGATGTTGGGAC |
| Reverse | CAGAAATGCCCTGGTTTTGGT | |
| csf2 | Forward | ATCAAAGAAGCCCTGAACCT |
| Reverse | GTGTTTCACAGTCCGTTTCC | |
| rorgt | Forward | AGCTTTGTGCAGATCTAAGG |
| Reverse | TGTCCTCCTCAGTAGGGTAG | |
| t-bet | Forward | AGCAAGGACGGCGAATGTT |
| Reverse | GTGGACATATAAGCGGTTCCC | |
| tnfa | Forward | CCGATGGGTTGTACCTTGTC |
| Reverse | CGGACTCCGCAAAGTCTAAG | |
| ccl2 | Forward | TTAAAAACCTGGATCGGAACCAA |
| Reverse | GCATTAGCTTCAGATTTACGGGT | |
| Il6 | Forward | TAGTCCTTCCTACCCCAATTTCC |
| Reverse | TTGGTCCTTAGCCACTCCTTC | |
| il8 | Forward | TGTTGAGCATGAAAAGCCTCTAT |
| Reverse | AGGTCTCCCGAATTGGAAAGG | |
| il1b | Forward | CTGTGACTCATGGGATGATGATG |
| Reverse | CGGAGCCTGTAGTGCAGTTG | |
| il10 | Forward | ATAACTGCACCCACTTCCCA |
| Reverse | GGGCATCACTTCTACCAGGT | |
| ccl20 | Forward | ATGGCCTGCGGTGGCAAGCGTCTG |
| Reverse | TAGGCTGAGGAGGTTCACAGCCCT | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Hu, C.; Zhong, Y.; Qiao, D.; Chi, W.; Shen, H.; Chong, W. Multifunctional Interleukin-24 Resolves Neuroretina Autoimmunity via Diverse Mechanisms. Int. J. Mol. Sci. 2022, 23, 11988. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms231911988
Zhang X, Hu C, Zhong Y, Qiao D, Chi W, Shen H, Chong W. Multifunctional Interleukin-24 Resolves Neuroretina Autoimmunity via Diverse Mechanisms. International Journal of Molecular Sciences. 2022; 23(19):11988. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms231911988
Chicago/Turabian StyleZhang, Xuan, Cuiping Hu, Yajie Zhong, Dijie Qiao, Wei Chi, Huangxuan Shen, and Waipo Chong. 2022. "Multifunctional Interleukin-24 Resolves Neuroretina Autoimmunity via Diverse Mechanisms" International Journal of Molecular Sciences 23, no. 19: 11988. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms231911988