
Anthocyanin Biosynthesis Associated with Natural Variation in Autumn Leaf Coloration in Quercus aliena Accessions
Abstract
:1. Introduction
2. Results
2.1. Changes of Autumn Leaves in Q. aliena Accessions
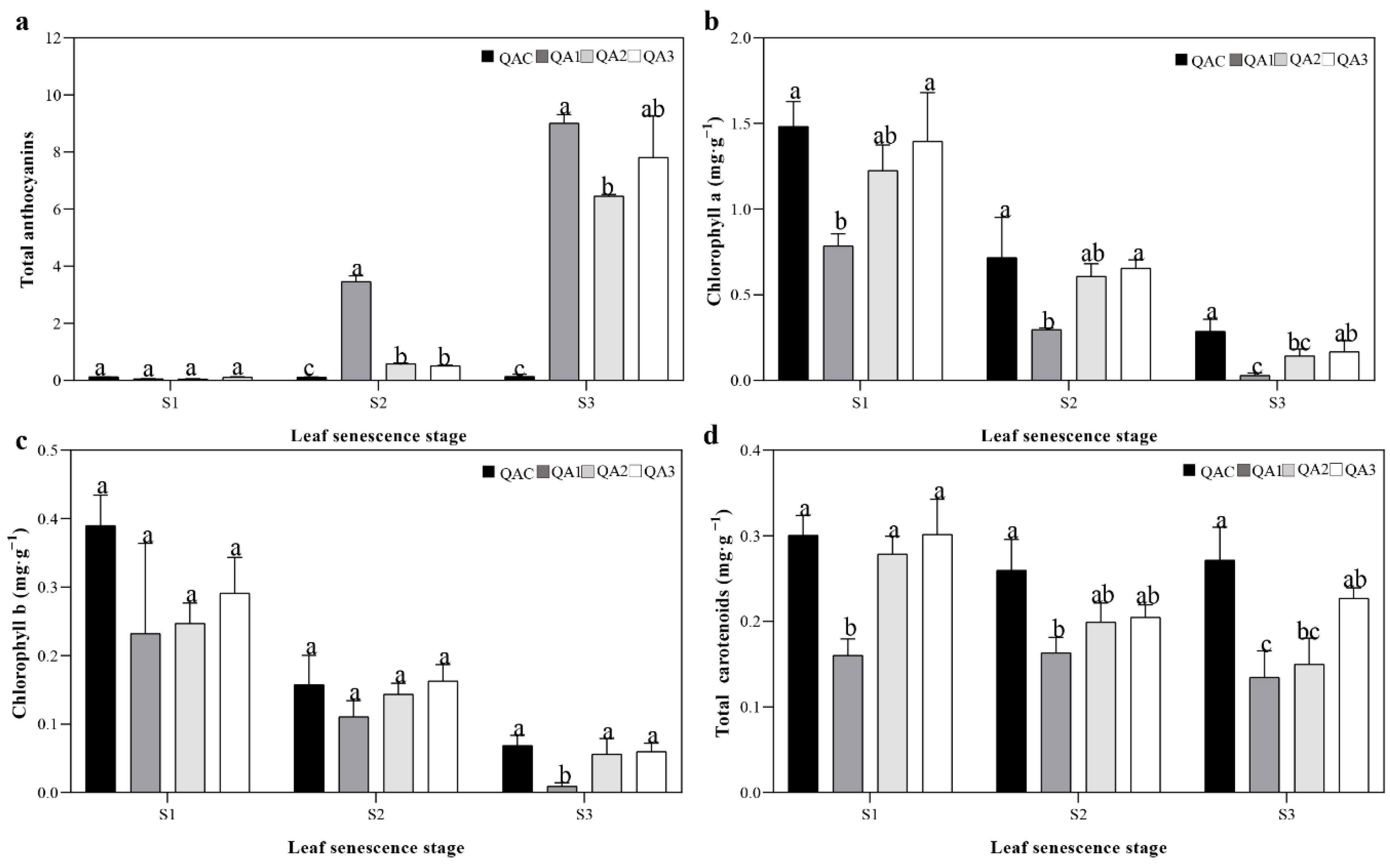
2.2. Contents of Total Anthocyanins, Total Carotenoids, and Chlorophyll in Q. aliena Accessions
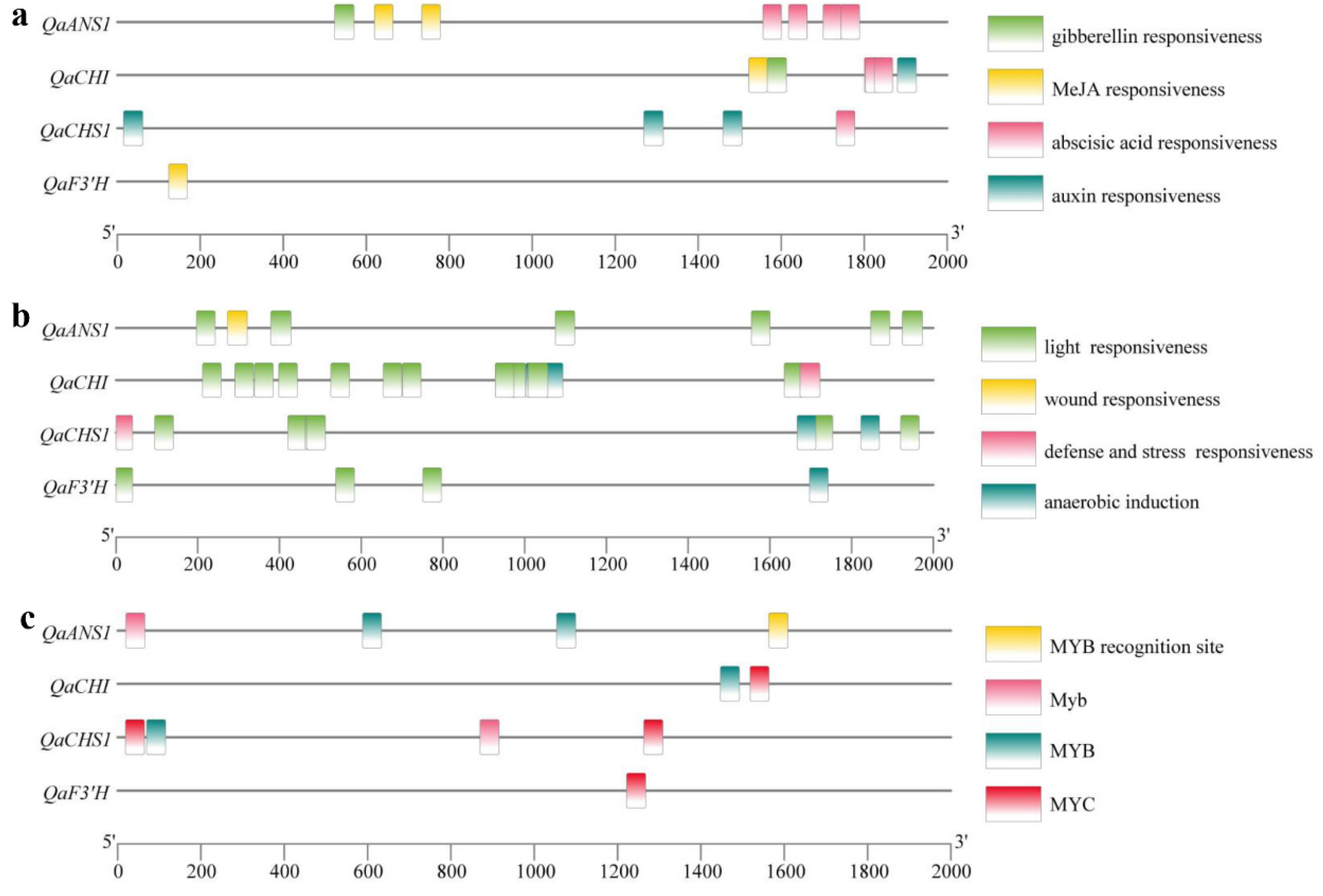
2.3. Expression of Genes Associated with Anthocyanin Biosynthesis in Q. aliena
2.4. Homology Alignment and Phylogenetic Analysis of MYB Transcription Factors
3. Discussion
3.1. Anthocyanin Accumulation Causes Reddening of Leaves in Q. aliena Variants
3.2. Changes in Anthocyanin Biosynthesis Alter Leaf Color in Q. aliena Variants
3.3. QaMYB1 Probably Regulates Anthocyanin Biosynthesis in Q. aliena
4. Materials and Methods
4.1. Plant Material
4.2. Leaves Phenotypic Analysis
4.3. Analysis of Simple Sequence Repeat Markers
4.4. Measurement of Chlorophyll Fluorescence
4.5. Quantification of Total Anthocyanins, Total Carotenoids, Chlorophyll a, and Chlorophyll b
4.6. Cloning of Genes Associated with Anthocyanin Biosynthesis
4.7. Semi-Quantitative and Quantitative Gene Expression Analyses
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shi, Q.; Zhou, L.; Li, K.; Wang, Y. Advances in transcriptional regulation of anthocyanin synthesis in plants. For. Sci. Res. 2015, 28, 570–576. [Google Scholar] [CrossRef]
- Boldt, J.; Erwin, J.E.; Meyer, M.H. Foliar Anthocyanins: A Horticultural Review. Hortic. Rev. 2014, 42, 209–252. [Google Scholar] [CrossRef]
- Karageorgou, P.; Manetas, Y. The importance of being red when young: Anthocyanins and the protection of young leaves of Quercus coccifera from insect herbivory and excess light. Tree Physiol. 2006, 26, 613–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabrita, L.; Fossen, T.; MAndersen, Ø. Colour and stability of the six common anthocyanidin 3-glucosides in aqueous solutions. Food Chem. 2000, 68, 101–107. [Google Scholar] [CrossRef]
- Grotewold, E. The genetics and biochemistry of floral pigments. Annu. Rev. Plant Biol. 2006, 57, 761–780. [Google Scholar] [CrossRef]
- Holton, T.A.; Cornish, E.C. Genetics and Biochemistry of Anthocyanin Biosynthesis. Plant Cell 1995, 7, 1071–1083. [Google Scholar] [CrossRef]
- Shi, M.-Z.; Xie, D.-Y. Biosynthesis and Metabolic Engineering of Anthocyanins in Arabidopsis thaliana. Recent Pat. Biotechnol. 2014, 8, 47–60. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.-S.; Nguyen, V.P.; Jeon, H.-W.; Kim, M.-H.; Eom, S.H.; Lim, Y.J.; Kim, W.-C.; Park, E.-J.; Choi, Y.-I.; Ko, J.-H. Overexpression of PtrMYB119, a R2R3-MYB transcription factor from Populus trichocarpa, promotes anthocyanin production in hybrid poplar. Tree Physiol. 2016, 36, 1162–1176. [Google Scholar] [CrossRef] [Green Version]
- An, J.; Zhang, X.; Liu, Y.; Wang, X.; You, C.; Hao, Y. ABI5 regulates ABA-induced anthocyanin biosynthesis by modulating the MYB1-bHLH3 complex in apple. J. Exp. Bot. 2020, 72, 1460–1472. [Google Scholar] [CrossRef]
- Azuma, A.; Yakushiji, H.; Koshita, Y.; Kobayashi, S. Flavonoid biosynthesis-related genes in grape skin are differentially regulated by temperature and light conditions. Planta 2012, 236, 1067–1080. [Google Scholar] [CrossRef]
- Schaart, J.G.; Dubos, C.; De La Fuente, I.R.; van Houwelingen, A.M.M.L.; de Vos, R.C.H.; Jonker, H.H.; Xu, W.; Routaboul, J.-M.; Lepiniec, L.; Bovy, A.G. Identification and characterization of MYB-bHLH-WD40 regulatory complexes controlling proanthocyanidin biosynthesis in strawberry (Fragaria × ananassa) fruits. New Phytol. 2013, 197, 454–467. [Google Scholar] [CrossRef] [PubMed]
- Jian, W.; Cao, H.; Yuan, S.; Liu, Y.; Lu, J.; Lu, W.; Li, N.; Wang, J.; Zou, J.; Tang, N.; et al. SlMYB75, an MYB-type transcription factor, promotes anthocyanin accumulation and enhances volatile aroma production in tomato fruits. Hortic. Res. 2019, 6, 15. [Google Scholar] [CrossRef] [Green Version]
- Jia, N.; Wang, J.; Wang, Y.; Ye, W.; Liu, J.; Jiang, J.; Sun, J.; Yan, P.; Wang, P.; Wang, F.; et al. The Light-Induced WD40-Repeat Transcription Factor DcTTG1 Regulates Anthocyanin Biosynthesis in Dendrobium candidum. Front. Plant Sci. 2021, 12, 633333. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.-H.; Chu, F.-H. A R2R3-MYB Gene LfMYB113 is Responsible for Autumn Leaf Coloration in Formosan sweet gum (Liquidambar formosana Hance). Plant Cell Physiol. 2017, 58, 508–521. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Zhang, Q.; Sun, Y.; Yang, L.; Wang, Z. Genome-wide identification and characterization of R2R3-MYB family in Hypericum perforatum under diverse abiotic stresses. Int. J. Biol. Macromol. 2019, 145, 341–354. [Google Scholar] [CrossRef]
- Hu, D.-G.; Sun, C.-H.; Zhang, Q.-Y.; Gu, K.-D.; Hao, Y.-J. The basic helix-loop-helix transcription factor MdbHLH3 modulates leaf senescence in apple via the regulation of dehydratase-enolase-phosphatase complex 1. Hortic. Res. 2020, 7, 50. [Google Scholar] [CrossRef] [Green Version]
- Wen, C.-H.; Lin, S.-S.; Chu, F.-H. Transcriptome Analysis of a Subtropical Deciduous Tree: Autumn Leaf Senescence Gene Expression Profile of Formosan Gum. Plant Cell Physiol. 2015, 56, 163–174. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Gu, M.; Deng, M. In Vitro Propagation of Oriental White Oak Quercus aliena Blume. Forests 2019, 10, 463. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Li, Y.; Song, J.; Zhang, R.; Yan, Y.; Wang, Y.; Du, F.K. Geometric morphometric analyses of leaf shapes in two sympatric Chinese oaks: Quercus dentata Thunberg and Quercus aliena Blume (Fagaceae). Ann. For. Sci. 2018, 75, 90. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Hou, M.; Du, F.K.; Li, J.; Yin, K. Complete chloroplast genome of the Oriental white oak: Quercus aliena Blume. Mitochondrial Dna 2015, 27, 2802–2804. [Google Scholar] [CrossRef]
- Jin, Y.-S.; Heo, S.-I.; Lee, M.-J.; Rhee, H.-I.; Wang, M.-H. Free radical scavenging and hepatoprotective actions of Quercus aliena acorn extract against CCl4-induced liver. Free Radic. Res. 2005, 39, 1351–1358. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q. Quercus aliena Bl. Seedling and silviculture techniques. Mod. Hortic. 2010, 11, 45. [Google Scholar] [CrossRef]
- Xu, Z.-S.; Yang, Q.-Q.; Kai, F.; Yu, X.; Xiong, A.-S. DcMYB113, a root-specific R2R3-MYB, conditions anthocyanin biosynthesis and modification in carrot. Wiley-Blackwell Online Open 2020, 18, 1585–1597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laura, M.-P.; Guadalupe, C.-L.; Francisco, A.-R.; Thomas, H.; Ludwig, R.; Antonio, R.-F.; Luis, C.J.; Wilfried, S.; Juan, M.-B.; Rosario, B.-P. MYB10 plays a major role in the regulation of flavonoid/phenylpropanoid metabolism during ripening of Fragaria × ananassa fruits. J. Exp. Bot. 2013, 65, 401. [Google Scholar] [CrossRef] [Green Version]
- An, J.; Xu, R.; Liu, X.; Zhang, J.; Wang, X.; You, C.; Hao, Y. Jasmonate induces biosynthesis of anthocyanin and proanthocyanidin in apple by mediating the JAZ1–TRB1–MYB9 complex. Plant J. 2021, 106, 1414–1430. [Google Scholar] [CrossRef]
- Iwashina, T. Contribution to flower colors of flavonoids including anthocyanins: A review. Nat. Prod. Commun. 2015, 10, 529–544. [Google Scholar] [CrossRef] [Green Version]
- Winkel-Shirley, B. Flavonoid Biosynthesis. A Colorful Model for Genetics, Biochemistry, Cell Biology, and Biotechnology. Plant Physiol. 2001, 126, 485–493. [Google Scholar] [CrossRef] [Green Version]
- Hu, R.; He, Y.; He, Z.; Wu, S. Progress in studies on absorption, metabolism and physiological functions of centaurea -3- glucoside. Chin. J. Anim. Nutr. 2019, 31, 2052–2062. [Google Scholar] [CrossRef]
- Tallis, M.J.; Lin, Y.; Rogers, A.; Zhang, J.; Street, N.R.; Miglietta, F.; Karnosky, D.F.; Angelis, P.D.; Calfapietra, C.; Taylor, G. The transcriptome of Populus in elevated CO2 reveals increased anthocyanin biosynthesis during delayed autumnal senescence. New Phytol. 2010, 186, 415–428. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.; O’Keefe, J.; MicheleHolbrook, N.; Feild, T. Pigment dynamics and autumn leaf senescence in a New England deciduous forest, eastern USA. Ecol. Res. 2003, 18, 677–694. [Google Scholar] [CrossRef]
- Hoch, W.A.; Singsaas, E.L.; McCown, B.H. Resorption Protection. Anthocyanins Facilitate Nutrient Recovery in Autumn by Shielding Leaves from Potentially Damaging Light Levels. Plant Physiol. 2003, 133, 1296–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.-G.; Long, S.P.; Ort, D.R. What is the maximum efficiency with which photosynthesis can convert solar energy into biomass? Curr. Opin. Biotechnol. 2008, 19, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Rachel, A.; Peter, R. Early Autumn Senescence in Red Maple (Acer rubrum L.) Is Associated with High Leaf Anthocyanin Content. Plants 2015, 4, 505–522. [Google Scholar] [CrossRef] [Green Version]
- He, Q.; Ren, Y.; Zhao, W.; Li, R.; Zhang, L. Low Temperature Promotes Anthocyanin Biosynthesis and Related Gene Expression in the Seedlings of Purple Head Chinese Cabbage (Brassica rapa L.). Genes 2020, 11, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, T.; Li, K.; Wu, T.; Wang, Y.; Zhang, X.; Xu, X.; Yao, Y.; Han, Z. Identification of new regulators through transcriptome analysis that regulate anthocyanin biosynthesis in apple leaves at low temperatures. PLoS ONE 2019, 14, e0210672. [Google Scholar] [CrossRef] [PubMed]
- Gao-Takaia, M.; Katayama-Ikegamia, A.; Matsudab, K.; Shindoa, H.; Uemaea, S.; Oyaizua, M. A low temperature promotes anthocyanin biosynthesis but does not accelerate endogenous abscisic acid accumulation in red-skinned grapes. Plant Sci. 2019, 283, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Liu, Z.; Wu, Y.; Zheng, L.; Zhang, G. Regulatory Mechanisms of Anthocyanin Biosynthesis in Apple and Pear. Int. J. Mol. Sci. 2021, 22, 8441. [Google Scholar] [CrossRef]
- Yu, R.; Du, X.; Li, J.; Liu, L.; Hu, C.; Yan, X.; Xia, Y.; Xu, H. Identification and differential expression analysis of anthocyanin biosynthetic genes in root-skin color variants of radish (Raphanus sativus L.). Genes Genom. 2020, 42, 413–424. [Google Scholar] [CrossRef]
- Muleke, E.M.M.; Fan, L.; Wang, Y.; Xu, L.; Zhu, X.; Zhang, W.; Cao, Y.; Karanja, K.B.; Liu, L. Coordinated Regulation of Anthocyanin Biosynthesis Genes Confers Varied Phenotypic and Spatial-Temporal Anthocyanin Accumulation in Radish (Raphanus sativus L.). Front. Plant Sci. 2017, 8, 1243. [Google Scholar] [CrossRef] [Green Version]
- Aza-González, C.; Herrera-Isidrón, L.; Núez-Palenius, H.G.; Vega, O.M.D.L.; Ochoa-Alejo, N. Anthocyanin accumulation and expression analysis of biosynthesis-related genes during chili pepper fruit development. Biol. Plant. 2012, 57, 49–55. [Google Scholar] [CrossRef]
- Ben-Yehudah, G.; Korchinsky, R.; Redel, G.; Ovadya, R.; Cohen, Y. Colour accumulation patterns and the anthocyanin biosynthetic pathway in ‘Red Delicious’ apple variants. J. Pomol. Hortic. Sci. 2005, 80, 187–192. [Google Scholar] [CrossRef]
- Chen, X.; Liu, H.; Wang, S.; Zhang, C.; Zhang, J. Combined Transcriptome and Proteome Analysis Provides Insights into Anthocyanin Accumulation in the Leaves of Red-Leaved Poplars. Plant Mol. Biol. 2021, 106, 491–503. [Google Scholar] [CrossRef]
- Song, X.; Yang, Q.; Liu, Y.; Li, J.; Chang, X.; Xian, L.; Zhang, J. Genome-wide identification of Pistacia R2R3-MYB gene family and function characterization of PcMYB113 during autumn leaf coloration in Pistacia chinensis. Int. J. Biol. Macromol. 2021, 192, 16–27. [Google Scholar] [CrossRef]
- Urao, T.; Yamaguchi-Shinozaki, K.; Urao, S.; Shinozaki, K. An Arabidopsis myb homolog is induced by dehydration stress and its gene product binds to the conserved MYB recognition sequence. Plant Cell 1993, 5, 1529–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, H.; Urao, T.; Ito, T.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in abscisic acid signaling. Plant Cell 2003, 15, 63–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solano, R.; Nieto, C.; Avila, J.; Cañas, L.; Diaz, I.; Paz-Ares, J. Dual DNA binding specificity of a petal epidermis-specific MYB transcription factor (MYB.Ph3) from Petunia hybrida. EMBO J. 1995, 14, 1773–1784. [Google Scholar] [CrossRef]
- Chinnusamy, V.; Schumaker, K.; Zhu, J.K. Molecular genetic perspectives on cross-talk and specificity in abiotic stress signalling in plants. J. Exp. Bot. 2004, 55, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Naing, A.H.; Kim, C.K. Roles of R2R3-MYB transcription factors in transcriptional regulation of anthocyanin biosynthesis in horticultural plants. Plant Mol. Biol. 2018, 98, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Poudel, P.; Azuma, A.; Kobayashi, S.; Koyama, K.; Goto-Yamamoto, N. VvMYBAs induce expression of a series of anthocyanin biosynthetic pathway genes in red grapes (Vitis vinifera L.). Sci. Hortic. 2021, 283, 110304. [Google Scholar] [CrossRef]
- Kobayashi, S.; Ishimaru, M.; Hiraoka, K.; Honda, C. Myb-related genes of the Kyoho grape (Vitis labruscana) regulate anthocyanin biosynthesis. Planta 2002, 215, 924–933. [Google Scholar] [CrossRef] [PubMed]
- Telias, A.; Lin-Wang, K.; Stevenson, D.E.; Cooney, J.M.; Bradeen, J.M. Apple skin patterning is associated with differential expression of MYB10. BMC Plant Biol. 2011, 11, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.; Grain, D.; Bobet, S.; Gourrierec, J.L.; Thévenin, J.; Kelemen, Z.; Lepiniec, L.; Dubos, C. Complexity and robustness of the flavonoid transcriptional regulatory network revealed by comprehensive analyses of MYB-bHLH-WDR complexes and their targets in Arabidopsis seed. New Phytol. 2014, 202, 15–18. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.; Zhao, M.; Leavitt, J.M.; Lloyd, A.M. Regulation of the anthocyanin biosynthetic pathway by the TTG1/bHLH/Myb transcriptional complex in Arabidopsis seedlings. Plant J. 2010, 53, 814–827. [Google Scholar] [CrossRef]
- Chen, G.; Pengbo, X.; Pan, J.; Li, Y.; Zhou, J.; Kuang, H.; Lian, H. Inhibition of FvMYB10 transcriptional activity promotes color loss in strawberry fruit. Plant Sci. 2020, 298, 110578. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, M.; Jun, N.; Hou, J.; Shu, X.; Zhao, W.; Su, P.; Wang, D.; Shah, F.A.; Huang, S.; et al. The R2R3-MYB transcription factor SsMYB1 positively regulates anthocyanin biosynthesis and determines leaf color in Chinese tallow (Sapium sebiferum Roxb.). Ind. Crop. Prod. 2021, 164, 113335. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, K.; Qi, Y.; Lv, G.; Ren, X.; Liu, Z.; Ma, F. Transcriptional Regulation of Anthocyanin Synthesis by MYB-bHLH-WDR Complexes in Kiwifruit (Actinidia chinensis). J. Agric. Food Chem. 2021, 69, 3677–3691. [Google Scholar] [CrossRef]
- Castillejo, C.; Waurich, V.; Wagner, H.; Ramos, R.; Oiza, N.; Muñoz, P.; Triviño, J.C.; Caruana, J.; Liu, Z.; Cobo, N.; et al. Allelic Variation of MYB10 Is the Major Force Controlling Natural Variation in Skin and Flesh Color in Strawberry (Fragaria spp.) Fruit. Plant Cell 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yang, Y.; Li, H.; Wang, W.; Zheng, H.; Tao, J. Genome-Wide Identification of Glutathione S-Transferase and Expression Analysis in Response to Anthocyanin Transport in the Flesh of the New Teinturier Grape Germplasm ‘Zhongshan-HongYu’. Int. J. Mol. Sci. 2022, 32, 7717. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Dong, W.; Zhu, Y.; Allan, A.C.; Lin-Wang, K.; Xu, C. PpGST1, an anthocyanin-related glutathione S-transferase gene, is essential for fruit coloration in peach. Plant Biotechnol. J. 2020, 18, 1284–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Qi, Y.; Zhang, A.; Wu, H.; Liu, Z.; Ren, X. Molecular cloning and functional characterization of AcGST1, an anthocyanin-related glutathione S-transferase gene in kiwifruit (Actinidia chinensis). Plant Mol. Biol. 2019, 100, 451–465. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Cao, H.; Pan, L.; Niu, L.; Wei, B.; Cui, G.; Wang, L.; Yao, J.L.; Zeng, W.; Wang, Z. Two loss-of-function alleles of the glutathione S-transferase (GST) gene cause anthocyanin deficiency in flower and fruit skin of peach (Prunus persica). Plant J. Cell Mol. Biol. 2021, 107, 1320–1331. [Google Scholar] [CrossRef] [PubMed]
- El-Sharkawy, I.; Liang, D.; Xu, K. Transcriptome analysis of an apple (Malus × domestica) yellow fruit somatic mutation identifies a gene network module highly associated with anthocyanin and epigenetic regulation. J. Exp. Bot. 2015, 66, 7359–7376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y. Natural Hybridization between Quercus dentata and Q. mongolica-liaotungensis Revealed by Microsatellite Markers. Master’s Thesis, Shandong University, Shandong, China, 2012. [Google Scholar]
- Yang, X.; Gao, K.; Chen, Z.; Yang, X.; Rao, P.; Zhao, T.; An, X. Development and application of EST-SSR markers in Koelreuteria paniculata Laxm. using a transcriptomic approach. Biotechnology 2017, 16, 45–56. [Google Scholar] [CrossRef]
- Rohlf, F. NTSYS-pc: Numerical Taxonomy System Ver. 2.1; Exeter Publishing Ltd.: Setauket, NY, USA, 2002. [Google Scholar]
- Zhang, K.M.; Wang, J.W.; Guo, M.L.; Du, W.L.; Wang, X. Short-day signals are crucial for the induction of anthocyanin biosynthesis in Begonia semperflorens under low temperature condition. J. Plant Physiol. 2016, 204, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Analysis 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Gambino, G.; Perrone, I.; Gribaudo, I. A Rapid and effective method for RNA extraction from different tissues of grapevine and other woody plants. Phytochem. Anal. 2008, 19, 520–525. [Google Scholar] [CrossRef] [PubMed]
- Plomion, C.; Aury, J.-M.; Amselem, J.; Leroy, T.; Murat, F.; Duplessis, S.; Faye, S.; Francillonne, N.; Labadie, K.; Le Provost, G.; et al. Oak genome reveals facets of long lifespan. Nat. Plants 2018, 4, 440–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, E.L.; Ruskin, P.E. MEGA: A Computer Assisted Educational Tool for Multidisciplinary Education in Geriatrics and Aging. Gerontol. Geriatr. Educ. 1997, 18, 23–31. [Google Scholar] [CrossRef]
- Pérez, M.; Cañal, M.J.; Toorop, P.E. Expression analysis of epigenetic and abscisic acid-related genes during maturation of Quercus suber somatic embryos. Plant Cell Tissue Organ Cult. (PCTOC) 2015, 121, 353–366. [Google Scholar] [CrossRef]
- Mallón, R.; Valladares, S.; Corredoira, E.; Vieitez, A.; Vidal, N. Overexpression of the chestnut CsTL1 gene coding for a thaumatin-like protein in somatic embryos of Quercus robur. Plant Cell Tissue Organ Cult. (PCTOC) 2014, 116, 141–151. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Class | Compound | Molecular Weight (g/mol) | QAC | QA1 | QA2 | QA3 |
|---|---|---|---|---|---|---|
| Cyanidin | Cyanidin 3-O-glucoside | 449.111 | 1.03 | 777.96 | 662.40 | 583.98 |
| Cyanidin | Cyanidin 3-O-sambubioside | 581.15 | 0.03 | 76.49 | 57.03 | 10.56 |
| Delphinidin | Delphinidin 3-O-glucoside | 465.10 | 0.35 | 3.86 | 3.03 | 36.74 |
| Procyanidin | Procyanidin B1 | 578.14 | 49.52 | 191.28 | 195.87 | 130.92 |
| Procyanidin | Procyanidin B3 | 578.14 | 133.55 | 521.95 | 481.77 | 229.01 |
| Flavonoid | Quercetin 3-O-glucoside | 464.10 | 543.53 | 796.13 | 418.63 | 439.02 |
| Gene | S2 Stage | S3 Stage | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| ACs | QaMYB1 | QaMYB2 | QaMYB3 | QaWD40 | ACs | QaMYB1 | QaMYB2 | QaMYB3 | QaWD40 | |
| QaPAL1 | 0.992 ** | 1.000 ** | 0.721 | 1.000 ** | 1.000 ** | 0.707 | 0.952 * | 0.865 | 0.980 * | 0.981 * |
| QaPAL2 | 0.894 | 0.872 | 0.509 | 0.863 | 0.861 | 0.799 | 0.953 * | 0.729 | 0.871 | 0.908 |
| Qa4CL1 | −0.556 | −0.511 | −0.959 * | −0.509 | −0.511 | −0.882 | −0.990 ** | −0.657 | −0.911 | −0.852 |
| Qa4CL2 | 0.940 | 0.941 | 0.518 | 0.936 | 0.935 | 0.782 | 0.969 * | 0.785 | 0.926 | 0.944 |
| QaCHS1 | 0.880 | 0.852 | 0.516 | 0.841 | 0.839 | 0.998 ** | 0.905 | 0.293 | 0.651 | 0.585 |
| QaCHS2 | 0.992 ** | 1.000 ** | 0.711 | 1.000 ** | 1.000 ** | 0.745 | 0.958 * | 0.809 | 0.982 * | 0.935 |
| QaCHI | 0.970 * | 0.952 * | 0.666 | 0.945 | 0.944 | 0.846 | 0.964 * | 0.674 | 0.850 | 0.876 |
| QaF3H | 0.992 ** | 0.999 ** | 0.702 | 0.999 ** | 0.999 ** | 0.719 | 0.934 | 0.818 | 0.914 | 0.955 * |
| QaF3′H | 0.946 | 0.935 | 0.569 | 0.929 | 0.927 | 0.961 * | 0.822 | 0.169 | 0.570 | 0.455 |
| QaDFR1 | 0.993 ** | 1.000 ** | 0.714 | 1.000 ** | 1.000 ** | 0.721 | 0.943 | 0.812 | 0.983 * | 0.927 |
| QaDFR2 | 0.993 ** | 1.000 ** | 0.722 | 1.000 ** | 1.000 ** | 0.601 | 0.900 | 0.924 | 0.996 ** | 0.991 ** |
| QaDFR3 | 0.993 ** | 1.000 ** | 0.729 | 1.000 ** | 1.000 ** | 0.549 | 0.853 | 0.899 | 0.988 * | 0.941 |
| QaANS1 | 0.971 * | 0.964 * | 0.619 | 0.959 * | 0.958 * | 0.946 | 0.988 * | 0.551 | 0.838 | 0.789 |
| QaUFGT1 | 0.992 ** | 0.998 ** | 0.688 | 0.997 ** | 0.997 ** | 0.497 | 0.826 | 0.928 | 0.986 * | 0.950 * |
| QaWD40 | 0.991 ** | 1.000 ** | 0.723 | 1.000 ** | 0.557 | 0.874 | 0.944 | 0.976 * | ||
| QaMYB3 | 0.992 ** | 1.000 ** | 0.722 | 0.617 | 0.905 | 0.905 | ||||
| QaMYB2 | 0.766 | 0.725 | 0.257 | 0.671 | ||||||
| QaMYB1 | 0.994 ** | 0.889 | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Yang, N.; Zhang, Q.; Pei, Z.; Chang, M.; Zhou, H.; Ge, Y.; Yang, Q.; Li, G. Anthocyanin Biosynthesis Associated with Natural Variation in Autumn Leaf Coloration in Quercus aliena Accessions. Int. J. Mol. Sci. 2022, 23, 12179. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232012179
Yang X, Yang N, Zhang Q, Pei Z, Chang M, Zhou H, Ge Y, Yang Q, Li G. Anthocyanin Biosynthesis Associated with Natural Variation in Autumn Leaf Coloration in Quercus aliena Accessions. International Journal of Molecular Sciences. 2022; 23(20):12179. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232012179
Chicago/Turabian StyleYang, Xiong, Ning Yang, Qian Zhang, Ziqi Pei, Muxi Chang, Huirong Zhou, Yaoyao Ge, Qinsong Yang, and Guolei Li. 2022. "Anthocyanin Biosynthesis Associated with Natural Variation in Autumn Leaf Coloration in Quercus aliena Accessions" International Journal of Molecular Sciences 23, no. 20: 12179. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232012179