
Binding of Gamma-Glutamyl Transferase to TLR4 Signalling Allows Tissue Factor Activation in Monocytes
 , , , , , and
, , , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
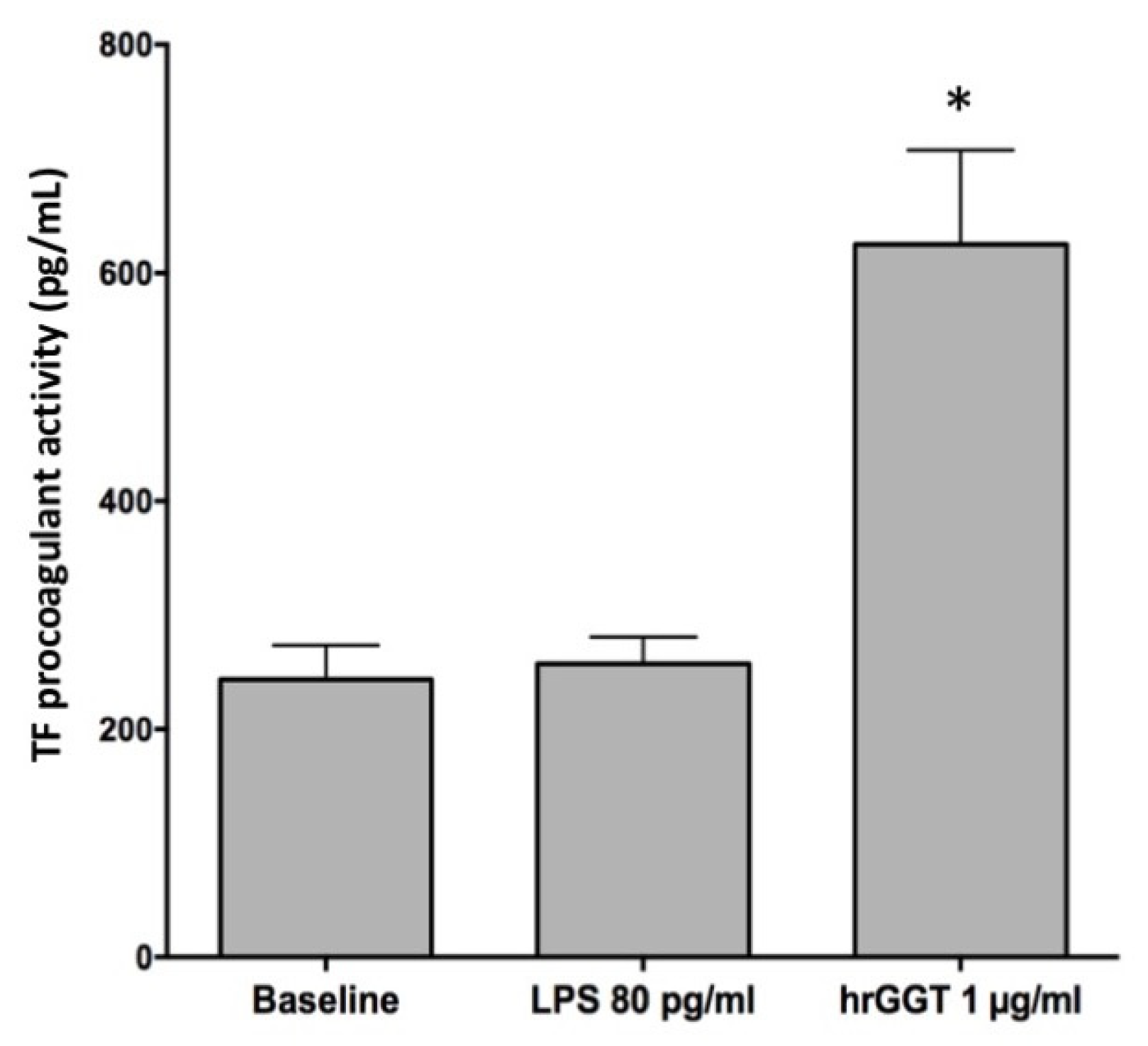
2.1. hrGGT Stimulates TF Activity in PBMCs and THP-1 Cell Lines
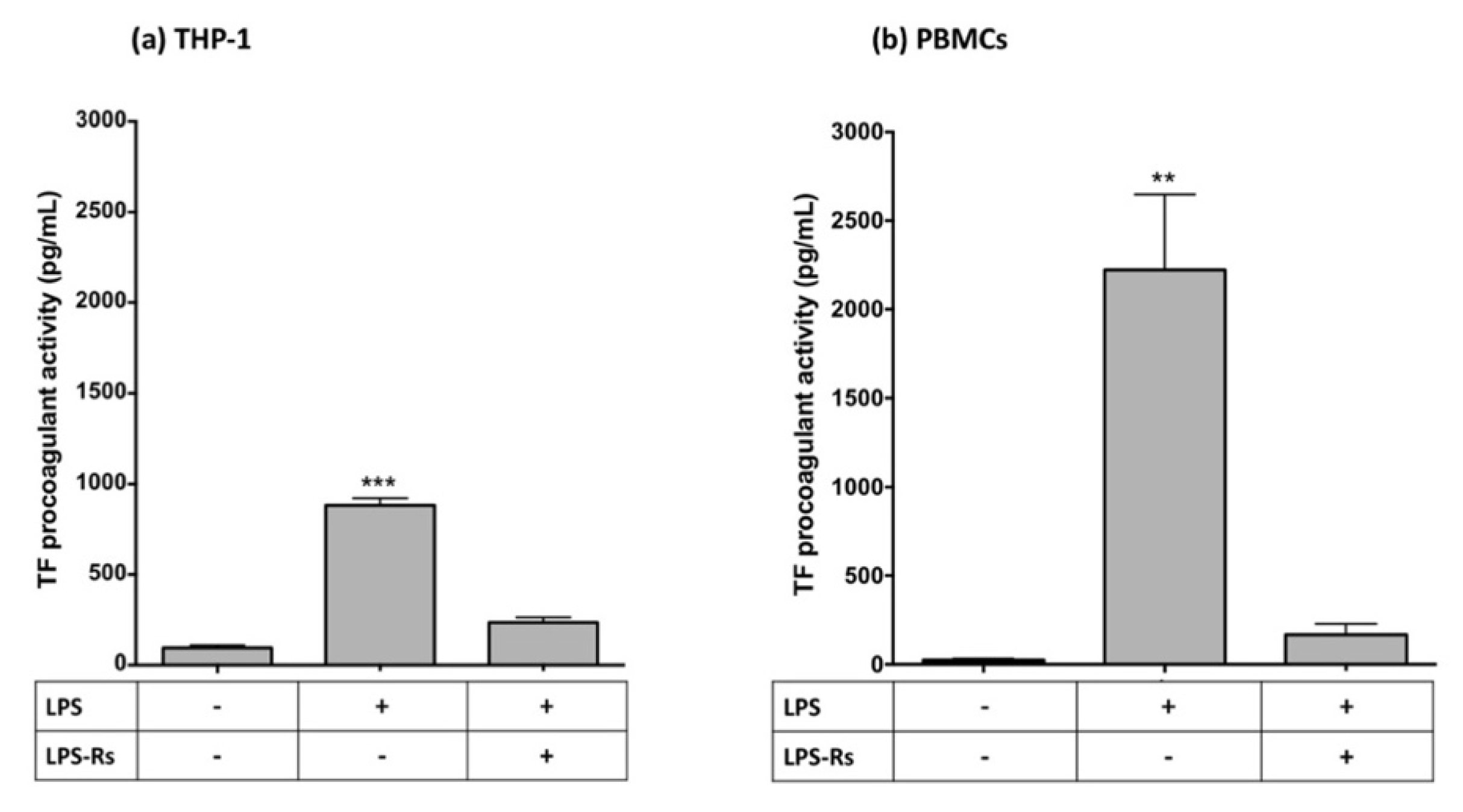
2.2. LPS Contamination Does Not Explain GGT-Induced TF Activity
2.3. Specificity of TF Procoagulant Activity
2.4. Cell Vitality: MTT Assay on THP-1 Cell Lines
2.5. Inhibition of the TLR4/NF-κB Pathway Downregulates LPS- and hrGGT-Induced TF Procoagulant Activity
2.6. Activation by hrGGT of NF-κB through TLR4 Signaling in HEK-Blue Cells
3. Discussion
3.1. GGT Upregulates Monocytic TF Expression
3.2. hrGGT Activates TLR4/NF-κB Signaling
4. Materials and Methods
4.1. Reagent and Materials
4.2. Cell Culture
4.2.1. PBMCs Preparations
4.2.2. THP-1 Cell Line as a Monocyte Response Model
4.2.3. HEK-Blue Cells
4.3. TF Procoagulant Activity
4.4. Western Blot Analysis
4.5. Evaluation of Endotoxin Contamination by a Limulus Amoebocyte Lysate (LAL) Assay
4.6. TLR4/NF-κB Pathway in hrGGT-Induced TF-Procoagulant Activity
4.7. Cell Vitality: Dimethyl Thiazolyl Diphenyl Tetrazolium (MTT) Assay
4.8. NF-κB Reporter Assay
4.9. Statistic Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Castonguay, R.; Halim, D.; Morin, M.; Furtos, A.; Lherbet, C.; Bonneil, E.; Thibault, P.; Keillor, J.W. Kinetic characterization and identification of the acylation and glycosylation sites of recombinant human gamma-glutamyltranspeptidase. Biochemistry 2007, 46, 12253–12262. [Google Scholar] [CrossRef]
- Koenig, G.; Seneff, S. Gamma-Glutamyltransferase: A Predictive Biomarker of Cellular Antioxidant Inadequacy and Disease Risk. Dis. Markers 2015, 2015, 818570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, S.; Jiang, D.; Tao, Y. Role of gamma-glutamyltransferase in cardiovascular diseases. Exp. Clin. Cardiol. 2013, 18, 53–56. [Google Scholar] [PubMed]
- Ruttmann, E.; Brant, L.J.; Concin, H.; Diem, G.; Rapp, K.; Ulmer, H.; Group, V.H.M.a.P.P.S. Gamma-glutamyltransferase as a risk factor for cardiovascular disease mortality: An epidemiological investigation in a cohort of 163,944 Austrian adults. Circulation 2005, 112, 2130–2137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ndrepepa, G.; Kastrati, A. Gamma-glutamyl transferase and cardiovascular disease. Ann. Transl. Med. 2016, 4, 481. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.; Aonuma, K. Gamma-Glutamyl Transferase as a Risk Biomarker of Cardiovascular Disease—Does It Have Another Face? Circ. J. 2017, 81, 783–785. [Google Scholar] [CrossRef] [Green Version]
- Franzini, M.; Corti, A.; Martinelli, B.; Del Corso, A.; Emdin, M.; Parenti, G.F.; Glauber, M.; Pompella, A.; Paolicchi, A. Gamma-glutamyltransferase activity in human atherosclerotic plaques—Biochemical similarities with the circulating enzyme. Atherosclerosis 2009, 202, 119–127. [Google Scholar] [CrossRef]
- Corti, A.; Belcastro, E.; Dominici, S.; Maellaro, E.; Pompella, A. The dark side of gamma-glutamyltransferase (GGT): Pathogenic effects of an ‘antioxidant’ enzyme. Free Radic. Biol. Med. 2020, 160, 807–819. [Google Scholar] [CrossRef] [PubMed]
- Paolicchi, A.; Minotti, G.; Tonarelli, P.; Tongiani, R.; De Cesare, D.; Mezzetti, A.; Dominici, S.; Comporti, M.; Pompella, A. Gamma-glutamyl transpeptidase-dependent iron reduction and LDL oxidation—A potential mechanism in atherosclerosis. J. Investig. Med. 1999, 47, 151–160. [Google Scholar] [PubMed]
- Ndrepepa, G.; Colleran, R.; Kastrati, A. Gamma-glutamyl transferase and the risk of atherosclerosis and coronary heart disease. Clin. Chim. Acta 2018, 476, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Paolicchi, A.; Emdin, M.; Ghliozeni, E.; Ciancia, E.; Passino, C.; Popoff, G.; Pompella, A. Images in cardiovascular medicine. Human atherosclerotic plaques contain gamma-glutamyl transpeptidase enzyme activity. Circulation 2004, 109, 1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pucci, A.; Franzini, M.; Matteucci, M.; Ceragioli, S.; Marconi, M.; Ferrari, M.; Passino, C.; Basolo, F.; Emdin, M.; Paolicchi, A. b-Gamma-glutamyltransferase activity in human vulnerable carotid plaques. Atherosclerosis 2014, 237, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Scalise, V.; Balia, C.; Cianchetti, S.; Neri, T.; Carnicelli, V.; Zucchi, R.; Franzini, M.; Corti, A.; Paolicchi, A.; Celi, A.; et al. Non enzymatic upregulation of tissue factor expression by gamma-glutamyl transferase in human peripheral blood mononuclear cells. Thromb. J. 2016, 14, 45. [Google Scholar] [CrossRef] [Green Version]
- West, M.B.; Wickham, S.; Quinalty, L.M.; Pavlovicz, R.E.; Li, C.; Hanigan, M.H. Autocatalytic cleavage of human gamma-glutamyl transpeptidase is highly dependent on N-glycosylation at asparagine 95. J. Biol. Chem. 2011, 286, 28876–28888. [Google Scholar] [CrossRef] [Green Version]
- West, M.B.; Chen, Y.; Wickham, S.; Heroux, A.; Cahill, K.; Hanigan, M.H.; Mooers, B.H. Novel insights into eukaryotic γ-glutamyltranspeptidase 1 from the crystal structure of the glutamate-bound human enzyme. J. Biol. Chem. 2013, 288, 31902–31913. [Google Scholar] [CrossRef] [Green Version]
- Mechiche, H.; Cornillet-Lefebvre, P.; Nguyen, P. A subpopulation of human B lymphocytes can express a functional Tissue Factor in response to phorbol myristate acetate. Thromb. Haemost. 2005, 94, 146–154. [Google Scholar] [CrossRef]
- Wilcox, J.N.; Smith, K.M.; Schwartz, S.M.; Gordon, D. Localization of tissue factor in the normal vessel wall and in the atherosclerotic plaque. Proc. Natl. Acad. Sci. USA 1989, 86, 2839–2843. [Google Scholar] [CrossRef] [Green Version]
- Belcastro, E.; Franzini, M.; Cianchetti, S.; Lorenzini, E.; Masotti, S.; Fierabracci, V.; Pucci, A.; Pompella, A.; Corti, A. Monocytes/macrophages activation contributes to b-gamma-glutamyltransferase accumulation inside atherosclerotic plaques. J. Transl. Med. 2015, 13, 325. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.D.; Ye, B.Q.; Zheng, S.X.; Wang, J.T.; Wang, J.G.; Chen, M.; Liu, J.G.; Pei, X.H.; Wang, L.J.; Lin, Z.X.; et al. NF-kappaB transcription factor p50 critically regulates tissue factor in deep vein thrombosis. J. Biol. Chem. 2009, 284, 4473–4483. [Google Scholar] [CrossRef]
- Riva, M.; Källberg, E.; Björk, P.; Hancz, D.; Vogl, T.; Roth, J.; Ivars, F.; Leanderson, T. Induction of nuclear factor-κB responses by the S100A9 protein is Toll-like receptor-4-dependent. Immunology 2012, 137, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Deierborg, T.; Burguillos, M.A. A new “sweet” ligand for Toll-like receptor 4. Oncotarget 2015, 6, 19928–19929. [Google Scholar] [CrossRef]
- Bhattacharyya, S.; Varga, J. Endogenous ligands of TLR4 promote unresolving tissue fibrosis: Implications for systemic sclerosis and its targeted therapy. Immunol. Lett. 2018, 195, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Moriwaki, S.; Into, T.; Suzuki, K.; Miyauchi, M.; Takata, T.; Shibayama, K.; Niida, S. γ-Glutamyltranspeptidase is an endogenous activator of Toll-like receptor 4-mediated osteoclastogenesis. Sci. Rep. 2016, 6, 35930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bar-Shavit, Z. The osteoclast: A multinucleated, hematopoietic-origin, bone-resorbing osteoimmune cell. J. Cell Biochem. 2007, 102, 1130–1139. [Google Scholar] [CrossRef]
- García, M.G.; Alaniz, L.; Lopes, E.C.; Blanco, G.; Hajos, S.E.; Alvarez, E. Inhibition of NF-kappaB activity by BAY 11-7082 increases apoptosis in multidrug resistant leukemic T-cell lines. Leuk. Res. 2005, 29, 1425–1434. [Google Scholar] [CrossRef]
- Hiller, E.; Saal, J.G.; Ostendorf, P.; Griffiths, G.W. The procoagulant activity of human granulocytes, lymphocytes and monocytes stimulated by endotoxin. Coagulation and electron microscopic studies. Klin. Wochenschr. 1977, 55, 751–757. [Google Scholar] [CrossRef]
- Mészáros, K.; Aberle, S.; Dedrick, R.; Machovich, R.; Horwitz, A.; Birr, C.; Theofan, G.; Parent, J.B. Monocyte tissue factor induction by lipopolysaccharide (LPS): Dependence on LPS-binding protein and CD14, and inhibition by a recombinant fragment of bactericidal/permeability-increasing protein. Blood 1994, 83, 2516–2525. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Deng, J.; Chiu, M.; Chen, Y.X.; O’Brien, E.R. Heat shock protein 27 immune complex altered signaling and transport (ICAST): Novel mechanisms of attenuating inflammation. FASEB J. 2020, 34, 14287–14301. [Google Scholar] [CrossRef]
- Cines, D.B.; Zaitsev, S.; Rauova, L.; Rux, A.H.; Stepanova, V.; Krishnaswamy, S.; Sarkar, A.; Kowalska, M.A.; Zhao, G.; Mast, A.E.; et al. FcRn augments induction of tissue factor activity by IgG-containing immune complexes. Blood 2020, 135, 2085–2093. [Google Scholar] [CrossRef]
- Gogesch, P.; Dudek, S.; van Zandbergen, G.; Waibler, Z.; Anzaghe, M. The Role of Fc Receptors on the Effectiveness of Therapeutic Monoclonal Antibodies. Int. J. Mol. Sci. 2021, 22, 8947. [Google Scholar] [CrossRef] [PubMed]
- Ehret, J.; Zimmermann, M.; Eichhorn, T.; Zimmer, A. Impact of cell culture media additives on IgG glycosylation produced in Chinese hamster ovary cells. Biotechnol. Bioeng. 2019, 116, 816–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Sato, S.; Hemmi, H.; Hoshino, K.; Kaisho, T.; Sanjo, H.; Takeuchi, O.; Sugiyama, M.; Okabe, M.; Takeda, K.; et al. Role of adaptor TRIF in the MyD88-independent toll-like receptor signaling pathway. Science 2003, 301, 640–643. [Google Scholar] [CrossRef]
- Gaikwad, S.; Agrawal-Rajput, R. Lipopolysaccharide from Rhodobacter sphaeroides Attenuates Microglia-Mediated Inflammation and Phagocytosis and Directs Regulatory T Cell Response. Int. J. Inflam. 2015, 2015, 361326. [Google Scholar] [CrossRef] [Green Version]
- Kawamoto, T.; Ii, M.; Kitazaki, T.; Iizawa, Y.; Kimura, H. TAK-242 selectively suppresses Toll-like receptor 4-signaling mediated by the intracellular domain. Eur. J. Pharmacol. 2008, 584, 40–48. [Google Scholar] [CrossRef]
- Schwarz, H.; Schmittner, M.; Duschl, A.; Horejs-Hoeck, J. Residual endotoxin contaminations in recombinant proteins are sufficient to activate human CD1c+ dendritic cells. PLoS ONE 2014, 9, e113840. [Google Scholar] [CrossRef] [Green Version]
- Idriss, H.T.; Naismith, J.H. TNF alpha and the TNF receptor superfamily: Structure-function relationship(s). Microsc. Res. Tech. 2000, 50, 184–195. [Google Scholar] [CrossRef]
- Bosshart, H.; Heinzelmann, M. THP-1 cells as a model for human monocytes. Ann. Transl. Med. 2016, 4, 438. [Google Scholar] [CrossRef] [Green Version]
- Scalise, V.; Sanguinetti, C.; Neri, T.; Cianchetti, S.; Lai, M.; Carnicelli, V.; Celi, A.; Pedrinelli, R. PCSK9 Induces Tissue Factor Expression by Activation of TLR4/NFkB Signaling. Int. J. Mol. Sci. 2021, 22, 12640. [Google Scholar] [CrossRef]
- Kielkopf, C.L.; Bauer, W.; Urbatsch, I.L. Bradford Assay for Determining Protein Concentration. Cold Spring Harb. Protoc. 2020, 2020, 102269. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Wakelin, S.J.; Sabroe, I.; Gregory, C.D.; Poxton, I.R.; Forsythe, J.L.; Garden, O.J.; Howie, S.E. “Dirty little secrets”—Endotoxin contamination of recombinant proteins. Immunol. Lett. 2006, 106, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, G.K.; Roslansky, P.F.; Novitsky, T.J. Single-step, chromogenic Limulus amebocyte lysate assay for endotoxin. J. Clin. Microbiol. 1989, 27, 947–951. [Google Scholar] [CrossRef] [Green Version]
- Poelzl, G.; Eberl, C.; Achrainer, H.; Doerler, J.; Pachinger, O.; Frick, M.; Ulmer, H. Prevalence and prognostic significance of elevated gamma-glutamyltransferase in chronic heart failure. Circ. Heart Fail 2009, 2, 294–302. [Google Scholar] [CrossRef]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanguinetti, C.; Scalise, V.; Neri, T.; Celi, A.; Susini, V.; Franzini, M.; Pedrinelli, R. Binding of Gamma-Glutamyl Transferase to TLR4 Signalling Allows Tissue Factor Activation in Monocytes. Int. J. Mol. Sci. 2022, 23, 12207. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232012207
Sanguinetti C, Scalise V, Neri T, Celi A, Susini V, Franzini M, Pedrinelli R. Binding of Gamma-Glutamyl Transferase to TLR4 Signalling Allows Tissue Factor Activation in Monocytes. International Journal of Molecular Sciences. 2022; 23(20):12207. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232012207
Chicago/Turabian StyleSanguinetti, Chiara, Valentina Scalise, Tommaso Neri, Alessandro Celi, Vanessa Susini, Maria Franzini, and Roberto Pedrinelli. 2022. "Binding of Gamma-Glutamyl Transferase to TLR4 Signalling Allows Tissue Factor Activation in Monocytes" International Journal of Molecular Sciences 23, no. 20: 12207. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232012207