
Deodorized Garlic Decreases Oxidative Stress Caused by Lipopolysaccharide in Rat Heart through Hydrogen Sulfide: Preliminary Findings
 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Results
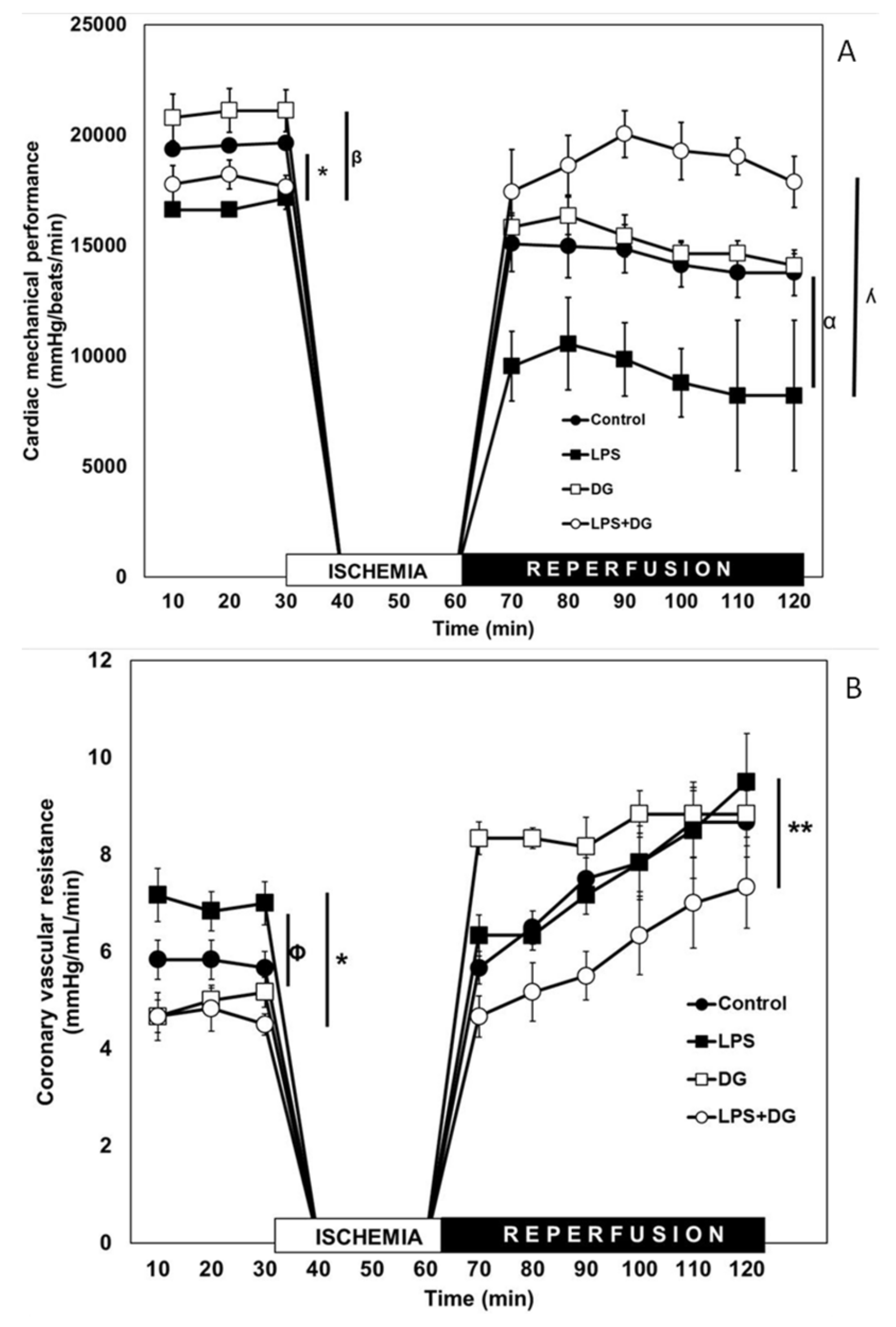
2.1. Cardiac Mechanical Performance (CMP)
2.2. Coronary Vascular Resistance (CVR)
2.3. Histological Analysis
2.4. Oxidative Stress Markers
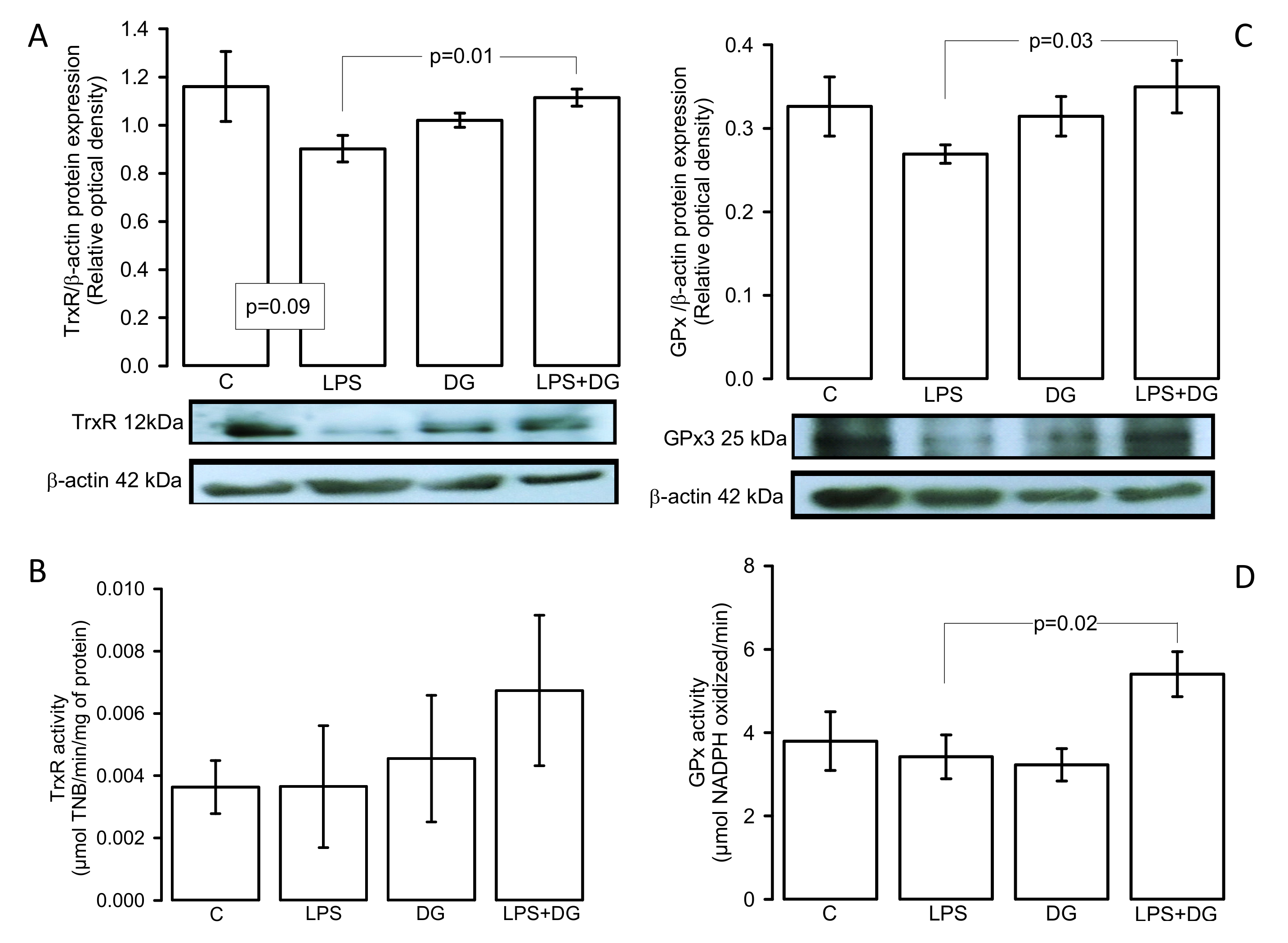
2.5. Enzymatic Activities and Expressions of the TxrR and GPx
2.6. Expressions of the CTH, CBS Enzymes and H2S Concentration
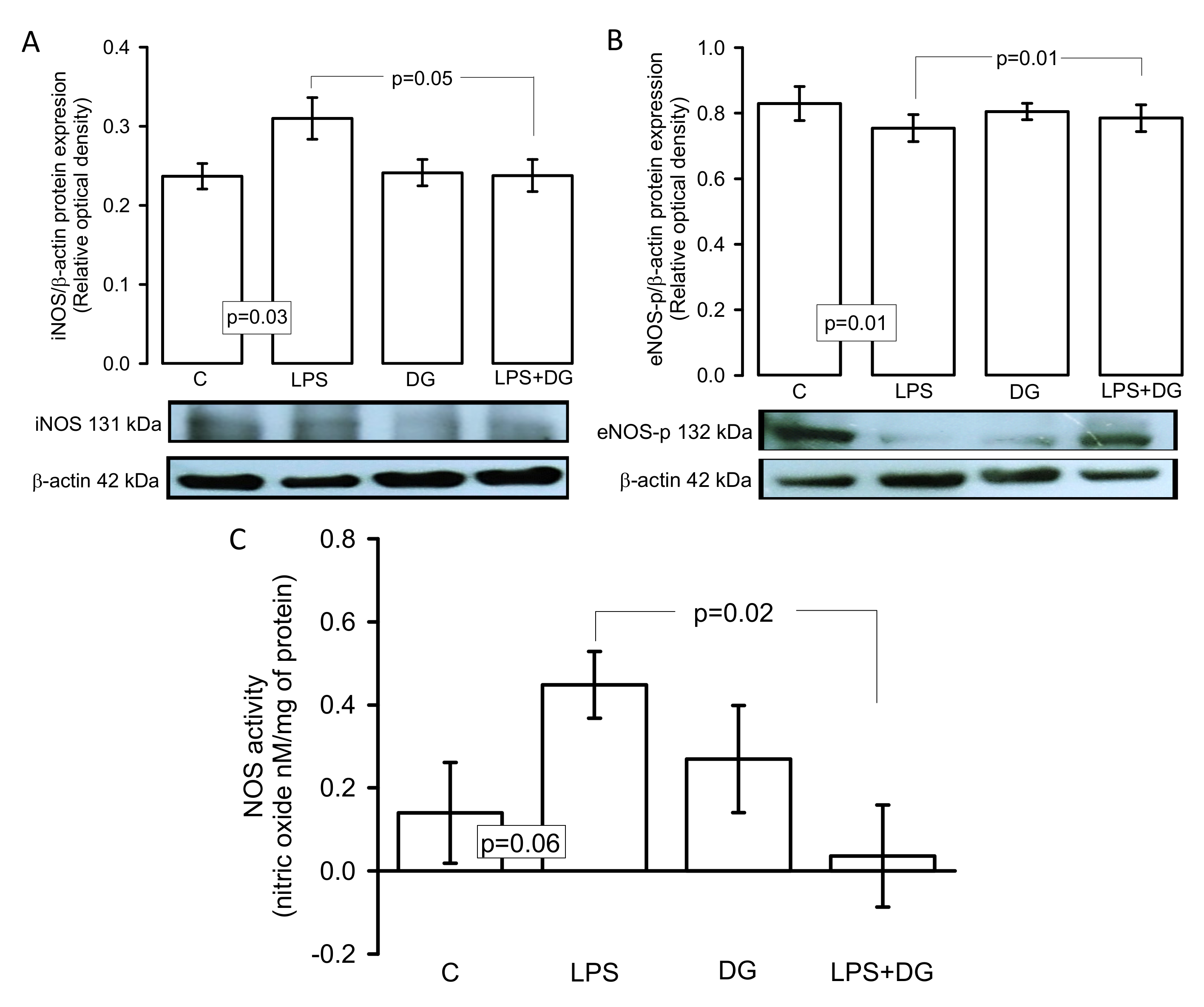
2.7. Expressions of the iNOS and eNOS Enzymes and eNOS-p Activity
3. Discussion
4. Materials and Methods
4.1. Deodorized Garlic
4.2. LPS Inoculation
4.3. Determinations of the Isolated and Perfused Hearts
4.4. Histological Preparation
4.5. Oxidative Stress Markers: Total Antioxidant Capacity, Glutathione, Selenium Lipid Peroxidation, and Thiols Groups
4.6. Determinations of Thioredoxin Reductase (TrxR), Glutathione Peroxidase (GPx), and Glutathione-S-Transferase (GST) Activities
4.7. Western Blotting for TrxR, GPx, CTH, CBS, iNOS, and eNOS-p
4.8. Nitric Oxide Synthase Activity
4.9. Hydrogen Sulfide Concentration
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shi, L.; Lin, Q.; Li, X.; Nie, Y.; Sun, S.; Deng, X.; Wang, L.; Lu, J.; Tang, Y.; Luo, F.A. Garlic organosulfur compound, ameliorates gut inflammation through MAPK-NF-kappaB/AP-1/STAT-1 inactivation and PPAR-gamma activation. Mol. Nutr. Food. Res. 2017, 61, 1601013. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Mathews, A.E.; Rodrigues, C.; Eudy, B.J.; Rowe, C.A.; O’Donoughue, A.; Percival, S.S. Aged garlic extract supplementation modifies inflammation and immunity of adults with obesity: A randomized, double-blind, placebo-controlled clinical trial. Clin. Nutr. ESPEN 2018, 24, 148–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.C.; Sheen, L.Y.; Chen, H.W.; Tsai, S.J.; Lii, C.K. Effects of organosulfur compounds from garlic oil on the antioxidation system in rat liver and red blood cells. Food. Chem. Toxicol. 2002, 39, 563–569. [Google Scholar] [CrossRef]
- Zhang, Z.; Huang, H.; Liu, P.; Tang, C.; Wang, J. Hydrogen sulfide contributes to cardioprotection during ischemia-reperfusion injury by opening K ATP channels. Can. J. Physiol. Pharmacol. 2007, 85, 1248–1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, X.L.; Murphy, R.; Thandapilly, S.J.; Yu, L.; Netticadan, T. Garlic extracts prevent oxidative stress, hypertrophy and apoptosis in cardiomyocytes: A role for nitric oxide and hydrogen sulfide. BMC. Complement. Altern. Med. 2012, 12, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, C.; Percival, S.S. Immunomodulatory effects of glutathione, garlic derivatives, and hydrogen sulfide. Nutrients 2019, 11, 295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stipanuk, M.H. Sulfur amino acid metabolism: Pathways for production and removal of homocysteine and cysteine. Annu. Rev. Nutr. 2004, 24, 539–577. [Google Scholar] [CrossRef]
- Pérez-Torres, I.; Soto, M.E.; Manzano-Pech, L.; Díaz-Diaz, E.; Soria-Castro, E.; Rubio-Ruíz, M.E.; Guarner-Lans, V. Oxidative stress in plasma from patients with marfan syndrome is modulated by deodorized garlic preliminary findings. Oxid. Med. Cell. Longev. 2022, 2022, 5492127. [Google Scholar] [CrossRef]
- Aisa-Alvarez, A.; Soto, M.E.; Guarner-Lans, V.; Camarena-Alejo, G.; Franco-Granillo, J.; Martínez-Rodríguez, E.A.; Ávila, R.G.; Manzano-Pech, L.; Pérez-Torres, I. Usefulness of antioxidants as adjuvant therapy for septic shock: A randomized clinical trial. Medicina 2020, 56, 619. [Google Scholar] [CrossRef]
- Rivers, E.P.; McIntyre, L.; Morro, D.C.; Rivers, K.K. Early and innovative interventions for severe sepsis and septic shock: Taking advantage of a window of opportunity. CMAJ 2005, 173, 1054–1065. [Google Scholar] [CrossRef]
- Russell, J.A. Management of sepsis. N. Engl. J. Med. 2006, 355, 1699–1713. [Google Scholar] [CrossRef] [Green Version]
- Flesch, M.; Kilter, H.; Cremers, B.; Laufs, U.; Südkamp, M.; Ortmann, M.; Müller, F.U.; Böhm, M. Effects of endotoxin on human myocardial contractility involvement of nitric oxide and peroxynitrite. Am. Coll. Cardiol. 1999, 33, 1062–1070. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.Y.; Yiang, G.T.; Liao, W.T.; Tsai, A.P.Y.; Cheng, Y.L.; Cheng, P.W.; Chia-Ying, L.; Chia-Jung, L. Current mechanistic concepts in ischemia and reperfusion injury. Cell. Physiol. Biochem. 2018, 46, 1650–1667. [Google Scholar] [CrossRef]
- Quintero-Fabian, S.; Ortuño-Sahagún, D.; Vázquez-Carrera, M.; López-Roa, R.I. Alliin, a garlic (Allium sativum) compound, prevents LPS-induced inflammation in 3T3-L1 adipocytes. Mediat. Inflamm. 2013, 2013, 381815. [Google Scholar] [CrossRef] [Green Version]
- Arreola, R.; Quintero-Fabián, S.; López-Roa, R.I.; Flores-Gutiérrez, E.O.; Reyes-Grajeda, J.P.; Carrera-Quintanar, L.; Ortuño-Sahagún, D. Immunomodulation and anti-inflammatory effects of garlic compounds. J. Immunol. Res. 2015, 2015, 401630. [Google Scholar] [CrossRef] [Green Version]
- Choumar, A.; Tarhuni, A.; Lettéron, P.; Reyl-Desmars, F.; Dauhoo, N.; Damasse, J.; Vadrot, N.; Nahon, P.; Moreau, R.; Pessayre, D.; et al. Lipopolysaccharide induced mitochondrial DNA depletion. Antioxid. Redox. Signal. 2011, 15, 2837–2854. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Torres, I.; Torres-Narváez, J.C.; Pedraza-Chaverri, J.; Rubio-Ruíz, M.E.; Díaz-Díaz, E.; Del Valle-Mondragón, L.; Martínez-Memije, R.; Varela López, E.; Guarner-Lans, V. Effect of the aged garlic extract on cardiovascular function in metabolic syndrome rats. Molecules 2016, 21, 1425. [Google Scholar] [CrossRef]
- Shang, A.; Cao, S.-Y.; Xu, X.-Y.; Gan, R.-Y.; Tang, G.-Y.; Corke, H.; Mavumengwana, V.; Li, H.-B. Bioactive compounds and biological functions of garlic (Allium sativum L.). Foods 2019, 8, 246. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.M.; Liu, G.Q.; Xian, H.B.; Si, J.L.; Qi, S.X.; Yu, Y.P. LncRNA NEAT1 alleviates sepsis-induced myocardial injury by regulating the TLR2/NF-κB signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 4898–4907. [Google Scholar]
- Goraca, A.; Piechota, A.; Huk-Kolega, H. Effect of alpha-lipoic acid n LPS-induced oxidative stress in the heart. J. Physiol. Pharmacol. 2009, 60, 61–68. [Google Scholar]
- Avula, P.R.; Asdaq, S.M.; Asad, M. Effect of aged garlic extract and S-allyl cysteine and their interaction with atenolol during isoproterenol induced myocardial toxicity in rats. Indian. J. Pharmacol. 2014, 46, 94–99. [Google Scholar] [PubMed] [Green Version]
- You, L.; Pan, Y.Y.; An, M.Y.; Chen, W.H.; Zhang, Y.; Wu, Y.N.; Li, Y.; Sun, K.; Yong-Qiang, Y.; Jian-Shi, L. The cardioprotective effects of remote ischemic conditioning in a rat model of acute myocardial infarction. Med. Sci. Monit. 2019, 25, 1769–1779. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.H.; Kang, D. Physicochemical properties, biological activity, health benefits, and general limitations of aged black garlic: A review. Molecules 2017, 22, 919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeremic, J.N.; Jakovljevic, V.; Zivkovic, V.I.; Srejovic, I.M.; Bradic, J.; Milosavljevic, I.M.; Mitrovic, S.J.; Jovicic, N.U.; Bolevich, S.B.; Svistunov, A.A.; et al. Garlic derived diallyl trisulfide in experimental metabolic syndrome: Metabolic effects and cardioprotective role. Int. J. Mol. Sci. 2020, 21, 9100. [Google Scholar] [CrossRef]
- Amagase, H.; Petesch, B.L.; Matsuura, H.; Kasuga, S.; Itakura, Y. Intake of garlic and its bioactive components. J. Nutr. 2001, 131, 955S–962S. [Google Scholar] [CrossRef] [Green Version]
- Franco-Enzástiga, U.; Santana-Martínez, R.A.; Silva-Islas, C.A.; Barrera-Oviedo, D.; Chánez-Cárdenas, M.E.; Maldonado, P.D. Chronic administration of S-allylcysteine activates Nrf2 factor and enhances the activity of antioxidant enzymes in the striatum, frontal cortex and hippocampus. Neurochem. Res. 2017, 42, 3041–3051. [Google Scholar] [CrossRef]
- Bradley, J.M.; Organ, C.L.; Lefer, D.J. Garlic-derived organic polysulfides and myocardial protection. J. Nutr. 2016, 146, 403S–409S. [Google Scholar] [CrossRef] [Green Version]
- Kimura, Y.; Goto, Y.; Kimura, H. Hydrogen sulfide increases glutathione production and suppresses oxidative stress in mitochondria. Antioxid. Redox. Signal. 2010, 12, 1–13. [Google Scholar] [CrossRef]
- Ide, N.; Lau, B.H. Garlic compounds minimize intracellular oxidative stress and inhibit nuclear factor-κB activation. J. Nutr. 2001, 131, 1020S–1026S. [Google Scholar] [CrossRef] [Green Version]
- Kaya-Yasar, Y.; Karaman, Y.; Bozjurt, T.E.; Onder, S.C.; Sahin-Erdemli, I. Effects of intranasal treatment with slow (GYY4137) and rapid (NaHS) donors of hydrogen sulfide in lipopolysaccharide-induced airway inflammation in mice. Pulm. Pharmacol. Ther. 2017, 45, 170–180. [Google Scholar] [CrossRef]
- Bekpinar, S.; Unlucerci, Y.; Uysal, M.; Gurdol, F. Propargylglycine aggravates liver damage in LPS-treated rats: Possible relation of nitrosative stress with the inhibition of H2S formation. Pharmacol. Rep. 2014, 66, 897–901. [Google Scholar] [CrossRef]
- Ried, K.; Fakler, P. Potential of garlic (Allium sativum) in lowering high blood pressure: Mechanisms of action and clinical relevance. Integr. Blood. Press. Control. 2014, 7, 71–82. [Google Scholar] [CrossRef]
- Predmore, B.L.; Kondo, K.; Bhushan, S.; Zlatopolsky, M.A.; King, A.L.; Aragon, J.P.; Grinsfelder, D.B.; Condit, M.E.; Lefer, D.J. The polysulfide diallyl trisulfide protects the ischemic myocardium by preservation of endogenous hydrogen sulfide and increasing nitric oxide bioavailability. Am. J. Physiol. Heart. Circ. Physiol. 2012, 302, H2410–H2418. [Google Scholar] [CrossRef]
- Morihara, N.; Sumioka, I.; Ide, N.; Moriguchi, T.; Uda, N.; Kyo, E. Aged garlic extract maintains cardiovascular homeostasis in mice and rats. J. Nutr. 2006, 136, 777S–781S. [Google Scholar] [CrossRef] [Green Version]
- Dirsch, V.M.; Kiemer, A.K.; Wagner, H.; Vollmar, A.M. Effect of allicin and ajoene, two compounds of garlic, on inducible nitric oxide synthase. Atherosclerosis 1998, 139, 333–339. [Google Scholar] [CrossRef]
- Ziolo, M.T.; Maier, L.S.; Piacentino, V.; Bossuyt, J.; Houser, S.T.; Bers, D.M. Myocyte nitric oxide synthase 2 contributes to blunted beta-adrenergic response in failing human hearts by decreasing Ca2+ transients. Circulation 2004, 109, 1886–1891. [Google Scholar] [CrossRef] [Green Version]
- Clapp, B.R.; Hingorani, A.D.; Kharbanda, R.K.; Mohamed-Ali, V.; Stephens, J.W.; Vallance, P.; MacAllister, R.J. Inflammation-induced endothelial dysfunction involves reduced nitric oxide bioavailability and increased oxidant stress. Cardiovasc. Res. 2004, 64, 172–178. [Google Scholar] [CrossRef]
- Rauf, A.; Abu-Izneid, T.; Thiruvengadam, M.; Imran, M.; Olatunde, A.; Shariati, M.A.; Bawazeer, S.; Naz, S.; Shirooie, S.; Sanches-Silva, A.; et al. Garlic (Allium sativum L.): Its Chemistry, Nutritional Composition, Toxicity, and Anticancer Properties. Curr. Top Med. Chem. 2022, 22, 957–972. [Google Scholar] [CrossRef]
- Ried, K.; Travica, N.; Sali, A. The effect of aged garlic extract on blood pressure and other cardiovascular risk factors in uncontrolled hypertensives: The AGE at heart trial. Integr. Blood. Press. Control 2016, 9, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Hayashida, R.; Kondo, K.; Morita, S.; Unno, K.; Shintani, S.; Shimizu, Y.; Calvert, J.W.; Shibata, R.; Murohara, T. Diallyl Trisulfide Augments Ischemia-Induced Angiogenesis via an Endothelial Nitric Oxide Synthase-Dependent Mechanism. Circ. J. 2017, 81, 870–878. [Google Scholar] [CrossRef] [Green Version]
- Borlinghaus, J.; Albrecht, F.; Gruhlke, M.C.; Nwachukwu, I.D.; Slusarenko, A.J. Allicin: Chemistry and biological properties. Molecules 2014, 8, 12591–12618. [Google Scholar] [CrossRef] [Green Version]
- Bayan, L.; Koulivand, P.H.; Gorji, A. Garlic: A review of potential therapeutic effects. Avicenna. J. Phytomed. 2014, 4, 1–14. [Google Scholar]
- Greasley, A.; Zhang, Y.; Wu, B.; Pei, Y.; Belzile, N.; Yang, G. H2S protects against cardiac cell hypertrophy through regulation of selenoproteins. Oxid. Med. Cell. Longev. 2019, 2019, 6494306. [Google Scholar] [CrossRef]
- Szabó, G.; Veres, G.; Radovits, T.; Gero, D.; Módis, K.; Miesel-Gröschel, C.; Horkay, F.; Karck, M.; Szabó, C. Cardioprotective effects of hydrogen sulfide. Nitric Oxide. 2011, 25, 201–210. [Google Scholar] [CrossRef] [Green Version]
- Gruhlke, M.C.H.; Slusarenko, A.J. The chemistry of alliums. Molecules 2018, 23, 143. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhao, Y.; Wei, Z.; Tao, L.; Sheng, X.; Wang, S. Targeting thioredoxin system with an organosulfur compound, diallyl trisulfide (DATS), attenuates progression and metastasis of triple-negative breast cancer (TNBC). Cell. Physiol. Biochem. 2018, 50, 1945–1963. [Google Scholar] [CrossRef]
- Kojima, R.; Toyama, Y.; Ohnishi, S.T. Protective effects of an aged garlic extract on doxorubicin-induced cardiotoxicity in the mouse. Nutr. Cancer 1994, 22, 163–173. [Google Scholar] [CrossRef]
- Chih-Chung, W.; Lee-Yan, S.; Haw-Wen, C.; Wei-Wen, K.; Shun-Jen, T.; Chong-Kuei, L. Differential effects of garlic oil and its three major organosulfur components on the hepatic detoxification system in rats. J. Agric. Food. Chem. 2002, 50, 378–383. [Google Scholar]
- Fukao, T.; Hosono, T.; Misawa, S.; Seki, T.; Ariga, T. The effects of allyl sulfides on the induction of phase II detoxification enzymes and liver injury by carbon tetrachloride. Food Chem. Toxicol. 2004, 42, 743–749. [Google Scholar] [CrossRef]
- Döring, H.J.; Dehvert, H. The Isolated Perfused Warm-Blooded Heart According to Langendorff. In Biological Measurement Techniques, 1st ed.; Dehnert, H., Ed.; Biomes Technik-Verlag: Baden-Wurtemberg, Germany, 1988; pp. 1–131. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of ‘antioxidant power’: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, I.; Kode, A.; Biswas, S. Assay for quantitative determination of glutathione and glutathione disulfide levels using enzymatic recycling method. Nat. Protoc. 2006, 1, 3159–3165. [Google Scholar] [CrossRef] [PubMed]
- Soto, M.E.; Manzano-Pech, L.G.; Guarner-Lans, V.; Díaz-Galindo, J.A.; Vásquez, X.; Castrejón-Tellez, V.; Gamboa, R.; Huesca, C.; Fuentevilla-Alvárez, G.; Pérez-Torres, I. Oxidant/antioxidant profile in the thoracic aneurysm of patients with the Loeys-Dietz syndrome. Oxid. Med. Cell. Longev. 2020, 2020, 5392454. [Google Scholar] [CrossRef] [PubMed]
- Erel, O.; Neselioglu, S. A novel and automated assay for thiol/disulphide homeostasis. Clin. Biochem. 2014, 49, 326–332. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione-S-transferases. The first enzymatic step in mercapturic acid formation. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Soto, M.E.; Iturriaga, H.A.V.; Guarner-Lans, V.; Zuñiga-Muñoz, A.; Aranda, F.A.; Velázquez, E.R.; Pérez-Torres, I. Participation of oleic acid in the formation of the aortic aneurysm in Marfan syndrome patients. Prostaglandins. Other. Lipid. Mediat. 2016, 123, 46–55. [Google Scholar] [CrossRef]
- Padiya, R.; Khatua, T.N.; Bagul, P.K.; Kuncha, M.; Banerjee, S.K. Garlic improves insulin sensitivity and associated metabolic syndromes in fructose fed rats. Nutr. Metab. 2011, 8, 53. [Google Scholar] [CrossRef] [Green Version]
- Padiya, R.; Chowdhury, D.; Borkar, R.; Srinivas, R.; Bhadra, M.P.; Banerjee, S.K. Garlic attenuates cardiac oxidative stress via activation of PI3K/AKT/Nrf2-Keap1 pathway in fructose-fed diabetic rat. PLoS ONE 2014, 9, e94228. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables (mg of Protein) | C | LPS | DG | LPS + DG |
|---|---|---|---|---|
| TAC (nM) | 691.30 ± 64.14 | 483.91 ± 51.20 * | 715.80 ± 52.86 | 663.54 ± 49.71 * |
| GSH (µM) | 0.009 ± 0.001 | 0.003 ± 0.0009 ** | 0.009 ± 0.001 | 0.010 ± 0.001 ** |
| Se (nM) | 0.022 ± 0.001 | 0.025 ± 0.003 | 0.022 ± 0.002 | 0.041 ± 0.001 ** |
| GST activity (GS + DNB µmol) | 0.001 ± 0.0001 | 0.0006 ± 0.00005 * | 0.001 ± 0.00007 | 0.001 ± 0.0001 * |
| LPO (nmol/MDA) | 2.89 ± 0.49 | 12.86 ± 5.25 ** | 3.56 ± 0.17 | 3.78 ± 0.41* |
| Thiols (µM) | 251.74 ± 16.74 | 166.46 ± 14.35 ** | 244.451 ± 8.53 | 272.28 ± 8.39 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Torres, I.; Manzano-Pech, L.; Guarner-Lans, V.; Soto, M.E.; Castrejón-Téllez, V.; Márquez-Velasco, R.; Vargas-González, Á.; Martínez-Memije, R.; Del Valle-Mondragón, L.; Díaz-Juárez, J.A.; et al. Deodorized Garlic Decreases Oxidative Stress Caused by Lipopolysaccharide in Rat Heart through Hydrogen Sulfide: Preliminary Findings. Int. J. Mol. Sci. 2022, 23, 12529. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232012529
Pérez-Torres I, Manzano-Pech L, Guarner-Lans V, Soto ME, Castrejón-Téllez V, Márquez-Velasco R, Vargas-González Á, Martínez-Memije R, Del Valle-Mondragón L, Díaz-Juárez JA, et al. Deodorized Garlic Decreases Oxidative Stress Caused by Lipopolysaccharide in Rat Heart through Hydrogen Sulfide: Preliminary Findings. International Journal of Molecular Sciences. 2022; 23(20):12529. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232012529
Chicago/Turabian StylePérez-Torres, Israel, Linaloe Manzano-Pech, Verónica Guarner-Lans, María Elena Soto, Vicente Castrejón-Téllez, Ricardo Márquez-Velasco, Álvaro Vargas-González, Raúl Martínez-Memije, Leonardo Del Valle-Mondragón, Julieta Anabell Díaz-Juárez, and et al. 2022. "Deodorized Garlic Decreases Oxidative Stress Caused by Lipopolysaccharide in Rat Heart through Hydrogen Sulfide: Preliminary Findings" International Journal of Molecular Sciences 23, no. 20: 12529. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232012529