
Trichostatin A-Mediated Epigenetic Modulation Predominantly Triggers Transcriptomic Alterations in the Ex Vivo Expanded Equine Chondrocytes
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Efficiency Alignment of NGS Reads
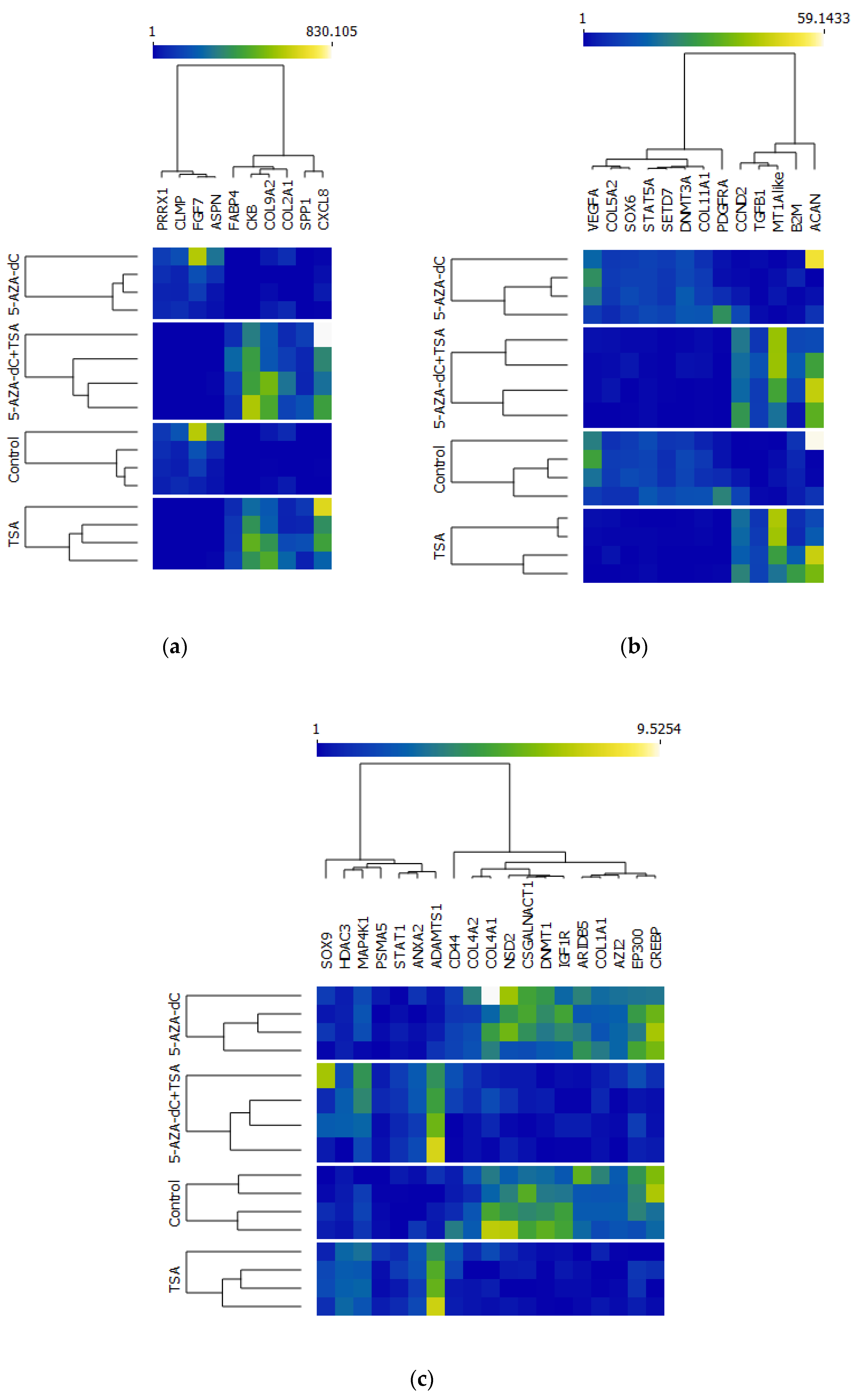
2.2. Differentially Expressed Genes (DEGs) Obtained Using 3′ mRNA-Seq
2.3. General Description of Differentially Expressed Genes upon Applying Chondrocyte Stimulation in Monolayer Culture
2.4. Results of Functional Overrepresentation of DEGs Using DAVID Annotation Tools
2.5. Validation in RNA-Seq Results
3. Discussion
4. Materials and Methods
4.1. Short Description of the Research
4.2. Chondrocyte Culture Conditions and Applied Stimulations
4.3. Preparation of RNA and 3′ RNA-seq Libraries, and Next-Generation Sequencing
4.4. Trimming, Filtering, Quantification, and Mapping of Demultiplexed NGS Reads, and Differential Analysis
4.5. DEGs’ Functional Annotation in KEGG Pathways
4.6. Real-Time PCR
5. Conclusions and Future Goals
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ortved, K.F.; Nixon, A.J. Cell-based cartilage repair strategies in the horse. Vet. J. 2016, 208, 1–12. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Lipa, K.E.; Alexander, P.G.; Clark, K.L.; Lin, H. Potential Methods of Targeting Cellular Aging Hallmarks to Reverse Osteoarthritic Phenotype of Chondrocytes. Biology 2022, 11, 996. [Google Scholar] [CrossRef] [PubMed]
- Ulivi, V.; Giannoni, P.; Gentili, C.; Cancedda, R.; Descalzi, F. p38/NF-kB-dependent expression of COX-2 during differentiation and inflammatory response of chondrocytes. J. Cell. Biochem. 2008, 104, 1393–1406. [Google Scholar] [CrossRef]
- Buhrmann, C.; Popper, B.; Aggarwal, B.B.; Shakibaei, M. Resveratrol downregulates inflammatory pathway activated by lymphotoxin α (TNF-β) in articular chondrocytes: Comparison with TNF-α. PLoS ONE 2017, 12, e0186993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buhrmann, C.; Brockmueller, A.; Mueller, A.L.; Shayan, P.; Shakibaei, M. Curcumin Attenuates Environment-Derived Osteoarthritis by Sox9/NF-kB Signaling Axis. Int. J. Mol. Sci. 2021, 22, 7645. [Google Scholar] [CrossRef] [PubMed]
- Hata, K. Epigenetic regulation of chondrocyte differentiation. Jpn. Dent. Sci. Rev. 2015, 51, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Michalowsky, L.A.; Jones, P.A. Differential nuclear protein binding to 5-azacytosine-containing DNA as a potential mechanism for 5-aza-2’-deoxycytidine resistance. Mol. Cell Biol. 1987, 7, 3076–3083. [Google Scholar]
- Vanhaecke, T.; Papeleu, P.; Elaut, G.; Rogiers, V. Trichostatin A-like hydroxamate histone deacetylase inhibitors as therapeutic agents: Toxicological point of view. Curr. Med. Chem. 2004, 11, 1629–1643. [Google Scholar] [CrossRef]
- Drummond, D.C.; Noble, C.O.; Kirpotin, D.B.; Guo, Z.; Scott, G.K.; Benz, C.C. Clinical development of histone deacetylase inhibitors as anticancer agents. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 495–528. [Google Scholar] [CrossRef]
- Vigushin, D.M.; Ali, S.; Pace, P.E.; Mirsaidi, N.; Ito, K.; Adcock, I.; Coombes, R.C. Trichostatin A is a histone deacetylase inhibitor with potent antitumor activity against breast cancer in vivo. Clin. Cancer Res. 2001, 7, 971–976. [Google Scholar]
- Haq, S.H. 5-Aza-2’-deoxycytidine acts as a modulator of chondrocyte hypertrophy and maturation in chick caudal region chondrocytes in culture. Anat. Cell Biol. 2016, 49, 107–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadler, S.; Vural, Ö.; Rosowski, J.; Reiners-Schramm, L.; Lauster, R.; Rosowski, M. Effects of 5-aza-2’-deoxycytidine on primary human chondrocytes from osteoarthritic patients. PLoS ONE 2020, 15, e0234641. [Google Scholar] [CrossRef]
- Young, D.A.; Lakey, R.L.; Pennington, C.J.; Jones, D.; Kevorkian, L.; Edwards, D.R.; Cawston, T.E.; Clark, I.M. Histone deacetylase inhibitors modulate metalloproteinase gene expression in chondrocytes and block cartilage resorption. Arthritis Res. Ther. 2005, 7, 503–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leoni, F.; Zaliani, A.; Bertolini, G.; Porro, G.; Pagani, P.; Pozzi, P.; Donà, G.; Fossati, G.; Sozzani, S.; Azam, T.; et al. The antitumor histone deacetylase inhibitor suberoylanilide hydroxamic acid exhibits antiinflammatory properties via suppression of cytokines. Proc. Natl. Acad. Sci. USA 2002, 99, 2995–3000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Im, G.I. Effects of Trichostatin A on the Chondrogenesis from Human Mesenchymal Stem Cells. Tissue Eng. Regen. Med. 2017, 14, 403–410. [Google Scholar] [CrossRef]
- Wan, C.; Zhang, F.; Yao, H.; Li, H.; Tuan, R.S. Histone Modifications and Chondrocyte Fate: Regulation and Therapeutic Implications. Front. Cell Dev. Biol. 2021, 9, 626708. [Google Scholar] [CrossRef] [PubMed]
- Miyagawa, J.; Muguruma, M.; Aoto, H.; Suetake, I.; Nakamura, M.; Tajima, S. Isolation of the novel cDNA of a gene of which expression is induced by a demethylating stimulus. Gene 1999, 240, 289–295. [Google Scholar] [CrossRef]
- Goldring, M.B. Human chondrocyte cultures as models of cartilage-specific gene regulation. Methods Mol. Med. 2005, 107, 69–95. [Google Scholar] [PubMed] [Green Version]
- Liu, C.F.; Samsa, W.E.; Zhou, G.; Lefebvre, V. Transcriptional control of chondrocyte specification and differentiation. Semin. Cell Dev. Biol. 2017, 62, 34–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loeser, R.F.; Im, H.J.; Richardson, B.; Lu, Q.; Chubinskaya, S. Methylation of the OP-1 promoter: Potential role in the age-related decline in OP-1 expression in cartilage. Osteoarthr. Cartil. 2009, 17, 513–517. [Google Scholar] [CrossRef] [Green Version]
- Rao, J.; Bhattacharya, D.; Banerjee, B.; Sarin, A.; Shivashankar, G.V. Trichostatin-A induces differential changes in histone protein dynamics and expression in HeLa cells. Biochem. Biophys. Res. Commun. 2007, 363, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Buhrmann, C.; Busch, F.; Shayan, P.; Shakibaei, M. Sirtuin-1 (SIRT1) is required for promoting chondrogenic differentiation of mesenchymal stem cells. J. Biol. Chem. 2014, 289, 22048–22062. [Google Scholar] [CrossRef] [PubMed]
- Korogi, W.; Yoshizawa, T.; Karim, M.F.; Tanoue, H.; Yugami, M.; Sobuz, S.U.; Hinoi, E.; Sato, Y.; Oike, Y.; Mizuta, H.; et al. SIRT7 is an important regulator of cartilage homeostasis and osteoarthritis development. Biochem. Biophys. Res. Commun. 2018, 496, 891–897. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Chen, B.F.; Chan, W.Y. An epigenetic regulator: Methyl-CpG-binding domain protein 1 (MBD1). Int. J. Mol. Sci. 2015, 16, 5125–5140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.F.; Lefebvre, V. The transcription factors SOX9 and SOX5/SOX6 cooperate genome-wide through super-enhancers to drive chondrogenesis. Nucleic Acids Res. 2015, 43, 8183–8203. [Google Scholar] [CrossRef] [Green Version]
- Imagawa, K.; de Andrés, M.C.; Hashimoto, K.; Itoi, E.; Otero, M.; Roach, H.I.; Goldring, M.B.; Oreffo, R.O. Association of reduced type IX collagen gene expression in human osteoarthritic chondrocytes with epigenetic silencing by DNA hypermethylation. Arthritis Rheumatol. 2014, 66, 3040–3051. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Liu, S.; Huang, J.; Guo, W.; Chen, J.; Zhang, L.; Zhao, B.; Peng, J.; Wang, A.; Wang, Y.; et al. The ECM-cell interaction of cartilage extracellular matrix on chondrocytes. Biomed. Res. Int. 2014, 2014, 648459. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.; Lee, M.N.; Choung, J.S.; Kim, S.; Choi, B.H.; Noh, M.; Shin, J.H. Focal Adhesion Assembly Induces Phenotypic Changes and Dedifferentiation in Chondrocytes. J. Cell Physiol. 2016, 231, 1822–1831. [Google Scholar] [CrossRef]
- Schipani, E.; Ryan, H.E.; Didrickson, S.; Kobayashi, T.; Knight, M.; Johnson, R.S. Hypoxia in cartilage: HIF-1alpha is essential for chondrocyte growth arrest and survival. Genes Dev. 2001, 15, 2865–2876. [Google Scholar] [CrossRef]
- Kita, K.; Kimura, T.; Nakamura, N.; Yoshikawa, H.; Nakano, T. PI3K/Akt signaling as a key regulatory pathway for chondrocyte terminal differentiation. Genes Cells. 2008, 13, 839–850. [Google Scholar] [CrossRef]
- Tekari, A.; Luginbuehl, R.; Hofstetter, W.; Egli, R.J. Transforming growth factor beta signaling is essential for the autonomous formation of cartilage-like tissue by expanded chondrocytes. PLoS ONE 2015, 10, e0120857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rokutanda, S.; Fujita, T.; Kanatani, N.; Yoshida, C.A.; Komori, H.; Liu, W.; Mizuno, A.; Komori, T. Akt regulates skeletal development through GSK3, mTOR, and FoxOs. Dev. Biol. 2009, 328, 78–93. [Google Scholar] [CrossRef] [PubMed]
- Stanton, L.A.; Underhill, T.M.; Beier, F. MAP kinases in chondrocyte differentiation. Dev. Biol. 2003, 263, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Hollander, J.M.; Zeng, L. The Emerging Role of Glucose Metabolism in Cartilage Development. Curr. Osteoporos. Rep. 2019, 17, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Wang, P.; Lee, N.H.; Goldring, M.B.; Konstantopoulos, K. Prolonged application of high fluid shear to chondrocytes recapitulates gene expression profiles associated with osteoarthritis. PLoS ONE 2010, 5, e15174. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Shao, Y.Y.; Ballock, R.T. Thyroid hormone-mediated growth and differentiation of growth plate chondrocytes involves IGF-1 modulation of beta-catenin signaling. J. Bone Min. Res. 2010, 25, 1138–1146. [Google Scholar] [CrossRef]
- Tseng, C.C.; Chen, Y.J.; Chang, W.A.; Tsai, W.C.; Ou, T.T.; Wu, C.C.; Sung, W.Y.; Yen, J.H.; Kuo, P.L. Dual Role of Chondrocytes in Rheumatoid Arthritis: The Chicken and the Egg. Int. J. Mol. Sci. 2020, 21, 1071. [Google Scholar] [CrossRef] [Green Version]
- Kelwick, R.; Desanlis, I.; Wheeler, G.N.; Edwards, D.R. The ADAMTS (A Disintegrin and Metalloproteinase with Thrombos-pondin motifs) family. Genome Biol. 2015, 16, 113. [Google Scholar] [CrossRef] [Green Version]
- Legeai-Mallet, L.; Benoist-Lasselin, C.; Munnich, A.; Bonaventure, J. Overexpression of FGFR3, Stat1, Stat5 and p21Cip1 correlates with phenotypic severity and defective chondrocyte differentiation in FGFR3-related chondrodysplasias. Bone 2004, 34, 26–36. [Google Scholar] [CrossRef]
- Caron, M.M.; Emans, P.J.; Coolsen, M.M.; Voss, L.; Surtel, D.A.; Cremers, A.; van Rhijn, L.W.; Welting, T.J. Redifferentiation of dedifferentiated human articular chondrocytes: Comparison of 2D and 3D cultures. Osteoarthr. Cartil. 2012, 20, 1170–1178. [Google Scholar] [CrossRef] [Green Version]
- Edwards, R.B.; Lu, Y.; Cole, B.J.; Muir, P.; Markel, M.D. Comparison of radiofrequency treatment and mechanical debride-ment of fibrillated cartilage in an equine model. Vet. Comp. Orthop. Traumatol. 2008, 21, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Edwards, R.B., 3rd; Lu, Y.; Uthamanthil, R.K.; Bogdanske, J.J.; Muir, P.; Athanasiou, K.A.; Markel, M.D. Comparison of me-chanical debridement and radiofrequency energy for chondroplasty in an in vivo equine model of partial thickness carti-lage injury. Osteoarthr. Cartil. 2007, 15, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Shakya, B.R.; Tiulpin, A.; Saarakkala, S.; Turunen, S.; Thevenot, J. Detection of experimental cartilage damage with acoustic. Equine Vet. J. 2020, 52, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Uthamanthil, R.K.; Edwards, R.B.; Lu, Y.; Manley, P.A.; Athanasiou, K.A.; Markel, M.D. In vivo study on the short-term effect of radiofrequency energy on chondromalacic patellar cartilage and its correlation with calcified cartilage pathology in an equine model. J. Orthop. Res. 2006, 24, 716–724. [Google Scholar] [CrossRef]
- Ryan, A.; Bertone, A.L.; Kaeding, C.C.; Backstrom, K.C.; Weisbrode, S.E. The effects of radiofrequency energy treatment on chondrocytes and matrix of fibrillated articular cartilage. Am. J. Sports Med. 2003, 31, 386–391. [Google Scholar] [CrossRef]
- Bodó, G.; Hangody, L.; Szabó, Z.; Peham, C.; Schinzel, M.; Girtler, D.; Sótonyi, P. Arthroscopic autologous osteochondral mosaicplasty for the treatment of subchondral cystic lesion in the medial femoral condyle in a horse. Acta Vet. Hung. 2000, 48, 343–354. [Google Scholar]
- Bodo, G.; Hangody, L.; Modis, L.; Hurtig, M. Autologous osteochondral grafting (mosaic arthroplasty) for treatment of subchondral cystic lesions in the equine stifle and fetlock joints. Vet. Surg. 2004, 33, 588–596. [Google Scholar] [CrossRef]
- Sparks, H.D.; Nixon, A.J.; Bogenrief, D.S. Reattachment of the articular cartilage component of type 1 subchondral cystic lesions of the medial femoral condyle with polydioxanone pins in 3 horses. J. Am. Vet. Med. Assoc. 2011, 238, 636–640. [Google Scholar] [CrossRef]
- Smith, M.A.; Walmsley, J.P.; Phillips, T.J.; Pinchbeck, G.L.; Booth, T.M.; Greet, T.R.; Richardson, D.W.; Ross, M.W.; Schramme, M.C.; Singer, E.R.; et al. Effect of age at presentation on outcome following arthroscopic debridement of sub-chondral cystic lesions of the medial femoral condyle: 85 horses (1993—2003). Equine Vet. J. 2005, 37, 175–180. [Google Scholar] [CrossRef]
- Frazer, L.L.; Santschi, E.M.; Fischer, K.J. The impact of subchondral bone cysts on local bone stresses in the medial femoral condyle of the equine stifle joint. Med. Eng. Phys. 2017, 48, 158–167. [Google Scholar] [CrossRef]
- Russell, J.W.; Hall, M.S.; Kelly, G.M. Osteochondroma on the cranial aspect of the distal radial metaphysis causing teno-synovitis of the extensor carpi radialis tendon sheath in a horse. Aust. Vet. J. 2017, 95, 46–48. [Google Scholar] [CrossRef] [PubMed]
- Olstad, K.; Østevik, L.; Carlson, C.S.; Ekman, S. Osteochondrosis Can Lead to Formation of Pseudocysts and True Cysts in the Subchondral Bone of Horses. Vet. Pathol. 2015, 52, 862–872. [Google Scholar] [CrossRef] [PubMed]
- Secombe, C.J.; Anderson, B.H. Diagnosis and treatment of an osteochondroma of the distal tibia in a 3-year-old horse. Aust. Vet. J. 2000, 78, 16–18. [Google Scholar] [CrossRef]
- Ząbek, T.; Witarski, W.; Semik-Gurgul, E.; Szmatoła, T.; Kowalska, K.; Bugno-Poniewierska, M. Chondrogenic expression and DNA methylation patterns in prolonged passages of chondrocyte cell lines of the horse. Gene 2019, 707, 58–64. [Google Scholar] [CrossRef]
- Miao, Z.; Lu, Z.; Wu, H.; Liu, H.; Li, M.; Lei, D.; Zheng, L.; Zhao, J. Collagen, agarose, alginate, and Matrigel hydrogels as cell substrates for culture of chondrocytes in vitro: A comparative study. J. Cell. Biochem. 2018, 119, 7924–7933. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, T.L.; Toh, W.S.; Pei, M. The role of laminins in cartilaginous tissues: From development to regeneration. Eur. Cell Mater. 2017, 34, 40–54. [Google Scholar] [CrossRef]
- Dodt, M.; Roehr, J.T.; Ahmed, R.; Dieterich, C. Flexbar—Flexible barcode and adapter processing for next-generation sequencing platforms. Biology 2012, 1, 895–905. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [Green Version]
- Putri, G.H.; Anders, S.; Pyl, P.T.; Pimanda, J.E.; Zanini, F. Analysing high-throughput sequencing data in Python with HTSeq 2.0. Bioinformatics 2022, 38, 2943–2945. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Dennis, G., Jr.; Sherman, B.T.; Hosack, D.A.; Yang, J.; Gao, W.; Lane, H.C.; Lempicki, R.A. DAVID: Database for Annotation, Visualization, and Integrated Discovery. Genome. Biol. 2003, 4, P3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveros, J.; Venny, C. An Interactive Tool for Comparing Lists with Venn’s Diagrams. 2007–2015. Available online: https://bioinfogp.cnb.csic.es/tools/venny/index.html (accessed on 8 August 2022).
- Demsar, J.; Curk, T.; Erjavec, A.; Gorup, C.; Hocevar, T.; Milutinovic, M.; Mozina, M.; Polajnar, M.; Toplak, M.; Staric, A.; et al. Orange: Data Mining Toolbox in Python. J. Mach. Learn. Res. 2013, 14, 2349–2353. [Google Scholar]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | 3rd Passage Chondrocyte Stimulation | 3’ mRNA-Seq Library | Total Reads after Filtering | Number of Uniquely Mapped Reads |
|---|---|---|---|---|
| I | 5-AZA-dc 1 | 2aza-1 | 3,114,172 | 2,541,510 (81.6%) |
| I | 5-AZA-dc | 3aza-1 | 2,685,011 | 2,216,276 (82.5%) |
| I | 5-AZA-dc | 4/DEO | 2,332,147 | 1,913,713 (82.1%) |
| I | 5-AZA-dc | 6/DEO | 2,603,715 | 2,137,736 (82.1%) |
| II | 5-AZA-dc+TSA 2 | 3tsaza-1 | 3,490,960 | 2,862,115 (82.0%) |
| II | 5-AZA-dc+TSA | 2tsaaza-1 | 2,915,748 | 2,336,842 (80.1%) |
| II | 5-AZA-dc+TSA | 4/Deo+TSA | 3,091,055 | 2,548,432 (82.4%) |
| II | 5-AZA-dc+TSA | 6/DEO+TSA | 2,821,472 | 2,329,453 (82.6%) |
| III | Control | 2k-1 | 2,361,866 | 1,954,158 (82.7%) |
| III | Control | 3k-1 | 3,062,350 | 2,511,373 (82.0%) |
| III | Control | 4/K | 3,312,867 | 2,732,432 (82.5%) |
| III | Control | 6/K | 3,207,522 | 2,627,829 (81.9%) |
| IV | TSA | 3tsa-1 | 3,699,592 | 3,054,305 (82.6%) |
| IV | TSA | 2tsa-1 | 2,753,414 | 2,261,816 (82.1%) |
| IV | TSA | 4/TSA | 3,158,031 | 2,620,954 (83.0%) |
| IV | TSA | 6/TSA | 2,858,574 | 2,361,644 (82.6%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ząbek, T.; Witarski, W.; Szmatoła, T.; Sawicki, S.; Mrozowicz, J.; Samiec, M. Trichostatin A-Mediated Epigenetic Modulation Predominantly Triggers Transcriptomic Alterations in the Ex Vivo Expanded Equine Chondrocytes. Int. J. Mol. Sci. 2022, 23, 13168. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113168
Ząbek T, Witarski W, Szmatoła T, Sawicki S, Mrozowicz J, Samiec M. Trichostatin A-Mediated Epigenetic Modulation Predominantly Triggers Transcriptomic Alterations in the Ex Vivo Expanded Equine Chondrocytes. International Journal of Molecular Sciences. 2022; 23(21):13168. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113168
Chicago/Turabian StyleZąbek, Tomasz, Wojciech Witarski, Tomasz Szmatoła, Sebastian Sawicki, Justyna Mrozowicz, and Marcin Samiec. 2022. "Trichostatin A-Mediated Epigenetic Modulation Predominantly Triggers Transcriptomic Alterations in the Ex Vivo Expanded Equine Chondrocytes" International Journal of Molecular Sciences 23, no. 21: 13168. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113168