
Protein Alterations in Cardiac Ischemia/Reperfusion Revealed by Spatial-Omics
 , ,
, ,
Abstract
:1. Introduction
2. Results
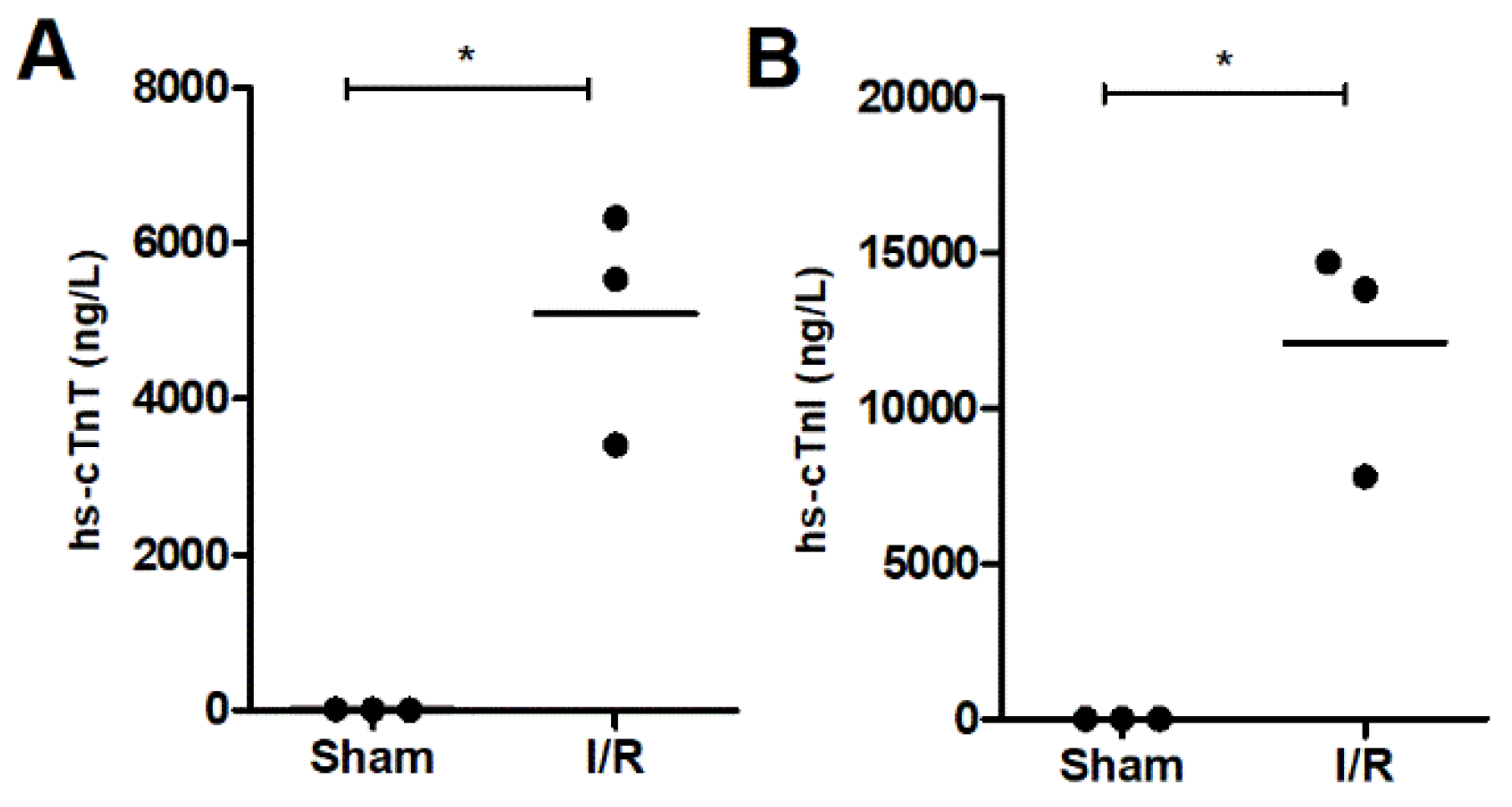
2.1. Cardiac Troponin as a Biomarker for Ischemic Heart Injury
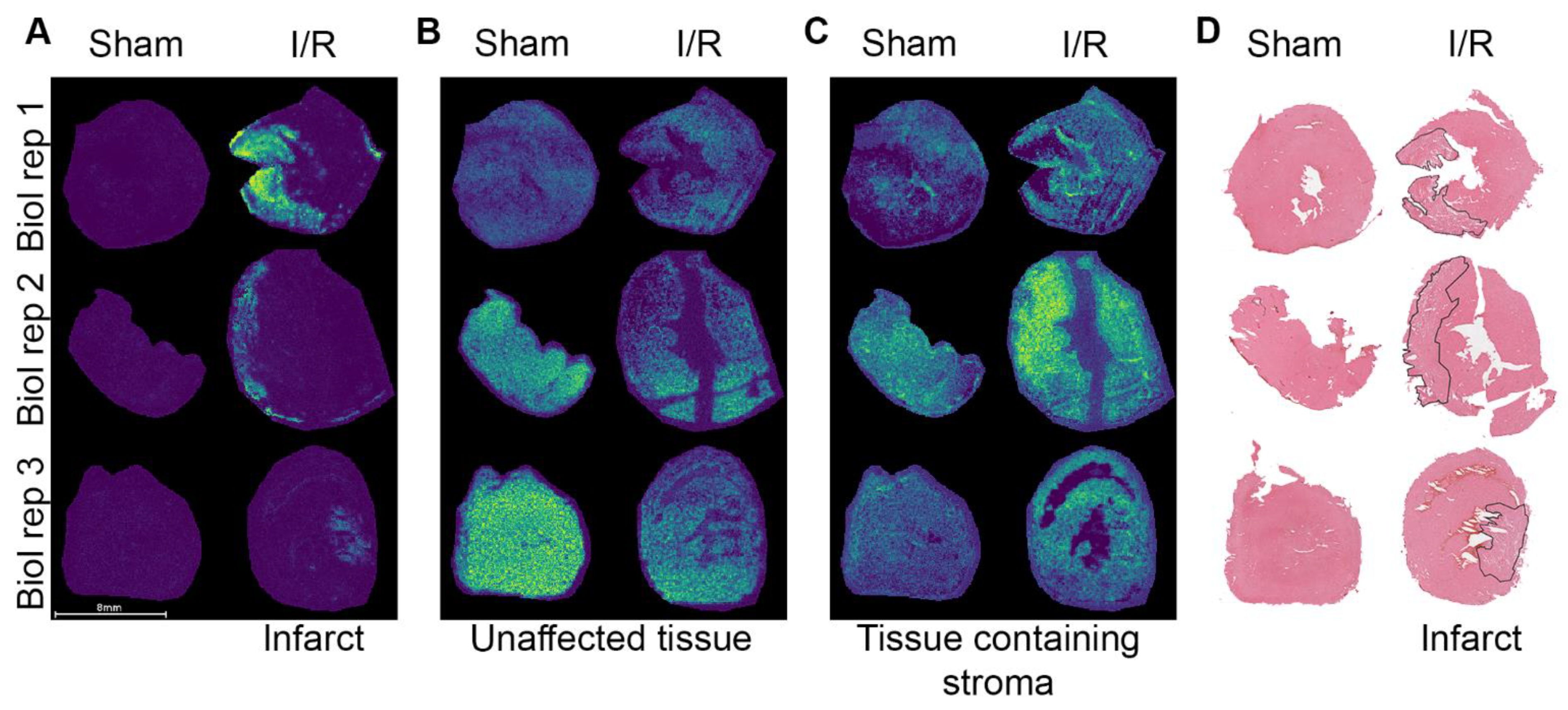
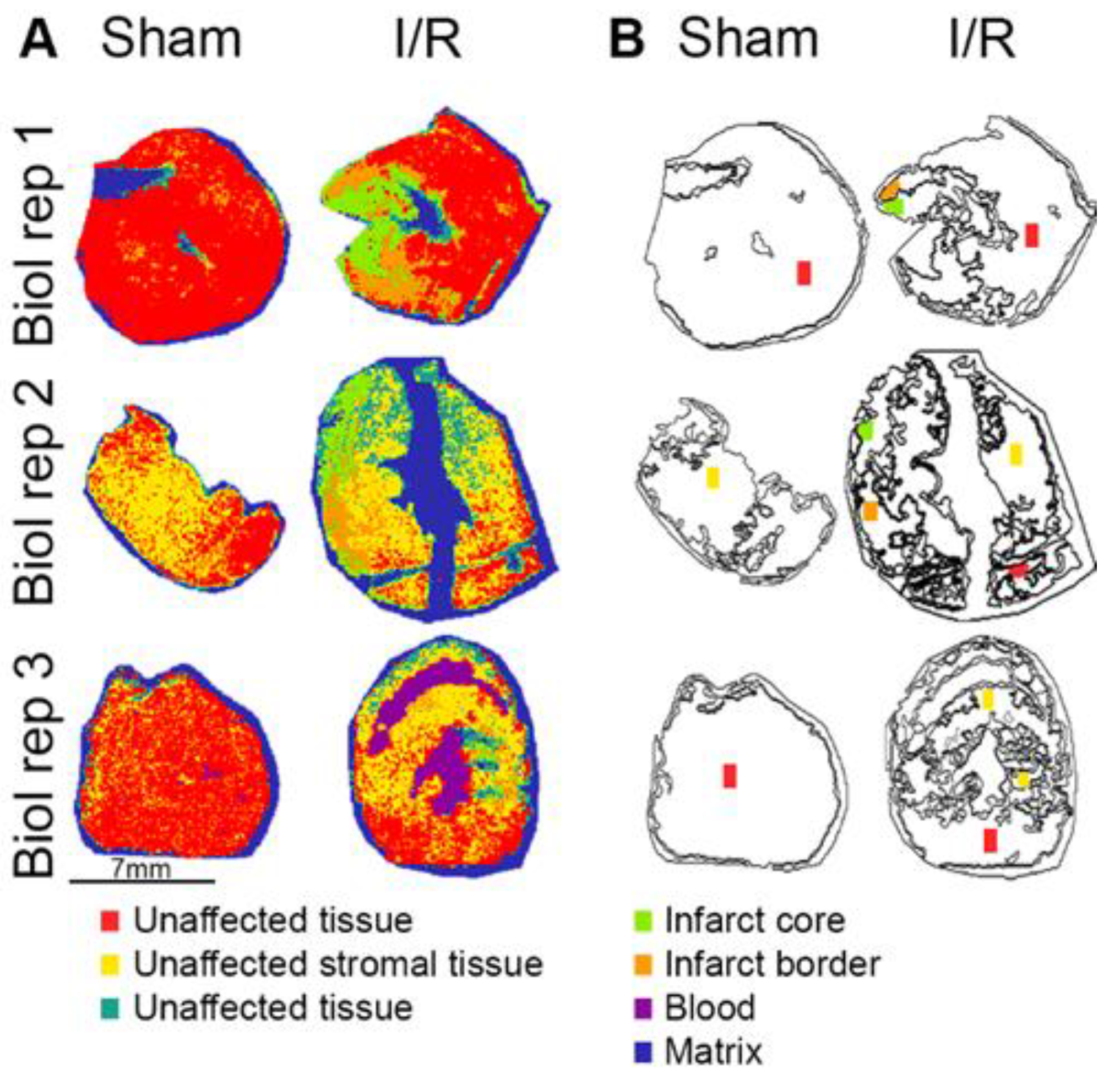
2.2. Spatial Distribution of Proteins in the I/R Heart
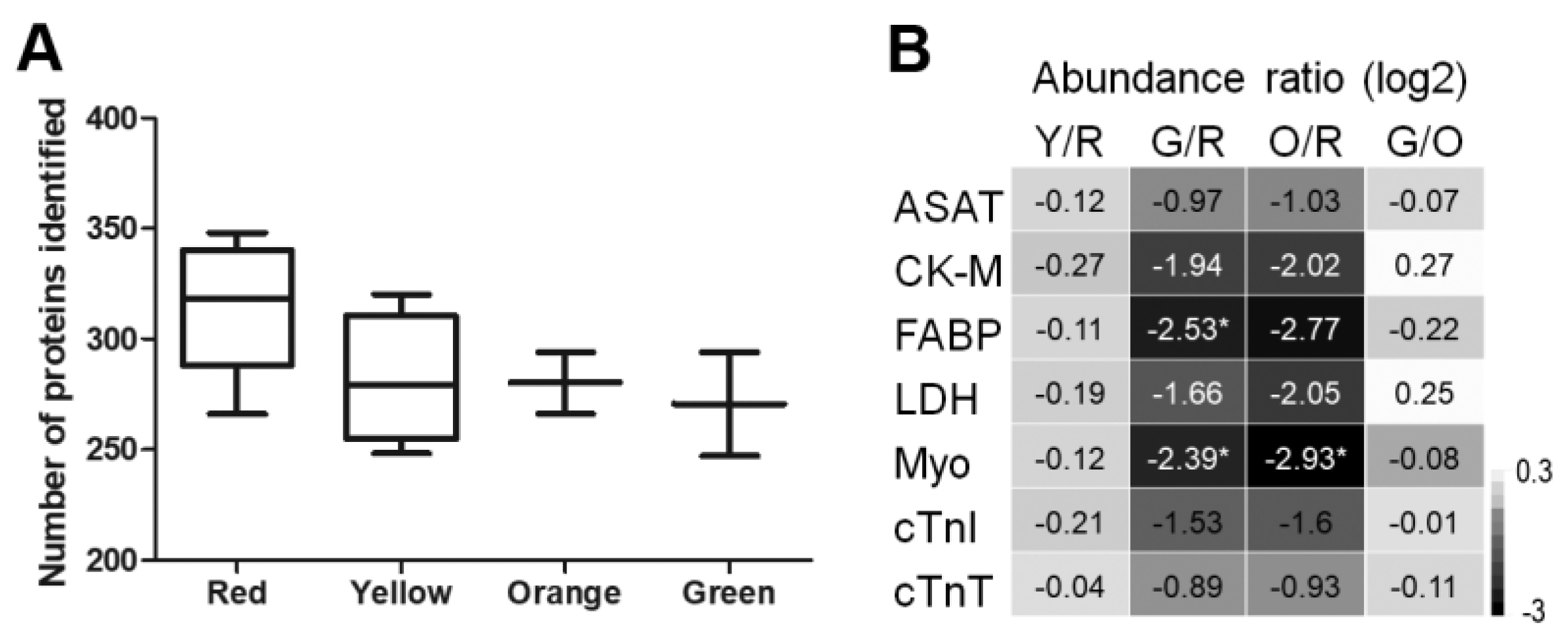
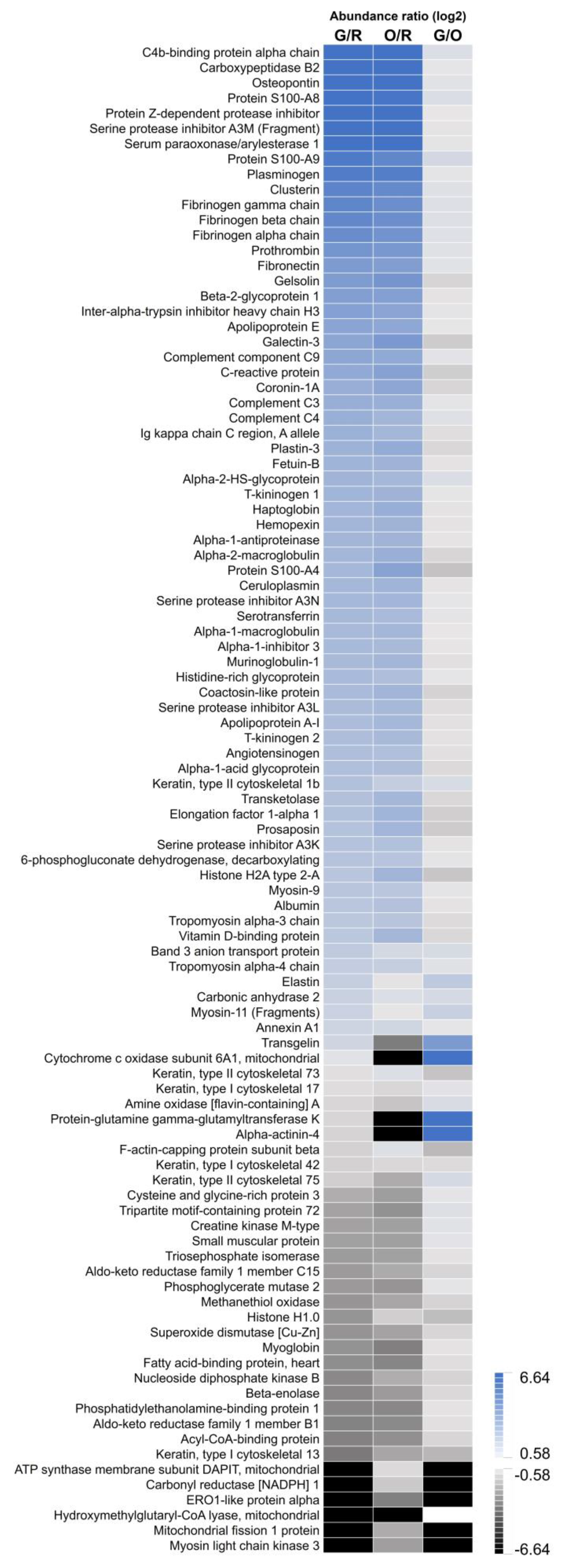
2.3. Image-Guided Proteomics Revealed Up- and Downregulation of Specific Proteins in the Infarct Area
3. Discussion
4. Materials and Methods
4.1. Chemicals and Solvents
4.2. Tissue and Blood Samples
4.3. Clinical Chemistry Measurements
4.4. Intact Protein MALDI Mass Spectrometry Imaging
4.5. MSI Data Analysis
4.6. Laser Capture Microdissection
4.7. Sample Processing for Proteomics
4.8. LC-MS/MS Anasis
4.9. Proteomics Data Analysis
4.10. Haematoxylin and Eosin (H&E) Staining
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yellon, D.M.; Hausenloy, D.J. Mechanisms of disease: Myocardial reperfusion injury. N. Engl. J. Med. 2007, 357, 1121–1135. [Google Scholar] [CrossRef] [PubMed]
- Braunwald, E.; Kloner, R.A. Myocardial reperfusion: A double-edged sword? J. Clin. Investig. 1985, 76, 1713–1719. [Google Scholar] [CrossRef]
- Frangogiannis, N.G. The inflammatory response in myocardial injury, repair, and remodelling. Nat. Rev. Cardiol. 2014, 11, 255–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boros, P.; Bromberg, J.S. New cellular and molecular immune pathways in ischemia/reperfusion injury. Am. J. Transplant. 2006, 6, 652–658. [Google Scholar] [CrossRef]
- Konstantinidis, K.; Whelan, R.S.; Kitsis, R.N. Mechanisms of cell death in heart disease. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1552–1562. [Google Scholar] [CrossRef] [Green Version]
- Sutton, M.G.; Sharpe, N. Left ventricular remodeling after myocardial infarction: Pathophysiology and therapy. Circulation 2000, 101, 2981–2988. [Google Scholar] [CrossRef] [PubMed]
- Binek, A.; Fernandez-Jimenez, R.; Jorge, I.; Camafeita, E.; Lopez, J.A.; Bagwan, N.; Galán-Arriola, C.; Pun, A.; Aguero, J.; Fuster, V.; et al. Proteomic footprint of myocardial ischemia/reperfusion injury: Longitudinal study of the at-risk and remote regions in the pig model. Sci. Rep. 2017, 7, 12343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Wang, C.; Li, T.; Ma, L.; Fan, F.; Jin, Y.; Shen, J. The whole transcriptome and proteome changes in the early stage of myocardial infarction. Cell Death Discov. 2019, 5, 73. [Google Scholar] [CrossRef] [Green Version]
- De Celle, T.; Vanrobaeys, F.; Lijnen, P.; Blankesteijn, W.M.; Heeneman, S.; Van Beeumen, J.; Devreese, B.; Smits, J.F.; Janssen, B.J. Alterations in mouse cardiac proteome after in vivo myocardial infarction: Permanent ischaemia versus ischaemia-reperfusion. Exp. Physiol. 2005, 90, 593–606. [Google Scholar] [CrossRef] [Green Version]
- Migrino, R.Q.; Zhu, X.; Morker, M.; Brahmbhatt, T.; Bright, M.; Zhao, M. Myocardial dysfunction in the periinfarct and remote regions following anterior infarction in rats quantified by 2D radial strain echocardiography: An observational cohort study. Cardiovasc. Ultrasound 2008, 6, 17. [Google Scholar] [CrossRef]
- Kuhn, D.E.; Roy, S.; Radtke, J.; Khanna, S.; Sen, C.K. Laser microdissection and capture of pure cardiomyocytes and fibroblasts from infarcted heart regions: Perceived hyperoxia induces p21 in peri-infarct myocytes. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H1245–H1253. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Gregorich, Z.R.; Cai, W.; Zhang, P.; Young, B.; Gu, Y.; Zhang, J.; Ge, Y. Quantitative Proteomics and Immunohistochemistry Reveal Insights into Cellular and Molecular Processes in the Infarct Border Zone One Month after Myocardial Infarction. J. Proteome Res. 2017, 16, 2101–2112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mezger, S.T.P.; Mingels, A.M.A.; Bekers, O.; Cillero-Pastor, B.; Heeren, R.M.A. Trends in mass spectrometry imaging for cardiovascular diseases. Anal. Bioanal. Chem. 2019, 411, 3709–3720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugiura, Y.; Katsumata, Y.; Sano, M.; Honda, K.; Kajimura, M.; Fukuda, K.; Suematsu, M. Visualization of in vivo metabolic flows reveals accelerated utilization of glucose and lactate in penumbra of ischemic heart. Sci. Rep. 2016, 6, 32361. [Google Scholar] [CrossRef]
- Khan, A.A.; Bararpour, N.; Gorka, M.; Joye, T.; Morel, S.; Montessuit, C.A.; Grabherr, S.; Fracasso, T.; Augsburger, M.; Kwak, B.R.; et al. Detecting early myocardial ischemia in rat heart by MALDI imaging mass spectrometry. Sci. Rep. 2021, 11, 5135. [Google Scholar] [CrossRef]
- Menger, R.F.; Stutts, W.L.; Anbukumar, D.S.; Bowden, J.A.; Ford, D.A.; Yost, R.A. MALDI mass spectrometric imaging of cardiac tissue following myocardial infarction in a rat coronary artery ligation model. Anal. Chem. 2012, 84, 1117–1125. [Google Scholar] [CrossRef] [Green Version]
- Kaya, I.; Sämfors, S.; Levin, M.; Borén, J.; Fletcher, J.S. Multimodal MALDI Imaging Mass Spectrometry Reveals Spatially Correlated Lipid and Protein Changes in Mouse Heart with Acute Myocardial Infarction. J. Am. Soc. Mass Spectrom. 2020, 31, 2133–2142. [Google Scholar] [CrossRef]
- Yajima, Y.; Hiratsuka, T.; Kakimoto, Y.; Ogawa, S.; Shima, K.; Yamazaki, Y.; Yoshikawa, K.; Tamaki, K.; Tsuruyama, T. Region of Interest analysis using mass spectrometry imaging of mitochondrial and sarcomeric proteins in acute cardiac infarction tissue. Sci. Rep. 2018, 8, 7493. [Google Scholar] [CrossRef] [Green Version]
- Lefcoski, S.; Kew, K.; Reece, S.; Torres, M.J.; Parks, J.; Reece, S.; Brás, L.E.D.C.; Virag, J.A.I. Anatomical-Molecular Distribution of EphrinA1 in Infarcted Mouse Heart Using MALDI Mass Spectrometry Imaging. J. Am. Soc. Mass Spectrom. 2018, 29, 527–534. [Google Scholar] [CrossRef]
- Longuespée, R.; Alberts, D.; Pottier, C.; Smargiasso, N.; Mazzucchelli, G.; Baiwir, D.; Kriegsmann, M.; Herfs, M.; Kriegsmann, J.; Delvenne, P.; et al. A laser microdissection-based workflow for FFPE tissue microproteomics: Important considerations for small sample processing. Methods 2016, 104, 154–162. [Google Scholar] [CrossRef]
- Dewez, F.; Oejten, J.; Henkel, C.; Hebeler, R.; Neuweger, H.; De Pauw, E.; Heeren, R.M.A.; Balluff, B. MS Imaging-Guided Microproteomics for Spatial Omics on a Single Instrument. Proteomics 2020, 20, e1900369. [Google Scholar] [CrossRef]
- Mezger, S.T.P.; Mingels, A.M.A.; Bekers, O.; Heeren, R.M.A.; Cillero-Pastor, B. Mass Spectrometry Spatial-Omics on a Single Conductive Slide. Anal. Chem. 2021, 93, 2527–2533. [Google Scholar] [CrossRef] [PubMed]
- Herman, E.; Knapton, A.; Rosen, E.; Zhang, J.; Estis, J.; Agee, S.J.; Lu, Q.-A.; Todd, J.A.; Lipshultz, S.E. Baseline serum cardiac troponin I concentrations in Sprague-Dawley, spontaneous hypertensive, Wistar, Wistar-Kyoto, and Fisher rats as determined with an ultrasensitive immunoassay. Toxicol. Pathol. 2011, 39, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Serfass, R.C.; Mackey-Bojack, S.M.; Kelly, K.L.; Titus, J.L.; Apple, F.S. Cardiac troponin T alterations in myocardium and serum of rats after stressful, prolonged intense exercise. J. Appl. Physiol. (1985) 2000, 88, 1749–1755. [Google Scholar] [CrossRef] [PubMed]
- Chaurand, P.; Cornett, D.S.; Caprioli, R.M. Caprioli, Molecular imaging of thin mammalian tissue sections by mass spectrometry. Curr. Opin. Biotechnol. 2006, 17, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Amann, J.M.; Chaurand, P.; Gonzalez, A.; Mobley, J.A.; Massion, P.P.; Carbone, D.P.; Caprioli, R.M. Selective profiling of proteins in lung cancer cells from fine-needle aspirates by matrix-assisted laser desorption ionization time-of-flight mass spectrometry. Clin. Cancer Res. 2006, 12, 5142–5150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heusch, G. Myocardial ischaemia-reperfusion injury and cardioprotection in perspective. Nat. Rev. Cardiol. 2020, 17, 773–789. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.C.; Pereira, C.; Fonseca, A.C.R.G.; Pinto-Do-Ó, P.; Nascimento, D.S. Bearing My Heart: The Role of Extracellular Matrix on Cardiac Development, Homeostasis, and Injury Response. Front. Cell Dev. Biol. 2020, 8, 621644. [Google Scholar] [CrossRef]
- Jourdan-LeSaux, C.; Zhang, J.; Lindsey, M.L. Lindsey, Extracellular matrix roles during cardiac repair. Life Sci. 2010, 87, 391–400. [Google Scholar] [CrossRef] [Green Version]
- Zhong, W.; Sun, B.; Gao, W.; Qin, Y.; Zhang, H.; Huai, L.; Tang, Y.; Liang, Y.; He, L.; Zhang, X.; et al. Salvianolic acid A targeting the transgelin-actin complex to enhance vasoconstriction. eBioMedicine 2018, 37, 246–258. [Google Scholar] [CrossRef]
- Shu, Y.N.; Zhang, F.; Bi, W.; Dong, L.H.; Zhang, D.D.; Chen, R.; Lv, P.; Xie, X.L.; Lin, Y.L.; Xue, Z.Y.; et al. SM22alpha inhibits vascular inflammation via stabilization of IkappaBalpha in vascular smooth muscle cells. J. Mol. Cell Cardiol. 2015, 84, 191–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nossuli, T.O.; Lakshminarayanan, V.; Baumgarten, G.; Taffet, G.E.; Ballantyne, C.M.; Michael, L.H.; Entman, M.L. A chronic mouse model of myocardial ischemia-reperfusion: Essential in cytokine studies. Am. J. Physiol. Heart Circ. Physiol. 2000, 278, H1049–H1055. [Google Scholar] [CrossRef] [PubMed]
- Katagiri, T. Changes of cardiac structural proteins in myocardial infarction. Jpn. Heart J. 1977, 18, 711–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piga, I.; Heijs, B.; Nicolardi, S.; Giusti, L.; Marselli, L.; Marchetti, P.; Mazzoni, M.R.; Lucacchini, A.; McDonnell, L.A. Ultra-high resolution MALDI-FTICR-MSI analysis of intact proteins in mouse and human pancreas tissue. Int. J. Mass Spectrom. 2019, 437, 10–16. [Google Scholar] [CrossRef]
- Dilillo, M.; Ait-Belkacem, R.; Esteve, C.; Pellegrini, D.; Nicolardi, S.; Costa, M.; Vannini, E.; de Graaf, E.L.; Caleo, M.; McDonnell, L.A. Ultra-High Mass Resolution MALDI Imaging Mass Spectrometry of Proteins and Metabolites in a Mouse Model of Glioblastoma. Sci. Rep. 2017, 7, 603. [Google Scholar] [CrossRef] [Green Version]
- Angel, P.M.; Comte-Walters, S.; Ball, L.E.; Talbot, K.; Mehta, A.; Brockbank, K.G.; Drake, R.R. Mapping Extracellular Matrix Proteins in Formalin-Fixed, Paraffin-Embedded Tissues by MALDI Imaging Mass Spectrometry. J. Proteome Res. 2018, 17, 635–646. [Google Scholar] [CrossRef]
- Clift, C.L.; Drake, R.R.; Mehta, A.; Angel, P.M. Multiplexed imaging mass spectrometry of the extracellular matrix using serial enzyme digests from formalin-fixed paraffin-embedded tissue sections. Anal. Bioanal. Chem. 2021, 413, 2709–2719. [Google Scholar] [CrossRef]
- Clift, C.L.; McLaughlin, S.; Muñoz, M.; Suuronen, E.J.; Rotstein, B.H.; Mehta, A.S.; Drake, R.R.; Alarcon, E.I.; Angel, P.M. Evaluation of Therapeutic Collagen-Based Biomaterials in the Infarcted Mouse Heart by Extracellular Matrix Targeted MALDI Imaging Mass Spectrometry. J. Am. Soc. Mass Spectrom. 2021, 32, 2746–2754. [Google Scholar] [CrossRef]
- Harouki, N.; Nicol, L.; Remy-Jouet, I.; Henry, J.P.; Dumesnil, A.; Lejeune, A.; Renet, S.; Golding, F.; Djerada, Z.; Wecker, D.; et al. The IL-1beta Antibody Gevokizumab Limits Cardiac Remodeling and Coronary Dysfunction in Rats With Heart Failure. JACC Basic Transl. Sci. 2017, 2, 418–430. [Google Scholar] [CrossRef]
- Richard, V.; Kaeffer, N.; Tron, C.; Thuillez, C. Ischemic preconditioning protects against coronary endothelial dysfunction induced by ischemia and reperfusion. Circulation 1994, 89, 1254–1261. [Google Scholar] [CrossRef]
- Strohalm, M.; Hassman, M.; Košata, B.; Kodíček, M. mMass data miner: An open source alternative for mass spectrometric data analysis. Rapid Commun. Mass Spectrom. 2008, 22, 905–908. [Google Scholar] [CrossRef] [PubMed]
- Verbeeck, N.; Caprioli, R.M.; Van De Plas, R. Unsupervised machine learning for exploratory data analysis in imaging mass spectrometry. Mass Spectrom. Rev. 2020, 39, 245–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, K.H.; O’Malley, A.J.; Mauri, L. Receiver-operating characteristic analysis for evaluating diagnostic tests and predictive models. Circulation 2007, 115, 654–657. [Google Scholar] [CrossRef] [Green Version]
- Dewez, F.; Martin-Lorenzo, M.; Herfs, M.; Baiwir, D.; Mazzucchelli, G.; De Pauw, E.; Heeren, R.M.; Balluff, B. Precise co-registration of mass spectrometry imaging, histology, and laser microdissection-based omics. Anal. Bioanal. Chem. 2019, 411, 5647–5653. [Google Scholar] [CrossRef] [Green Version]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’Ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [Green Version]
- Bankhead, P.; Loughrey, M.B.; Fernández, J.A.; Dombrowski, Y.; McArt, D.G.; Dunne, P.D.; McQuaid, S.; Gray, R.T.; Murray, L.J.; Coleman, H.G.; et al. QuPath: Open source software for digital pathology image analysis. Sci. Rep. 2017, 7, 16878. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Upregulated Pathways | Overlap | Combined Score |
|---|---|---|
| Platelet degranulation | 16/105 | 5135.92 |
| Response to elevated platelet cytosolic Ca2+ | 16/110 | 4791.00 |
| GRB2:SOS provides linkage to MAPK signaling for Integrins | 4/15 | 2403.01 |
| p130Cas linkage to MAPK signaling for integrins | 4/15 | 2403.01 |
| Platelet activation, signaling and aggregation | 17/253 | 1679.42 |
| Common pathway of fibrin clot formation | 4/22 | 1320.54 |
| Terminal pathway of complement | 2/8 | 1089.78 |
| Platelet aggregation (Plug Formation) | 5/37 | 1069.55 |
| Formation of fibrin clot (Clotting Cascade) | 5/39 | 990.15 |
| Integrin alphaIIb beta3 signaling | 4/27 | 973.26 |
| Upregulated Pathways | Overlap | Combined Score |
|---|---|---|
| RHO GTPases activate CIT | 1/16 | 1422.78 |
| RHO GTPases Activate ROCKs | 1/17 | 1318.68 |
| RHO GTPases activate PAKs | 1/21 | 1012.63 |
| Nephrin interactions | 1/22 | 955.53 |
| Sema4D induced cell migration and growth-cone collapse | 1/24 | 857.29 |
| Sema4D in semaphorin signaling | 1/27 | 740.22 |
| Smooth Muscle Contraction | 1/33 | 576.31 |
| EPHA-mediated growth cone collapse | 1/34 | 555.22 |
| Elastic fibre formation | 1/41 | 439.30 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mezger, S.T.P.; Mingels, A.M.A.; Soulié, M.; Peutz-Kootstra, C.J.; Bekers, O.; Mulder, P.; Heeren, R.M.A.; Cillero-Pastor, B. Protein Alterations in Cardiac Ischemia/Reperfusion Revealed by Spatial-Omics. Int. J. Mol. Sci. 2022, 23, 13847. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232213847
Mezger STP, Mingels AMA, Soulié M, Peutz-Kootstra CJ, Bekers O, Mulder P, Heeren RMA, Cillero-Pastor B. Protein Alterations in Cardiac Ischemia/Reperfusion Revealed by Spatial-Omics. International Journal of Molecular Sciences. 2022; 23(22):13847. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232213847
Chicago/Turabian StyleMezger, Stephanie T. P., Alma M. A. Mingels, Matthieu Soulié, Carine J. Peutz-Kootstra, Otto Bekers, Paul Mulder, Ron M. A. Heeren, and Berta Cillero-Pastor. 2022. "Protein Alterations in Cardiac Ischemia/Reperfusion Revealed by Spatial-Omics" International Journal of Molecular Sciences 23, no. 22: 13847. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232213847