
Inhibition of Calpain Attenuates Degeneration of Substantia Nigra Neurons in the Rotenone Rat Model of Parkinson’s Disease

 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Increased Expression of Phosphorylated α-syn and Induction of Gliosis in Dorsal Striatum of Rotenone-Parkinsonian Rats
2.2. Inhibition of Calpain Prevents Rotenone-Induced Loss of DA Neurons in the Substantia Nigra of Rotenone-Parkinsonian Rats
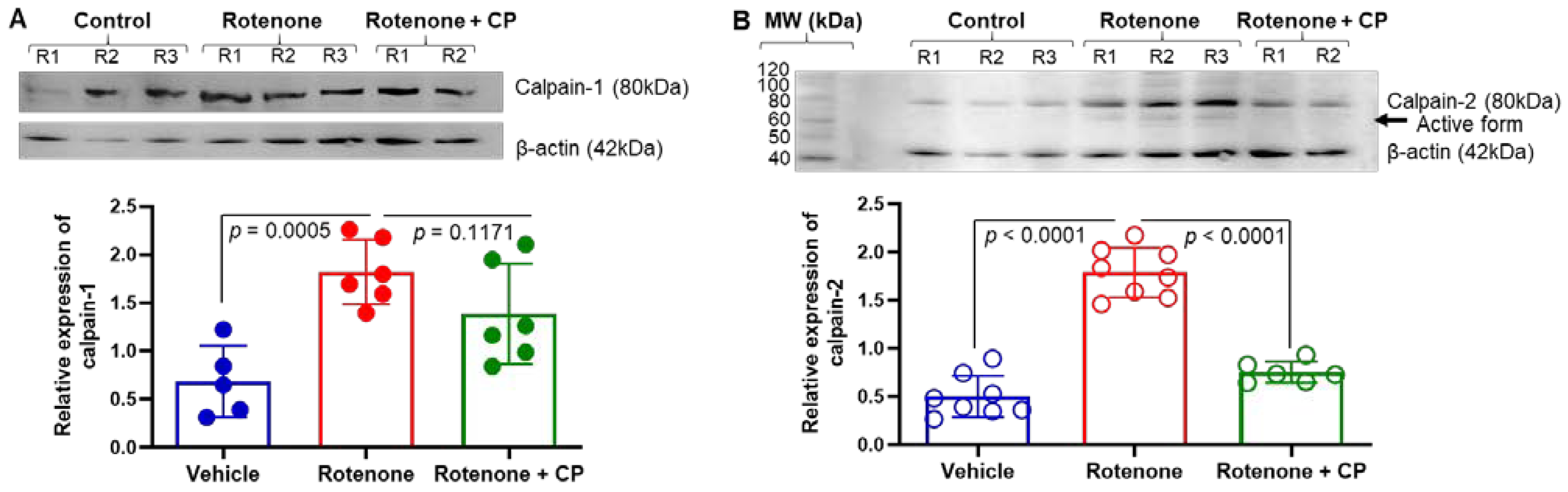
2.3. Increased Expression of Calpain-1 Was Detected in Substantia Nigra DA Neurons of Rotenone-Parkinsonian Rats
2.4. Calpeptin Treatment Reduced the Enhanced Expression of Calpain-2 in SN DA Neurons of Rotenone-Parkinsonian Rats
2.5. Calpeptin Treatment Attenuated Reactive Gliosis in the Dorsal Striatum of Rotenone-Parkinsonian Rats
2.6. Calpain Inhibitor Attenuates Rotenone-Induced Calpain-2 in Dorsal Striatum of Rotenone-Parkinsonian Rats
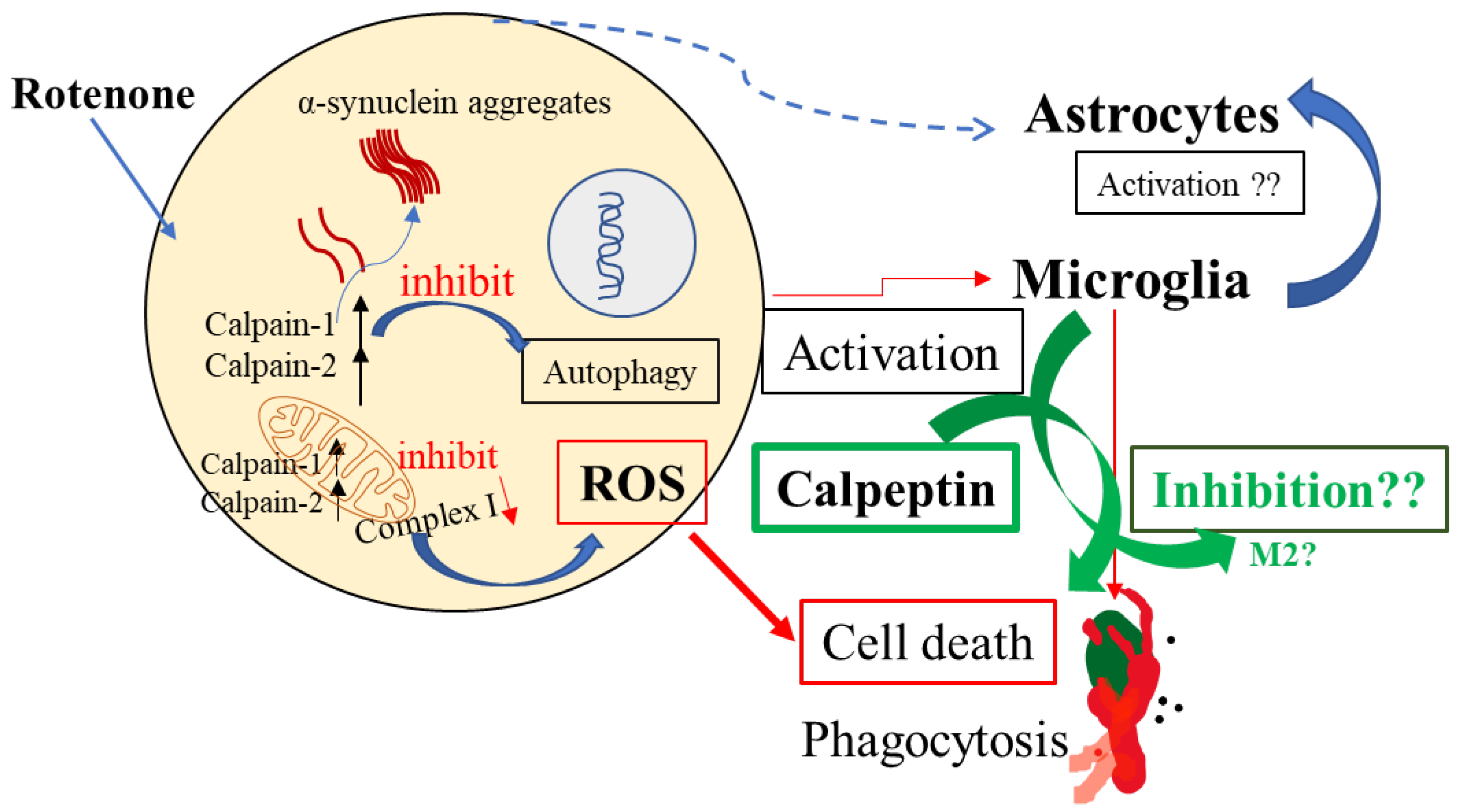
3. Discussion
4. Material and Methods
4.1. Animals
4.2. Rotenone Administration
4.3. Tissue Processing
4.4. Immunohistochemistry
4.5. Western Blot Analysis
4.6. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AD | Alzheimer disease |
| ANOVA | Analysis of variance |
| CP | Calpeptin |
| DMSO | Dimethylsulfoxide |
| EDTA | Ethylenediaminetetraacetic acid |
| GFAP | Glial fibrillary acidic protein |
| IACUC | Institutional Animal Care and Use Committee |
| PBS | Phosphate-buffered saline |
| PD | Parkinson’s disease |
| ROS | Reactive oxygen species |
| SN | Substantia nigra |
| SNpc | Substantia nigra pars compacta |
| SNpr | Substantia nigra pars reticulata |
| TH | Tyrosine hydroxylase |
References
- Wong, Y.C.; Krainc, D. alpha-synuclein toxicity in neurodegeneration: Mechanism and therapeutic strategies. Nat. Med. 2017, 23, 1–13. [Google Scholar] [CrossRef]
- Zaman, V.; Shields, D.C.; Shams, R.; Drasites, K.P.; Matzelle, D.; Haque, A.; Banik, N.L. Cellular and molecular pathophysiology in the progression of Parkinson’s disease. Metab. Brain Dis. 2021, 36, 815–827. [Google Scholar] [PubMed]
- Liang, C.L.; Sinton, C.M.; German, D.C. Midbrain dopaminergic neurons in the mouse: Co-localization with Calbindin-D28K and calretinin. Neuroscience 1996, 75, 523–533. [Google Scholar] [CrossRef]
- Nemoto, C.; Hida, T.; Arai, R. Calretinin and calbindin-D28k in dopaminergic neurons of the rat midbrain: A triple-labeling immunohistochemical study. Brain Res. 1999, 846, 129–136. [Google Scholar] [CrossRef]
- German, D.C.; Manaye, K.F.; Sonsalla, P.K.; Brooks, B.A. Midbrain dopaminergic cell loss in Parkinson’s disease and MPTP-induced parkinsonism: Sparing of calbindin-D28k-containing cells. Ann. N. Y. Acad. Sci. 1992, 648, 42–62. [Google Scholar] [CrossRef]
- Vosler, P.S.; Sun, D.; Wang, S.; Gao, Y.; Kintner, D.B.; Signore, A.P.; Cao, G.; Chen, J. Calcium dysregulation induces apoptosis-inducing factor release: Cross-talk between PARP-1- and calpain-signaling pathways. Exp. Neurol. 2009, 218, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Ono, Y.; Sorimachi, H. Calpains: An elaborate proteolytic system. Biochim. Biophys. Acta 2012, 1824, 224–236. [Google Scholar] [CrossRef] [Green Version]
- Grant, R.J.; Sellings, L.H.; Crocker, S.J.; Melloni, E.; Park, D.S.; Clarke, P.B. Effects of calpain inhibition on dopaminergic markers and motor function following intrastriatal 6-hydroxydopamine administration in rats. Neuroscience 2009, 158, 558–569. [Google Scholar] [CrossRef]
- Gao, A.; McCoy, H.M.; Zaman, V.; Shields, D.C.; Banik, N.L.; Haque, A. Calpain activation and progression of inflammatory cycles in Parkinson’s disease. Front. Biosci. 2022, 27, 20. [Google Scholar] [CrossRef]
- Mouatt-Prigent, A.; Karlsson, J.O.; Yelnik, J.; Agid, Y.; Hirsch, E.C. Calpastatin immunoreactivity in the monkey and human brain of control subjects and patients with Parkinson’s disease. J. Comp. Neurol. 2000, 419, 175–192. [Google Scholar] [CrossRef]
- Samantaray, S.; Knaryan, V.H.; Angelo, M.D.R.; Woodward, J.J.; Shields, D.C.; Azuma, M.; Inoue, J.; Ray, S.K.; Banik, N.L. Cell-Permeable Calpain Inhibitor SJA6017 Provides Functional Protection to Spinal Motoneurons Exposed to MPP. Neurotox Res. 2020, 38, 640–649. [Google Scholar] [CrossRef] [PubMed]
- Shams, R.; Banik, N.L.; Haque, A. Calpain in the cleavage of alpha-synuclein and the pathogenesis of Parkinson’s disease. Prog. Mol. Biol. Transl. Sci. 2019, 167, 107–124. [Google Scholar] [PubMed]
- Dufty, B.M.; Warner, L.R.; Hou, S.T.; Jiang, S.X.; Gomez-Isla, T.; Leenhouts, K.M.; Oxford, J.T.; Feany, M.B.; Masliah, E.; Rohn, T.T. Calpain-cleavage of alpha-synuclein: Connecting proteolytic processing to disease-linked aggregation. Am. J. Pathol. 2007, 170, 1725–1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, V.; Junn, E.; Mouradian, M.M. The role of oxidative stress in Parkinson’s disease. J. Park. Dis. 2013, 3, 461–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, R.G.; Waymire, J.C.; Lin, E.; Liu, J.J.; Guo, F.; Zigmond, M.J. A role for alpha-synuclein in the regulation of dopamine biosynthesis. J. Neurosci. 2002, 22, 3090–3099. [Google Scholar] [CrossRef] [PubMed]
- Alerte, T.N.; Akinfolarin, A.A.; Friedrich, E.E.; Mader, S.A.; Hong, C.S.; Perez, R.G. Alpha-synuclein aggregation alters tyrosine hydroxylase phosphorylation and immunoreactivity: Lessons from viral transduction of knockout mice. Neurosci. Lett. 2008, 435, 24–29. [Google Scholar] [CrossRef] [Green Version]
- Betarbet, R.; Sherer, T.B.; MacKenzie, G.; Garcia-Osuna, M.; Panov, A.V.; Greenamyre, J.T. Chronic systemic pesticide exposure reproduces features of Parkinson’s disease. Nat. Neurosci. 2000, 3, 1301–1306. [Google Scholar] [CrossRef] [Green Version]
- Betarbet, R.; Canet-Aviles, R.M.; Sherer, T.B.; Mastroberardino, P.G.; McLendon, C.; Kim, J.H.; Lund, S.; Na, H.M.; Taylor, G.; Bence, N.F.; et al. Intersecting pathways to neurodegeneration in Parkinson’s disease: Effects of the pesticide rotenone on DJ-1, alpha-synuclein, and the ubiquitin-proteasome system. Neurobiol. Dis. 2006, 22, 404–420. [Google Scholar] [CrossRef]
- Cannon, J.R.; Tapias, V.; Na, H.M.; Honick, A.S.; Drolet, R.E.; Greenamyre, J.T. A highly reproducible rotenone model of Parkinson’s disease. Neurobiol. Dis. 2009, 34, 279–290. [Google Scholar] [CrossRef] [Green Version]
- Sherer, T.B.; Kim, J.H.; Betarbet, R.; Greenamyre, J.T. Subcutaneous rotenone exposure causes highly selective dopaminergic degeneration and alpha-synuclein aggregation. Exp. Neurol. 2003, 179, 9–16. [Google Scholar] [CrossRef]
- Greenamyre, J.T.; Betarbet, R.; Sherer, T.B. The rotenone model of Parkinson’s disease: Genes, environment and mitochondria. Park. Relat. Disord. 2003, 9 (Suppl. S2), S59–S64. [Google Scholar] [CrossRef]
- Liu, H.F.; Ho, P.W.; Leung, G.C.; Lam, C.S.; Pang, S.Y.; Li, L.; Kung, M.H.; Ramsden, D.B.; Ho, S.L. Combined LRRK2 mutation, aging and chronic low dose oral rotenone as a model of Parkinson’s disease. Sci. Rep. 2017, 7, 40887. [Google Scholar] [CrossRef] [Green Version]
- Samantaray, S.; Knaryan, V.H.; Shields, D.C.; Cox, A.A.; Haque, A.; Banik, N.L. Inhibition of Calpain Activation Protects MPTP-Induced Nigral and Spinal Cord Neurodegeneration, Reduces Inflammation, and Improves Gait Dynamics in Mice. Mol. Neurobiol. 2015, 52, 1054–1066. [Google Scholar] [CrossRef] [Green Version]
- Haque, A.; Samantaray, S.; Knaryan, V.H.; Capone, M.; Hossain, A.; Matzelle, D.; Chandran, R.; Shields, D.C.; Farrand, A.Q.; Boger, H.A.; et al. Calpain mediated expansion of CD4+ cytotoxic T cells in rodent models of Parkinson’s disease. Exp. Neurol. 2020, 330, 113315. [Google Scholar] [CrossRef] [PubMed]
- Samantaray, S.; Knaryan, V.H.; Guyton, M.K.; Matzelle, D.D.; Ray, S.K.; Banik, N.L. The parkinsonian neurotoxin rotenone activates calpain and caspase-3 leading to motoneuron degeneration in spinal cord of Lewis rats. Neuroscience 2007, 146, 741–755. [Google Scholar] [CrossRef] [Green Version]
- Knaryan, V.H.; Samantaray, S.; Park, S.; Azuma, M.; Inoue, J.; Banik, N.L. SNJ-1945, a calpain inhibitor, protects SH-SY5Y cells against MPP(+) and rotenone. J. Neurochem. 2014, 130, 280–290. [Google Scholar] [CrossRef] [Green Version]
- Canerina-Amaro, A.; Pereda, D.; Diaz, M.; Rodriguez-Barreto, D.; Casanas-Sanchez, V.; Heffer, M.; Garcia-Esparcia, P.; Ferrer, I.; Puertas-Avendano, R.; Marin, R. Differential Aggregation and Phosphorylation of Alpha Synuclein in Membrane Compartments Associated With Parkinson Disease. Front. Neurosci. 2019, 13, 382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazaki, I.; Asanuma, M. Neuron-Astrocyte Interactions in Parkinson’s Disease. Cells 2020, 9, 2623. [Google Scholar] [CrossRef]
- Wang, Y.; Lopez, D.; Davey, P.G.; Cameron, D.J.; Nguyen, K.; Tran, J.; Marquez, E.; Liu, Y.; Bi, X.; Baudry, M. Calpain-1 and calpain-2 play opposite roles in retinal ganglion cell degeneration induced by retinal ischemia/reperfusion injury. Neurobiol. Dis. 2016, 93, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Bi, X.; Baudry, M. Calpain-2 as a therapeutic target for acute neuronal injury. Expert Opin. Targets 2018, 22, 19–29. [Google Scholar] [CrossRef]
- Baudry, M. Calpain-1 and Calpain-2 in the Brain: Dr. Jekill and Mr Hyde? Curr. Neuropharmacol. 2019, 17, 823–829. [Google Scholar] [CrossRef]
- Alluri, H.; Grimsley, M.; Anasooya Shaji, C.; Varghese, K.P.; Zhang, S.L.; Peddaboina, C.; Robinson, B.; Beeram, M.R.; Huang, J.H.; Tharakan, B. Attenuation of Blood-Brain Barrier Breakdown and Hyperpermeability by Calpain Inhibition. J. Biol. Chem. 2016, 291, 26958–26969. [Google Scholar] [CrossRef] [Green Version]
- Camins, A.; Verdaguer, E.; Folch, J.; Pallas, M. Involvement of calpain activation in neurodegenerative processes. CNS Drug. Rev. 2006, 12, 135–148. [Google Scholar] [CrossRef]
- Du, S.; Rubin, A.; Klepper, S.; Barrett, C.; Kim, Y.C.; Rhim, H.W.; Lee, E.B.; Park, C.W.; Markelonis, G.J.; Oh, T.H. Calcium influx and activation of calpain I mediate acute reactive gliosis in injured spinal cord. Exp. Neurol. 1999, 157, 96–105. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y.; Nham, A.; Sherbaf, A.; Quach, D.; Yahya, E.; Ranburger, D.; Bi, X.; Baudry, M. Calpain-2 as a therapeutic target in repeated concussion-induced neuropathy and behavioral impairment. Sci. Adv. 2020, 6, eaba5547. [Google Scholar] [CrossRef]
- Gonzalez, M.I. Calpain-dependent cleavage of GABAergic proteins during epileptogenesis. Epilepsy Res. 2019, 157, 106206. [Google Scholar] [CrossRef]
- Saito, K.; Elce, J.S.; Hamos, J.E.; Nixon, R.A. Widespread activation of calcium-activated neutral proteinase (calpain) in the brain in Alzheimer disease: A potential molecular basis for neuronal degeneration. Proc. Natl. Acad. Sci. USA 1993, 90, 2628–2632. [Google Scholar] [CrossRef] [Green Version]
- Banik, N.L.; Shields, D.C.; Ray, S.; Davis, B.; Matzelle, D.; Wilford, G.; Hogan, E.L. Role of calpain in spinal cord injury: Effects of calpain and free radical inhibitors. Ann. N. Y. Acad. Sci. 1998, 844, 131–137. [Google Scholar] [CrossRef] [Green Version]
- Yildiz-Unal, A.; Korulu, S.; Karabay, A. Neuroprotective strategies against calpain-mediated neurodegeneration. Neuropsychiatr. Dis. Treat. 2015, 11, 297–310. [Google Scholar] [CrossRef] [Green Version]
- Newcomb, J.K.; Kampfl, A.; Posmantur, R.M.; Zhao, X.; Pike, B.R.; Liu, S.J.; Clifton, G.L.; Hayes, R.L. Immunohistochemical study of calpain-mediated breakdown products to alpha-spectrin following controlled cortical impact injury in the rat. J. Neurotrauma 1997, 14, 369–383. [Google Scholar] [CrossRef]
- Ryu, M.; Nakazawa, T. Calcium and Calpain Activation. In Neuroprotection and Neuroregeneration for Retinal Diseases; Springer: Berlin/Heidelberg, Germany, 2014; pp. 13–24. [Google Scholar]
- Crocker, S.J.; Smith, P.D.; Jackson-Lewis, V.; Lamba, W.R.; Hayley, S.P.; Grimm, E.; Callaghan, S.M.; Slack, R.S.; Melloni, E.; Przedborski, S.; et al. Inhibition of calpains prevents neuronal and behavioral deficits in an MPTP mouse model of Parkinson’s disease. J. Neurosci. Off. J. Soc. Neurosci. 2003, 23, 4081–4091. [Google Scholar] [CrossRef] [Green Version]
- Banik, N.L.; Matzelle, D.; Gantt-Wilford, G.; Hogan, E.L. Role of calpain and its inhibitors in tissue degeneration and neuroprotection in spinal cord injury. Ann. N. Y. Acad. Sci. 1997, 825, 120–127. [Google Scholar] [CrossRef]
- Ray, S.K.; Banik, N.L. Calpain and its involvement in the pathophysiology of CNS injuries and diseases: Therapeutic potential of calpain inhibitors for prevention of neurodegeneration. Curr. Drug Targets. CNS Neurol. Disord. 2003, 2, 173–189. [Google Scholar] [CrossRef]
- Ray, S.K.; Wilford, G.G.; Ali, S.F.; Banik, N.L. Calpain upregulation in spinal cords of mice with 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-induced Parkinson’s disease. Ann. N. Y. Acad. Sci. 2000, 914, 275–283. [Google Scholar] [CrossRef]
- Samantaray, S.; Ray, S.K.; Ali, S.F.; Banik, N.L. Calpain activation in apoptosis of motoneurons in cell culture models of experimental parkinsonism. Ann. N. Y. Acad. Sci. 2006, 1074, 349–356. [Google Scholar] [CrossRef]
- Shields, D.C.; Banik, N.L. Putative role of calpain in the pathophysiology of experimental optic neuritis. Exp. Eye Res. 1998, 67, 403–410. [Google Scholar] [CrossRef]
- Shields, D.C.; Schaecher, K.E.; Hogan, E.L.; Banik, N.L. Calpain activity and expression increased in activated glial and inflammatory cells in penumbra of spinal cord injury lesion. J. Neurosci. Res. 2000, 61, 146–150. [Google Scholar] [CrossRef]
- Siman, R.; Gall, C.; Perlmutter, L.S.; Christian, C.; Baudry, M.; Lynch, G. Distribution of calpain I, an enzyme associated with degenerative activity, in rat brain. Brain Res. 1985, 347, 399–403. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y.; Bi, X.; Baudry, M. Calpain-1 and Calpain-2 in the Brain: New Evidence for a Critical Role of Calpain-2 in Neuronal Death. Cells 2020, 9, 2698. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y.; Lopez, D.; Lee, M.; Dayal, S.; Hurtado, A.; Bi, X.; Baudry, M. Protection against TBI-Induced Neuronal Death with Post-Treatment with a Selective Calpain-2 Inhibitor in Mice. J. Neurotrauma 2018, 35, 105–117. [Google Scholar] [CrossRef]
- Zhao, X.; Posmantur, R.; Kampfl, A.; Liu, S.J.; Wang, K.K.; Newcomb, J.K.; Pike, B.R.; Clifton, G.L.; Hayes, R.L. Subcellular localization and duration of mu-calpain and m-calpain activity after traumatic brain injury in the rat: A casein zymography study. J. Cereb. Blood Flow Metab. 1998, 18, 161–167. [Google Scholar] [CrossRef] [Green Version]
- Shinkai-Ouchi, F.; Shindo, M.; Doi, N.; Hata, S.; Ono, Y. Calpain-2 participates in the process of calpain-1 inactivation. Biosci. Rep. 2020, 40, BSR20200552. [Google Scholar] [CrossRef]
- Shoffner, J.M.; Watts, R.L.; Juncos, J.L.; Torroni, A.; Wallace, D.C. Mitochondrial oxidative phosphorylation defects in Parkinson’s disease. Ann. Neurol. 1991, 30, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Parker, W.D., Jr.; Boyson, S.J.; Parks, J.K. Abnormalities of the electron transport chain in idiopathic Parkinson’s disease. Ann. Neurol. 1989, 26, 719–723. [Google Scholar] [CrossRef]
- Greenamyre, J.T.; Sherer, T.B.; Betarbet, R.; Panov, A.V. Complex I and Parkinson’s disease. IUBMB Life 2001, 52, 135–141. [Google Scholar] [CrossRef]
- Betarbet, R.; Sherer, T.B.; Greenamyre, J.T. Animal models of Parkinson’s disease. Bioessays 2002, 24, 308–318. [Google Scholar]
- Samantaray, S.; Knaryan, V.H.; Shields, D.C.; Banik, N.L. Critical role of calpain in spinal cord degeneration in Parkinson’s disease. J. Neurochem. 2013, 127, 880–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blomgren, K.; Zhu, C.; Wang, X.; Karlsson, J.O.; Leverin, A.L.; Bahr, B.A.; Mallard, C.; Hagberg, H. Synergistic activation of caspase-3 by m-calpain after neonatal hypoxia-ischemia: A mechanism of “pathological apoptosis”? J. Biol. Chem. 2001, 276, 10191–10198. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.Y.; Wang, S.C.; Lei, M.; Wang, Z.; Xiong, K. Regulatory role of calpain in neuronal death. Neural Regen. Res. 2018, 13, 556–562. [Google Scholar]
- Russo, R.; Berliocchi, L.; Adornetto, A.; Varano, G.P.; Cavaliere, F.; Nucci, C.; Rotiroti, D.; Morrone, L.A.; Bagetta, G.; Corasaniti, M.T. Calpain-mediated cleavage of Beclin-1 and autophagy deregulation following retinal ischemic injury in vivo. Cell Death Dis. 2011, 2, e144. [Google Scholar] [CrossRef] [Green Version]
- Watchon, M.; Yuan, K.C.; Mackovski, N.; Svahn, A.J.; Cole, N.J.; Goldsbury, C.; Rinkwitz, S.; Becker, T.S.; Nicholson, G.A.; Laird, A.S. Calpain Inhibition Is Protective in Machado-Joseph Disease Zebrafish Due to Induction of Autophagy. J. Neurosci. 2017, 37, 7782–7794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iacopino, A.; Christakos, S.; German, D.; Sonsalla, P.K.; Altar, C.A. Calbindin-D28K-containing neurons in animal models of neurodegeneration: Possible protection from excitotoxicity. Brain Res. Mol. Brain Res. 1992, 13, 251–261. [Google Scholar] [CrossRef]
- Liang, C.L.; Sinton, C.M.; Sonsalla, P.K.; German, D.C. Midbrain dopaminergic neurons in the mouse that contain calbindin-D28k exhibit reduced vulnerability to MPTP-induced neurodegeneration. Neurodegeneration 1996, 5, 313–318. [Google Scholar] [CrossRef]
- Ahmad, F.; Das, D.; Kommaddi, R.P.; Diwakar, L.; Gowaikar, R.; Rupanagudi, K.V.; Bennett, D.A.; Ravindranath, V. Isoform-specific hyperactivation of calpain-2 occurs presymptomatically at the synapse in Alzheimer’s disease mice and correlates with memory deficits in human subjects. Sci. Rep. 2018, 8, 13119. [Google Scholar] [CrossRef] [Green Version]
- Gilhus, N.E.; Deuschl, G. Neuroinflammation—A common thread in neurological disorders. Nat. Rev. Neurol. 2019, 15, 429–430. [Google Scholar] [CrossRef]
- McGeer, P.L.; Itagaki, S.; Boyes, B.E.; McGeer, E.G. Reactive microglia are positive for HLA-DR in the substantia nigra of Parkinson’s and Alzheimer’s disease brains. Neurology 1988, 38, 1285–1291. [Google Scholar] [CrossRef]
- Kim, W.G.; Mohney, R.P.; Wilson, B.; Jeohn, G.H.; Liu, B.; Hong, J.S. Regional difference in susceptibility to lipopolysaccharide-induced neurotoxicity in the rat brain: Role of microglia. J. Neurosci. 2000, 20, 6309–6316. [Google Scholar] [CrossRef]
- Grabert, K.; Michoel, T.; Karavolos, M.H.; Clohisey, S.; Baillie, J.K.; Stevens, M.P.; Freeman, T.C.; Summers, K.M.; McColl, B.W. Microglial brain region-dependent diversity and selective regional sensitivities to aging. Nat. Neurosci. 2016, 19, 504–516. [Google Scholar] [CrossRef] [Green Version]
- Lawson, L.J.; Perry, V.H.; Dri, P.; Gordon, S. Heterogeneity in the distribution and morphology of microglia in the normal adult mouse brain. Neuroscience 1990, 39, 151–170. [Google Scholar] [CrossRef]
- Haque, A.; Das, A.; Samantaray, S.; Matzelle, D.; Capone, M.; Wallace, G.; Husarik, A.N.; Taheri, S.; Reiter, R.J.; Varma, A.; et al. Premarin Reduces Neurodegeneration and Promotes Improvement of Function in an Animal Model of Spinal Cord Injury. Int. J. Mol. Sci. 2022, 23, 2384. [Google Scholar] [CrossRef]
- Hathaway-Schrader, J.D.; Doonan, B.P.; Hossain, A.; Radwan, F.F.Y.; Zhang, L.; Haque, A. Autophagy-dependent crosstalk between GILT and PAX-3 influences radiation sensitivity of human melanoma cells. J. Cell Biochem. 2018, 119, 2212–2221. [Google Scholar] [CrossRef] [PubMed]
- Haque, A.; Drasites, K.P.; Cox, A.; Capone, M.; Myatich, A.I.; Shams, R.; Matzelle, D.; Garner, D.P.; Bredikhin, M.; Shields, D.C.; et al. Protective Effects of Estrogen via Nanoparticle Delivery to Attenuate Myelin Loss and Neuronal Death after Spinal Cord Injury. Neurochem. Res. 2021, 46, 2979–2990. [Google Scholar] [CrossRef] [PubMed]
- Radwan, F.F.; Zhang, L.; Hossain, A.; Doonan, B.P.; God, J.M.; Haque, A. Mechanisms regulating enhanced human leukocyte antigen class II-mediated CD4 + T cell recognition of human B-cell lymphoma by resveratrol. Leuk Lymphoma 2012, 53, 305–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaman, V.; Drasites, K.P.; Myatich, A.; Shams, R.; Shields, D.C.; Matzelle, D.; Haque, A.; Banik, N.L. Inhibition of Calpain Attenuates Degeneration of Substantia Nigra Neurons in the Rotenone Rat Model of Parkinson’s Disease. Int. J. Mol. Sci. 2022, 23, 13849. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232213849
Zaman V, Drasites KP, Myatich A, Shams R, Shields DC, Matzelle D, Haque A, Banik NL. Inhibition of Calpain Attenuates Degeneration of Substantia Nigra Neurons in the Rotenone Rat Model of Parkinson’s Disease. International Journal of Molecular Sciences. 2022; 23(22):13849. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232213849
Chicago/Turabian StyleZaman, Vandana, Kelsey P. Drasites, Ali Myatich, Ramsha Shams, Donald C. Shields, Denise Matzelle, Azizul Haque, and Narendra L. Banik. 2022. "Inhibition of Calpain Attenuates Degeneration of Substantia Nigra Neurons in the Rotenone Rat Model of Parkinson’s Disease" International Journal of Molecular Sciences 23, no. 22: 13849. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232213849