

Transcriptome Analysis of Two Tetraploid Potato Varieties under Water-Stress Conditions

Abstract
:1. Introduction
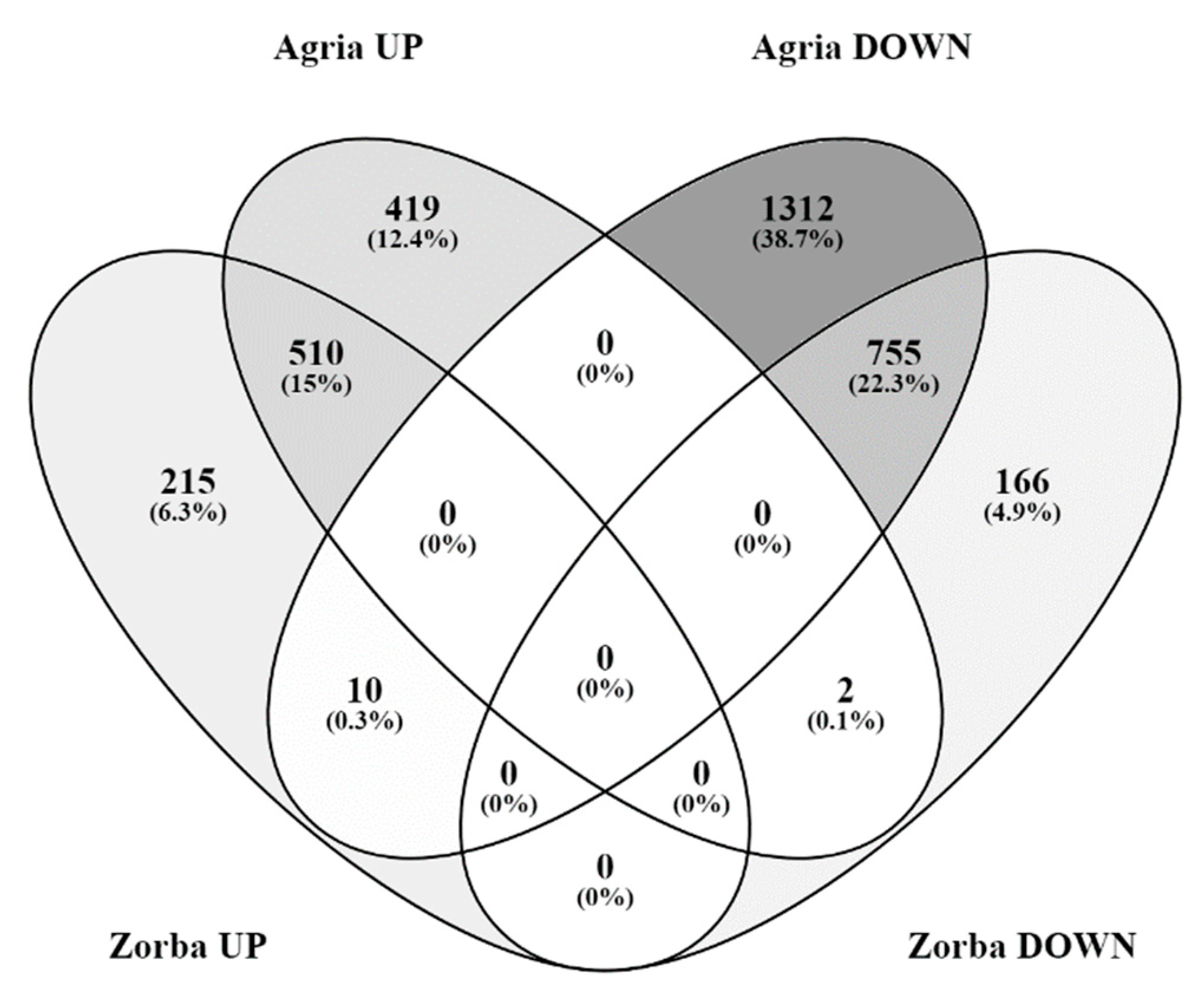
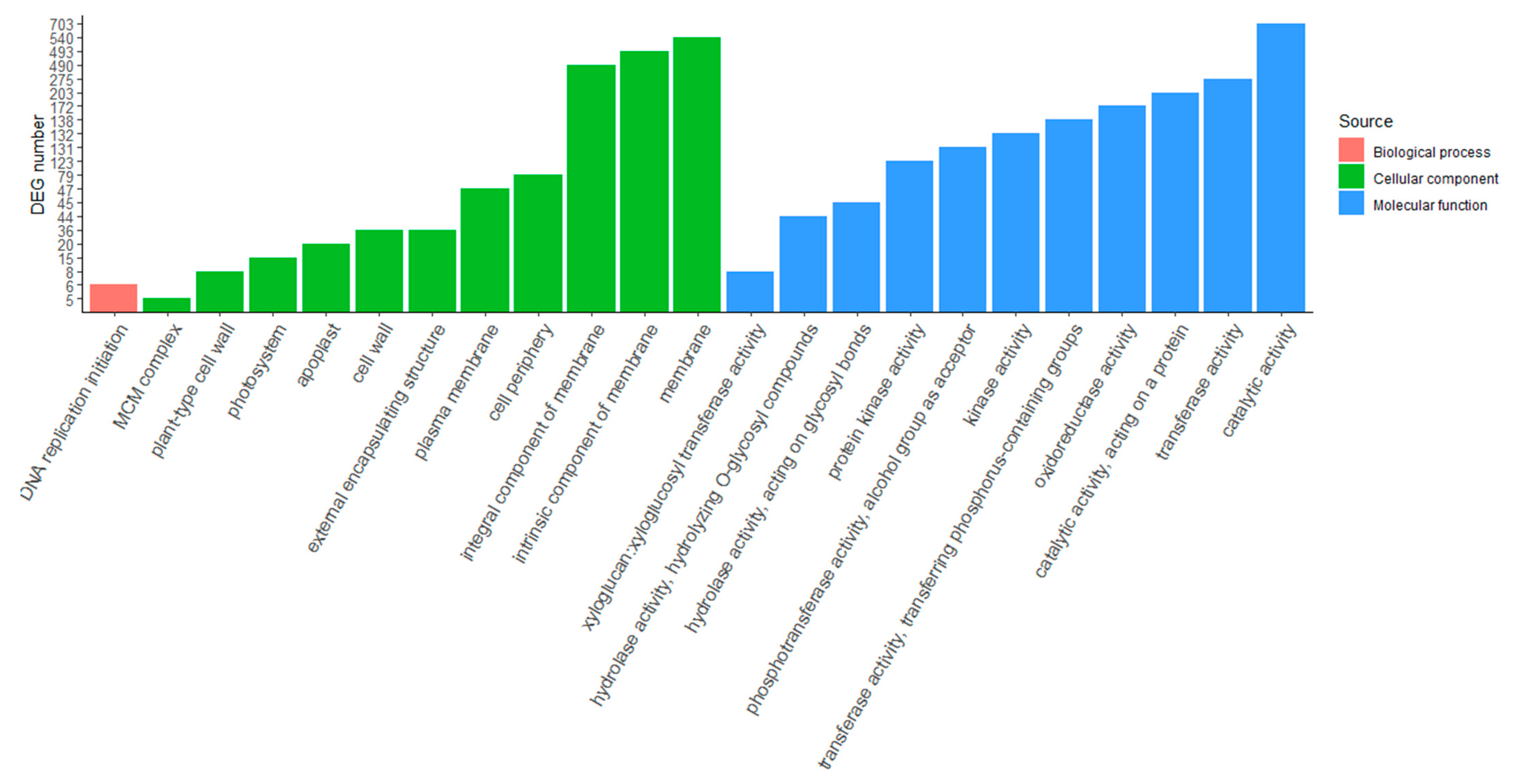
2. Results
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. RNA Extraction and cDNA Library Preparation for Sequencing
4.3. Transcriptome Analysis
4.4. GO Enrichment Analysis
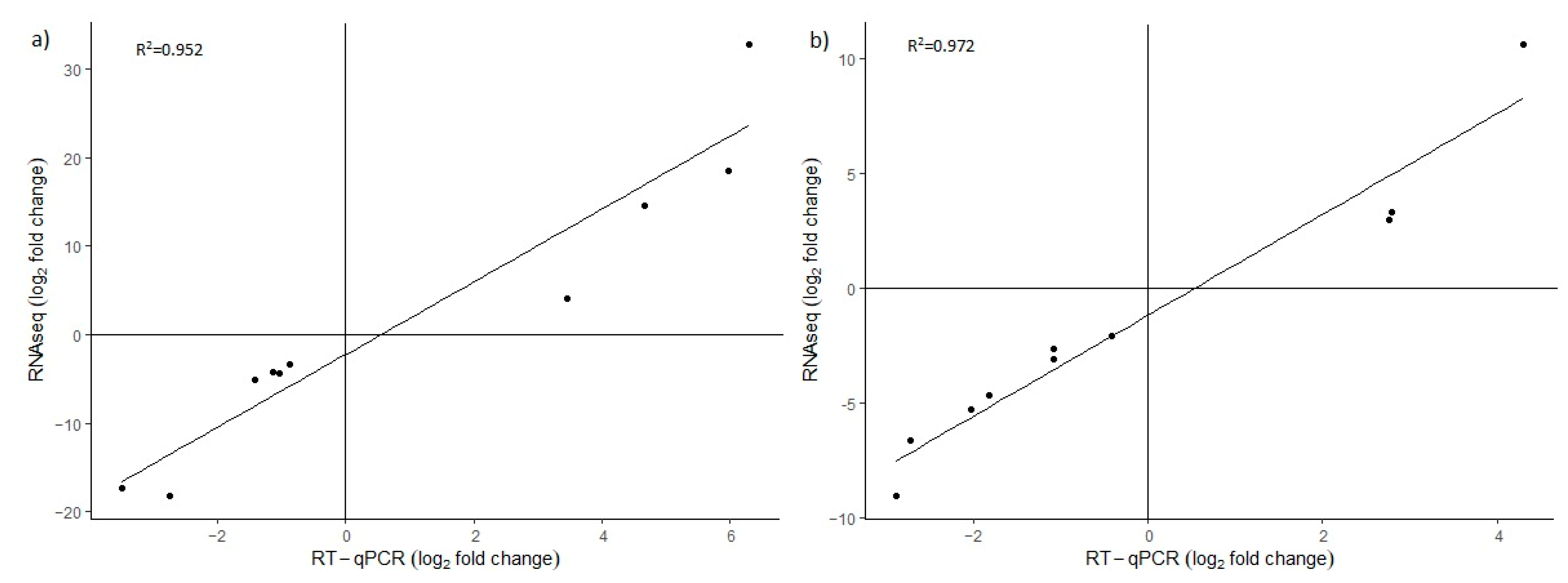
4.5. Validation of Differentially Expressed Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aksoy, E.; Demirel, U.; Öztürk, Z.N.; Çalişkan, S.; Çalişkan, M.E. Recent advances in potato genomics, transcriptomics, and transgenics under drought and heat stresses: A review. Turk. J. Bot. 2015, 39, 920–940. [Google Scholar] [CrossRef]
- Hill, D.; Nelson, D.; Hammond, J.; Bell, L. Morphophysiology of Potato (Solanum tuberosum) in Response to Drought Stress: Paving the Way Forward. Front. Plant Sci. 2021, 11, 597554. [Google Scholar] [CrossRef] [PubMed]
- Dahal, K.; Li, X.Q.; Tai, H.; Creelman, A.; Bizimungu, B. Improving potato stress tolerance and tuber yield under a climate change scenario—A current overview. Front. Plant Sci. 2019, 10, 563. [Google Scholar] [CrossRef] [PubMed]
- Boguszewska-Mańkowska, D.; Pieczyński, M.; Wyrzykowska, A.; Kalaji, H.M.; Sieczko, L.; Szweykowska-Kulińska, Z.; Zagdańska, B. Divergent strategies displayed by potato (Solanum tuberosum L.) cultivars to cope with soil drought. J. Agron. Crop Sci. 2018, 204, 13–30. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Pandey, S.; Muthamilarasan, M.; Chaudhry, V.; Dulani, P.; Prasad, M. Genomics Resources for Abiotic Stress Tolerance in Solanaceae Crops. In The Potato Genome; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 195–216. [Google Scholar]
- Barra, M.; Meneses, C.; Riquelme, S.; Pinto, M.; Lagüe, M.; Davidson, C.; Tai, H.H. Transcriptome Profiles of Contrasting Potato (Solanum tuberosum L.) Genotypes under Water Stress. Agronomy 2019, 9, 848. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Xiong, L. Genetic engineering and breeding of drought-resistant crops. Annu. Rev. Plant Biol. 2014, 65, 715–741. [Google Scholar] [CrossRef]
- George, T.S.; Taylor, M.A.; Dodd, I.C.; White, P.J. Climate Change and Consequences for Potato Production: A Review of Tolerance to Emerging Abiotic Stress. Potato Res. 2017, 60, 239–268. [Google Scholar] [CrossRef]
- Patil, V.U.; Sharma, N.N.; Chakrabarti, S.K. High-Throughput Sequencing of the Potato Genome. In The Potato Genome; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 95–107. [Google Scholar]
- Saidi, A.; Hajibarat, Z. Application of Next Generation Sequencing, GWAS, RNA seq, WGRS, for genetic improvement of potato (Solanum tuberosum L.) under drought stress. Biocatal. Agric. Biotechnol. 2020, 29, 101801. [Google Scholar] [CrossRef]
- Iquebal, M.A.; Sharma, P.; Jasrotia, R.S.; Jaiswal, S.; Kaur, A.; Saroha, M.; Angadi, U.B.; Sheoran, S.; Singh, R.; Singh, G.P.; et al. RNAseq analysis reveals drought-responsive molecular pathways with candidate genes and putative molecular markers in root tissue of wheat. Sci. Rep. 2019, 9, 13917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haider, M.S.; Zhang, C.; Kurjogi, M.M.; Pervaiz, T.; Zheng, T.; Zhang, C.; Lide, C.; Shangguan, L.; Fang, J. Insights into grapevine defense response against drought as revealed by biochemical, physiological and RNA-Seq analysis. Sci. Rep. 2017, 7, 13134. [Google Scholar] [CrossRef] [PubMed]
- Zenda, T.; Liu, S.; Wang, X.; Liu, G.; Jin, H.; Dong, A.; Yang, Y.; Duan, H. Key maize drought-responsive genes and pathways revealed by comparative transcriptome and physiological analyses of contrasting inbred lines. Int. J. Mol. Sci. 2019, 20, 1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, P.J.; Jung, H.; Jeong, D.H.; Ha, S.H.; Choi, Y.D.; Kim, J.K. Transcriptome profiling of drought responsive noncoding RNAs and their target genes in rice. BMC Genom. 2016, 17, 563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varshney, R.K.; Singh, V.K.; Kumar, A.; Powell, W.; Sorrells, M.E. Can genomics deliver climate-change ready crops? Curr. Opin. Plant Biol. 2018, 45, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Lavin Trueba, J.L.; Aransay, A. The High-Throughput Sequencing Technologies Triple-W Discussion: Why Use HTS, What Is the Optimal HTS Method to Use, and Which Data Analysis Workflow to Follow. In Field Guidelines for Genetic Experimental Designs in High-Throughput Sequencing; Springer: Berlin/Heidelberg, Germany, 2016; pp. 1–12. [Google Scholar]
- Bykova, I.V.; Shmakov, N.A.; Afonnikov, D.A.; Kochetov, A.V.; Khlestkina, E.K. Achievements and prospects of applying high-throughput sequencing techniques to potato genetics and breeding. Russ. J. Genet. Appl. Res. 2017, 7, 736–743. [Google Scholar] [CrossRef]
- Jain, M. Next-generation sequencing technologies for gene expression profiling in plants. Brief. Funct. Genom. 2012, 11, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Duan, Y.; Duan, S.; Armstrong, M.R.; Xu, J.; Zheng, J.; Hu, J.; Chen, X.; Hein, I.; Li, G.; Jin, L. Comparative transcriptome profiling reveals compatible and incompatible patterns of potato toward Phytophthora infestans. G3 Genes Genomes Genet. 2020, 10, 623–634. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekar, S.; Natarajan, P.; Mhatre, P.H.; Mahajan, M.; Nivitha, S.; Palanisamy, V.E.; Reddy, U.K.; Sundararaj, P. RNA-Seq of Cyst Nematode Infestation of Potato (Solanum tuberosum L.): A Comparative Transcriptome Analysis of Resistant and Susceptible Cultivars. Plants 2022, 11, 1008. [Google Scholar] [CrossRef]
- Li, Q.; Qin, Y.; Hu, X.; Li, G.; Ding, H.; Xiong, X.; Wang, W. Transcriptome analysis uncovers the gene expression profile of salt-stressed potato (Solanum tuberosum L.). Sci. Rep. 2020, 10, 5411. [Google Scholar] [CrossRef] [Green Version]
- Ahn, J.Y.; Kim, J.; Yang, J.Y.; Lee, H.J.; Kim, S.; Cho, K.-S.; Lee, S.H.; Kim, J.H.; Lee, T.H.; Hur, Y.; et al. Comparative Transcriptome Analysis between Two Potato Cultivars in Tuber Induction to Reveal Associated Genes with Anthocyanin Accumulation. Int. J. Mol. Sci. 2022, 23, 3681. [Google Scholar] [CrossRef]
- Gálvez, J.H.; Tai, H.H.; Lagüe, M.; Zebarth, B.J.; Strömvik, M.V. The nitrogen responsive transcriptome in potato (Solanum tuberosum L.) reveals significant gene regulatory motifs. Sci. Rep. 2016, 6, 26090. [Google Scholar] [CrossRef]
- Gong, L.; Zhang, H.; Gan, X.; Zhang, L.; Chen, Y.; Nie, F.; Shi, L.; Li, M.; Guo, Z.; Zhang, G.; et al. Transcriptome profiling of the potato (Solanum tuberosum L.) plant under drought stress and water-stimulus conditions. PLoS ONE 2015, 10, e0128041. [Google Scholar] [CrossRef] [PubMed]
- Moon, K.B.; Ahn, D.J.; Park, J.S.; Jung, W.Y.; Cho, H.S.; Kim, H.R.; Jeon, J.H.; Park, Y.I.; Kim, H.S. Transcriptome profiling and characterization of drought-tolerant potato plant (Solanum tuberosum L.). Mol. Cells 2018, 41, 979–992. [Google Scholar]
- Sprenger, H.; Erban, A.; Seddig, S.; Rudack, K.; Thalhammer, A.; Le, M.Q.; Walther, D.; Zuther, E.; Köhl, K.I.; Kopka, J.; et al. Metabolite and transcript markers for the prediction of potato drought tolerance. Plant Biotechnol. J. 2018, 16, 939–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, N.; Liu, B.; Ma, C.; Zhang, G.; Chang, J.; Si, H.; Wang, D. Transcriptome characterization and sequencing-based identification of drought-responsive genes in potato. Mol. Biol. Rep. 2014, 41, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Pieczynski, M.; Wyrzykowska, A.; Milanowska, K.; Boguszewska-Mankowska, D.; Zagdanska, B.; Karlowski, W.; Jarmolowski, A.; Szweykowska-Kulinska, Z. Genomewide identification of genes involved in the potato response to drought indicates functional evolutionary conservation with Arabidopsis plants. Plant Biotechnol. J. 2018, 16, 603–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krannich, C.T.; Maletzki, L.; Kurowsky, C.; Horn, R. Network candidate genes in breeding for drought tolerant crops. Int. J. Mol. Sci. 2015, 16, 16378–16400. [Google Scholar] [CrossRef]
- Chen, Y.; Li, C.; Yi, J.; Yang, Y.; Lei, C.; Gong, M. Transcriptome response to drought, rehydration and re-dehydration in potato. Int. J. Mol. Sci. 2020, 21, 159. [Google Scholar] [CrossRef] [Green Version]
- Chacón-Cerdas, R.; Barboza-Barquero, L.; Albertazzi, F.J.; Rivera-Méndez, W. Transcription factors controlling biotic stress response in potato plants. Physiol. Mol. Plant Pathol. 2020, 112, 101527. [Google Scholar] [CrossRef]
- Gonçalves, S. MYB transcription factors for enhanced drought tolerance in plants. In Water Stress and Crop Plants: A Sustainable Approach; John Wiley & Sons: Hoboken, NJ, USA, 2016; pp. 194–205. [Google Scholar]
- Ambrosone, A.; Batelli, G.; Bostan, H.; D’Agostino, N.; Chiusano, M.L.; Perrotta, G.; Leone, A.; Grillo, S.; Costa, A. Distinct gene networks drive differential response to abrupt or gradual water deficit in potato. Gene 2017, 597, 30–39. [Google Scholar] [CrossRef] [Green Version]
- Lata, C.; Yadav, A.; Prasad, M. Role of Plant Transcription Factors in Abiotic Stress Tolerance. In Abiotic Stress Response in Plants—Physiological, Biochemical and Genetic Perspectives; IntechOpen: London, UK, 2011. [Google Scholar]
- Manna, M.; Thakur, T.; Chirom, O.; Mandlik, R.; Deshmukh, R.; Salvi, P. Transcription factors as key molecular target to strengthen the drought stress tolerance in plants. Physiol Plant. 2021, 172, 847–868. [Google Scholar] [CrossRef]
- Shin, D.; Moon, S.J.; Han, S.; Kim, B.G.; Park, S.R.; Lee, S.K.; Yoon, H.J.; Lee, H.E.; Kwon, H.B.; Baek, D.; et al. Expression of StMYB1R-1, a novel potato single MYB-like domain transcription factor, increases drought tolerance. Plant Physiol. 2011, 155, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Joshi, R.; Wani, S.H.; Singh, B.; Bohra, A.; Dar, Z.A.; Lone, A.A.; Pareek, A.; Singla-Pareek, S.L. Transcription factors and plants response to drought stress: Current understanding and future directions. Front. Plant Sci. 2016, 7, 1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez-Morezuelas, A.; Barandalla, L.; Ritter, E.; Lacuesta, M.; Ruiz de Galarreta, J.I. Physiological response and yield components under greenhouse drought stress conditions in potato. J. Plant Physiol. 2022, 278, 153790. [Google Scholar] [CrossRef] [PubMed]
- Tang, R.; Gupta, S.K.; Niu, S.; Li, X.Q.; Yang, Q.; Chen, G.; Zhu, W.; Haroon, M. Transcriptome analysis of heat stress response genes in potato leaves. Mol. Biol. Rep. 2020, 47, 4311–4321. [Google Scholar] [CrossRef] [PubMed]
- van Muijen, D.; Anithakumari, A.M.; Maliepaard, C.; Visser, R.G.F.; van der Linden, C.G. Systems genetics reveals key genetic elements of drought induced gene regulation in diploid potato. Plant Cell Environ. 2016, 39, 1895–1908. [Google Scholar] [CrossRef]
- Muñoz-Mayor, A.; Pineda, B.; Garcia-Abellán, J.O.; Antón, T.; Garcia-Sogo, B.; Sanchez-Bel, P.; Flores, F.B.; Atarés, A.; Angosto, T.; Pintor-Toro, J.A.; et al. Overexpression of dehydrin tas14 gene improves the osmotic stress imposed by drought and salinity in tomato. J. Plant Physiol. 2012, 169, 459–468. [Google Scholar] [CrossRef]
- Aliche, E.B.; Gengler, T.; Hoendervangers, I.; Oortwijn, M.; Bachem, C.W.B.; Borm, T.; Visser, R.G.F.; van der Linden, C.G. Transcriptomic Responses of Potato to Drought Stress. Potato Res. 2022, 65, 289–305. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. g:Profiler: A web server for functional enrichment analysis and conversions of gene lists. Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicot, N.; Hausman, J.F.; Hoffmann, L.; Evers, D. Housekeeping gene selection for real-time RT-PCR normalization in potato during biotic and abiotic stress. J. Exp. Bot. 2005, 56, 2907–2914. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. Relative quantification. In Real-Time PCR; International University Line: La Jolla, CA, USA, 2001; pp. 63–82. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Drought | ||
|---|---|---|---|
| Agria | Raw reads | 57,004,343 | 54,509,300 |
| Clean reads | 56,473,479 | 54,042,913 | |
| Total mapped | 50,569,589 (89.54%) | 48,485,736 (89.71%) | |
| GC content (%) | 43.63 | 43.38 | |
| Q30 (%) | 96.08 | 96.18 | |
| Zorba | Raw reads | 56,278,132 | 53,964,119 |
| Clean reads | 55,798,228 | 53,513,496 | |
| Total mapped | 50,271,909 (90.09%) | 48,158,218 (89.99%) | |
| GC content (%) | 43.48 | 43.41 | |
| Q30 (%) | 96.12 | 96.14 |
| Gene Name | Description | log2fold change | p-Value |
|---|---|---|---|
| Downregulated genes | |||
| LOC102603621 | - | −126.51 | 4.56 × 10−7 |
| LOC102594756 | 36.4 kDa proline-rich protein-like | −39.22 | 5.85 × 10−6 |
| LOC102596805 | fasciclin-like arabinogalactan protein 2 | −37.45 | 3.65 × 10−8 |
| LOC102604005 | putative calcium-binding protein CML19 | −32.55 | 1.62 × 10−4 |
| LOC102602308 | hyoscyamine 6-dioxygenase | −31.44 | 3.14 × 10−4 |
| LOC102582168 | L-ascorbate oxidase-like | −30.82 | 5.63 × 10−7 |
| LOC102603929 | probable xyloglucan endotransglucosylase/hydrolase 1 | −30.46 | 1.54 × 10−4 |
| LOC102586959 | probable WRKY transcription factor 53 | −30.40 | 5.99 × 10−7 |
| LOC102583042 | 1-aminocyclopropane-1-carboxylate synthase-like | −28.26 | 1.62 × 10−7 |
| LOC102606325 | GDSL esterase/lipase At5g33370-like | −27.13 | 3.40 × 10−8 |
| Upregulated genes | |||
| LOC107057685 | abscisic acid and environmental stress-inducible protein TAS14-like | 3449.52 | 1.28 × 10−19 |
| LOC102583792 | abscisic stress-ripening protein 2 | 149.50 | 2.89 × 10−13 |
| LOC102606049 | fidgetin-like protein 1 | 140.15 | 6.90 × 10−10 |
| LOC102598218 | translocator protein homolog | 109.83 | 6.19 × 10−18 |
| LOC102590433 | - | 73.36 | 5.58 × 10−9 |
| LOC102606174 | bZIP transcription factor 53-like | 50.69 | 2.22 × 10−15 |
| LOC102598306 | SNF1-related protein kinase regulatory subunit gamma-like PV42a | 48.76 | 3.16 × 10−14 |
| LOC102592988 | - | 48.09 | 2.74 × 10−16 |
| LOC102584616 | - | 42.46 | 2.71 × 10−13 |
| LOC102591763 | branched-chain-amino-acid aminotransferase 2, chloroplastic-like | 39.31 | 1.50 × 10−8 |
| Gene Name | Description | log2fold change | p-Value |
|---|---|---|---|
| Downregulated genes | |||
| LOC102598924 | protein PMR5 | −19.17 | 8.26 × 10−18 |
| LOC102592481 | - | −16.05 | 7.36 × 10−13 |
| LOC102594756 | 36.4 kDa proline-rich protein-like | −15.42 | 1.85 × 10−3 |
| LOC102599076 | beta-hexosaminidase 3 | −11.63 | 9.22 × 10−10 |
| LOC102587252 | protein STRICTOSIDINE SYNTHASE-LIKE 11-like | −11.33 | 5.95 × 10−9 |
| LOC102605226 | glucan endo-1,3-beta-glucosidase, basic isoform 1-like | −11.33 | 5.66 × 10−8 |
| LOC102605560 | glucan endo-1,3-beta-glucosidase, basic isoform 1 | −10.91 | 5.85 × 10−7 |
| LOC102590679 | tropinone reductase homolog | −10.66 | 7.47 × 10−14 |
| LOC102594482 | delta(7)-sterol-C5(6)-desaturase-like | −10.37 | 2.30 × 10−6 |
| LOC102592844 | peroxidase 51 | −10.31 | 1.63 × 10−15 |
| Upregulated genes | |||
| LOC107057685 | abscisic acid and environmental stress-inducible protein TAS14-like | 692.09 | 1.47 × 10−12 |
| LOC102606049 | fidgetin-like protein 1 | 177.36 | 2.71 × 10−10 |
| LOC102590433 | - | 140.98 | 8.29 × 10−11 |
| LOC102580665 | O-acyltransferase WSD1-like | 69.42 | 4.71 × 10−22 |
| LOC102577501 | nonspecific lipid transfer protein a7 | 53.62 | 2.18 × 10−9 |
| LOC102596984 | nonspecific lipid-transfer protein 2-like | 50.35 | 1.77 × 10−8 |
| LOC102597309 | nonspecific lipid-transfer protein 2-like | 48.98 | 5.38 × 10−11 |
| LOC102598306 | SNF1-related protein kinase regulatory subunit gamma-like PV42a | 48.30 | 2.75 × 10−13 |
| LOC102582408 | probable protein phosphatase 2C 51 | 37.80 | 5.86 × 10−10 |
| LOC102587411 | MLO-like protein 6 | 36.71 | 7.14 × 10−6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvarez-Morezuelas, A.; Barandalla, L.; Ritter, E.; Ruiz de Galarreta, J.I. Transcriptome Analysis of Two Tetraploid Potato Varieties under Water-Stress Conditions. Int. J. Mol. Sci. 2022, 23, 13905. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232213905
Alvarez-Morezuelas A, Barandalla L, Ritter E, Ruiz de Galarreta JI. Transcriptome Analysis of Two Tetraploid Potato Varieties under Water-Stress Conditions. International Journal of Molecular Sciences. 2022; 23(22):13905. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232213905
Chicago/Turabian StyleAlvarez-Morezuelas, Alba, Leire Barandalla, Enrique Ritter, and Jose Ignacio Ruiz de Galarreta. 2022. "Transcriptome Analysis of Two Tetraploid Potato Varieties under Water-Stress Conditions" International Journal of Molecular Sciences 23, no. 22: 13905. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232213905