
Altered Cortical Palmitoylation Induces Widespread Molecular Disturbances in Parkinson’s Disease
 , , , and
, , , and 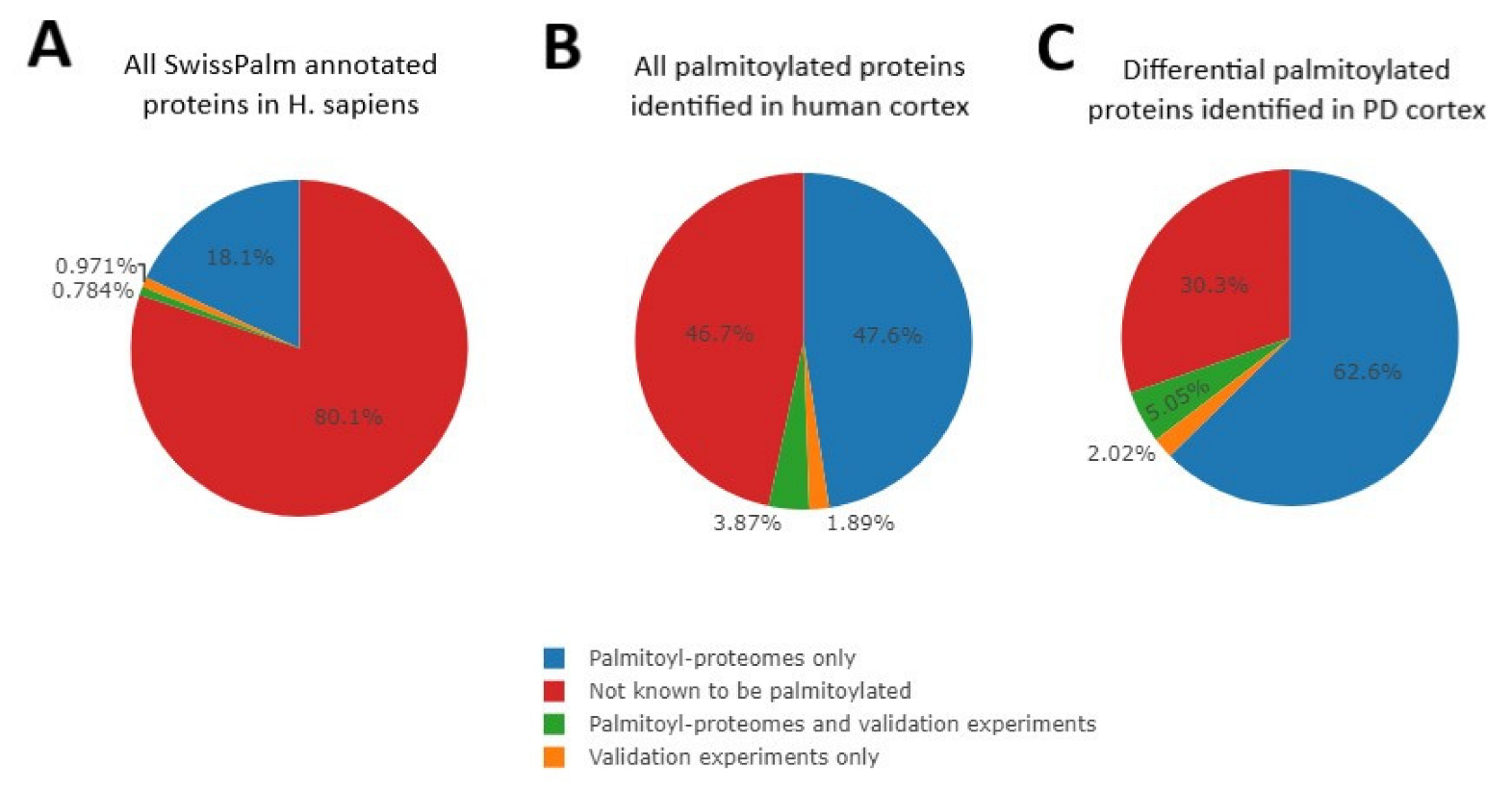{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
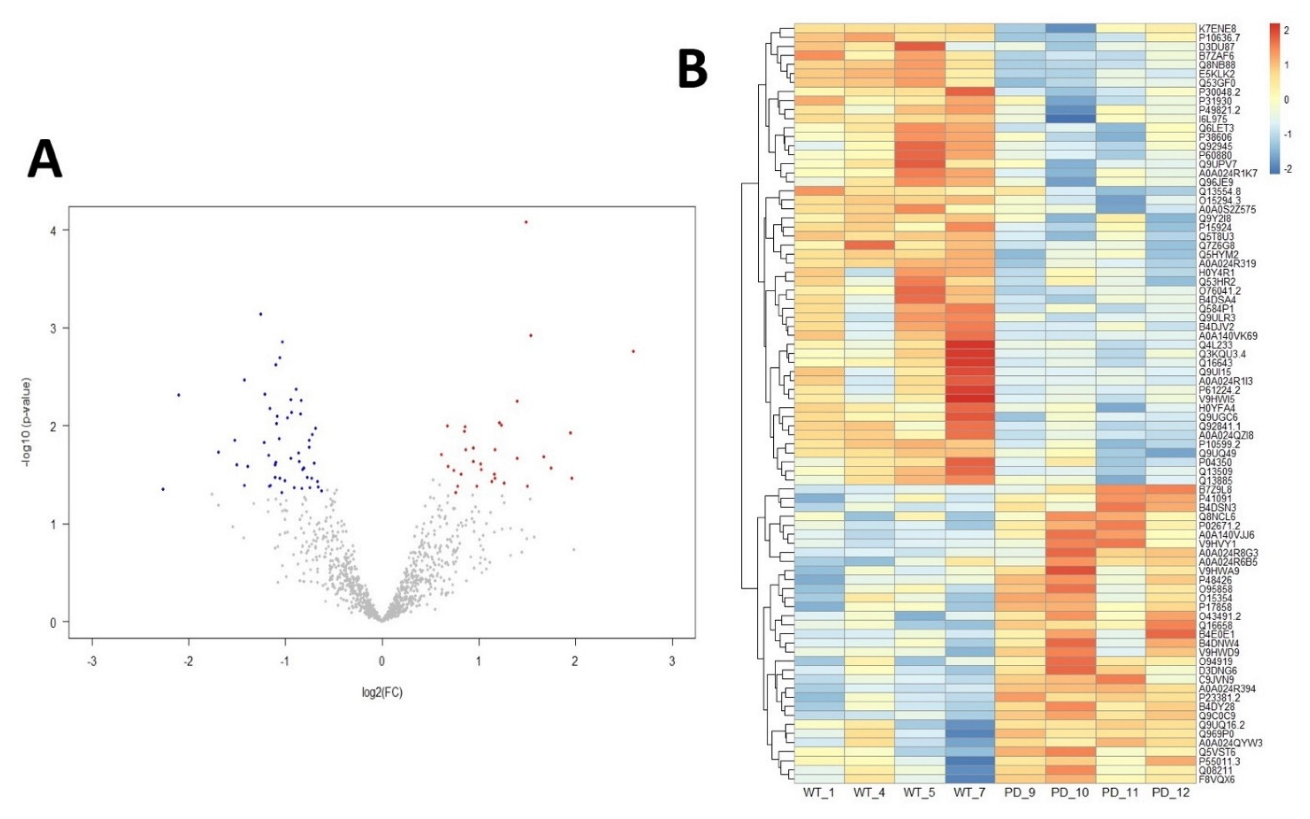
2.1. Altered Palmitoylation Levels of Cortical Proteins in PD
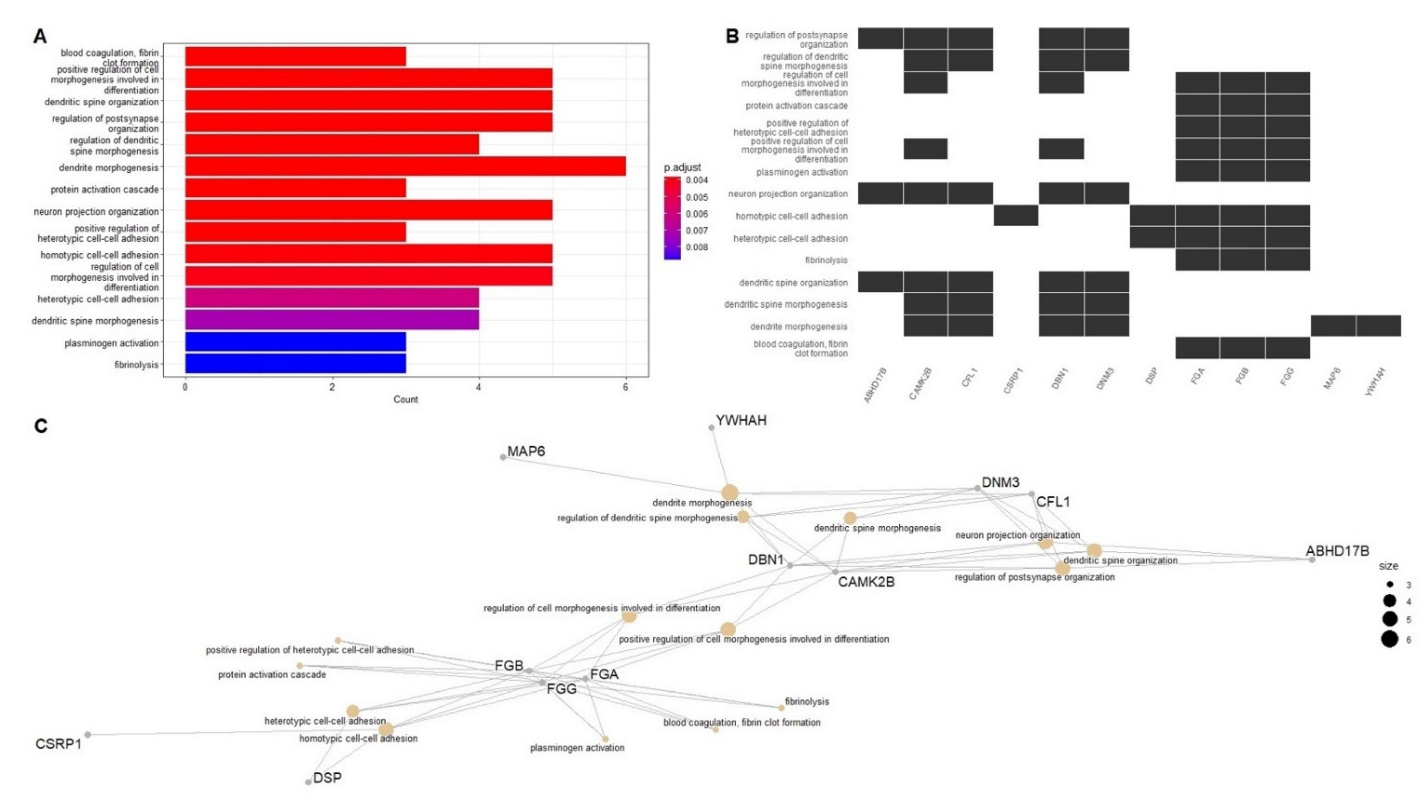
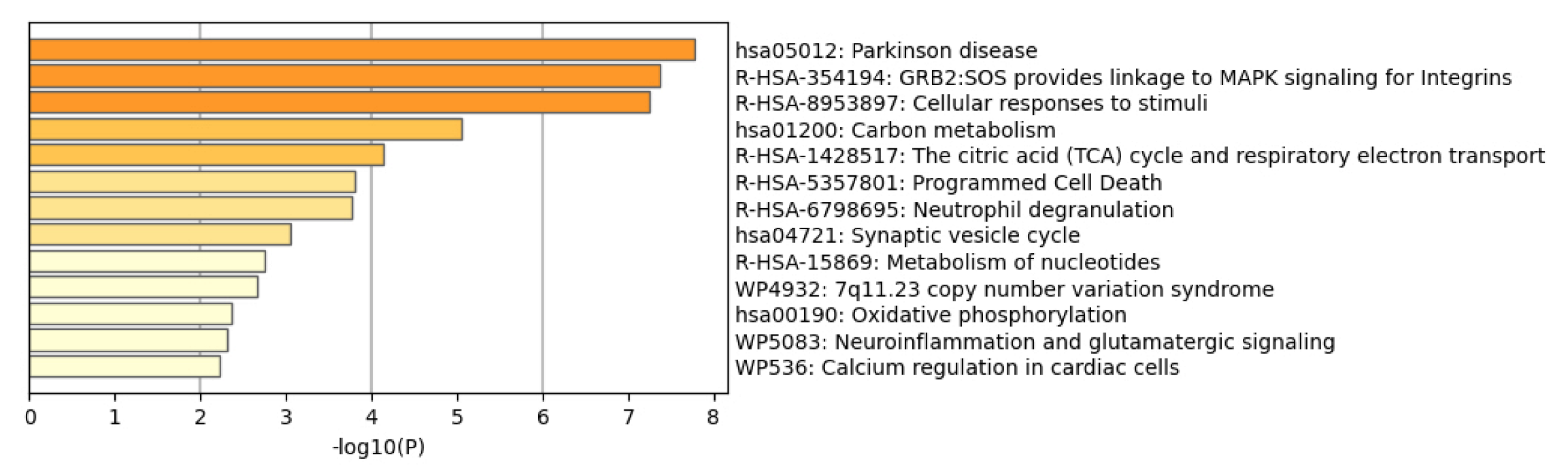
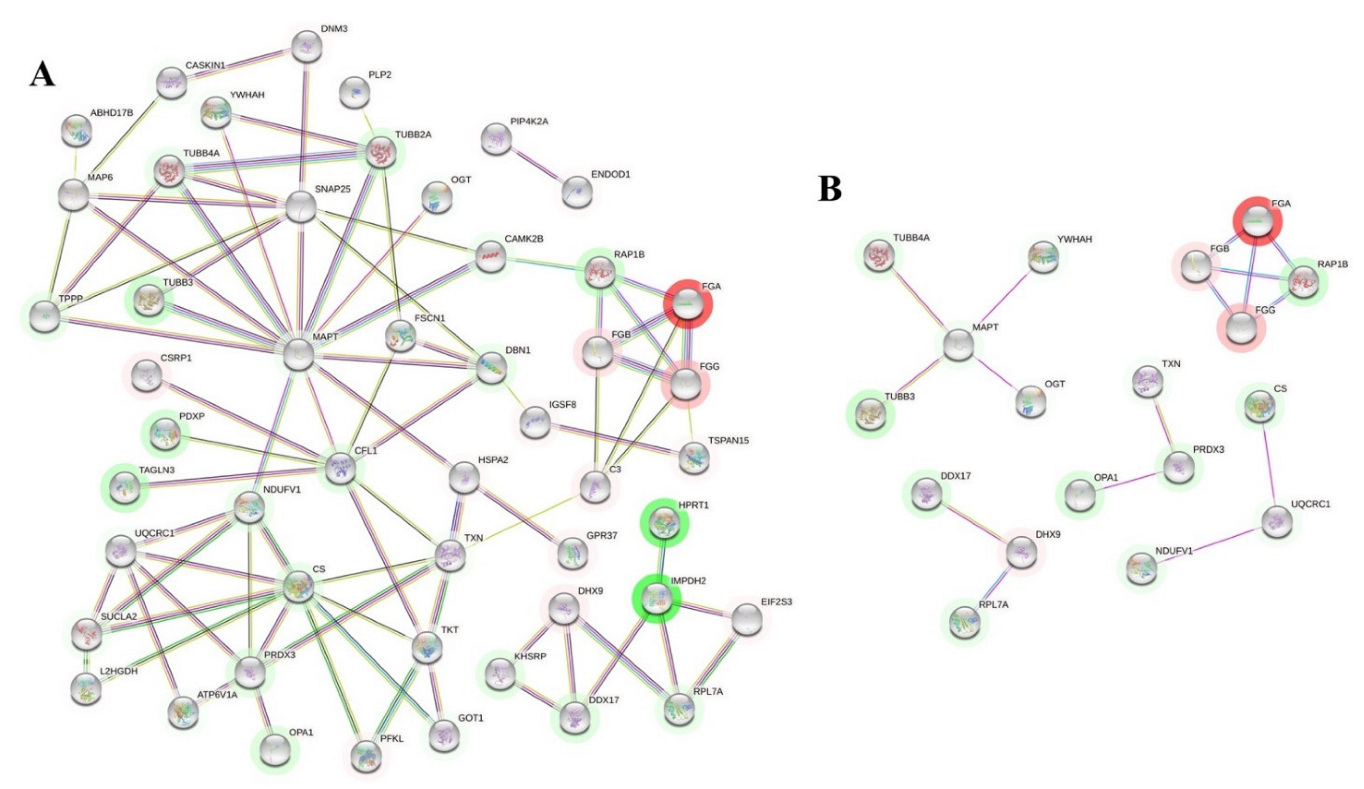
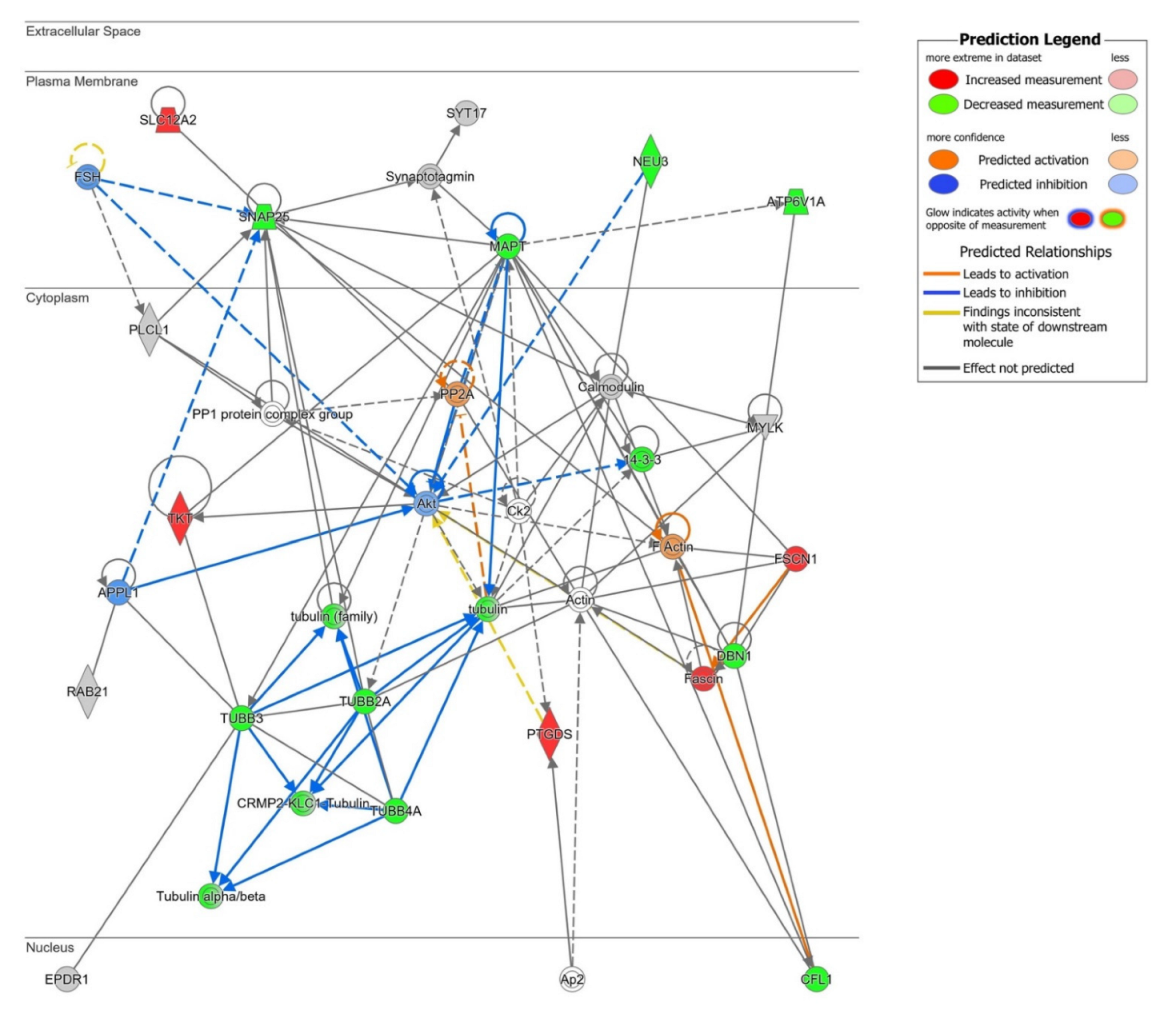
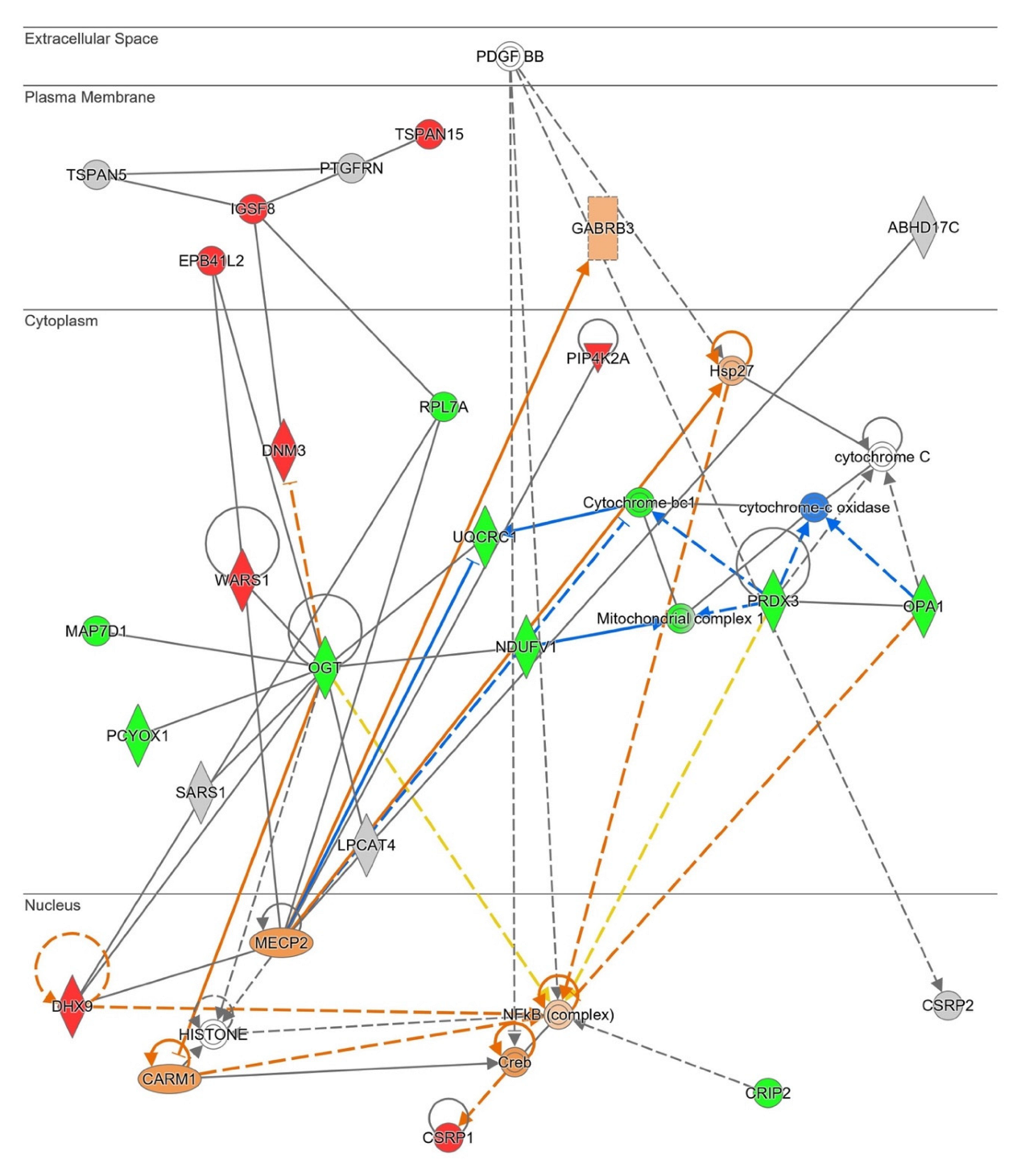
2.2. Network-Driven Palmitoyl-Proteomics Reveals an Imbalance in Cytoskeletal Architecture and Mitochondrial Homeostasis in PD
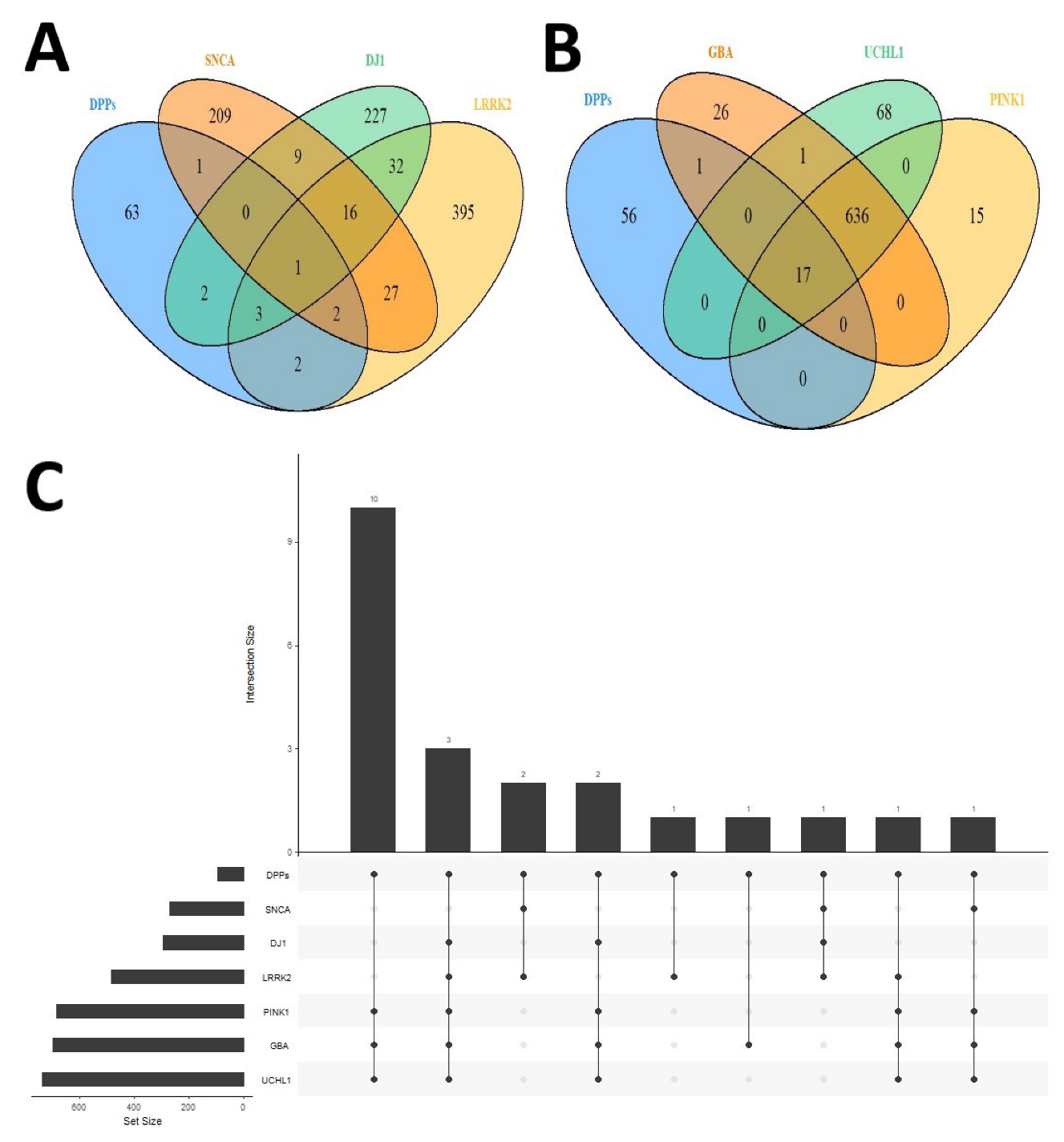
2.3. Differential Palmitoyl-Proteins Are Components of Interactomes Associated to PD Genes and Are Located in Multiple Cell Types
3. Materials and Methods
3.1. Subjects and Samples
3.2. Acyl-RAC
3.3. Label Free Proteomics
3.4. Data Preprocessing
3.5. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Balestrino, R.; Schapira, A.H.V. Parkinson Disease. Eur. J. Neurol. 2020, 27, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Foffani, G.; Obeso, J.A. A Cortical Pathogenic Theory of Parkinson’s Disease. Neuron 2018, 99, 1116–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schapira, A.H.V.; Chaudhuri, K.R.; Jenner, P. Non-Motor Features of Parkinson Disease. Nat. Rev. Neurosci. 2017, 18, 435–450. [Google Scholar] [CrossRef] [PubMed]
- Panicker, N.; Ge, P.; Dawson, V.L.; Dawson, T.M. The Cell Biology of Parkinson’s Disease. J. Cell Biol. 2021, 220, e202012095. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, M.J.; Okun, M.S. Diagnosis and Treatment of Parkinson Disease: A Review. JAMA 2020, 323, 548–560. [Google Scholar] [CrossRef]
- Hornemann, T. Palmitoylation and Depalmitoylation Defects. J. Inherit. Metab. Dis. 2015, 38, 179–186. [Google Scholar] [CrossRef]
- Zaręba-Kozioł, M.; Figiel, I.; Bartkowiak-Kaczmarek, A.; Włodarczyk, J. Insights Into Protein S-Palmitoylation in Synaptic Plasticity and Neurological Disorders: Potential and Limitations of Methods for Detection and Analysis. Front. Mol. Neurosci. 2018, 11, 175. [Google Scholar] [CrossRef] [Green Version]
- Fukata, Y.; Fukata, M. Protein Palmitoylation in Neuronal Development and Synaptic Plasticity. Nat. Rev. Neurosci. 2010, 11, 161–175. [Google Scholar] [CrossRef]
- Qu, M.; Zhou, X.; Wang, X.; Li, H. Lipid-Induced S-Palmitoylation as a Vital Regulator of Cell Signaling and Disease Development. Int. J. Biol. Sci. 2021, 17, 4223–4237. [Google Scholar] [CrossRef]
- Jin, J.; Zhi, X.; Wang, X.; Meng, D. Protein Palmitoylation and Its Pathophysiological Relevance. J. Cell. Physiol. 2021, 236, 3220–3233. [Google Scholar] [CrossRef]
- Cho, E.; Park, M. Palmitoylation in Alzheimer’s Disease and Other Neurodegenerative Diseases. Pharmacol. Res. 2016, 111, 133–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, F.B.; Butland, S.L.; Sanders, S.S.; Sutton, L.M.; Hayden, M.R. Putting Proteins in Their Place: Palmitoylation in Huntington Disease and Other Neuropsychiatric Diseases. Prog. Neurobiol. 2012, 97, 220–238. [Google Scholar] [CrossRef] [PubMed]
- Pinner, A.L.; Tucholski, J.; Haroutunian, V.; McCullumsmith, R.E.; Meador-Woodruff, J.H. Decreased Protein S-Palmitoylation in Dorsolateral Prefrontal Cortex in Schizophrenia. Schizophr. Res. 2016, 177, 78–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanders, S.S.; Hayden, M.R. Aberrant Palmitoylation in Huntington Disease. Biochem. Soc. Trans. 2015, 43, 205–210. [Google Scholar] [CrossRef]
- Virlogeux, A.; Scaramuzzino, C.; Lenoir, S.; Carpentier, R.; Louessard, M.; Genoux, A.; Lino, P.; Hinckelmann, M.V.; Perrier, A.L.; Humbert, S.; et al. Increasing Brain Palmitoylation Rescues Behavior and Neuropathology in Huntington Disease Mice. Sci. Adv. 2021, 7, eabb0799. [Google Scholar] [CrossRef]
- Henderson, M.X.; Wirak, G.S.; Zhang, Y.Q.; Dai, F.; Ginsberg, S.D.; Dolzhanskaya, N.; Staropoli, J.F.; Nijssen, P.C.G.; Lam, T.K.T.; Roth, A.F.; et al. Neuronal Ceroid Lipofuscinosis with DNAJC5/CSPα Mutation Has PPT1 Pathology and Exhibit Aberrant Protein Palmitoylation. Acta Neuropathol. 2016, 131, 621–637. [Google Scholar] [CrossRef] [Green Version]
- Kang, R.; Wan, J.; Arstikaitis, P.; Takahashi, H.; Huang, K.; Bailey, A.O.; Thompson, J.X.; Roth, A.F.; Drisdel, R.C.; Mastro, R.; et al. Neural Palmitoyl-Proteomics Reveals Dynamic Synaptic Palmitoylation. Nature 2008, 456, 904–909. [Google Scholar] [CrossRef] [Green Version]
- Prescott, G.R.; Gorleku, O.A.; Greaves, J.; Chamberlain, L.H. Palmitoylation of the Synaptic Vesicle Fusion Machinery. J. Neurochem. 2009, 110, 1135–1149. [Google Scholar] [CrossRef]
- Ji, B.; Skup, M. Roles of Palmitoylation in Structural Long-Term Synaptic Plasticity. Mol. Brain 2021, 14, 8. [Google Scholar] [CrossRef]
- Globa, A.K.; Bamji, S.X. Protein Palmitoylation in the Development and Plasticity of Neuronal Connections. Curr. Opin. Neurobiol. 2017, 45, 210–220. [Google Scholar] [CrossRef]
- Rastedt, D.E.; Vaughan, R.A.; Foster, J.D. Palmitoylation Mechanisms in Dopamine Transporter Regulation. J. Chem. Neuroanat. 2017, 83–84, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Kim, J.S.; Park, J.Y.; Suh, Y.H.; Jou, I.; Joe, E.H.; Park, S.M. DJ-1 Associates with Lipid Rafts by Palmitoylation and Regulates Lipid Rafts-Dependent Endocytosis in Astrocytes. Hum. Mol. Genet. 2013, 22, 4805–4817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schapansky, J.; Nardozzi, J.D.; Felizia, F.; LaVoie, M.J. Membrane Recruitment of Endogenous LRRK2 Precedes Its Potent Regulation of Autophagy. Hum. Mol. Genet. 2014, 23, 4201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, G.P.H.; Ramalingam, N.; Imberdis, T.; Wilkie, E.C.; Dettmer, U.; Selkoe, D.J. Upregulation of Cellular Palmitoylation Mitigates α-Synuclein Accumulation and Neurotoxicity. Mov. Disord. 2021, 36, 348–359. [Google Scholar] [CrossRef] [PubMed]
- Hosp, F.; Mann, M. A Primer on Concepts and Applications of Proteomics in Neuroscience. Neuron 2017, 96, 558–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanders, S.S.; Martin, D.D.O.; Butland, S.L.; Lavallée-Adam, M.; Calzolari, D.; Kay, C.; Yates, J.R.; Hayden, M.R. Curation of the Mammalian Palmitoylome Indicates a Pivotal Role for Palmitoylation in Diseases and Disorders of the Nervous System and Cancers. PLoS Comput. Biol. 2015, 11, e1004405. [Google Scholar] [CrossRef] [Green Version]
- Aghamaleky Sarvestany, A.; Hunter, G.; Tavendale, A.; Lamont, D.J.; Llavero Hurtado, M.; Graham, L.C.; Wishart, T.M.; Gillingwater, T.H. Label-Free Quantitative Proteomic Profiling Identifies Disruption of Ubiquitin Homeostasis as a Key Driver of Schwann Cell Defects in Spinal Muscular Atrophy. J. Proteome Res. 2014, 13, 4546–4557. [Google Scholar] [CrossRef]
- Cox, J.; Hein, M.Y.; Luber, C.A.; Paron, I.; Nagaraj, N.; Mann, M. Accurate Proteome-Wide Label-Free Quantification by Delayed Normalization and Maximal Peptide Ratio Extraction, Termed MaxLFQ. Mol. Cell. Proteom. 2014, 13, 2513–2526. [Google Scholar] [CrossRef] [Green Version]
- Blanc, M.; David, F.P.A.; Abrami, L.; Migliozzi, D.; Armand, F.; Jérôme, B.; van der Goot, F.G. SwissPalm: Protein Palmitoylation Database. Available online: https://swisspalm.org/reference (accessed on 9 October 2022).
- Carbon, S.; Douglass, E.; Good, B.M.; Unni, D.R.; Harris, N.L.; Mungall, C.J.; Basu, S.; Chisholm, R.L.; Dodson, R.J.; Hartline, E.; et al. The Gene Ontology Resource: Enriching a GOld Mine. Nucleic Acids Res. 2021, 49, D325–D334. [Google Scholar] [CrossRef]
- Lin, D.T.S.; Conibear, E. ABHD17 Proteins Are Novel Protein Depalmitoylases That Regulate N-Ras Palmitate Turnover and Subcellular Localization. Elife 2015, 4, e11306. [Google Scholar] [CrossRef]
- Davalos, D.; Akassoglou, K. Fibrinogen as a Key Regulator of Inflammation in Disease. Semin. Immunopathol. 2012, 34, 43–62. [Google Scholar] [CrossRef] [PubMed]
- Kattula, S.; Byrnes, J.R.; Wolberg, A.S. Fibrinogen and Fibrin in Hemostasis and Thrombosis. Arterioscler. Thromb. Vasc. Biol. 2017, 37, e13–e21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halper, J.; Kjaer, M. Basic Components of Connective Tissues and Extracellular Matrix: Elastin, Fibrillin, Fibulins, Fibrinogen, Fibronectin, Laminin, Tenascins and Thrombospondins. Adv. Exp. Med. Biol. 2014, 802, 31–47. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Fu, Y.; Cui, Y.; He, Y.; Zeng, X.; Ploplis, V.A.; Castellino, F.J.; Luo, Y. Fibrinogen Has Chaperone-like Activity. Biochem. Biophys. Res. Commun. 2009, 378, 662–667. [Google Scholar] [CrossRef] [Green Version]
- Petersen, M.A.; Ryu, J.K.; Akassoglou, K. Fibrinogen in Neurological Diseases: Mechanisms, Imaging and Therapeutics. Nat. Rev. Neurosci. 2018, 19, 283–301. [Google Scholar] [CrossRef]
- Sulimai, N.; Lominadze, D. Fibrinogen and Neuroinflammation During Traumatic Brain Injury. Mol. Neurobiol. 2020, 57, 4692–4703. [Google Scholar] [CrossRef]
- Merlini, M.; Rafalski, V.A.; Rios Coronado, P.E.; Gill, T.M.; Ellisman, M.; Muthukumar, G.; Subramanian, K.S.; Ryu, J.K.; Syme, C.A.; Davalos, D.; et al. Fibrinogen Induces Microglia-Mediated Spine Elimination and Cognitive Impairment in an Alzheimer’s Disease Model. Neuron 2019, 101, 1099–1108.e6. [Google Scholar] [CrossRef] [Green Version]
- Davalos, D.; Mahajan, K.R.; Trapp, B.D. Brain Fibrinogen Deposition Plays a Key Role in MS Pathophysiology—Yes. Mult. Scler. 2019, 25, 1434–1435. [Google Scholar] [CrossRef] [Green Version]
- Pajares, M.; Rojo, A.I.; Manda, G.; Boscá, L.; Cuadrado, A. Inflammation in Parkinson’s Disease: Mechanisms and Therapeutic Implications. Cells 2020, 9, 1687. [Google Scholar] [CrossRef]
- Sweeney, M.D.; Sagare, A.P.; Zlokovic, B.V. Blood-Brain Barrier Breakdown in Alzheimer Disease and Other Neurodegenerative Disorders. Nat. Rev. Neurol. 2018, 14, 133–150. [Google Scholar] [CrossRef]
- Sato, Y.; Kaji, M.; Metoki, N.; Yoshida, H.; Satoh, K. Coagulation-Fibrinolysis Abnormalities in Patients Receiving Antiparkinsonian Agents. J. Neurol. Sci. 2003, 212, 55–58. [Google Scholar] [CrossRef]
- Chen, H.; O’Reilly, E.J.; Schwarzschild, M.A.; Ascherio, A. Peripheral Inflammatory Biomarkers and Risk of Parkinson’s Disease. Am. J. Epidemiol. 2008, 167, 90–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, K.T.; Grove, J.S.; Grandinetti, A.; Curb, J.D.; Yee, M.; Blanchette, P.; Ross, G.W.; Rodriguez, B.I. Association of Fibrinogen with Parkinson Disease in Elderly Japanese-American Men: A Prospective Study. Neuroepidemiology 2010, 34, 50–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, I.U.; Kim, Y.D.; Cho, H.J.; Chung, S.W. Is Neuroinflammation Involved in the Development of Dementia in Patients with Parkinson’s Disease? Intern. Med. 2013, 52, 1787–1792. [Google Scholar] [CrossRef] [Green Version]
- Maarouf, C.L.; Beach, T.G.; Adler, C.H.; Shill, H.A.; Sabbagh, M.N.; Wu, T.; Walker, D.G.; Kokjohn, T.A.; Roher, A.E. Cerebrospinal Fluid Biomarkers of Neuropathologically Diagnosed Parkinson’s Disease Subjects. Neurol. Res. 2012, 34, 669–676. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape Provides a Biologist-Oriented Resource for the Analysis of Systems-Level Datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- González-Rodríguez, P.; Zampese, E.; Stout, K.A.; Guzman, J.N.; Ilijic, E.; Yang, B.; Tkatch, T.; Stavarache, M.A.; Wokosin, D.L.; Gao, L.; et al. Disruption of Mitochondrial Complex I Induces Progressive Parkinsonism. Nat. 2021 5997886 2021, 599, 650–656. [Google Scholar] [CrossRef]
- Haas, R.H.; Nasirian, F.; Nakano, K.; Ward, D.; Pay, M.; Hill, R.; Shults, C.W. Low Platelet Mitochondrial Complex I and Complex II/III Activity in Early Untreated Parkinson’s Disease. Ann. Neurol. 1995, 37, 714–722. [Google Scholar] [CrossRef]
- Yoshino, H.; Nakagawa-Hattori, Y.; Kondo, T.; Mizuno, Y. Mitochondrial Complex I and II Activities of Lymphocytes and Platelets in Parkinson’s Disease. J. Neural Transm. Park. Dis. Dement. Sect. 1992, 4, 27–34. [Google Scholar] [CrossRef]
- Liu, Z.; Ye, Q.; Wang, F.; Guo, Y.; Cui, R.; Wang, J.; Wang, D. Protective Effect of Thioredoxin Reductase 1 in Parkinson’s Disease. Neurosci. Lett. 2021, 741, 135457. [Google Scholar] [CrossRef]
- Xu, Z.; Zhong, L. New Insights into the Posttranslational Regulation of Human Cytosolic Thioredoxin by S-Palmitoylation. Biochem. Biophys. Res. Commun. 2015, 460, 949–956. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Liang, W.; Xu, Z.; Ye, F.; Li, X.; Zhong, L. Mechanistic Insights into the Inhibitory Effects of Palmitoylation on Cytosolic Thioredoxin Reductase and Thioredoxin. Biochimie 2015, 110, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Zhao, J.; Feng, J. Parkin Binds to Alpha/Beta Tubulin and Increases Their Ubiquitination and Degradation. J. Neurosci. 2003, 23, 3316–3324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zambito, A.M.; Wolff, J. Palmitoylation of Tubulin. Biochem. Biophys. Res. Commun. 1997, 239, 650–654. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING Database in 2021: Customizable Protein-Protein Networks, and Functional Characterization of User-Uploaded Gene/Measurement Sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Binarová, P.; Tuszynski, J. Tubulin: Structure, Functions and Roles in Disease. Cells 2019, 8, 1294. [Google Scholar] [CrossRef] [Green Version]
- Saha, P.; Sen, N. Tauopathy: A Common Mechanism for Neurodegeneration and Brain Aging. Mech. Ageing Dev. 2019, 178, 72–79. [Google Scholar] [CrossRef]
- Alquezar, C.; Arya, S.; Kao, A.W. Tau Post-Translational Modifications: Dynamic Transformers of Tau Function, Degradation, and Aggregation. Front. Neurol. 2021, 11, 1826. [Google Scholar] [CrossRef]
- Strang, K.H.; Golde, T.E.; Giasson, B.I. MAPT Mutations, Tauopathy, and Mechanisms of Neurodegeneration. Lab. Investig. 2019, 99, 912–928. [Google Scholar] [CrossRef]
- Leveille, E.; Ross, O.A.; Gan-Or, Z. Tau and MAPT Genetics in Tauopathies and Synucleinopathies. Park. Relat. Disord. 2021, 90, 142–154. [Google Scholar] [CrossRef]
- Zhang, X.; Gao, F.; Wang, D.; Li, C.; Fu, Y.; He, W.; Zhang, J. Tau Pathology in Parkinson’s Disease. Front. Neurol. 2018, 9, 809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; James, S.; Lei, P. Interactions Between α-Synuclein and Tau Protein: Implications to Neurodegenerative Disorders. J. Mol. Neurosci. 2016, 60, 298–304. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, H.; Lasda, E.; Bayer, K.U. CaMKIIβ Association with the Actin Cytoskeleton Is Regulated by Alternative Splicing. Mol. Biol. Cell 2006, 17, 4656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fink, C.C.; Bayer, K.U.; Myers, J.W.; Ferrell, J.E.; Schulman, H.; Meyer, T. Selective Regulation of Neurite Extension and Synapse Formation by the β but Not the α Isoform of CaMKII. Neuron 2003, 39, 283–297. [Google Scholar] [CrossRef] [Green Version]
- Pellegrini, L.; Wetzel, A.; Grannó, S.; Heaton, G.; Harvey, K. Back to the Tubule: Microtubule Dynamics in Parkinson’s Disease. Cell. Mol. Life Sci. 2017, 74, 409. [Google Scholar] [CrossRef] [Green Version]
- Calogero, A.M.; Mazzetti, S.; Pezzoli, G.; Cappelletti, G. Neuronal Microtubules and Proteins Linked to Parkinson’s Disease: A Relevant Interaction? Biol. Chem. 2019, 400, 1099–1112. [Google Scholar] [CrossRef]
- Oliveira da Silva, M.I.; Liz, M.A. Linking Alpha-Synuclein to the Actin Cytoskeleton: Consequences to Neuronal Function. Front. Cell Dev. Biol. 2020, 8, 787. [Google Scholar] [CrossRef]
- Bertoni, A.; Tadokoro, S.; Eto, K.; Pampori, N.; Parise, L.V.; White, G.C.; Shattil, S.J. Relationships between Rap1b, Affinity Modulation of Integrin Alpha IIbbeta 3, and the Actin Cytoskeleton. J. Biol. Chem. 2002, 277, 25715–25721. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Xiang, B.; Ye, S.; Chrzanowska-Wodnicka, M.; Morris, A.J.; Gartner, T.K.; Whiteheart, S.W.; White, G.C.; Smyth, S.S.; Li, Z. Distinct Roles for Rap1b Protein in Platelet Secretion and Integrin AIIbβ3 Outside-in Signaling. J. Biol. Chem. 2011, 286, 39466–39477. [Google Scholar] [CrossRef] [Green Version]
- Rai, S.N.; Dilnashin, H.; Birla, H.; Singh, S.S.; Zahra, W.; Rathore, A.S.; Singh, B.K.; Singh, S.P. The Role of PI3K/Akt and ERK in Neurodegenerative Disorders. Neurotox. Res. 2019, 35, 775–795. [Google Scholar] [CrossRef]
- Su, J.; Zhang, J.; Bao, R.; Xia, C.; Zhang, Y.; Zhu, Z.; Lv, Q.; Qi, Y.; Xue, J. Mitochondrial Dysfunction and Apoptosis Are Attenuated through Activation of AMPK/GSK-3β/PP2A Pathway in Parkinson’s Disease. Eur. J. Pharmacol. 2021, 907, 174202. [Google Scholar] [CrossRef] [PubMed]
- Heo, K.; Basu, H.; Gutnick, A.; Wei, W.; Shlevkov, E.; Schwarz, T.L. Serine/Threonine Protein Phosphatase 2A Regulates the Transport of Axonal Mitochondria. Front. Cell. Neurosci. 2022, 16, 88. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.Y.; Han, Z.D.; Li, W.; Yue, F.; Ye, J.; Li, B.; Cai, Z.; Lu, J.M.; Dong, W.; Jiang, X.; et al. Mitochondrion-Associated Protein Peroxiredoxin 3 Promotes Benign Prostatic Hyperplasia through Autophagy Suppression and Pyroptosis Activation. Oncotarget 2017, 8, 80295–80302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trist, B.G.; Hare, D.J.; Double, K.L. Oxidative Stress in the Aging Substantia Nigra and the Etiology of Parkinson’s Disease. Aging Cell 2019, 18, e13031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plain, F.; Howie, J.; Kennedy, J.; Brown, E.; Shattock, M.J.; Fraser, N.J.; Fuller, W. Control of Protein Palmitoylation by Regulating Substrate Recruitment to a ZDHHC-Protein Acyltransferase. Commun. Biol. 2020 31 2020, 3, 411. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Li, H.; He, Y.; Hao, J. The Emerging Link between O-GlcNAcylation and Neurological Disorders. Cell. Mol. Life Sci. 2017, 74, 3667–3686. [Google Scholar] [CrossRef]
- Yuzwa, S.A.; Shan, X.; MacAuley, M.S.; Clark, T.; Skorobogatko, Y.; Vosseller, K.; Vocadlo, D.J. Increasing O-GlcNAc Slows Neurodegeneration and Stabilizes Tau against Aggregation. Nat. Chem. Biol. 2012, 8, 393–399. [Google Scholar] [CrossRef]
- Liu, F.; Iqbal, K.; Grundke-Iqbal, I.; Hart, G.W.; Gong, C.X. O-GlcNAcylation Regulates Phosphorylation of Tau: A Mechanism Involved in Alzheimer’s Disease. Proc. Natl. Acad. Sci. USA 2004, 101, 10804–10809. [Google Scholar] [CrossRef] [Green Version]
- Marotta, N.P.; Lin, Y.H.; Lewis, Y.E.; Ambroso, M.R.; Zaro, B.W.; Roth, M.T.; Arnold, D.B.; Langen, R.; Pratt, M.R. O-GlcNAc Modification Blocks the Aggregation and Toxicity of the Protein α-Synuclein Associated with Parkinson’s Disease. Nat. Chem. 2015, 7, 913–920. [Google Scholar] [CrossRef] [Green Version]
- Lewis, Y.E.; Galesic, A.; Levine, P.M.; De Leon, C.A.; Lamiri, N.; Brennan, C.K.; Pratt, M.R. O-GlcNAcylation of α-Synuclein at Serine 87 Reduces Aggregation without Affecting Membrane Binding. ACS Publ. 2017, 12, 1020–1027. [Google Scholar] [CrossRef]
- Cao, Y.; Qiu, T.; Kathayat, R.S.; Azizi, S.A.; Thorne, A.K.; Ahn, D.; Fukata, Y.; Fukata, M.; Rice, P.A.; Dickinson, B.C. ABHD10 Is an S-Depalmitoylase Affecting Redox Homeostasis through Peroxiredoxin-5. Nat. Chem. Biol. 2019, 15, 1232. [Google Scholar] [CrossRef] [PubMed]
- Lanzillotta, A.; Porrini, V.; Bellucci, A.; Benarese, M.; Branca, C.; Parrella, E.; Spano, P.F.; Pizzi, M. NF-ΚB in Innate Neuroprotection and Age-Related Neurodegenerative Diseases. Front. Neurol. 2015, 6, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellucci, A.; Bubacco, L.; Longhena, F.; Parrella, E.; Faustini, G.; Porrini, V.; Bono, F.; Missale, C.; Pizzi, M. Nuclear Factor-ΚB Dysregulation and α-Synuclein Pathology: Critical Interplay in the Pathogenesis of Parkinson’s Disease. Front. Aging Neurosci. 2020, 12, 68. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.S.; Rai, S.N.; Birla, H.; Zahra, W.; Rathore, A.S.; Singh, S.P. NF-ΚB-Mediated Neuroinflammation in Parkinson’s Disease and Potential Therapeutic Effect of Polyphenols. Neurotox. Res. 2020, 37, 491–507. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Roy, A.; Liu, X.; Kordower, J.H.; Mufson, E.J.; Hartley, D.M.; Ghosh, S.; Mosley, R.L.; Gendelman, H.E.; Pahan, K. Selective Inhibition of NF-ΚB Activation Prevents Dopaminergic Neuronal Loss in a Mouse Model of Parkinson’s Disease. Proc. Natl. Acad. Sci. USA 2007, 104, 18754–18759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ordonez, D.G.; Lee, M.K.; Feany, M.B. α-Synuclein Induces Mitochondrial Dysfunction through Spectrin and the Actin Cytoskeleton. Neuron 2018, 97, 108. [Google Scholar] [CrossRef]
- Singh, A.; Zhi, L.; Zhang, H. LRRK2 and Mitochondria: Recent Advances and Current Views. Brain Res. 2019, 1702, 96–104. [Google Scholar] [CrossRef]
- Angeles, D.C.; Gan, B.H.; Onstead, L.; Zhao, Y.; Lim, K.L.; Dachsel, J.; Melrose, H.; Farrer, M.; Wszolek, Z.K.; Dickson, D.W.; et al. Mutations in LRRK2 Increase Phosphorylation of Peroxiredoxin 3 Exacerbating Oxidative Stress-Induced Neuronal Death. Hum. Mutat. 2011, 32, 1390–1397. [Google Scholar] [CrossRef]
- Angeles, D.C.; Ho, P.; Chua, L.L.; Wang, C.; Yap, Y.W.; Ng, C.; Zhou, Z.D.; Lim, K.L.; Wszolek, Z.K.; Wang, H.Y.; et al. Thiol Peroxidases Ameliorate LRRK2 Mutant-Induced Mitochondrial and Dopaminergic Neuronal Degeneration in Drosophila. Hum. Mol. Genet. 2014, 23, 3157–3165. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, L.; Rust, M.B.; Culmsee, C. Actin(g) on Mitochondria—A Role for Cofilin1 in Neuronal Cell Death Pathways. Biol. Chem. 2019, 400, 1089–1097. [Google Scholar] [CrossRef]
- Sekino, Y.; Kojima, N.; Shirao, T. Role of Actin Cytoskeleton in Dendritic Spine Morphogenesis. Neurochem. Int. 2007, 51, 92–104. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kang, H.; Lee, Y.; Park, C.H.; Jo, A.; Khang, R.; Shin, J.H. Identification of Transketolase as a Target of PARIS in Substantia Nigra. Biochem. Biophys. Res. Commun. 2017, 493, 1050–1056. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.; Schmitt, S.; Bergner, C.G.; Tyanova, S.; Kannaiyan, N.; Manrique-Hoyos, N.; Kongi, K.; Cantuti, L.; Hanisch, U.K.; Philips, M.A.; et al. Cell Type- and Brain Region-Resolved Mouse Brain Proteome. Nat. Neurosci. 2015, 18, 1819–1831. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, K.; Sloan, S.A.; Bennett, M.L.; Scholze, A.R.; O’Keeffe, S.; Phatnani, H.P.; Guarnieri, P.; Caneda, C.; Ruderisch, N.; et al. An RNA-Sequencing Transcriptome and Splicing Database of Glia, Neurons, and Vascular Cells of the Cerebral Cortex. J. Neurosci. 2014, 34, 11929–11947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tewari, R.; West, S.J.; Shayahati, B.; Akimzhanov, A.M. Detection of Protein S-Acylation Using Acyl-Resin Assisted Capture. J. Vis. Exp. 2020, 2020, e61016. [Google Scholar] [CrossRef] [PubMed]
- Gök, C.; Main, A.; Gao, X.; Kerekes, Z.; Plain, F.; Kuo, C.W.; Robertson, A.D.; Fraser, N.J.; Fuller, W. Insights into the Molecular Basis of the Palmitoylation and Depalmitoylation of NCX1. Cell Calcium 2021, 97, 102408. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Cox, J. The MaxQuant Computational Platform for Mass Spectrometry-Based Shotgun Proteomics. Nat. Protoc. 2016, 11, 2301–2319. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2020. Available online: http://www.r-project.org/index.html (accessed on 9 October 2022).
- Hornik, K. The Comprehensive R Archive Network. Wiley Interdiscip. Rev. Comput. Stat. 2012, 4, 394–398. [Google Scholar] [CrossRef]
- Gentleman, R.C.; Carey, V.J.; Bates, D.M.; Bolstad, B.; Dettling, M.; Dudoit, S.; Ellis, B.; Gautier, L.; Ge, Y.; Gentry, J.; et al. Bioconductor: Open Software Development for Computational Biology and Bioinformatics. Genome Biol. 2004 510 2004, 5, R80. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; D’, L.; Mcgowan, A.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef] [Green Version]
- Wilke, C.O. Streamlined Plot Theme and Plot Annotations for “ggplot2” [R Package Cowplot Version 1.1.1]. Available online: https://cran.r-project.org/package=cowplot (accessed on 9 October 2022).
- Auguie, B. Miscellaneous Functions for “Grid” Graphics [R Package GridExtra Version 2.3]. 2017. Available online: https://search.r-project.org/CRAN/refmans/gridExtra/html/00Index.html (accessed on 3 November 2022).
- Kassambara, A. Ggplot2 Based Publication Ready Plots • Ggpubr. Available online: https://rpkgs.datanovia.com/ggpubr/ (accessed on 9 October 2022).
- Wickham, H.; Bryan, J. Read Excel Files [R Package Readxl Version 1.4.1]. Available online: https://cran.r-project.org/package=readxl (accessed on 9 October 2022).
- Willforss, J.; Chawade, A.; Levander, F. NormalyzerDE: Online Tool for Improved Normalization of Omics Expression Data and High-Sensitivity Differential Expression Analysis. J. Proteome Res. 2019, 18, 732–740. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma Powers Differential Expression Analyses for RNA-Sequencing and Microarray Studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Kolde, R. Pheatmap: Pretty Heatmaps. Available online: https://cran.r-project.org/web/packages/pheatmap/index.html (accessed on 9 October 2022).
- Jain, A.; Nandakumar, K.; Ross, A. Score Normalization in Multimodal Biometric Systems. Pattern Recognit. 2005, 38, 2270–2285. [Google Scholar] [CrossRef]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. ClusterProfiler 4.0: A Universal Enrichment Tool for Interpreting Omics Data. Innov. Camb. 2021, 2, 100141. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. ClusterProfiler: An R Package for Comparing Biological Themes among Gene Clusters. OMICS 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Carlson, M. Org.Hs.Eg.Db: Genome Wide Annotation for Human. R Package Version 3.8.2. Available online: https://bioconductor.org/packages/release/data/annotation/html/org.Hs.eg.db.html (accessed on 10 October 2022).
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the Unification of Biology. Nat. Genet. 2000 251 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Benjaminit, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. B 1995, 57, 289–300. [Google Scholar]
- Kanehisa, M. Toward Understanding the Origin and Evolution of Cellular Organisms. Protein Sci. 2019, 28, 1947–1951. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Ishiguro-Watanabe, M.; Tanabe, M. KEGG: Integrating Viruses and Cellular Organisms. Nucleic Acids Res. 2021, 49, D545–D551. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Gillespie, M.; Jassal, B.; Stephan, R.; Milacic, M.; Rothfels, K.; Senff-Ribeiro, A.; Griss, J.; Sevilla, C.; Matthews, L.; Gong, C.; et al. The Reactome Pathway Knowledgebase 2022. Nucleic Acids Res. 2022, 50, D687–D692. [Google Scholar] [CrossRef] [PubMed]
- Martens, M.; Ammar, A.; Riutta, A.; Waagmeester, A.; Slenter, D.N.; Hanspers, K.; Miller, R.A.; Digles, D.; Lopes, E.N.; Ehrhart, F.; et al. WikiPathways: Connecting Communities. Nucleic Acids Res. 2021, 49, D613–D621. [Google Scholar] [CrossRef] [PubMed]
- Mudunuri, U.; Che, A.; Yi, M.; Stephens, R.M. BioDBnet: The Biological Database Network. Bioinformatics 2009, 25, 555–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krämer, A.; Green, J.; Pollard, J.; Tugendreich, S. Causal Analysis Approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef]
- Oughtred, R.; Rust, J.; Chang, C.; Breitkreutz, B.J.; Stark, C.; Willems, A.; Boucher, L.; Leung, G.; Kolas, N.; Zhang, F.; et al. The BioGRID Database: A Comprehensive Biomedical Resource of Curated Protein, Genetic, and Chemical Interactions. Protein Sci. 2021, 30, 187. [Google Scholar] [CrossRef]
- Guo, K.; Maintainer, B.M. Package “VennDetail” Type Package Title A Package for Visualization and Extract Details Version 1.12.0. 2022. Available online: https://webcache.googleusercontent.com/search?q=cache:VazZUzsMo70J:https://www.bioconductor.org/packages/release/bioc//manuals/VennDetail/man/VennDetail.pdf&cd=1&hl=zh-CN&ct=clnk&gl=hk (accessed on 3 November 2022).
- Conway, J.R.; Lex, A.; Gehlenborg, N. UpSetR: An R Package for the Visualization of Intersecting Sets and Their Properties. Bioinformatics 2017, 33, 2938–2940. [Google Scholar] [CrossRef] [Green Version]
- Okuda, S.; Watanabe, Y.; Moriya, Y.; Kawano, S.; Yamamoto, T.; Matsumoto, M.; Takami, T.; Kobayashi, D.; Araki, N.; Yoshizawa, A.C.; et al. JPOSTrepo: An International Standard Data Repository for Proteomes. Nucleic Acids Res. 2017, 45, D1107–D1111. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cervilla-Martínez, J.F.; Rodríguez-Gotor, J.J.; Wypijewski, K.J.; Fontán-Lozano, Á.; Wang, T.; Santamaría, E.; Fuller, W.; Mejías, R. Altered Cortical Palmitoylation Induces Widespread Molecular Disturbances in Parkinson’s Disease. Int. J. Mol. Sci. 2022, 23, 14018. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232214018
Cervilla-Martínez JF, Rodríguez-Gotor JJ, Wypijewski KJ, Fontán-Lozano Á, Wang T, Santamaría E, Fuller W, Mejías R. Altered Cortical Palmitoylation Induces Widespread Molecular Disturbances in Parkinson’s Disease. International Journal of Molecular Sciences. 2022; 23(22):14018. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232214018
Chicago/Turabian StyleCervilla-Martínez, Juan F., Juan J. Rodríguez-Gotor, Krzysztof J. Wypijewski, Ángela Fontán-Lozano, Tao Wang, Enrique Santamaría, William Fuller, and Rebeca Mejías. 2022. "Altered Cortical Palmitoylation Induces Widespread Molecular Disturbances in Parkinson’s Disease" International Journal of Molecular Sciences 23, no. 22: 14018. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232214018