
SMILE Downregulation during Melanogenesis Induces MITF Transcription in B16F10 Cells
 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. SMILE Inversely Correlates with MITF in Human Melanoma Biopsies
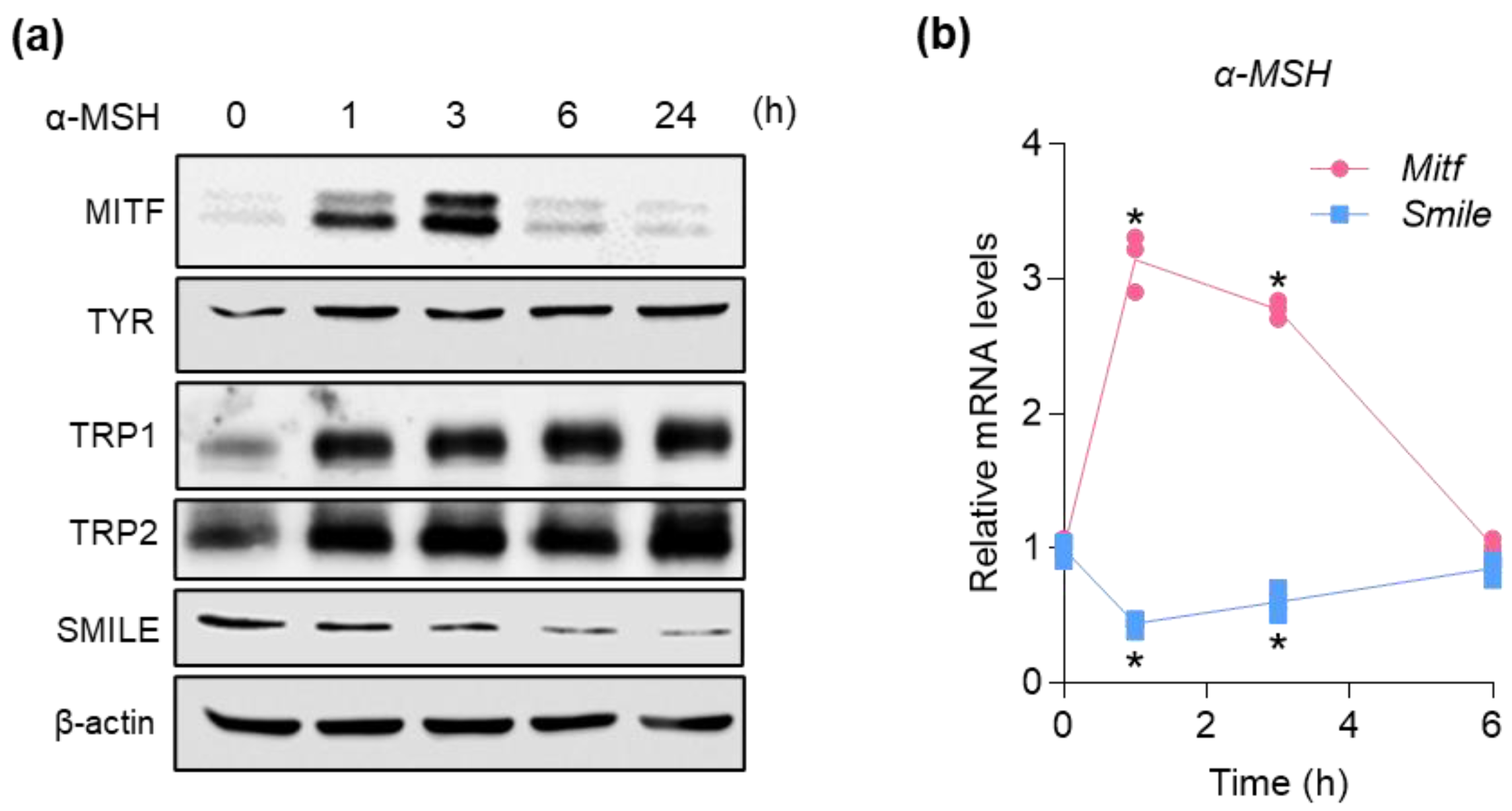
2.2. SMILE and MITF Are Reciprocally Regulated by α-MSH in B16F10 Mouse Melanoma Cells
2.3. SMILE Downregulates Melanogenesis in Melanoma Cells
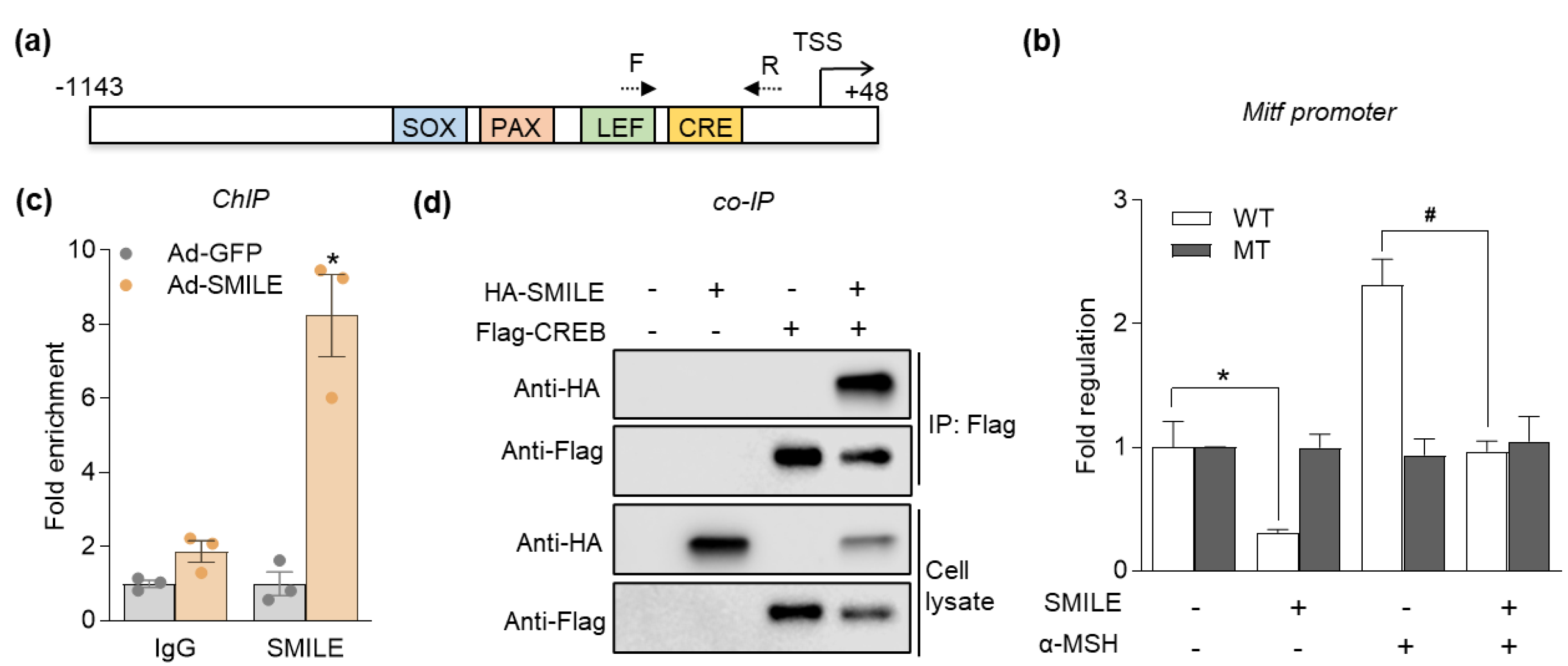
2.4. SMILE Suppresses Transcription of MITF via Binding to CREB
3. Discussion
4. Materials and Methods
4.1. Dataset Preparation for SMILE and MITF Expression Analysis
4.2. Materials and Cell Culture
4.3. RNA Isolation and Analysis
4.4. Immunoblotting
4.5. Luciferase Reporter Assay
4.6. Determination of Melanin Content
4.7. Measurement of Tyrosinase Activity
4.8. Chromatin Immunoprecipitation Assay
4.9. Co-Immunoprecipitation
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jablonski, N.G. The evolution of human skin and skin color. Annu. Rev. Anthropol. 2004, 33, 585–623. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.Y. Skin pigmentation abnormalities and their possible relationship with skin aging. Int. J. Mol. Sci. 2021, 22, 3727. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.Y.; Fisher, D.E. Melanocyte biology and skin pigmentation. Nature 2007, 445, 843–850. [Google Scholar] [CrossRef]
- D’Mello, S.A.; Finlay, G.J.; Baguley, B.C.; Askarian-Amiri, M.E. Signaling pathways in melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar] [CrossRef] [Green Version]
- Levy, C.; Khaled, M.; Fisher, D.E. MITF: Master regulator of melanocyte development and melanoma oncogene. Trends Mol. Med. 2006, 12, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Hartman, M.L.; Czyz, M. MITF in melanoma: Mechanisms behind its expression and activity. Cell. Mol. Life Sci. 2015, 72, 1249–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber, W.E.; Price, E.R.; Widlund, H.R.; Du, J.; Davis, I.J.; Wegner, M.; Fisher, D.E. A tissue-restricted cAMP transcriptional response: SOX10 modulates alpha-melanocyte-stimulating hormone-triggered expression of microphthalmia-associated transcription factor in melanocytes. J. Biol. Chem. 2003, 278, 45224–45230. [Google Scholar] [CrossRef] [Green Version]
- Shoag, J.; Haq, R.; Zhang, M.; Liu, L.; Rowe, G.C.; Jiang, A.; Koulisis, N.; Farrel, C.; Amos, C.I.; Wei, Q.; et al. PGC-1 coactivators regulate MITF and the tanning response. Mol. Cell 2013, 49, 145–157. [Google Scholar] [CrossRef] [Green Version]
- Lu, R.; Misra, V. Zhangfei: A second cellular protein interacts with herpes simplex virus accessory factor HCF in a manner similar to Luman and VP16. Nucleic Acids Res. 2000, 28, 2446–2454. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.B.; Lee, O.H.; Nedumaran, B.; Seong, H.A.; Lee, K.M.; Ha, H.; Lee, I.K.; Yun, Y.; Choi, H.S. SMILE, a new orphan nuclear receptor SHP-interacting protein, regulates SHP-repressed estrogen receptor transactivation. Biochem. J. 2008, 416, 463–473. [Google Scholar] [CrossRef]
- Lee, J.M.; Seo, W.Y.; Han, H.S.; Oh, K.J.; Lee, Y.S.; Kim, D.K.; Choi, S.; Choi, B.H.; Harris, R.A.; Lee, C.H.; et al. Insulin-inducible SMILE inhibits hepatic gluconeogenesis. Diabetes 2016, 65, 62–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, J.; Jiang, G.; Mao, W.; Huang, L.; Huang, C.; Wang, S.; Xue, H.; Ke, J.; Ni, Q. Up-regulation of long noncoding RNA MBNL1-AS1 suppresses breast cancer progression by modulating miR-423-5p/CREBZF axis. Bioengineered 2022, 13, 3707–3723. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Jeong, S.; Jung, W.Y.; Choi, J.W.; Hwang, K.C.; Kim, S.W.; Lee, Y.C. miRNAs as potential biomarkers for the progression of gastric cancer inhibit CREBZF and regulate migration of gastric adenocarcinoma cells. Int. J. Med. Sci. 2020, 17, 693–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.Y.; Song, C.H.; Xie, Y.B.; Jung, C.; Choi, H.S.; Lee, K. SMILE upregulated by metformin inhibits the function of androgen receptor in prostate cancer cells. Cancer Lett. 2014, 354, 390–397. [Google Scholar] [CrossRef]
- Yasumoto, K.; Yokoyama, K.; Takahashi, K.; Tomita, Y.; Shibahara, S. Functional analysis of microphthalmia-associated transcription factor in pigment cell-specific transcription of the human tyrosinase family genes. J. Biol. Chem. 1997, 272, 503–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.M.; Han, H.S.; Jung, Y.S.; Harris, R.A.; Koo, S.H.; Choi, H.S. The SMILE transcriptional corepressor inhibits cAMP response element-binding protein (CREB)-mediated transactivation of gluconeogenic genes. J. Biol. Chem. 2018, 293, 13125–13133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goding, C.R.; Arnheiter, H. MITF-the first 25 years. Genes Dev. 2019, 33, 983–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slominski, A.; Tobin, D.J.; Shibahara, S.; Wortsman, J. Melanin pigmentation in mammalian skin and its hormonal regulation. Physiol. Rev. 2004, 84, 1155–1228. [Google Scholar] [CrossRef]
- Bang, S.; Won, K.H.; Moon, H.R.; Yoo, H.; Hong, A.; Song, Y.; Chang, S.E. Novel regulation of melanogenesis by adiponectin via the AMPK/CRTC pathway. Pigment Cell Melanoma Res. 2017, 30, 553–557. [Google Scholar] [CrossRef]
- Lee, J.M.; Gang, G.T.; Kim, D.K.; Kim, Y.D.; Koo, S.H.; Lee, C.H.; Choi, H.S. Ursodeoxycholic acid inhibits liver X receptor alpha-mediated hepatic lipogenesis via induction of the nuclear corepressor SMILE. J. Biol. Chem. 2014, 289, 1079–1091. [Google Scholar] [CrossRef]
- Lim, H.Y.; Kim, E.; Park, S.H.; Hwang, K.H.; Kim, D.; Jung, Y.J.; Kopalli, S.R.; Hong, Y.D.; Sung, G.H.; Cho, J.Y. Antimelanogenesis effects of theasinensin A. Int. J. Mol. Sci. 2021, 22, 7453. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Kim, K.S.; Lim, D.; Yang, D.J.; Park, J.I.; Kim, K.W.; Jeong, J.H.; Choi, H.S.; Kim, D.K. Epigallocatechin-3-gallate (EGCG)-inducible SMILE inhibits STAT3-mediated hepcidin gene expression. Antioxidants 2020, 9, 514. [Google Scholar] [CrossRef] [PubMed]
- Moon, I.J.; Yoo, H.; Paik, S.H.; Kim, H.T.; Kim, S.Y.; Song, Y.; Chang, S.E. Ursodeoxycholic acid may inhibit environmental aging-associated hyperpigmentation. Antioxidants 2021, 10, 267. [Google Scholar] [CrossRef] [PubMed]
- Lehraiki, A.; Abbe, P.; Cerezo, M.; Rouaud, F.; Regazzetti, C.; Chignon-Sicard, B.; Passeron, T.; Bertolotto, C.; Ballotti, R.; Rocchi, S. Inhibition of melanogenesis by the antidiabetic metformin. J. Investig. Dermatol. 2014, 134, 2589–2597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Park, J.S.; Hwang, S.H.; Cho, K.H.; Na, H.S.; Choi, J.; Jhun, J.; Kim, S.J.; Lee, B.I.; Park, S.H.; et al. Metformin-inducible small heterodimer partner interacting leucine zipper protein ameliorates intestinal inflammation. Front. Immunol. 2021, 12, 652709. [Google Scholar] [CrossRef] [PubMed]
- Talantov, D.; Mazumder, A.; Yu, J.X.; Briggs, T.; Jiang, Y.; Backus, J.; Atkins, D.; Wang, Y. Novel genes associated with malignant melanoma but not benign melanocytic lesions. Clin. Cancer Res. 2005, 11, 7234–7242. [Google Scholar] [CrossRef] [Green Version]
- Truong, X.T.; Park, S.H.; Lee, Y.G.; Jeong, H.Y.; Moon, J.H.; Jeon, T.I. Protocatechuic acid from pear inhibits melanogenesis in melanoma cells. Int. J. Mol. Sci. 2017, 18, 1809. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Truong, X.T.; Lee, Y.-S.; Nguyen, T.T.P.; Kim, H.-J.; Kim, S.-H.; Moon, C.; Kim, D.-K.; Choi, H.-S.; Jeon, T.-I. SMILE Downregulation during Melanogenesis Induces MITF Transcription in B16F10 Cells. Int. J. Mol. Sci. 2022, 23, 15094. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232315094
Truong XT, Lee Y-S, Nguyen TTP, Kim H-J, Kim S-H, Moon C, Kim D-K, Choi H-S, Jeon T-I. SMILE Downregulation during Melanogenesis Induces MITF Transcription in B16F10 Cells. International Journal of Molecular Sciences. 2022; 23(23):15094. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232315094
Chicago/Turabian StyleTruong, Xuan T., Young-Seung Lee, Thuy T. P. Nguyen, Hyun-Jin Kim, Sung-Hak Kim, Changjong Moon, Don-Kyu Kim, Hueng-Sik Choi, and Tae-Il Jeon. 2022. "SMILE Downregulation during Melanogenesis Induces MITF Transcription in B16F10 Cells" International Journal of Molecular Sciences 23, no. 23: 15094. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232315094