
TRPML2 Mucolipin Channels Drive the Response of Glioma Stem Cells to Temozolomide and Affect the Overall Survival in Glioblastoma Patients

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
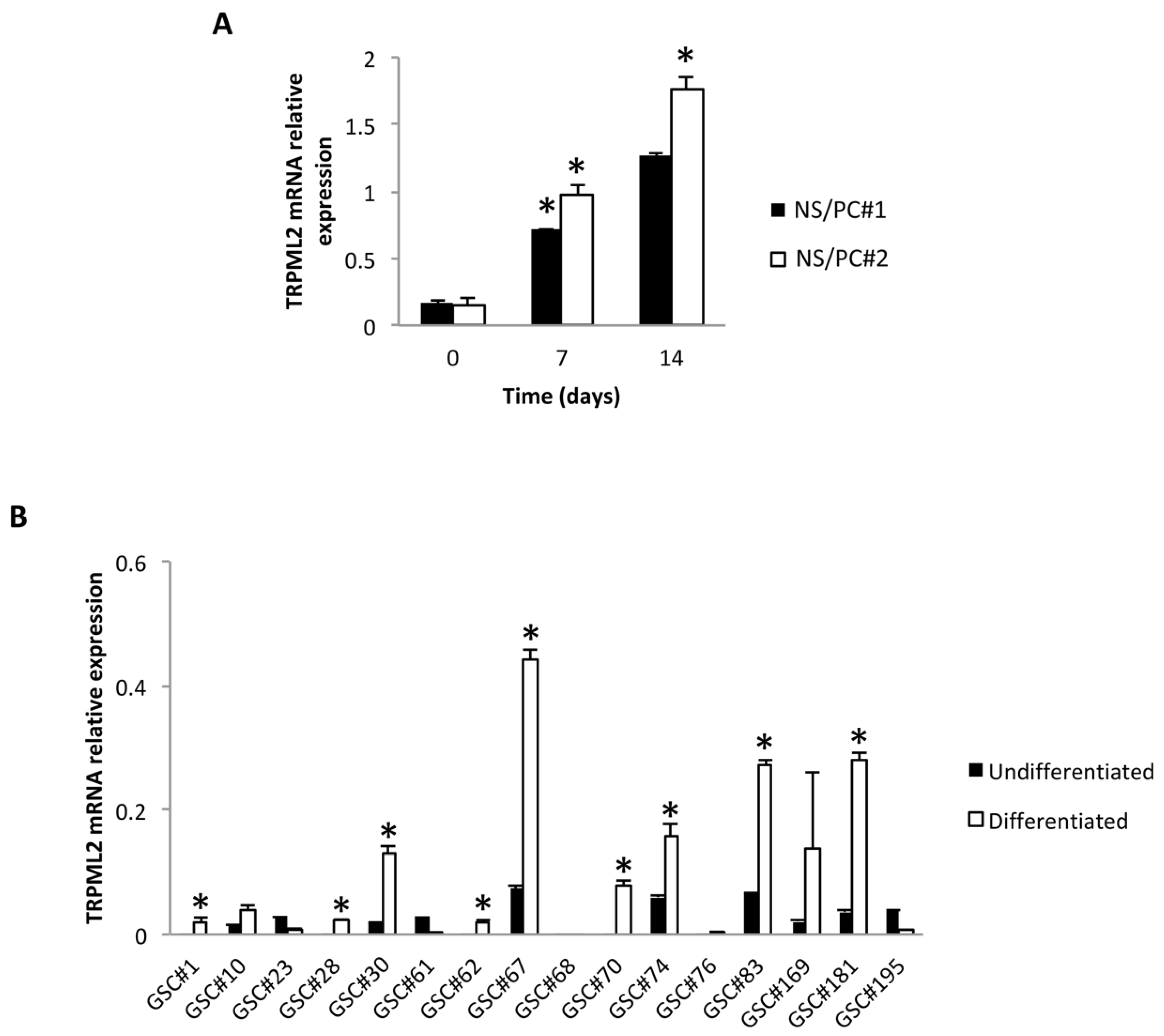
2.1. TRPML2 mRNA Expression during Differentiation of Neural Stem/Progenitor Cells (NS/PCs)
2.2. TRPML2 mRNA Expression in Undifferentiated (GSCs) and Differentiated (D-GSCs) Glioma Stem-like Cell Lines
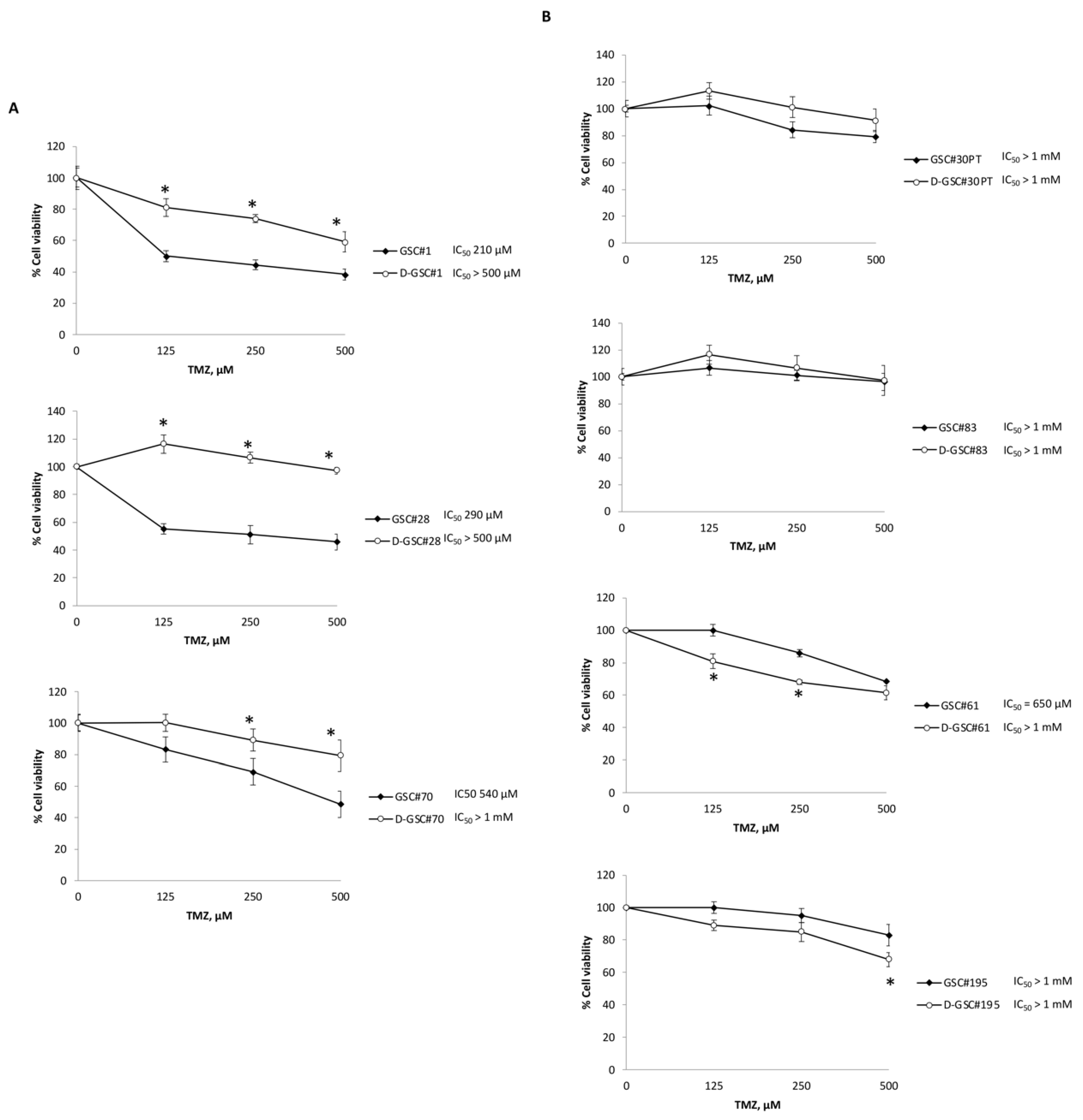
2.3. TMZ Resistance Is Associated with TRPML2 mRNA Expression in Undifferentiated and Differentiated GSCs Lines
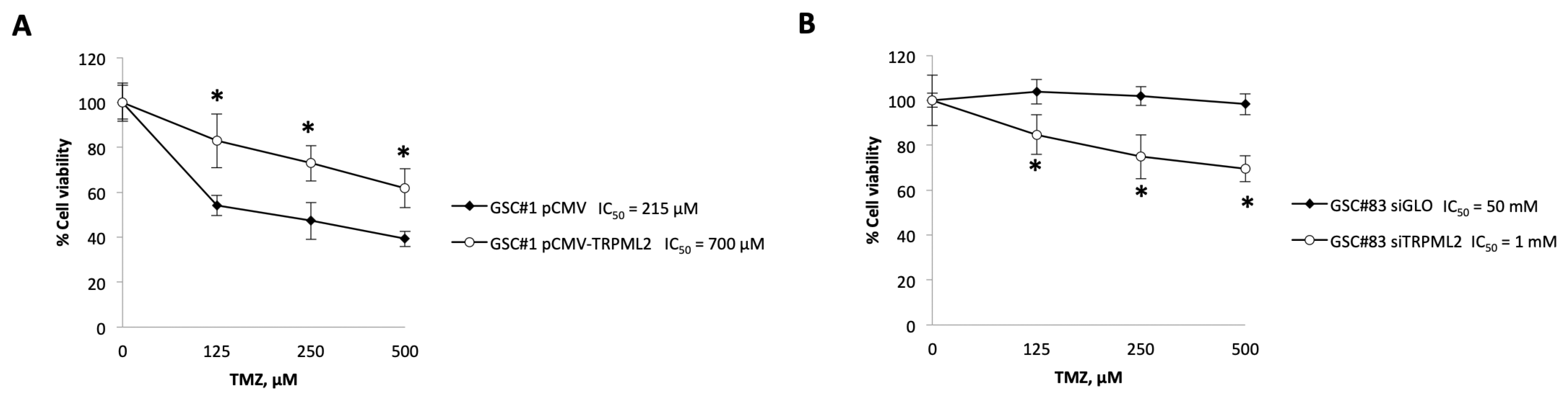
2.4. Changes in the TRPML2 mRNA Expression Result in Modulation of the Responsivity of GSC Lines to TMZ Treatment
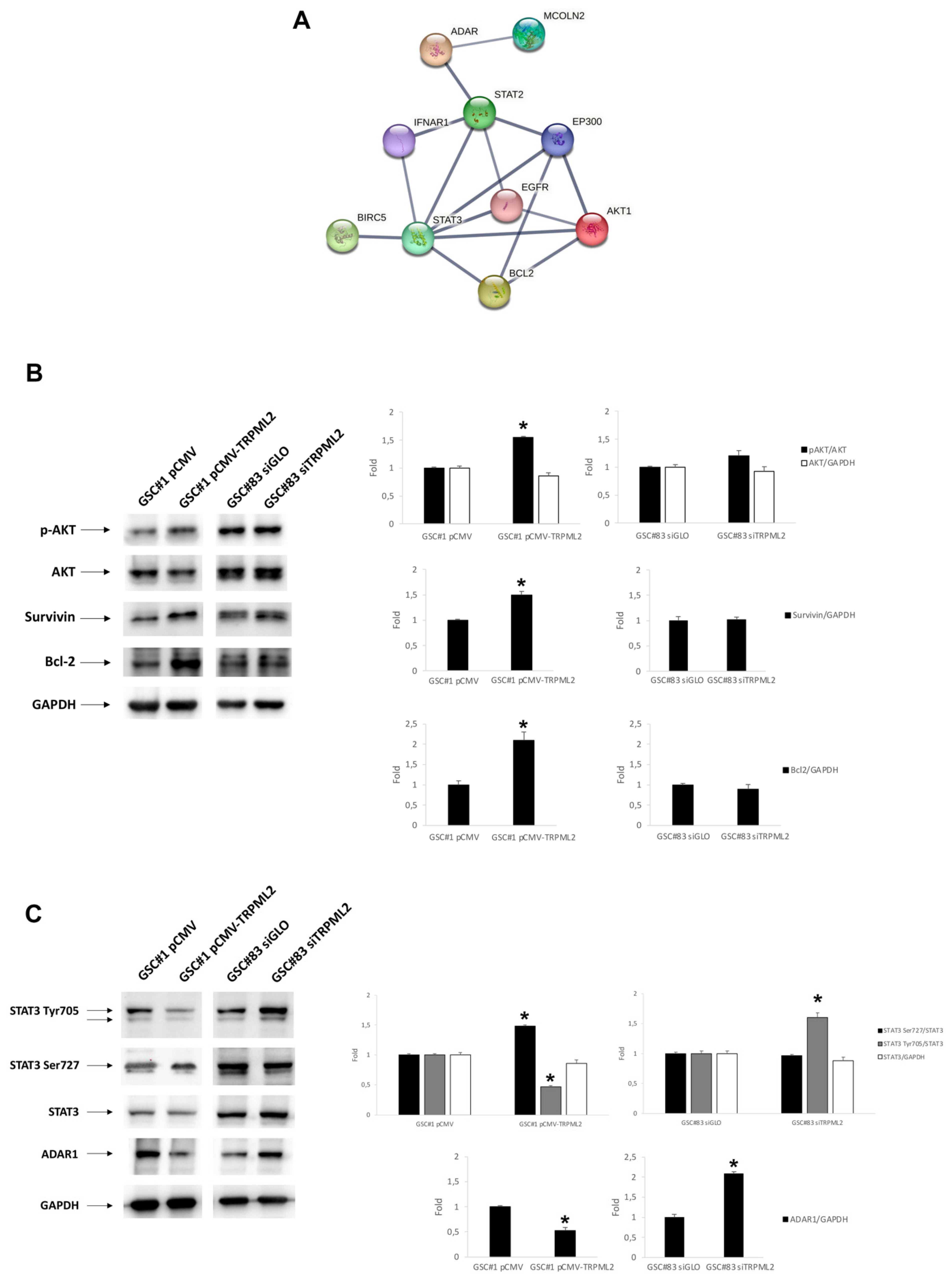
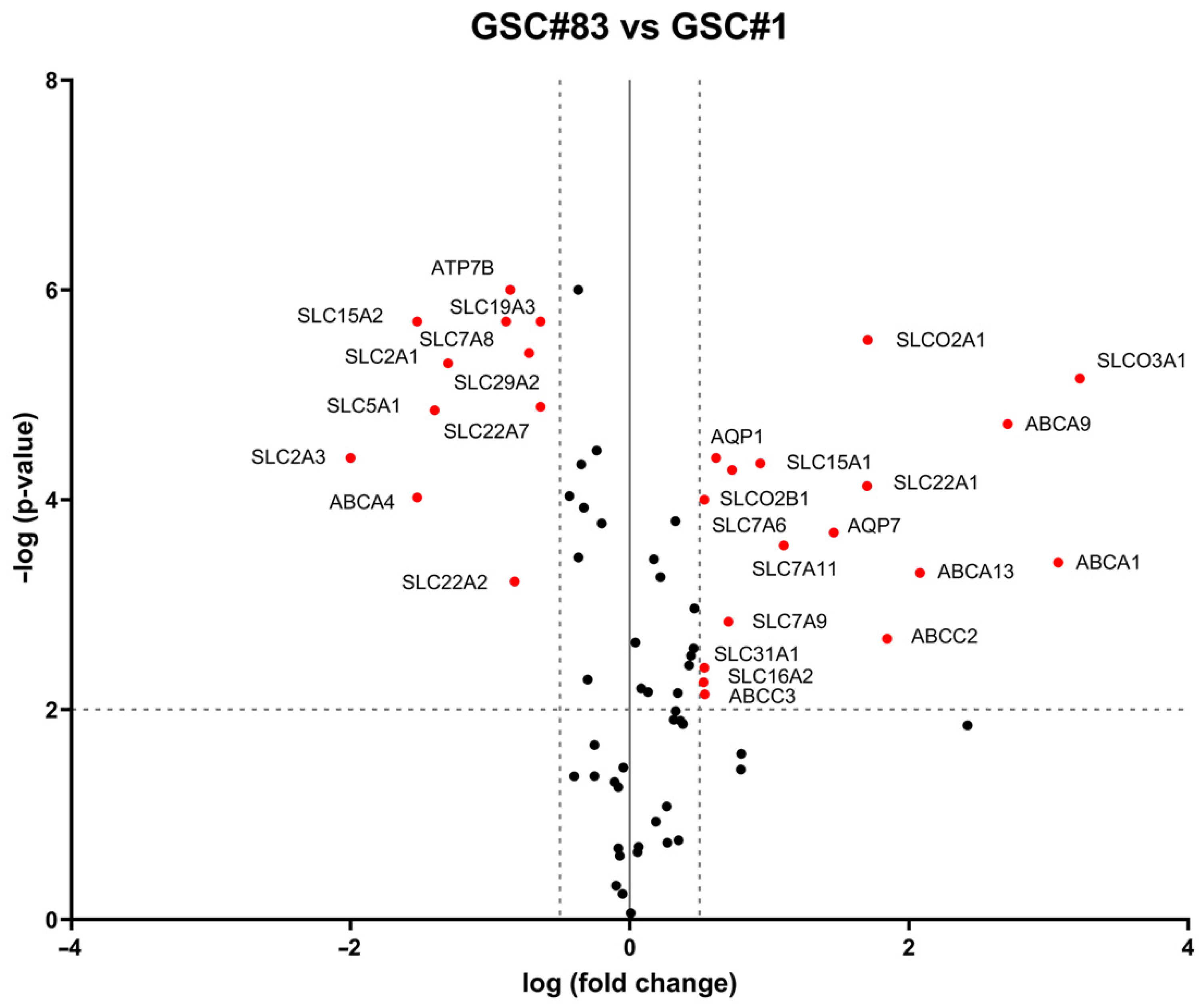
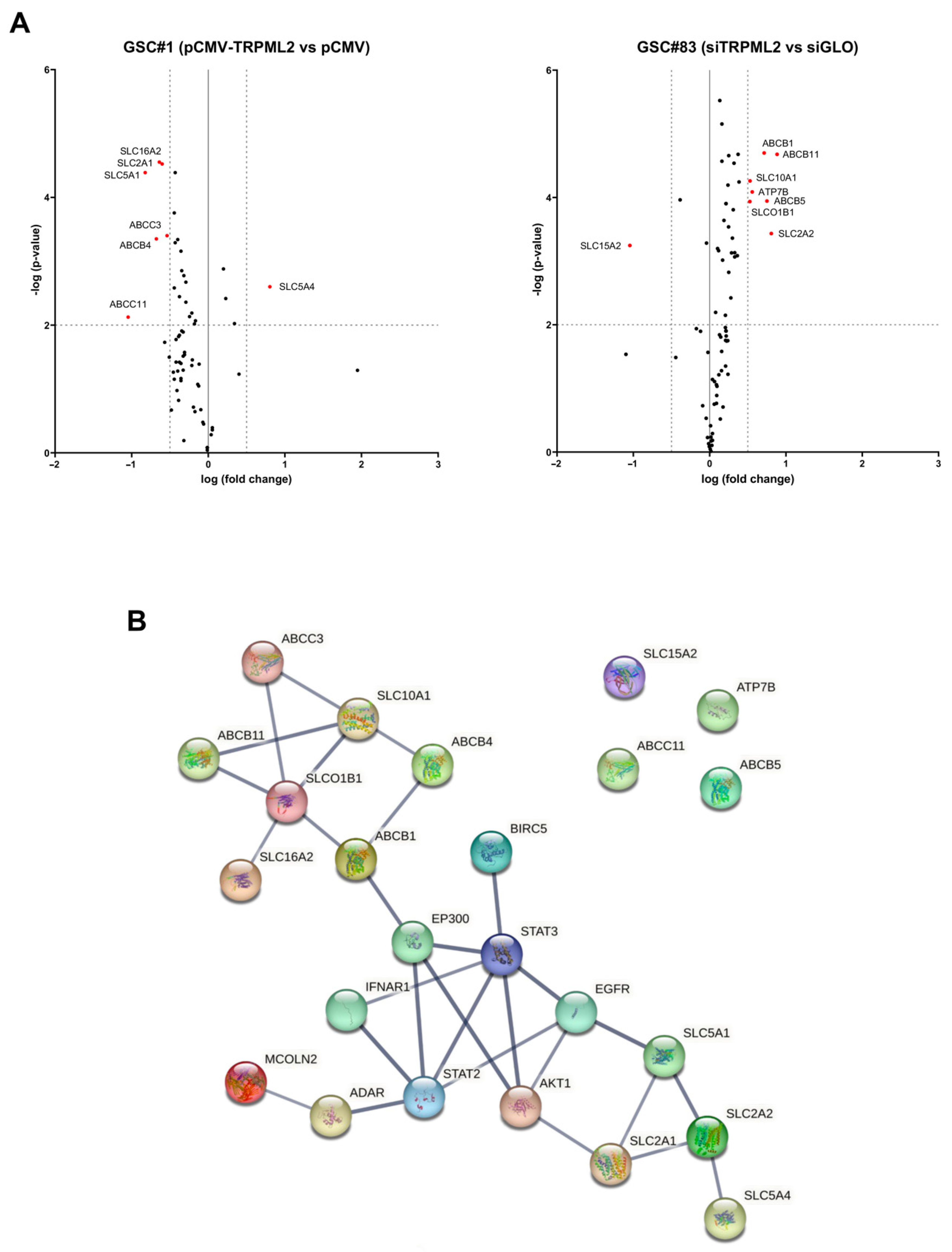
2.5. Drug-Transporter Gene Profile in TRPML2-Negative GSC#1 and TRPML2-Positive GSC#83 Lines
2.6. Drug Transporter Genes Correlated to TRPML2 Expression in GSC Lines
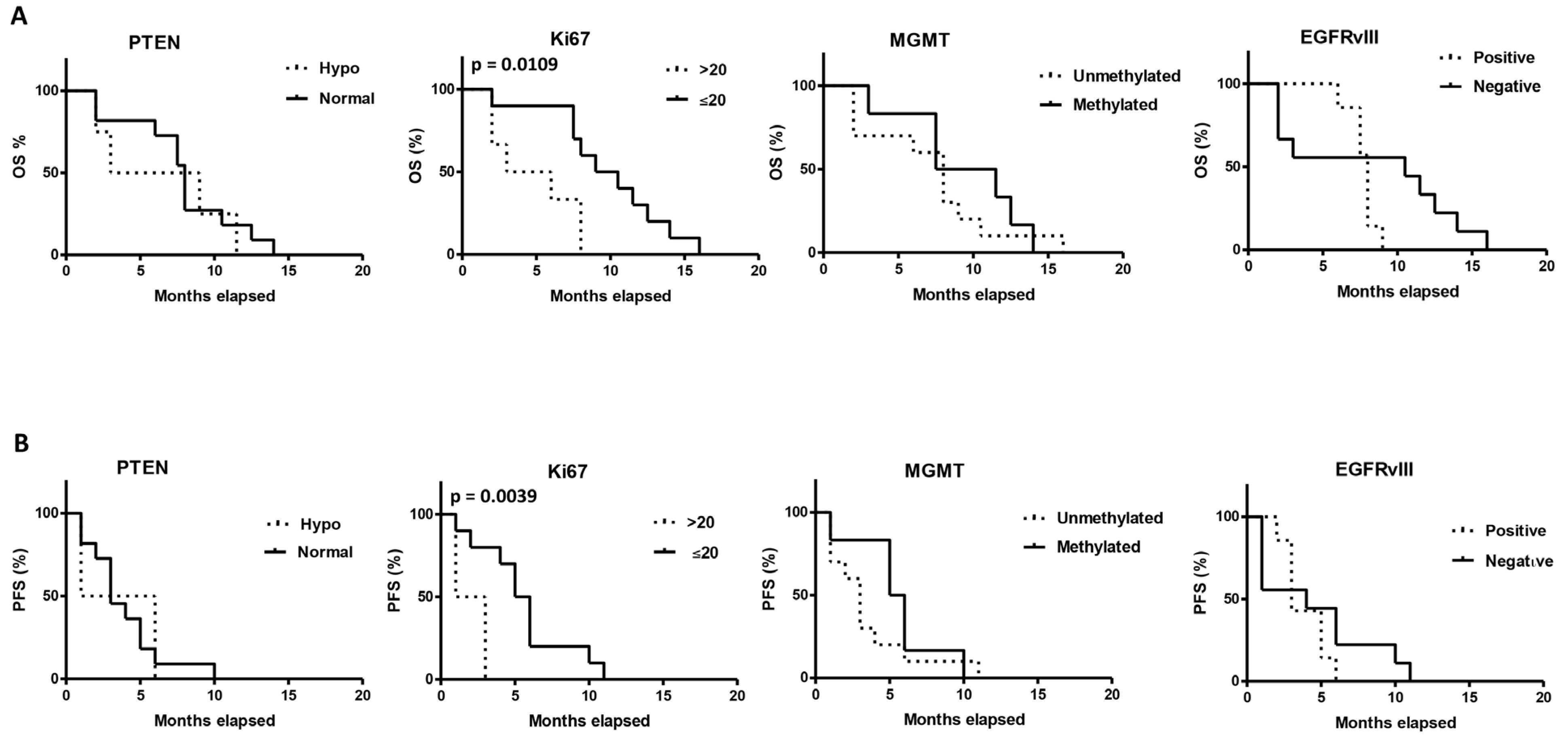
2.7. Correlation between the Tumor and Clinical Characteristics and OS and PFS
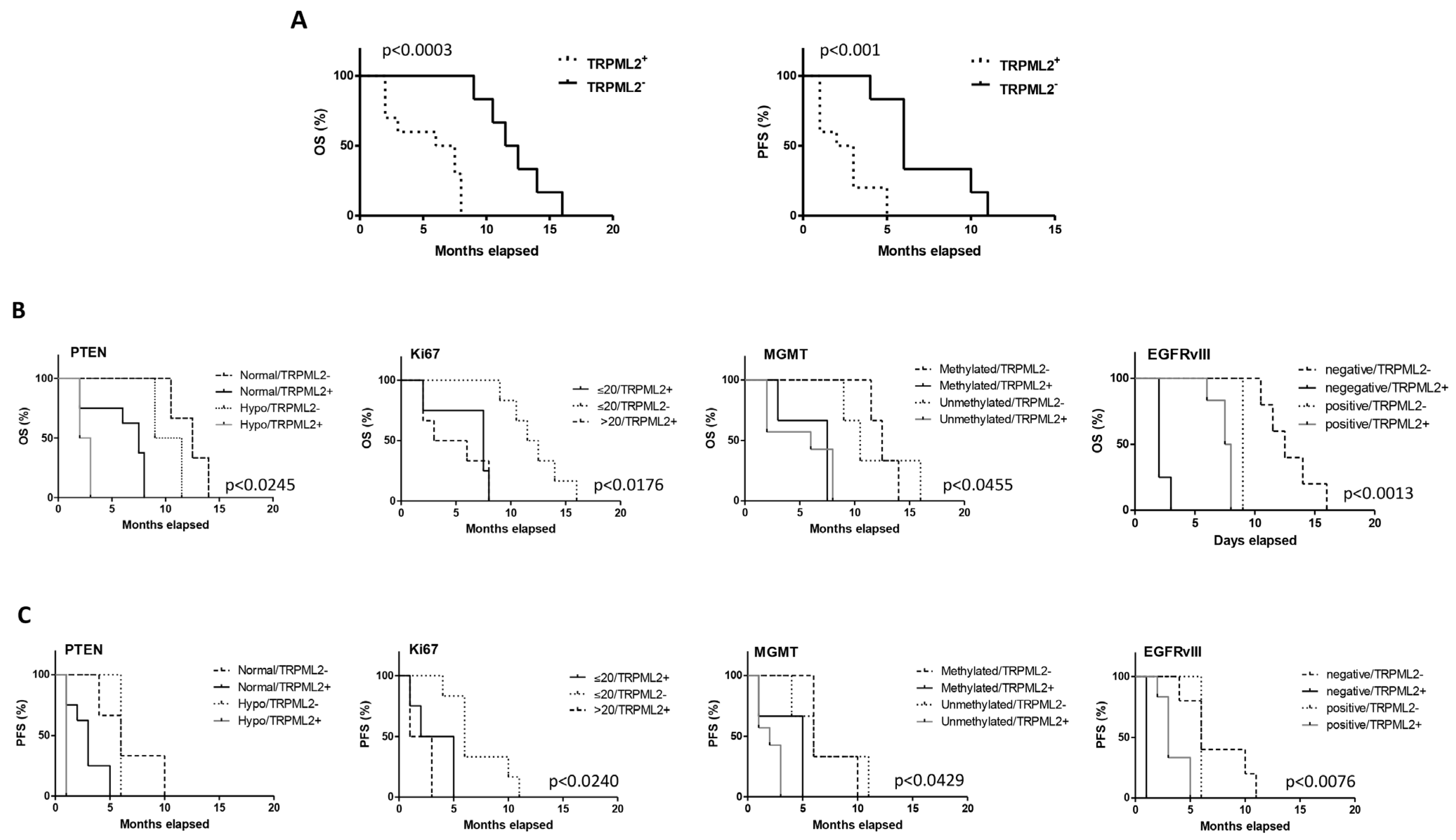
2.8. The TRPML2 mRNA Expression Correlates with Poor OS and PFS in GBM Stem Cell-Derived Patients
3. Discussion
4. Materials and Methods
4.1. Cell Cultures
4.2. Chemical and Reagents
4.3. MTT Assay
4.4. TRPML2 Transfection Models
4.5. Gene Expression Analysis
4.6. RT–PCR Profiler Array
4.7. Western Blot Analysis
4.8. Protein–Protein Interaction (PPI) Network Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maher, E.A.; Furnari, F.B.; Bachoo, R.M.; Rowitch, D.H.; Louis, D.N.; Cavenee, W.K.; DePinho, R.A. Malignant Glioma: Genetics and Biology of a Grave Matter. Genes Dev. 2001, 15, 1311–1333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanai, N.; Alvarez-Buylla, A.; Berger, M.S. Neural Stem Cells and the Origin of Gliomas. N. Engl. J. Med. 2005, 353, 811–822. [Google Scholar] [CrossRef] [PubMed]
- Hemmati, H.D.; Nakano, I.; Lazareff, J.A.; Masterman-Smith, M.; Geschwind, D.H.; Bronner-Fraser, M.; Kornblum, H.I. Cancerous Stem Cells Can Arise from Pediatric Brain Tumors. Proc. Natl. Acad. Sci. USA 2003, 100, 15178–15183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galli, R.; Binda, E.; Orfanelli, U.; Cipelletti, B.; Gritti, A.; de Vitis, S.; Fiocco, R.; Foroni, C.; Dimeco, F.; Vescovi, A. Isolation and Characterization of Tumorigenic, Stem-like Neural Precursors from Human Glioblastoma. Cancer Res. 2004, 64, 7011–7021. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.K.; Clarke, I.D.; Terasaki, M.; Bonn, V.E.; Hawkins, C.; Squire, J.; Dirks, P.B. Identification of a Cancer Stem Cell in Human Brain Tumors. Cancer Res. 2003, 63, 5821–5828. [Google Scholar]
- Kondo, T.; Setoguchi, T.; Taga, T. Persistence of a Small Subpopulation of Cancer Stem-like Cells in the C6 Glioma Cell Line. Proc. Natl. Acad. Sci. USA 2004, 101, 781–786. [Google Scholar] [CrossRef] [Green Version]
- Patrawala, L.; Calhoun, T.; Schneider-Broussard, R.; Zhou, J.; Claypool, K.; Tang, D.G. Side Population Is Enriched in Tumorigenic, Stem-Like Cancer Cells, Whereas ABCG2+ and ABCG2− Cancer Cells Are Similarly Tumorigenic. Cancer Res. 2005, 65, 6207–6219. [Google Scholar] [CrossRef] [Green Version]
- Dirks, P.B. Stem Cells and Brain Tumours. Nature 2006, 444, 687–688. [Google Scholar] [CrossRef]
- Guo, J.; Shan, C.; Xu, J.; Li, M.; Zhao, J.; Cheng, W. New Insights into TRP Ion Channels in Stem Cells. Int. J. Mol. Sci. 2022, 23, 7766. [Google Scholar] [CrossRef]
- Sun, L.; Hua, Y.; Vergarajauregui, S.; Diab, H.I.; Puertollano, R. Novel Role of TRPML2 in the Regulation of the Innate Immune Response. J. Immunol. 2015, 195, 4922–4932. [Google Scholar] [CrossRef] [Green Version]
- Cuajungco, M.P.; Silva, J.; Habibi, A.; Valadez, J.A. The Mucolipin-2 (TRPML2) Ion Channel: A Tissue-Specific Protein Crucial to Normal Cell Function. Pflug. Arch. 2016, 468, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Almamun, M.; Levinson, B.T.; van Swaay, A.C.; Johnson, N.T.; McKay, S.D.; Arthur, G.L.; Davis, J.W.; Taylor, K.H. Integrated Methylome and Transcriptome Analysis Reveals Novel Regulatory Elements in Pediatric Acute Lymphoblastic Leukemia. Epigenetics 2015, 10, 882–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, D.; Luo, X. Identification of Four Methylation-Driven Genes as Candidate Biomarkers for Monitoring Single-Walled Carbon Nanotube-Induced Malignant Transformation of the Lung. Toxicol. Appl. Pharm. 2021, 412, 115391. [Google Scholar] [CrossRef] [PubMed]
- Han, P.; Liu, G.; Lu, X.; Cao, M.; Yan, Y.; Zou, J.; Li, X.; Wang, G. CDH1 Rs9929218 Variant at 16q22.1 Contributes to Colorectal Cancer Susceptibility. Oncotarget 2016, 7, 47278–47286. [Google Scholar] [CrossRef]
- Jung, J.; Venkatachalam, K. TRPML1 and RAS-Driven Cancers—Exploring a Link with Great Therapeutic Potential. Channels 2019, 13, 374–381. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Xie, M.; Meng, Z.; Lo, C.-Y.; Chan, F.L.; Jiang, L.; Meng, X.; Yao, X. Endolysosomal Ion Channel MCOLN2 (Mucolipin-2) Promotes Prostate Cancer Progression via IL-1β/NF-ΚB Pathway. Br. J. Cancer 2021, 125, 1420–1431. [Google Scholar] [CrossRef]
- Huang, C.-C.; Tu, S.-H.; Lien, H.-H.; Jeng, J.-Y.; Huang, C.-S.; Huang, C.-J.; Lai, L.-C.; Chuang, E.Y. Concurrent Gene Signatures for Han Chinese Breast Cancers. PLoS ONE 2013, 8, e76421. [Google Scholar] [CrossRef] [Green Version]
- Morelli, M.B.; Nabissi, M.; Amantini, C.; Rossi, F.; Cardinali, C.; Santoni, M.; Oliva, M.A.; Santoni, A.; Polidori, C.; Mariani, M.P.; et al. Overexpression of Transient Receptor Potential Mucolipin-2 Ion Channels in Gliomas: Role in Tumor Growth and Progression. Oncotarget 2016, 7, 43654–43668. [Google Scholar] [CrossRef] [Green Version]
- Santoni, G.; Amantini, C.; Nabissi, M.; Arcella, A.; Maggi, F.; Santoni, M.; Morelli, M.B. Functional In Vitro Assessment of VEGFA/NOTCH2 Signaling Pathway and PRB Proteasomal Degradation and the Clinical Relevance of Mucolipin TRPML2 Overexpression in Glioblastoma Patients. Int. J. Mol. Sci. 2022, 23, 688. [Google Scholar] [CrossRef]
- Santoni, G.; Maggi, F.; Amantini, C.; Arcella, A.; Marinelli, O.; Nabissi, M.; Santoni, M.; Morelli, M.B. Coexpression of TRPML1 and TRPML2 Mucolipin Channels Affects the Survival of Glioblastoma Patients. Int. J. Mol. Sci. 2022, 23, 7741. [Google Scholar] [CrossRef]
- Fang, C.; Xu, H.; Liu, Y.; Huang, C.; Wang, X.; Zhang, Z.; Xu, Y.; Yuan, L.; Zhang, A.; Shao, A.; et al. TRP Family Genes Are Differently Expressed and Correlated with Immune Response in Glioma. Brain Sci. 2022, 12, 662. [Google Scholar] [CrossRef] [PubMed]
- Ricci-Vitiani, L.; Pallini, R.; Biffoni, M.; Todaro, M.; Invernici, G.; Cenci, T.; Maira, G.; Parati, E.A.; Stassi, G.; Larocca, L.M.; et al. Tumour Vascularization via Endothelial Differentiation of Glioblastoma Stem-like Cells. Nature 2010, 468, 824–828. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Miner, A.; Hennis, L.; Mittal, S. Mechanisms of Temozolomide Resistance in Glioblastoma—A Comprehensive Review. Cancer Drug Resist. 2021, 4, 17–43. [Google Scholar] [CrossRef]
- Kang, H.; Lee, H.; Kim, D.; Kim, B.; Kang, J.; Kim, H.Y.; Youn, H.; Youn, B. Targeting Glioblastoma Stem Cells to Overcome Chemoresistance: An Overview of Current Therapeutic Strategies. Biomedicines 2022, 10, 1308. [Google Scholar] [CrossRef] [PubMed]
- Borgström, A.; Peinelt, C.; Stokłosa, P. TRPM4 in Cancer—A New Potential Drug Target. Biomolecules 2021, 11, 229. [Google Scholar] [CrossRef]
- Elzamzamy, O.M.; Penner, R.; Hazlehurst, L.A. The Role of TRPC1 in Modulating Cancer Progression. Cells 2020, 9, 388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koso, H.; Takeda, H.; Yew, C.C.K.; Ward, J.M.; Nariai, N.; Ueno, K.; Nagasaki, M.; Watanabe, S.; Rust, A.G.; Adams, D.J.; et al. Transposon Mutagenesis Identifies Genes That Transform Neural Stem Cells into Glioma-Initiating Cells. Proc. Natl. Acad. Sci. USA 2012, 109, E2998–E3007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hersh, A.M.; Gaitsch, H.; Alomari, S.; Lubelski, D.; Tyler, B.M. Molecular Pathways and Genomic Landscape of Glioblastoma Stem Cells: Opportunities for Targeted Therapy. Cancers 2022, 14, 3743. [Google Scholar] [CrossRef] [PubMed]
- Colwell, N.; Larion, M.; Giles, A.J.; Seldomridge, A.N.; Sizdahkhani, S.; Gilbert, M.R.; Park, D.M. Hypoxia in the Glioblastoma Microenvironment: Shaping the Phenotype of Cancer Stem-like Cells. Neuro Oncol. 2017, 19, 887–896. [Google Scholar] [CrossRef] [Green Version]
- Matarredona, E.R.; Pastor, A.M. Neural Stem Cells of the Subventricular Zone as the Origin of Human Glioblastoma Stem Cells. Therapeutic Implications. Front. Oncol. 2019, 9, 779. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Li, M.; Xu, J.; Cheng, W. The Modulation of Ion Channels in Cancer Chemo-Resistance. Front. Oncol. 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Strobel, H.; Baisch, T.; Fitzel, R.; Schilberg, K.; Siegelin, M.D.; Karpel-Massler, G.; Debatin, K.-M.; Westhoff, M.-A. Temozolomide and Other Alkylating Agents in Glioblastoma Therapy. Biomedicines 2019, 7, 69. [Google Scholar] [CrossRef] [Green Version]
- Coronas, V.; Terrié, E.; Déliot, N.; Arnault, P.; Constantin, B. Calcium Channels in Adult Brain Neural Stem Cells and in Glioblastoma Stem Cells. Front. Cell. Neurosci. 2020, 14, 600018. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.-C. The Molecular Mechanisms of Chemoresistance in Cancers. Oncotarget 2017, 8, 59950–59964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Z.; Tindall, D.J. FOXOs, Cancer and Regulation of Apoptosis. Oncogene 2008, 27, 2312–2319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvesen, G.S.; Duckett, C.S. IAP Proteins: Blocking the Road to Death’s Door. Nat. Rev. Mol. Cell Biol. 2002, 3, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Pan, Y.; Ling, G.; Wang, S.; Huang, M.; Jiang, X.; Ke, Y. Escape of U251 Glioma Cells from Temozolomide-Induced Senescence Was Modulated by CDK1/Survivin Signaling. Am. J. Transl. Res. 2017, 9, 2163–2180. [Google Scholar]
- Fedele, M.; Cerchia, L.; Pegoraro, S.; Sgarra, R.; Manfioletti, G. Proneural-Mesenchymal Transition: Phenotypic Plasticity to Acquire Multitherapy Resistance in Glioblastoma. Int. J. Mol. Sci. 2019, 20, 2746. [Google Scholar] [CrossRef] [Green Version]
- Segerman, A.; Niklasson, M.; Haglund, C.; Bergström, T.; Jarvius, M.; Xie, Y.; Westermark, A.; Sönmez, D.; Hermansson, A.; Kastemar, M.; et al. Clonal Variation in Drug and Radiation Response among Glioma-Initiating Cells Is Linked to Proneural-Mesenchymal Transition. Cell Rep. 2016, 17, 2994–3009. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Varn, F.S.; Park, S.-H.; Yoon, B.W.; Park, H.R.; Lee, C.; Verhaak, R.G.W.; Paek, S.H. Perspective of Mesenchymal Transformation in Glioblastoma. Acta Neuropathol. Commun. 2021, 9, 50. [Google Scholar] [CrossRef]
- Androutsellis-Theotokis, A.; Leker, R.R.; Soldner, F.; Hoeppner, D.J.; Ravin, R.; Poser, S.W.; Rueger, M.A.; Bae, S.-K.; Kittappa, R.; McKay, R.D.G. Notch Signalling Regulates Stem Cell Numbers in vitro and in vivo. Nature 2006, 442, 823–826. [Google Scholar] [CrossRef]
- Lee, E.-S.; Ko, K.-K.; Joe, Y.A.; Kang, S.-G.; Hong, Y.-K. Inhibition of STAT3 Reverses Drug Resistance Acquired in Temozolomide-Resistant Human Glioma Cells. Oncol. Lett. 2011, 2, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Mannoor, K.; Gao, L.; Tan, A.; Guarnera, M.A.; Zhan, M.; Shetty, A.; Stass, S.A.; Xing, L.; Jiang, F. Characterization of MicroRNA Transcriptome in Lung Cancer by Next-Generation Deep Sequencing. Mol. Oncol. 2014, 8, 1208–1219. [Google Scholar] [CrossRef]
- Ding, N.; Wang, S.; Yang, Q.; Li, Y.; Cheng, H.; Wang, J.; Wang, D.; Deng, Y.; Yang, Y.; Hu, S.; et al. Deep Sequencing Analysis of MicroRNA Expression in Human Melanocyte and Melanoma Cell Lines. Gene 2015, 572, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Mannion, N.; Arieti, F.; Gallo, A.; Keegan, L.; O’Connell, M. New Insights into the Biological Role of Mammalian ADARs; the RNA Editing Proteins. Biomolecules 2015, 5, 2338–2362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galeano, F.; Tomaselli, S.; Locatelli, F.; Gallo, A. A-to-I RNA Editing: The “ADAR” Side of Human Cancer. Semin. Cell. Dev. Biol. 2012, 23, 244–250. [Google Scholar] [CrossRef]
- Gumireddy, K.; Li, A.; Kossenkov, A.V.; Sakurai, M.; Yan, J.; Li, Y.; Xu, H.; Wang, J.; Zhang, P.J.; Zhang, L.; et al. The MRNA-Edited Form of GABRA3 Suppresses GABRA3-Mediated Akt Activation and Breast Cancer Metastasis. Nat. Commun. 2016, 7, 10715. [Google Scholar] [CrossRef] [Green Version]
- Shoshan, E.; Mobley, A.K.; Braeuer, R.R.; Kamiya, T.; Huang, L.; Vasquez, M.E.; Salameh, A.; Lee, H.J.; Kim, S.J.; Ivan, C.; et al. Reduced Adenosine-to-Inosine MiR-455-5p Editing Promotes Melanoma Growth and Metastasis. Nat. Cell Biol. 2015, 17, 311–321. [Google Scholar] [CrossRef]
- Jiang, L.; Hao, Y.; Shao, C.; Wu, Q.; Prager, B.C.; Gimple, R.C.; Sulli, G.; Kim, L.J.; Zhang, G.; Qiu, Z.; et al. ADAR1-Mediated RNA Editing Links Ganglioside Catabolism to Glioblastoma Stem Cell Maintenance. J. Clin. Investig. 2022, 132. [Google Scholar] [CrossRef]
- Luo, J.; Bai, R.; Liu, Y.; Bi, H.; Shi, X.; Qu, C. Long Non-Coding RNA ATXN8OS Promotes Ferroptosis and Inhibits the Temozolomide-Resistance of Gliomas through the ADAR/GLS2 Pathway. Brain Res. Bull. 2022, 186, 27–37. [Google Scholar] [CrossRef]
- Hussein, N.A.; Malla, S.; Pasternak, M.A.; Terrero, D.; Brown, N.G.; Ashby, C.R.; Assaraf, Y.G.; Chen, Z.-S.; Tiwari, A.K. The Role of Endolysosomal Trafficking in Anticancer Drug Resistance. Drug Resist. Update 2021, 57, 100769. [Google Scholar] [CrossRef] [PubMed]
- Szakacs, G.; Abele, R. An Inventory of Lysosomal ABC Transporters. FEBS Lett. 2020, 594, 3965–3985. [Google Scholar] [CrossRef] [PubMed]
- Dean, M.; Fojo, T.; Bates, S. Tumour Stem Cells and Drug Resistance. Nat. Rev. Cancer 2005, 5, 275–284. [Google Scholar] [CrossRef]
- Riganti, C.; Salaroglio, I.C.; Caldera, V.; Campia, I.; Kopecka, J.; Mellai, M.; Annovazzi, L.; Bosia, A.; Ghigo, D.; Schiffer, D. Temozolomide Downregulates P-Glycoprotein Expression in Glioblastoma Stem Cells by Interfering with the Wnt3a/Glycogen Synthase-3 Kinase/-Catenin Pathway. Neuro Oncol. 2013, 15, 1502–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pessina, S.; Cantini, G.; Kapetis, D.; Cazzato, E.; di Ianni, N.; Finocchiaro, G.; Pellegatta, S. The Multidrug-Resistance Transporter Abcc3 Protects NK Cells from Chemotherapy in a Murine Model of Malignant Glioma. Oncoimmunology 2016, 5, e1108513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talty, R.; Bosenberg, M. The Role of Ferroptosis in Melanoma. Pigment Cell Melanoma Res. 2022, 35, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Mi, Y.; Qian, H.; Guo, N.; Yan, A.; Zhang, Y.; Gao, X. A Potential Mechanism of Temozolomide Resistance in Glioma–Ferroptosis. Front. Oncol. 2020, 10, 897. [Google Scholar] [CrossRef]
- Li, Q.; Shu, Y. Role of Solute Carriers in Response to Anticancer Drugs. Mol. Cell. Ther. 2014, 2, 15. [Google Scholar] [CrossRef] [Green Version]
- Flavahan, W.A.; Wu, Q.; Hitomi, M.; Rahim, N.; Kim, Y.; Sloan, A.E.; Weil, R.J.; Nakano, I.; Sarkaria, J.N.; Stringer, B.W.; et al. Brain Tumor Initiating Cells Adapt to Restricted Nutrition through Preferential Glucose Uptake. Nat. Neurosci. 2013, 16, 1373–1382. [Google Scholar] [CrossRef]
- Hassen, W.; Kassambara, A.; Reme, T.; Sahota, S.; Seckinger, A.; Vincent, L.; Cartron, G.; Moreaux, J.; Hose, D.; Klein, B. Drug Metabolism and Clearance System in Tumor Cells of Patients with Multiple Myeloma. Oncotarget 2015, 6, 6431–6447. [Google Scholar] [CrossRef] [Green Version]
- Villalva, C.; Cortes, U.; Wager, M.; Tourani, J.-M.; Rivet, P.; Marquant, C.; Martin, S.; Turhan, A.G.; Karayan-Tapon, L. O6-Methylguanine-Methyltransferase (MGMT) Promoter Methylation Status in Glioma Stem-Like Cells Is Correlated to Temozolomide Sensitivity Under Differentiation-Promoting Conditions. Int. J. Mol. Sci. 2012, 13, 6983–6994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, K.; Eldred, B.S.C.; Kevan, B.; Pianka, S.; Eldred, B.A.; Zapanta Rinonos, S.; Yong, W.H.; Liau, L.M.; Nghiemphu, P.L.; Cloughesy, T.F.; et al. Prognostic Value of O6-Methylguanine-DNA Methyltransferase Methylation in Isocitrate Dehydrogenase Mutant Gliomas. Neurooncol. Adv. 2022, 4, vdac030. [Google Scholar] [CrossRef] [PubMed]
- Montano, N.; Cenci, T.; Martini, M.; D’Alessandris, Q.G.; Pelacchi, F.; Ricci-Vitiani, L.; Maira, G.; de Maria, R.; Larocca, L.M.; Pallini, R. Expression of EGFRvIII in Glioblastoma: Prognostic Significance Revisited. Neoplasia 2011, 13, 1113–1121, IN3–IN6. [Google Scholar] [CrossRef] [PubMed]
- Cominelli, M.; Grisanti, S.; Mazzoleni, S.; Branca, C.; Buttolo, L.; Furlan, D.; Liserre, B.; Bonetti, M.F.; Medicina, D.; Pellegrini, V.; et al. EGFR Amplified and Overexpressing Glioblastomas and Association with Better Response to Adjuvant Metronomic Temozolomide. JNCI J. Natl. Cancer Inst. 2015, 107, djv041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, W.; Chen, J.; Guo, X.; Su, Z. Letter Regarding Xiao WZ et al. entitled “Relationships between PTEN Gene Mutations and Prognosis in Glioma: A Meta-Analysis”. Tumor Biol. 2015, 36, 4089. [Google Scholar] [CrossRef] [PubMed]
- Armocida, D.; Frati, A.; Salvati, M.; Santoro, A.; Pesce, A. Is Ki-67 Index Overexpression in IDH Wild Type Glioblastoma a Predictor of Shorter Progression Free Survival? A Clinical and Molecular Analytic Investigation. Clin. Neurol. Neurosurg. 2020, 198, 106126. [Google Scholar] [CrossRef]
- Kleihues, P.; Ohgaki, H. Phenotype vs Genotype in the Evolution of Astrocytic Brain Tumors. Toxicol. Pathol. 2000, 28, 164–170. [Google Scholar] [CrossRef]
- Visconti, P.; Parodi, F.; Parodi, B.; Casarino, L.; Romano, P.; Buccarelli, M.; Pallini, R.; D’Alessandris, Q.G.; Montori, A.; Pilozzi, E.; et al. Short Tandem Repeat Profiling for the Authentication of Cancer Stem-like Cells. Int. J. Cancer 2021, 148, 1489–1498. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING V11: Protein–Protein Association Networks with Increased Coverage, Supporting Functional Discovery in Genome-Wide Experimental Datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morelli, M.B.; Nabissi, M.; Amantini, C.; Maggi, F.; Ricci-Vitiani, L.; Pallini, R.; Santoni, G. TRPML2 Mucolipin Channels Drive the Response of Glioma Stem Cells to Temozolomide and Affect the Overall Survival in Glioblastoma Patients. Int. J. Mol. Sci. 2022, 23, 15356. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232315356
Morelli MB, Nabissi M, Amantini C, Maggi F, Ricci-Vitiani L, Pallini R, Santoni G. TRPML2 Mucolipin Channels Drive the Response of Glioma Stem Cells to Temozolomide and Affect the Overall Survival in Glioblastoma Patients. International Journal of Molecular Sciences. 2022; 23(23):15356. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232315356
Chicago/Turabian StyleMorelli, Maria Beatrice, Massimo Nabissi, Consuelo Amantini, Federica Maggi, Lucia Ricci-Vitiani, Roberto Pallini, and Giorgio Santoni. 2022. "TRPML2 Mucolipin Channels Drive the Response of Glioma Stem Cells to Temozolomide and Affect the Overall Survival in Glioblastoma Patients" International Journal of Molecular Sciences 23, no. 23: 15356. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232315356