


Effect of Overexpression of γ-Tocopherol Methyltransferase on α-Tocopherol and Fatty Acid Accumulation and Tolerance to Salt Stress during Seed Germination in Brassica napus L.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
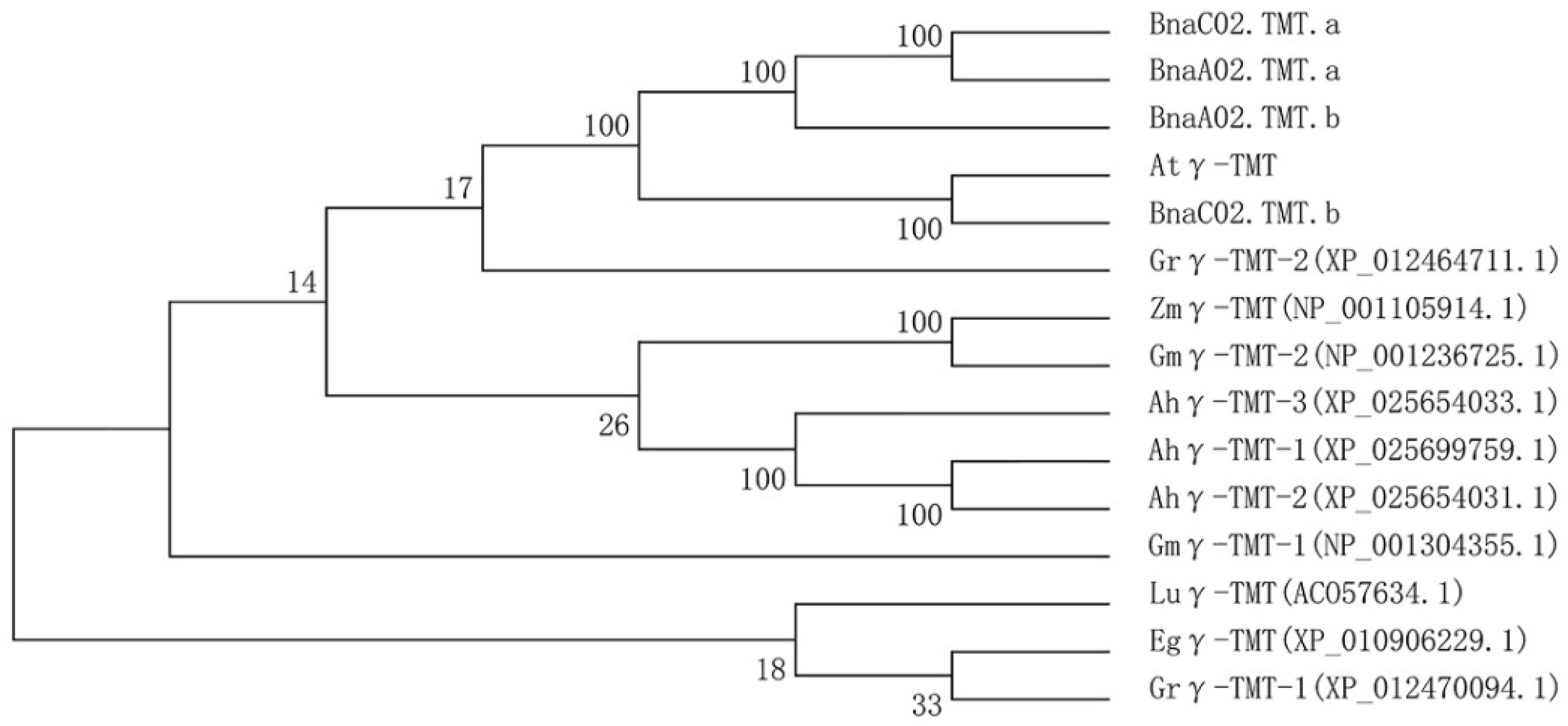
2.1. Sequence Analysis of Bnγ-TMT Paralogs
2.2. Analysis of the Expression Pattern of Bnγ-TMT
2.3. BnaC02.TMT.a Increased α-Tocopherol Content in B. napus Seeds
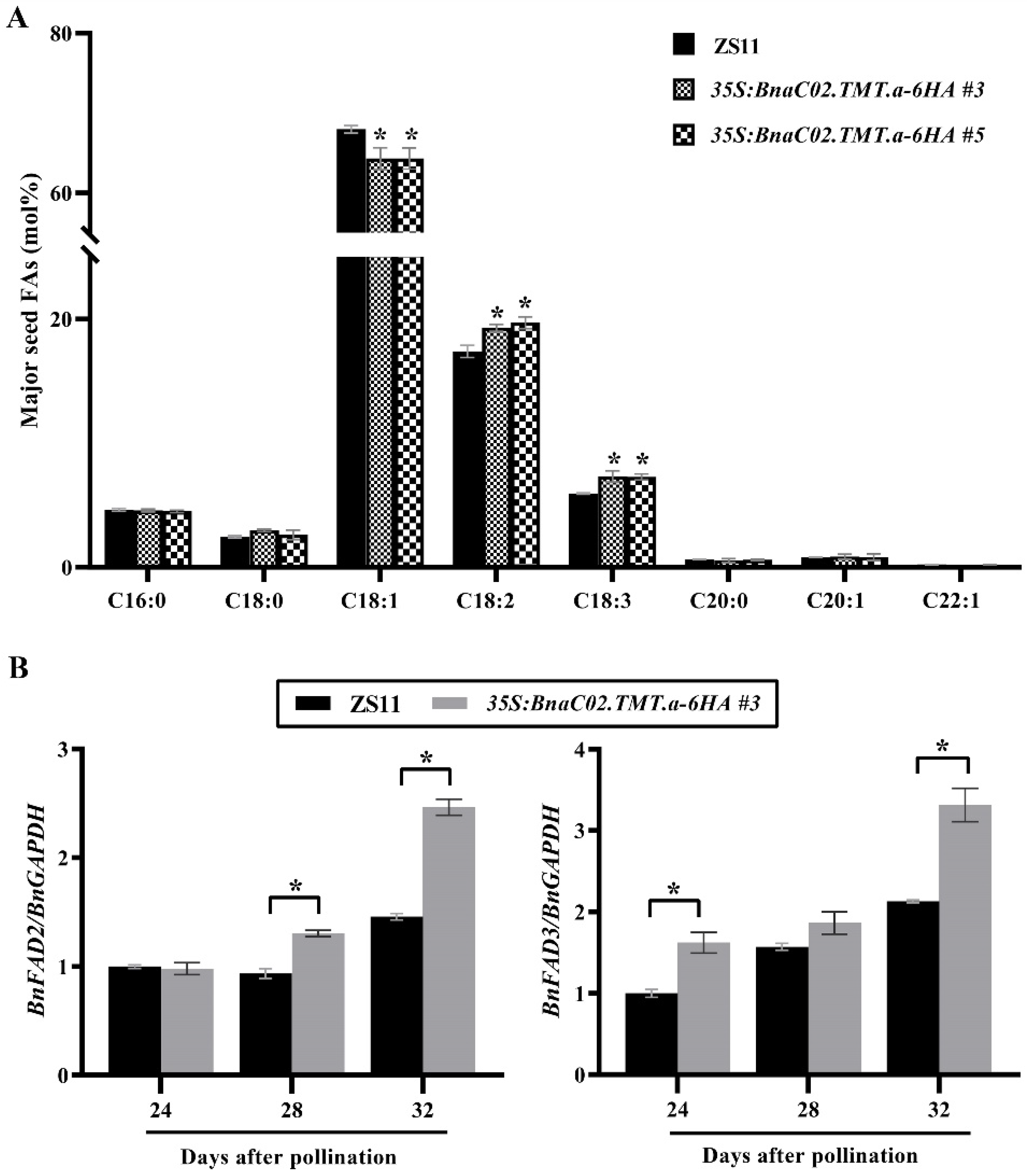
2.4. BnaC02.TMT.a Promoted PUFA Biosynthesis in B. napus Seeds
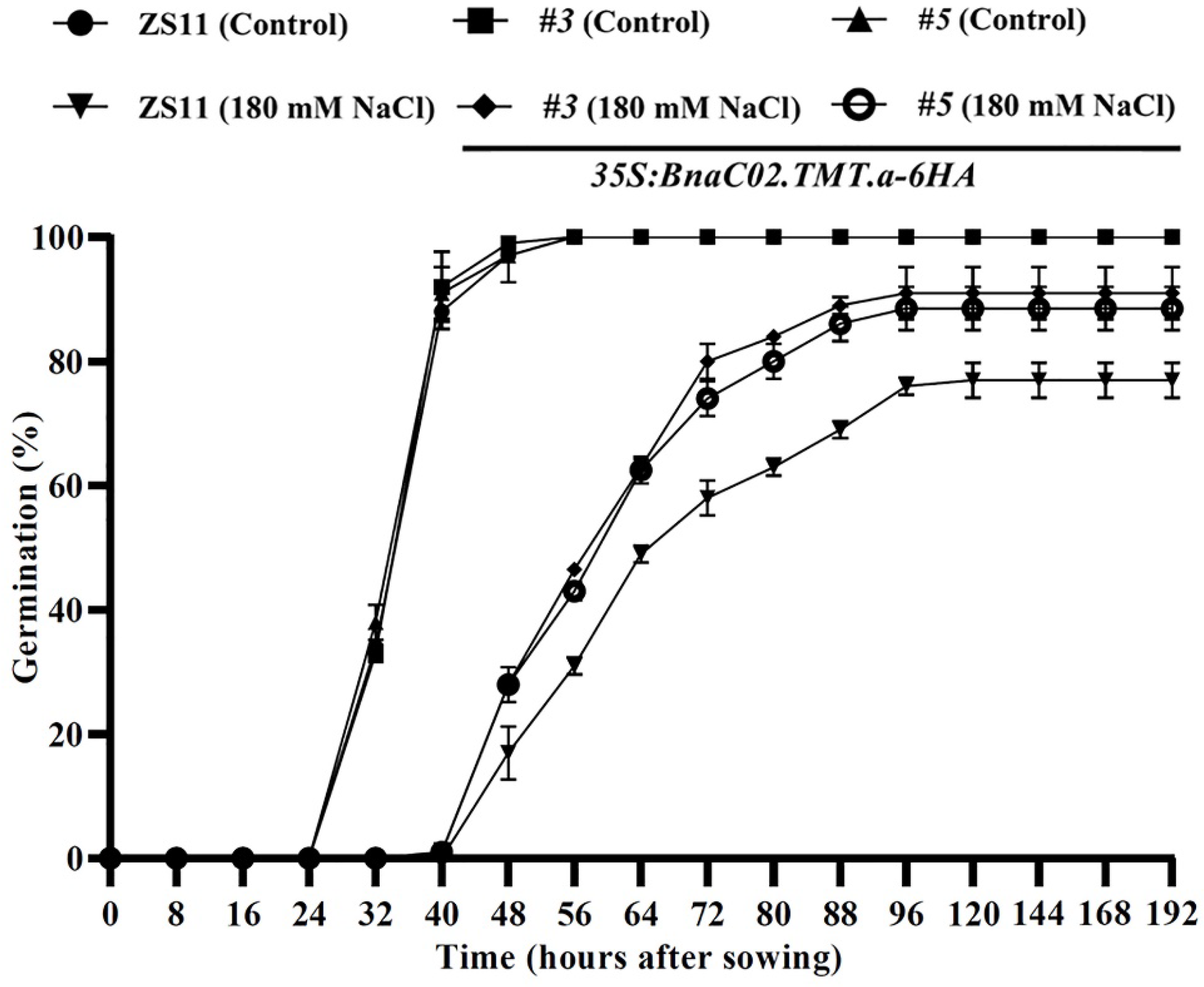
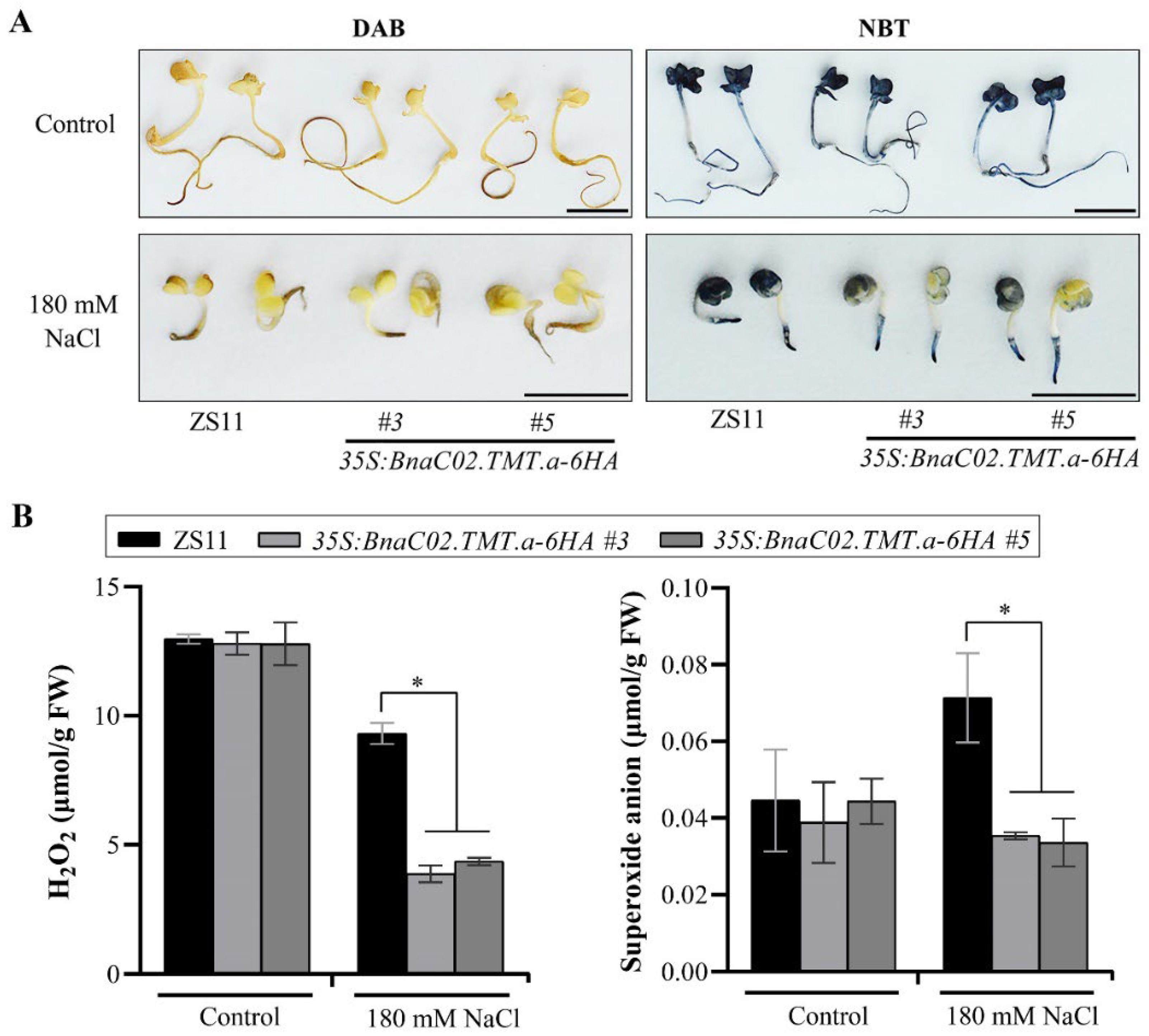
2.5. Functional Characterisation of BnaC02.TMT.a in the Seed Germination under Conditions Involving Salt Stress in B. napus
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Protein Sequence and Phylogenetic Analysis
4.3. Gene Cloning and Plasmid Construction
4.4. Generation of B. napus Transgenic Plants
4.5. Morphological Observation of Mature Seeds
4.6. Measurement of the FA Content
4.7. HPLC Analysis of the Tocopherol Content
4.8. ROS Staining
4.9. ROS Quantification
4.10. Germination Test
4.11. Gene Expression Analysis
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| A. thaliana | Arabidopsis thaliana |
| ANOVA | analysis of variance |
| BHT | butylated hydroxy toluene |
| BnPIR | Brassica napus pan-genome information resource |
| B. juncea | Brassica juncea |
| B. napus | Brassica napus |
| B. oleracea | Brassica oleracea |
| B. rapa | Brassica rapa |
| CDS | coding sequence |
| DAB | diaminobenzidine |
| DAP | days after pollination |
| DMBPQ | 2,3-dimethyl-6-phytyl-1,4-benzoquinol |
| ER | endoplasmic reticulum |
| FAs | fatty acids |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
| GGPP | geranylgeranyl diphosphate |
| GGR | geranylgeranyl reductase |
| HA | haemagglutinin |
| HAS | h after sowing |
| HGA | homogentisic acid |
| H2O2 | hydrogen peroxide |
| HPP | p-hydroxyphenylpyruvic acid |
| HPPD | HPP dioxygenase |
| HPT | HGA phytyl transferase |
| LD | long day |
| MEP | methylerythritol phosphate |
| MES | 2-(N-morpholino) ethanesulfonic acid |
| MBPQ | 2-methyl-6-phytyl-1,4-benzoquinol |
| MPBQ-MT | MPBQ methyltransferase |
| NaCl | sodium chloride |
| NBT | nitroblue tetrazolium |
| NCBI | National Center for Biotechnology Information |
| O2- | superoxide anion |
| PDP | phytyl diphosphate |
| Phytyl-P | phytyl phosphate |
| PK | phytol kinase |
| PPK | phytyl phosphate kinase |
| PUFAs | polyunsaturated FAs |
| ROS | reactive oxygen species |
| γ-TMT | γ-tocopherol methyltransferase |
| RT-qPCR | quantitative reverse transcription PCR |
| SAM | S-adenosylmethionine |
| SD | standard deviation |
| TAT | tyrosine aminotransferase |
| TC | tocopherol cyclase |
| ZS11 | Zhongshuang11 |
References
- Chalhoub, B.; Denoeud, F.; Liu, S.; Parkin, I.A.P.; Tang, H.; Wang, X.; Chiquet, J.; Belcram, H.; Tong, C.; Samans, B.; et al. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science 2014, 345, 950–953. [Google Scholar] [CrossRef] [Green Version]
- Fu, T.D. Variety improvement in rapeseed. Crop. Res. 2007, 3, 159–162. (In Chinese) [Google Scholar]
- Nestel, P.; Clifton, P.; Colquhoun, D.; Noakes, M.; Mori, T.A.; Sullivan, D.; Thomas, B. Indications for omega-3 long chain polyunsaturated fatty acid in the prevention and treatment of cardiovascular disease. Heart Lung Circ. 2015, 24, 769–779. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.V.; Schooling, C.M. Role of linoleic acid in autoimmune disorders: A Mendelian randomisation study. Ann. Rheum. Dis. 2019, 78, 711. [Google Scholar] [CrossRef]
- Djuricic, I.; Calder, P.C. Beneficial outcomes of omega-6 and omega-3 polyunsaturated fatty acids on human health: An update for 2021. Nutrients 2021, 13, 2421. [Google Scholar] [CrossRef]
- Weinstein, S.J.; Wright, M.E.; Lawson, K.A.; Snyder, K.; Mannisto, S.; Taylor, P.R.; Virtamo, J.; Albanes, D. Serum and dietary vitamin E in relation to prostate cancer risk. Cancer Epidemiol. Biomark. Prev. 2007, 16, 1253–1259. [Google Scholar] [CrossRef] [Green Version]
- Euch-Fayache, G.E.; Bouhlal, Y.; Amouri, R.; Feki, M.; Hentati, F. Molecular, clinical and peripheral neuropathy study of Tunisian patients with ataxia with vitamin E deficiency. Brain 2014, 137, 402–410. [Google Scholar] [CrossRef]
- Hunter, S.; Cahoon, E. Enhancing vitamin E in oilseeds: Unraveling tocopherol and tocotrienol biosynthesis. Lipids 2007, 42, 97–108. [Google Scholar] [CrossRef]
- Horvath, G.; Wessjohann, L.; Bigirimana, J.; Jansen, M.; Guisez, Y.; Caubergs, R.; Horemans, N. Differential distribution of tocopherols and tocotrienols in photosynthetic and non-photosynthetic tissues. Phytochemistry 2006, 67, 1185–1195. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Alegre, L. The function of tocopherols and tocotrienols in plants. Crit. Rev. Plant Sci. 2002, 21, 31–57. [Google Scholar] [CrossRef]
- DellaPenna, D.; Last, R.L. Progress in the dissection and manipulation of plant vitamin E biosynthesis. Physiol. Plant. 2006, 126, 356–368. [Google Scholar] [CrossRef]
- Garcia, I.; Rodgers, M.; Lenne, C.; Rolland, A.; Sailland, A.; Matringe, M. Subcellular localization and purification of a p-hydroxyphenylpyruvate dioxygenase from cultured carrot cells and characterization of the corresponding cDNA. Biochem. J. 1997, 325, 761–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riewe, D.; Koohi, M.; Lisec, J.; Pfeiffer, M.; Lippmann, R.; Schmeichel, J.; Willmitzer, L.; Altmann, T. A tyrosine aminotransferase involved in tocopherol synthesis in Arabidopsis. Plant J. 2012, 71, 850–859. [Google Scholar] [CrossRef] [PubMed]
- Keller, Y.; Bouvier, F.; D’Harlingue, A.; Camara, B. Metabolic compartmentation of plastid prenyllipid biosynthesis evidence for the involvement of a multifunctional geranylgeranyl reductase. Eur. J. Biochem. 1998, 251, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Kimura, E.; Abe, T.; Murata, K.; Kimura, T.; Otoki, Y.; Yoshida, T.; Miyazawa, T.; Nakagawa, K. Identification of OsGGR2, a second geranylgeranyl reductase involved in α-tocopherol synthesis in rice. Sci. Rep. 2018, 8, 1870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valentin, H.E.; Lincoln, K.; Moshiri, F.; Jensen, P.K.; Qi, Q.; Venkatesh, T.V.; Karunanandaa, B.; Baszis, S.R.; Norris, S.R.; Savidge, B.; et al. The Arabidopsis vitamin E pathway gene5-1 mutant reveals a critical role for phytol kinase in seed tocopherol biosynthesis. Plant Cell 2006, 18, 212–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorp, K.V.; Hölzl, G.; Plohmann, C.; Eisenhut, M.; Abraham, M.; Weber, A.P.M.; Hanson, A.D.; Dörmann, P. Remobilization of phytol from chlorophyll degradation is essential for tocopherol synthesis and growth of Arabidopsis. Plant Cell 2015, 27, 2846–2859. [Google Scholar]
- Mène-Saffrané, L.; DellaPenna, D. Biosynthesis, regulation and functions of tocochromanols in plants. Plant Physiol. Bioch. 2010, 48, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Fritsche, S.; Wang, X.; Jung, C. Recent advances in our understanding of tocopherol biosynthesis in plants: An overview of key genes, functions, and breeding of vitamin E improved crops. Antioxidants 2017, 6, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shintani, D.; DellaPenna, D. Elevating the vitamin E content of plants through metabolic engineering. Science 1998, 282, 2098–2100. [Google Scholar] [CrossRef]
- Bergmüller, E.; Porfirova, S.; Dörmann, P. Characterization of an Arabidopsis mutant deficient in γ-tocopherol methyltransferase. Plant Mol. Biol. 2003, 52, 1181–1190. [Google Scholar] [CrossRef]
- Kim, Y.J.; Seo, H.Y.; Park, T.I.; Baek, S.H.; Shin, W.C.; Kim, H.S.; Kim, J.G.; Choi, Y.E.; Yun, S.J. Enhanced biosynthesis of α-tocopherol in transgenic soybean by introducing γ-TMT gene. Plant Biotechnol. J. 2005, 7, 203–209. [Google Scholar]
- Kim, M.J.; Baek, S.H.; Yoo, N.H.; Yun, S.J. Transformation of Arabidopsis gamma-tocopherol methyltransferase into lettuce (Lactuca sativa L.). Korean J. Plant Tiss. Cult. 2000, 27, 435–439. [Google Scholar]
- Cho, E.A.; Lee, C.A.; Kim, Y.S.; Baek, S.H.; de Los, R.B.; Yun, S.J. Expression of γ-tocopherol methyltransferase transgene improves tocopherol composition in lettuce (Latuca sativa L.). Mol. Cell 2005, 19, 16–22. [Google Scholar]
- Zhang, G.Y.; Liu, R.R.; Xu, G.; Zhang, P.; Li, Y.; Tang, K.X.; Liang, G.H.; Liu, Q.Q. Increased α-tocotrienol content in seeds of transgenic rice overexpressing Arabidopsis γ-tocopherol methyltransferase. Transgenic Res. 2013, 22, 89–99. [Google Scholar] [CrossRef]
- Ghimire, B.K.; Seong, E.S.; Lee, C.O.; Lim, J.D.; Lee, J.G.; Yoo, J.H.; Chung, I.M.; Kim, N.Y.; Yu, C.Y. Enhancement of α-tocopherol content in transgenic Perilla frutescens containing the γ-TMT gene. Afr. J. Biotechnol. 2011, 10, 2430–2439. [Google Scholar]
- Tavva, V.S.; Kim, Y.; Kagan, I.A.; Dinkins, R.D.; Kim, K.; Collins, G.B. Increased α-tocopherol content in soybean seed overexpressing the Perilla frutescens γ-tocopherol methyltransferase gene. Plant Cell Rep. 2007, 26, 61–70. [Google Scholar] [CrossRef]
- Yusuf, M.A.; Sarin, N.B. Antioxidant value addition in human diets: Genetic transformation of Brassica juncea with γ-TMT gene for increased α-tocopherol content. Transgenic Res. 2007, 16, 109–113. [Google Scholar] [CrossRef]
- Grusak, M.A.; DellaPenna, D. Improving the nutrient composition of plants to enhance human nutrition and health. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 133–161. [Google Scholar] [CrossRef]
- Krieger-Liszkay, A.; Fufezan, C.; Trebst, A. Singlet oxygen production in photosystem II and related protection mechanism. Photosynth. Res. 2008, 98, 551–564. [Google Scholar] [CrossRef]
- Liebler, D.C. The role of metabolism in the antioxidant function of vitamin E. Crit. Rev. Toxicol. 1993, 23, 147–169. [Google Scholar] [CrossRef] [PubMed]
- Kamal-Eldin, A.; Appelqvist, L.A. The chemistry and antioxidant properties of tocopherols and tocotrienols. Lipids 1996, 31, 671–701. [Google Scholar] [CrossRef] [PubMed]
- Kamal-Eldin, A.; Andersson, R. A multivariate study of the correlation between tocopherol content and fatty acid composition in vegetable oils. J. Am. Oil Chem. Soc. 1997, 74, 375–380. [Google Scholar] [CrossRef]
- Goffman, F.D.; Böhme, T. Relationship between fatty acid profile and vitamin E content in maize hybrids (Zea mays L.). J. Agric. Food Chem. 2001, 49, 4990–4994. [Google Scholar] [CrossRef]
- Li, Y.; Hussain, N.; Zhang, L.; Chen, X.; Ali, E.; Jiang, L. Correlations between tocopherol and fatty acid components in germplasm collections of Brassica oilseeds. J. Agri. Food Chem. 2013, 61, 34–40. [Google Scholar] [CrossRef]
- Sattler, S.E.; Gilliland, L.U.; Magallanes-Lundback, M.; Pollard, M.; DellaPenna, D. Vitamin E is essential for seed longevity and for preventing lipid peroxidation during germination. Plant Cell 2004, 16, 1419–1432. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Weiler, E.W.; Alegre, L.; Müller, M.; Düchting, P.; Falk, J. α-Tocopherol may influence cellular signaling by modulating jasmonic acid levels in plants. Planta 2007, 225, 681–691. [Google Scholar] [CrossRef]
- Stahl, E.; Hartmann, M.; Scholten, N.; Zeier, J. A role for tocopherol biosynthesis in Arabidopsis basal immunity to bacterial infection. Plant Physiol. 2019, 181, 1008–1028. [Google Scholar] [CrossRef] [Green Version]
- Abbasi, A.R.; Hajirezaei, M.; Hofius, D.; Sonnewald, U.; Voll, L.M. Specific roles of α- and γ-tocopherol in abiotic stress responses of transgenic tobacco. Plant Physiol. 2007, 143, 1720–1738. [Google Scholar] [CrossRef] [Green Version]
- Yusuf, M.A.; Kumar, D.; Rajwanshi, R.; Strasser, R.J.; Tsimilli-Michael, M.; Govindjee; Sarin, N.B. Overexpression of γ-tocopherol methyl transferase gene in transgenic Brassica juncea plants alleviates abiotic stress: Physiological and chlorophyll a fluorescence measurements. Biochim. Biophys. Acta Bioenerg. 2010, 1797, 1428–1438. [Google Scholar] [CrossRef] [Green Version]
- Cela, J.; Chang, C.; Munné-Bosch, S. Accumulation of γ- rather than α-tocopherol alters ethylene signaling gene expression in the vte4 mutant of Arabidopsis thaliana. Plant Cell Physiol. 2011, 52, 1389–1400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellouzi, H.; Hamed, K.B.; Cela, J.; Müller, M.; Abdelly, C.; Munné-Bosch, S. Increased sensitivity to salt stress in tocopherol-deficient Arabidopsis mutants growing in a hydroponic system. Plant Signal Behav. 2013, 8, e23136. [Google Scholar] [CrossRef] [Green Version]
- Jin, S.; Daniell, H. Expression of γ-tocopherol methyltransferase in chloroplasts results in massive proliferation of the inner envelope membrane and decreases susceptibility to salt and metal-induced oxidative stresses by reducing reactive oxygen species. Plant Biotechnol. J. 2014, 12, 1274–1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Qiu, D.; Xu, L.; Cui, M.; Gao, H.; Pang, Y.; Qin, Y.; Wang, X. Overexpression of alfalfa γ-tocopherol methyltransferase (γ-TMT) gene increases salt susceptibility of transgenic Arabidopsis in seed germination. Environ. Exp. Bot. 2020, 180, 104264. [Google Scholar] [CrossRef]
- Maeda, H.; Song, W.; Sage, T.L.; DellaPenna, D. Tocopherols play a crucial role in low-temperature adaptation and phloem loading in Arabidopsis. Plant Cell 2006, 18, 2710–2732. [Google Scholar] [CrossRef] [Green Version]
- Maeda, H.; Sage, T.L.; Isaac, G.; Welti, R.; DellaPenna, D. Tocopherols modulate extraplastidic polyunsaturated fatty acid metabolism in Arabidopsis at low temperature. Plant Cell 2008, 20, 452–470. [Google Scholar] [CrossRef]
- Ma, J.; Qiu, D.; Gao, H.; Wen, H.; Wu, Y.; Pang, Y.; Wang, X.; Qin, Y. Over-expression of a γ-tocopherol methyltransferase gene in vitamin E pathway confers PEG-simulated drought tolerance in alfalfa. BMC Plant Biol. 2020, 20, 226. [Google Scholar] [CrossRef]
- Collin, V.C.; Eymery, F.; Genty, B.; Rey, P.; Havaux, M. Vitamin E is essential for the tolerance of Arabidopsis thaliana to metal-induced oxidative stress. Plant Cell Environ. 2008, 31, 244–257. [Google Scholar] [CrossRef]
- Zhu, Q.; Zhang, J.; Yu, H.; Li, L.; Chen, X.; Jiang, M.; Tan, M. Maize Cd-tolerant ZmVTE4 encoding γ-tocopherol-methyl-transferase alleviated Cd-toxicity through its product α-tocopherol. Environ. Exp. Bot. 2019, 158, 171–179. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef]
- Schranz, M.E.; Lysak, M.A.; Mitchell-olds, T. The ABC’s of comparative genomics in the Brassicaceae: Building blocks of crucifer genomes. Trends Plant Sci. 2006, 11, 535–542. [Google Scholar] [CrossRef]
- Grillo, M.A.; Colombatto, S. S-adenosylmethionine and its products. Amino Acids 2008, 34, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Goffman, F.D.; Velasco, L.; Becker, H.C. Tocopherols accumulation in developing seeds and pods of rapeseed (Brassica napus L.). Lipid/Fett 1999, 101, 400–403. [Google Scholar] [CrossRef]
- Endrigkeit, J.; Wang, X.; Cai, D.; Zhang, C.; Long, Y.; Meng, J.; Jung, C. Genetic mapping, cloning, and functional characterization of the BnaX.VTE4 gene encoding a γ-tocopherol methyltransferase from oilseed rape. Theor. Appl. Genet. 2009, 119, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Okuley, J.; Lightner, J.; Feldmann, K.; Yadav, N.; Lark, E.; Browse, J. Arabidopsis FAD2 gene encodes the enzyme that is essential for polyunsaturated lipid synthesis. Plant Cell 1994, 6, 147–158. [Google Scholar]
- Lee, K.R.; Lee, Y.; Kim, E.H.; Lee, S.B.; Roh, K.H.; Kim, J.B.; Kang, H.C.; Kim, H.U. Functional identification of oleate 12-desaturase and ω-3 fatty acid desaturase genes from Perilla frutescens var. Frutescens. Plant Cell Rep. 2016, 35, 2523–2537. [Google Scholar] [CrossRef]
- Shah, S.; Xin, Z.G.; Browse, J. Overexpression of the FAD3 desaturase gene in a mutant of Arabidopsis. Plant Physiol. 1997, 114, 1533–1539. [Google Scholar] [CrossRef] [Green Version]
- Peng, Z.; Ruan, J.; Tian, H.; Shan, L.; Meng, J.; Guo, F.; Zhang, Z.; Ding, H.; Wan, S.; Li, X. The family of peanut fatty acid desaturase genes and a functional analysis of four ω-3 AhFAD3 members. Plant Mol. Biol. Rep. 2020, 38, 209–221. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef]
- Miller, G.; Shulaev, V.; Mittler, R. Reactive oxygen signaling and abiotic stress. Physiol. Plant. 2008, 133, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Ohlrogge, J.B.; Jaworski, J.G. Regulation of fatty acid synthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 109–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, J.K.; Choi, H.W.; Hwang, I.S.; Kim, D.S.; Kim, N.H.; Choi, D.S.; Kim, Y.J.; Hwang, B.K. Function of a novel GDSL-type pepper lipase gene, CaGLIP1, in disease susceptibility and abiotic stress tolerance. Planta 2008, 227, 539–558. [Google Scholar] [CrossRef] [PubMed]
- Mu, J.; Tan, H.; Zheng, Q.; Fu, F.; Liang, Y.; Zhang, J.; Yang, X.; Wang, T.; Chong, K.; Wang, X.J.; et al. LEAFY COTYLEDON1 is a key regulator of fatty acid biosynthesis in Arabidopsis. Plant Physiol. 2008, 148, 1042–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, M.J. Enzymes involved in jasmonic acid biosynthesis. Physiol. Plant. 1997, 100, 653–663. [Google Scholar] [CrossRef]
- Wang, J.; Song, L.; Gong, X.; Xu, J.; Li, M. Functions of jasmonic acid in plant regulation and response to abiotic stress. Int. J. Mol. Sci. 2020, 21, 1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortés, A.J.; López-Hernández, F. Harnessing crop wild diversity for climate change adaptation. Genes 2021, 12, 783. [Google Scholar] [CrossRef] [PubMed]
- Cortés, A.J.; López-Hernández, F.; Blair, M.W. Genome-environment associations, an innovative tool for studying heritable evolutionary adaptation in orphan crops and wild relatives. Front. Genet. 2022, 13, 910386. [Google Scholar] [CrossRef]
- Burbano-Erazo, E.; León-Pacheco, R.I.; Cordero-Cordero, C.C.; López-Hernández, F.; Cortés, A.J.; Tofiño-Rivera, A.P. Multi-environment yield components in advanced common bean (Phaseolus vulgaris L.) × Tepary bean (P. acutifolius A. Gray) interspecific lines for heat and drought tolerance. Agronomy 2021, 11, 1978. [Google Scholar] [CrossRef]
- Liu, C.; Chen, H.; Er, H.L.; Soo, H.M.; Kumar, P.P.; Han, J.; Liou, Y.C.; Yu, H. Direct interaction of AGL24 and SOC1 integrates flowering signals in Arabidopsis. Development 2008, 135, 1481–1491. [Google Scholar] [CrossRef] [Green Version]
- Cardoza, V.; Stewart, C.N. Increased Agrobacterium-mediated transformation and rooting efficiencies in canola (Brassica napus L.) from hypocotyl segment explants. Plant Cell Rep. 2003, 21, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Poirier, Y.; Ventre, G.; Caldelari, D. Increased flow of fatty acids toward β-oxidation in developing seeds of Arabidopsis deficient in diacylglycerol acyltransferase activity or synthesizing medium-chain-length fatty acids. Plant Physiol. 1999, 121, 1359–1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Wang, Z.; Zhu, Y.; Li, Z.; Hussain, N.; Xuan, L.; Guo, W.; Zhang, G.; Jiang, L. The effect of transparent TESTA2 on seed fatty acid biosynthesis and tolerance to environmental stresses during young seedling establishment in Arabidopsis. Plant Physiol. 2012, 160, 1023–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Wang, J.; Li, S.; Kakan, X.; Zhou, Y.; Miao, Y.; Wang, F.; Qin, H.; Huang, R. Ascorbic acid integrates the antagonistic modulation of ethylene and abscisic acid in the accumulation of reactive oxygen species. Plant Physiol. 2019, 179, 1861–1875. [Google Scholar] [CrossRef] [Green Version]
- Kakan, X.; Yu, Y.; Li, S.; Li, X.; Huang, R.; Wang, J. Ascorbic acid modulation by ABI4 transcriptional repression of VTC2 in the salt tolerance of Arabidopsis. BMC Plant Biol. 2021, 21, 112. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Dai, Y.; Zheng, C.; Yang, Y.; Chen, W.; Wang, Q.; Chandrasekaran, U.; Du, J.; Liu, W.; Shu, K. The ABI4-RbohD/VTC2 regulatory module promotes Reactive Oxygen Species (ROS) accumulation to decrease seed germination under salinity stress. New Phytol. 2021, 229, 950–962. [Google Scholar] [CrossRef] [PubMed]
- Assmann, S.M. G protein signaling in the regulation of Arabidopsis seed germination. Sci. STKE 2005, 308, cm11. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST©) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef]
- Küpper, H.; Seib, L.O.; Sivaguru, M.; Hoekenga, O.A.; Kochian, L.V. A method for cellular localization of gene expression via quantitative in situ hybridization in plants. Plant J. 2007, 50, 159–175. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Y.; Li, D.; Liu, T.; Liao, M.; Li, Y.; Zhang, W.; Liu, Z.; Chen, M. Effect of Overexpression of γ-Tocopherol Methyltransferase on α-Tocopherol and Fatty Acid Accumulation and Tolerance to Salt Stress during Seed Germination in Brassica napus L. Int. J. Mol. Sci. 2022, 23, 15933. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232415933
Guo Y, Li D, Liu T, Liao M, Li Y, Zhang W, Liu Z, Chen M. Effect of Overexpression of γ-Tocopherol Methyltransferase on α-Tocopherol and Fatty Acid Accumulation and Tolerance to Salt Stress during Seed Germination in Brassica napus L. International Journal of Molecular Sciences. 2022; 23(24):15933. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232415933
Chicago/Turabian StyleGuo, Yuan, Dong Li, Tiantian Liu, Meifang Liao, Yuxin Li, Weitang Zhang, Zijin Liu, and Mingxun Chen. 2022. "Effect of Overexpression of γ-Tocopherol Methyltransferase on α-Tocopherol and Fatty Acid Accumulation and Tolerance to Salt Stress during Seed Germination in Brassica napus L." International Journal of Molecular Sciences 23, no. 24: 15933. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232415933