
Drosophila melanogaster Uncoupling Protein-4A (UCP4A) Catalyzes a Unidirectional Transport of Aspartate

 , , , , , and
, , , , , and 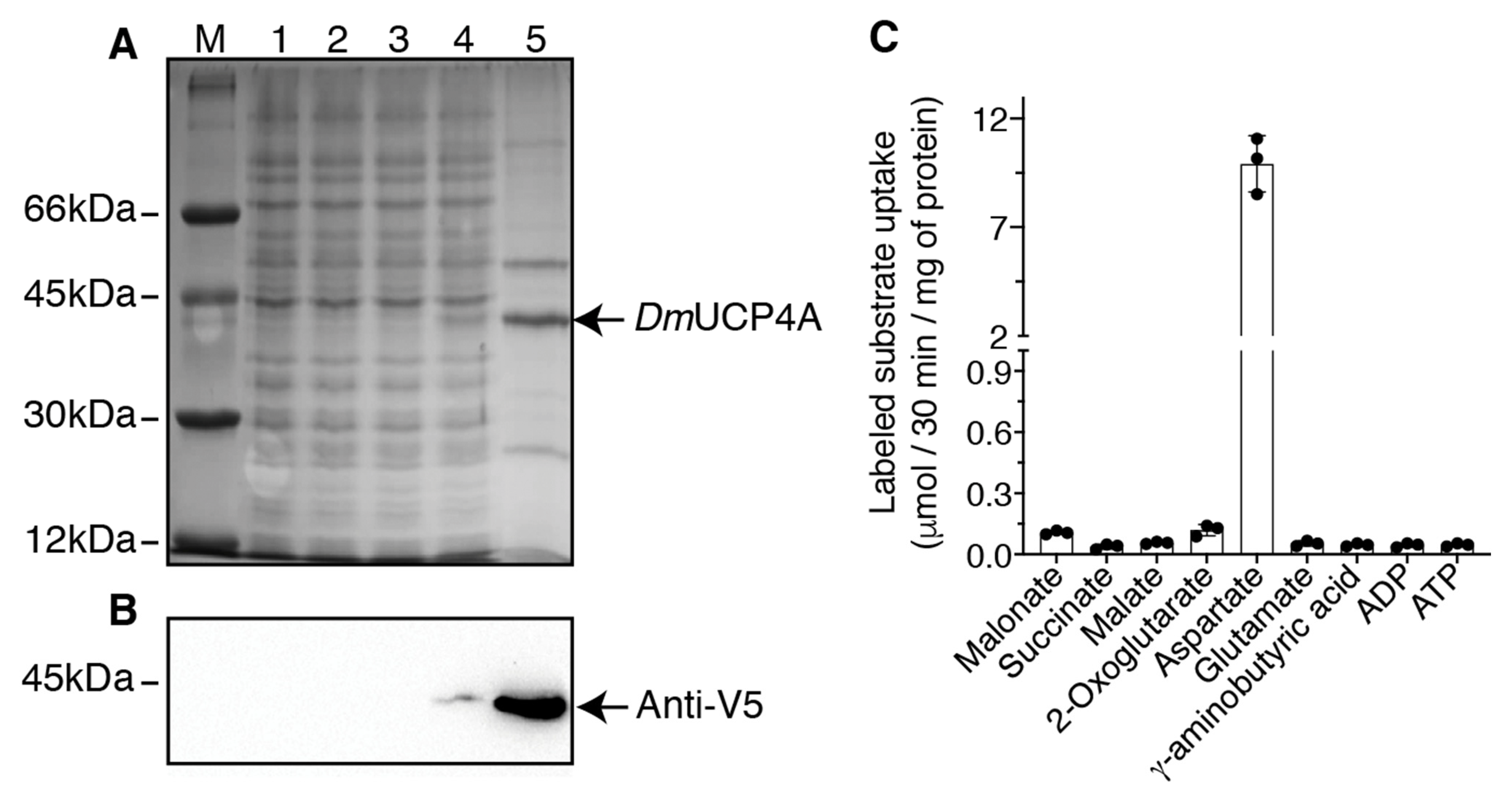{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
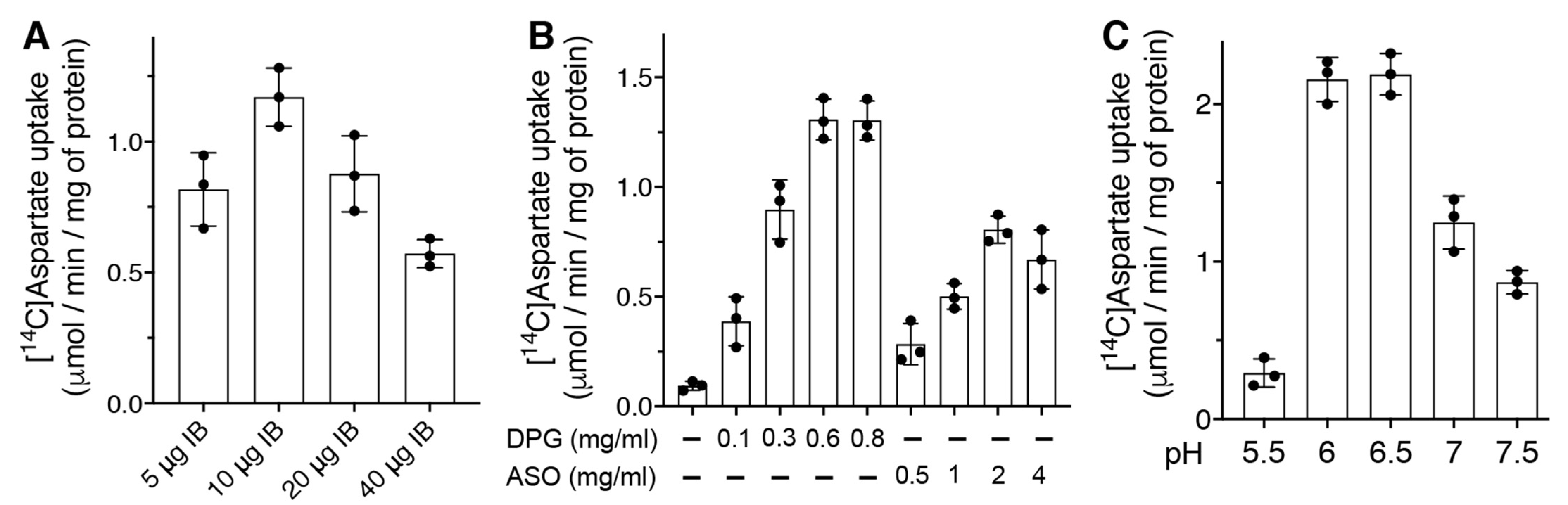
2.1. Bacterial Expression of DmUCP4A and Optimization of the Transport Assay into Liposomes
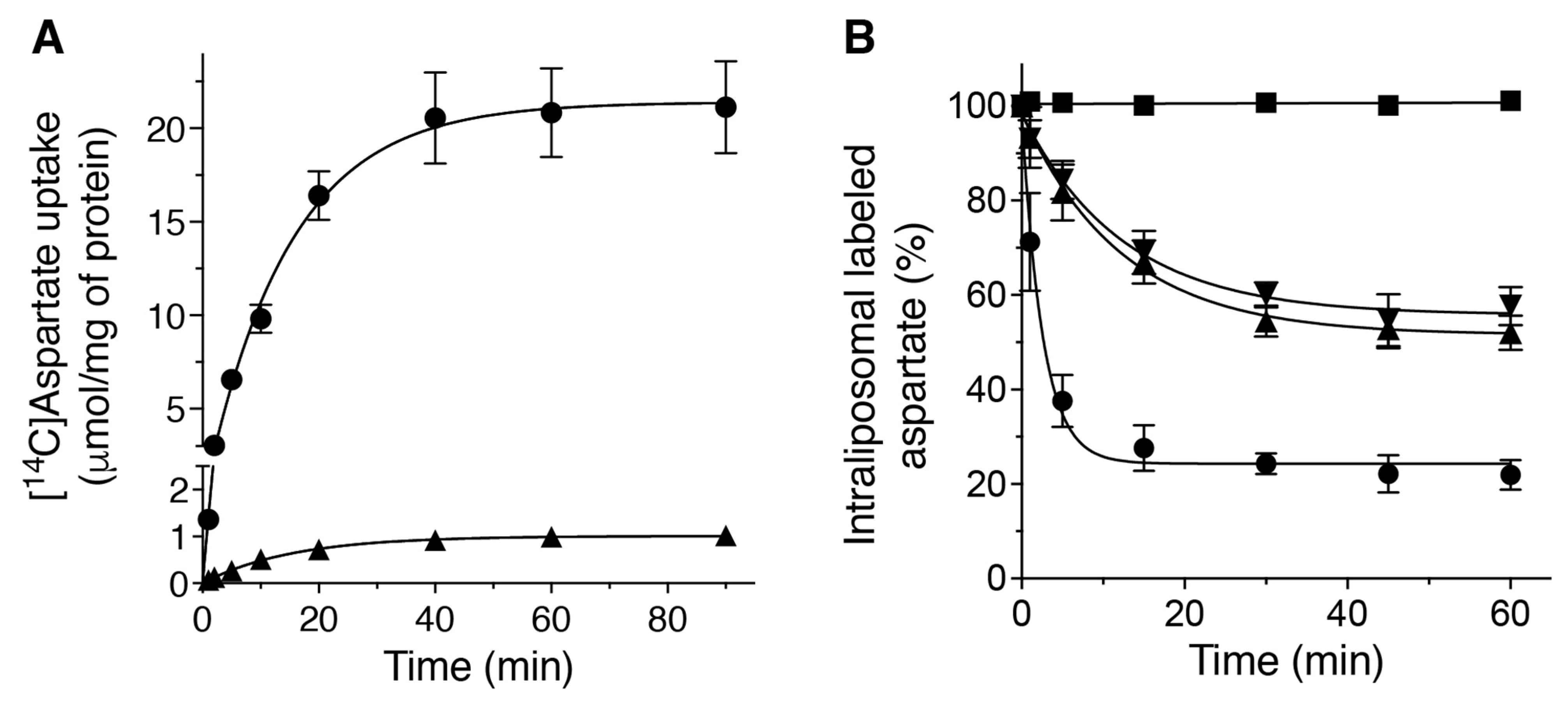
2.2. Kinetic Properties of the Recombinant DmUCP4A Protein
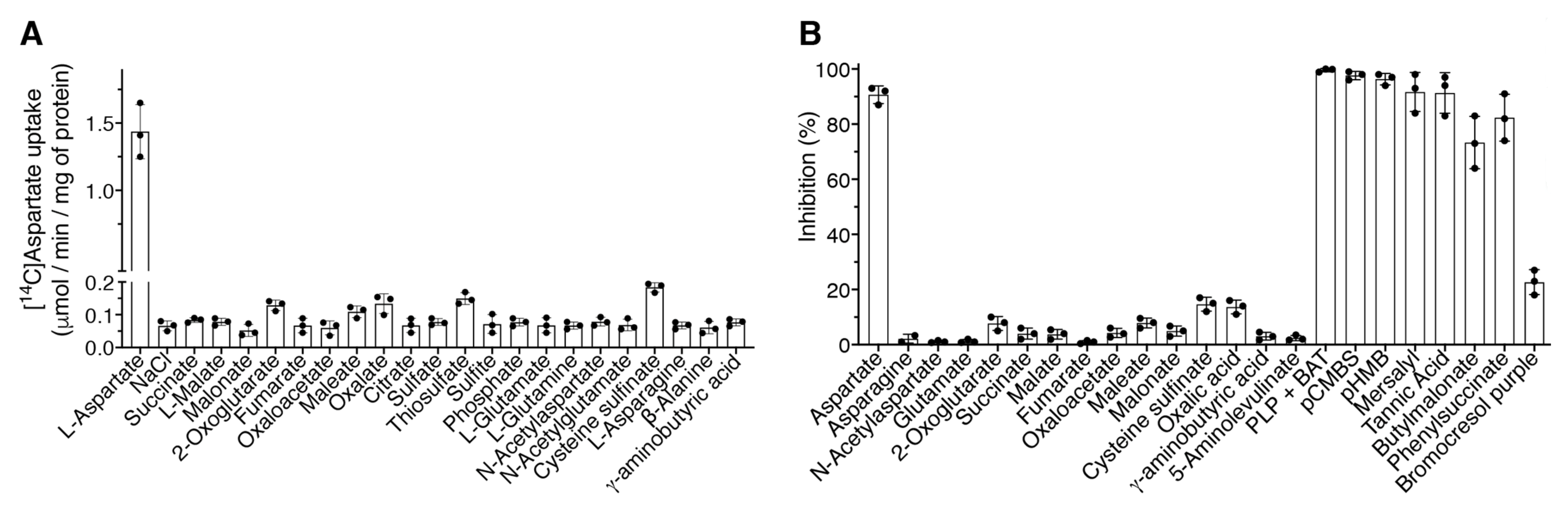
2.3. Substrate Specificity and Inhibitor Sensitivity of the Recombinant DmUCP4A Protein
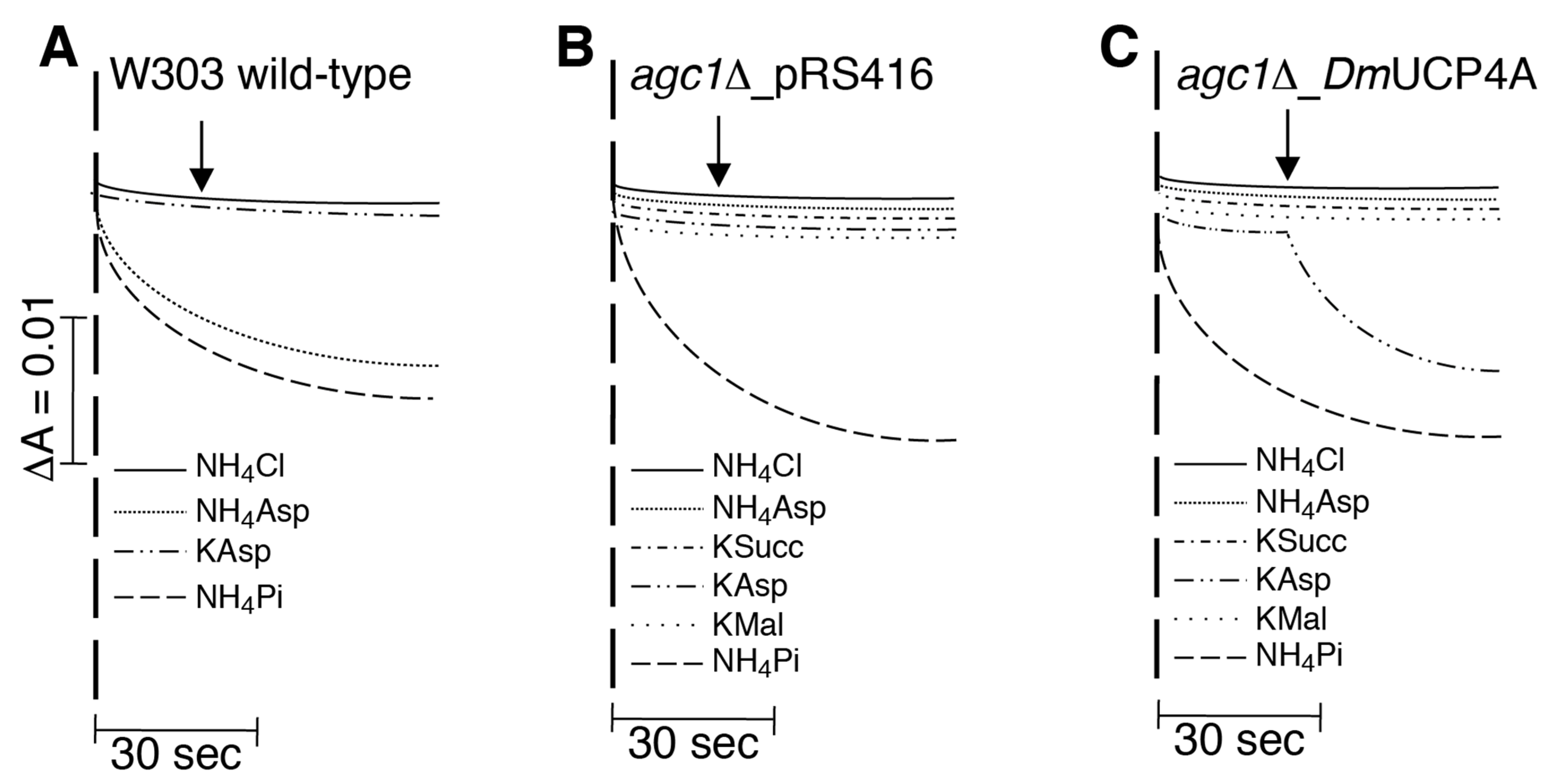
2.4. DmUCP4A Catalyzes a Unidirectional Transport of Aspartate in Saccharomyces Cerevisiae Mitochondria
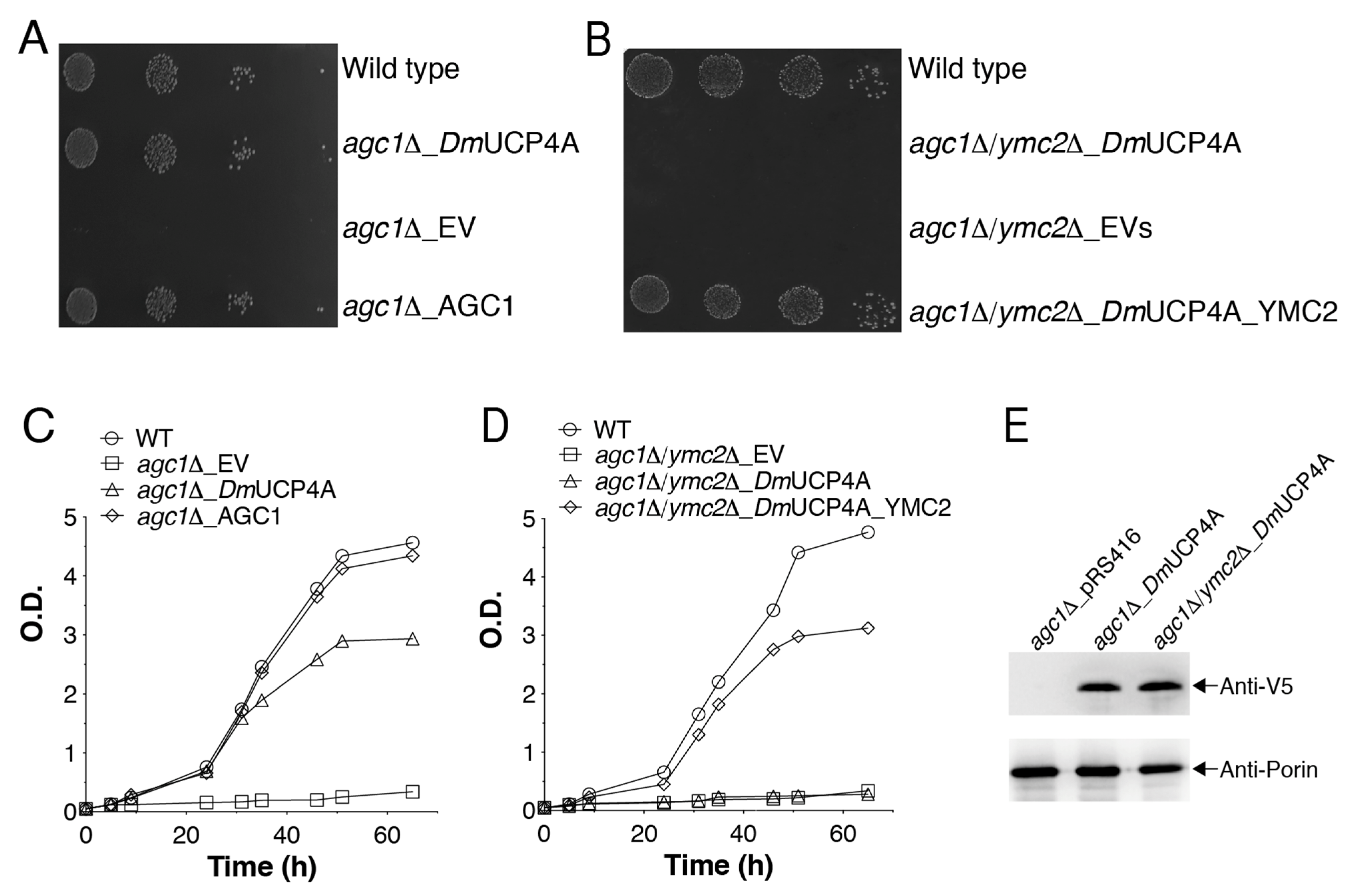
2.5. DmUCP4A Catalyzes a Unidirectional Transport of Aspartate in a Yeast Cell Model
3. Discussion
4. Materials and Methods
4.1. Construction of Plasmids for D. melanogaster DmUCP4A Expression in E. coli and S. cerevisiae
4.2. Bacterial Expression and Purification of DmUCP4A
4.3. Reconstitution into Liposomes and Transport Assays
4.4. Yeast Strains, Growth Conditions, and Functional Complementation
4.5. Isolation and Swelling of Yeast Mitochondria
4.6. Other Methods
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ASO | asolectin |
| AGC1p | yeast mitochondrial aspartate/glutamate transporter |
| BAT | bathophenanthroline |
| DPG | cardiolipin |
| DTE | dithioerythritol |
| YMC2p | yeast mitochondrial glutamate/H+ symporter |
| IB | inclusion bodies |
| MAS | malate-aspartate shuttle |
| MCF | mitochondrial carrier family |
| PCMBS | p-chloromercuribenzenesulfonate |
| pHMB | p-hydroxymercuribenzoate |
| PD | Parkinson’s disease |
| PCR | polymerase chain reaction |
| PLP | pyridoxal-5′-phosphate |
| TAN | tannic acid |
| UCPs | uncoupling proteins |
| PIPES | piperazine-N,N′-bis(2-ethanesulfonic acid) |
| EDTA | ethylenediaminetetraacetic acid |
References
- Curcio, R.; Lunetti, P.; Zara, V.; Ferramosca, A.; Marra, F.; Fiermonte, G.; Cappello, A.R.; De Leonardis, F.; Capobianco, L.; Dolce, V. Drosophila melanogaster Mitochondrial Carriers: Similarities and Differences with the Human Carriers. Int. J. Mol. Sci. 2020, 21, 6052. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, F. The mitochondrial transporter family SLC25: Identification, properties and physiopathology. Mol. Asp. Med. 2013, 34, 465–484. [Google Scholar] [CrossRef]
- Bertholet, A.M.; Chouchani, E.T.; Kazak, L.; Angelin, A.; Fedorenko, A.; Long, J.Z.; Vidoni, S.; Garrity, R.; Cho, J.; Terada, N.; et al. H(+) transport is an integral function of the mitochondrial ADP/ATP carrier. Nature 2019, 571, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Vozza, A.; Parisi, G.; De Leonardis, F.; Lasorsa, F.M.; Castegna, A.; Amorese, D.; Marmo, R.; Calcagnile, V.M.; Palmieri, L.; Ricquier, D.; et al. UCP2 transports C4 metabolites out of mitochondria, regulating glucose and glutamine oxidation. Proc. Natl. Acad. Sci. USA 2014, 111, 960–965. [Google Scholar] [CrossRef] [Green Version]
- Gorgoglione, R.; Porcelli, V.; Santoro, A.; Daddabbo, L.; Vozza, A.; Monne, M.; Di Noia, M.A.; Palmieri, L.; Fiermonte, G.; Palmieri, F. The human uncoupling proteins 5 and 6 (UCP5/SLC25A14 and UCP6/SLC25A30) transport sulfur oxyanions, phosphate and dicarboxylates. Biochim. Biophys. Acta Bioenerg. 2019, 1860, 724–733. [Google Scholar] [CrossRef]
- Monne, M.; Daddabbo, L.; Gagneul, D.; Obata, T.; Hielscher, B.; Palmieri, L.; Miniero, D.V.; Fernie, A.R.; Weber, A.P.M.; Palmieri, F. Uncoupling proteins 1 and 2 (UCP1 and UCP2) from Arabidopsis thaliana are mitochondrial transporters of aspartate, glutamate, and dicarboxylates. J. Biol. Chem. 2018, 293, 4213–4227. [Google Scholar] [CrossRef] [Green Version]
- Hanak, P.; Jezek, P. Mitochondrial uncoupling proteins and phylogenesis--UCP4 as the ancestral uncoupling protein. FEBS Lett 2001, 495, 137–141. [Google Scholar] [CrossRef]
- Slocinska, M.; Barylski, J.; Jarmuszkiewicz, W. Uncoupling proteins of invertebrates: A review. IUBMB Life 2016, 68, 691–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jezek, P.; Urbankova, E. Specific sequence of motifs of mitochondrial uncoupling proteins. IUBMB Life 2000, 49, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Liu, J.; Zhuang, N.; Wang, T. UCP4A protects against mitochondrial dysfunction and degeneration in pink1/parkin models of Parkinson’s disease. FASEB J. 2014, 28, 5111–5121. [Google Scholar] [CrossRef]
- Imai, Y.; Inoshita, T.; Meng, H.; Shiba-Fukushima, K.; Hara, K.Y.; Sawamura, N.; Hattori, N. Light-driven activation of mitochondrial proton-motive force improves motor behaviors in a Drosophila model of Parkinson’s disease. Commun. Biol. 2019, 2, 424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da-Re, C.; De Pitta, C.; Zordan, M.A.; Teza, G.; Nestola, F.; Zeviani, M.; Costa, R.; Bernardi, P. UCP4C mediates uncoupled respiration in larvae of Drosophila melanogaster. EMBO Rep. 2014, 15, 586–591. [Google Scholar] [CrossRef] [Green Version]
- Ulgherait, M.; Chen, A.; McAllister, S.F.; Kim, H.X.; Delventhal, R.; Wayne, C.R.; Garcia, C.J.; Recinos, Y.; Oliva, M.; Canman, J.C.; et al. Circadian regulation of mitochondrial uncoupling and lifespan. Nat. Commun. 2020, 11, 1927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fridell, Y.W.; Sanchez-Blanco, A.; Silvia, B.A.; Helfand, S.L. Functional characterization of a Drosophila mitochondrial uncoupling protein. J. Bioenerg. Biomembr. 2004, 36, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Blanco, A.; Fridell, Y.W.; Helfand, S.L. Involvement of Drosophila uncoupling protein 5 in metabolism and aging. Genetics 2006, 172, 1699–1710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeiffer, M.; Kayzer, E.-B.; Yang, X.; Abramson, E.; Kenaston, M.A.; Lago, C.U.; Lo, H.-H.; Sedensky, M.M.; Lunceford, A.; Clarke, C.F.; et al. Caenorhabditis elegans UCP4 protein controls complex II-mediated oxidative phosphorylation through succinate transport. J. Biol. Chem. 2011, 286, 37712–37720. [Google Scholar] [CrossRef] [Green Version]
- Ho, P.W.-L.; Ho, J.W.-M.; Liu, H.-F.; Chan, K.-H.; Ramsden, D.B.; Ho, S.-L. Uncoupling Protein-4 (UCP4) Increases ATP Supply by Interacting with Mitochondrial Complex II in Neuroblastoma Cells. PLoS ONE 2012, 7, e32810. [Google Scholar] [CrossRef] [Green Version]
- Fiermonte, G.; Walker, J.E.; Palmieri, F. Abundant bacterial expression and reconstitution of an intrinsic membrane-transport protein from bovine mitochondria. Biochem. J. 1993, 294 Pt 1, 293–299. [Google Scholar] [CrossRef]
- Fiermonte, G.; Dolce, V.; Palmieri, F. Expression in Escherichia coli, functional characterization, and tissue distribution of isoforms A and B of the phosphate carrier from bovine mitochondria. J. Biol. Chem. 1998, 273, 22782–22787. [Google Scholar] [CrossRef] [Green Version]
- Vozza, A.; De Leonardis, F.; Paradies, E.; De Grassi, A.; Pierri, C.L.; Parisi, G.; Marobbio, C.M.; Lasorsa, F.M.; Muto, L.; Capobianco, L.; et al. Biochemical characterization of a new mitochondrial transporter of dephosphocoenzyme A in Drosophila melanogaster. Biochim. Biophys. Acta 2017, 1858, 137–146. [Google Scholar] [CrossRef]
- Dolce, V.; Cappello, A.R.; Capobianco, L. Mitochondrial tricarboxylate and dicarboxylate-Tricarboxylate carriers: From animals to plants. IUBMB Life 2014, 66, 462–471. [Google Scholar] [CrossRef]
- Curcio, R.; Muto, L.; Pierri, C.L.; Montalto, A.; Lauria, G.; Onofrio, A.; Fiorillo, M.; Fiermonte, G.; Lunetti, P.; Vozza, A.; et al. New insights about the structural rearrangements required for substrate translocation in the bovine mitochondrial oxoglutarate carrier. Biochim. Biophys. Acta 2016, 1864, 1473–1480. [Google Scholar] [CrossRef] [PubMed]
- Picault, N.; Palmieri, L.; Pisano, I.; Hodges, M.; Palmieri, F. Identification of a novel transporter for dicarboxylates and tricarboxylates in plant mitochondria. Bacterial expression, reconstitution, functional characterization, and tissue distribution. J. Biol. Chem. 2002, 277, 24204–24211. [Google Scholar] [CrossRef] [Green Version]
- Fiermonte, G.; Palmieri, L.; Todisco, S.; Agrimi, G.; Palmieri, F.; Walker, J.E. Identification of the mitochondrial glutamate transporter. Bacterial expression, reconstitution, functional characterization, and tissue distribution of two human isoforms. J. Biol. Chem. 2002, 277, 19289–19294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmieri, F.; Indiveri, C.; Bisaccia, F.; Iacobazzi, V. [25] Mitochondrial metabolite carrier proteins: Purification, reconstitution, and transport studies. Meth. Enzymol. 1995, 260, 349–369. [Google Scholar] [CrossRef]
- Lunetti, P.; Marsano, R.M.; Curcio, R.; Dolce, V.; Fiermonte, G.; Cappello, A.R.; Marra, F.; Moschetti, R.; Li, Y.; Aiello, D.; et al. The mitochondrial aspartate/glutamate carrier (AGC or Aralar1) isoforms in D. melanogaster: Biochemical characterization, gene structure, and evolutionary analysis. Biochim. Biophys. Acta Gen. Subj. 2021, 1865, 129854. [Google Scholar] [CrossRef]
- Fiermonte, G.; Palmieri, L.; Dolce, V.; Lasorsa, F.M.; Palmieri, F.; Runswick, M.J.; Walker, J.E. The sequence, bacterial expression, and functional reconstitution of the rat mitochondrial dicarboxylate transporter cloned via distant homologs in yeast and Caenorhabditis elegans. J. Biol. Chem. 1998, 273, 24754–24759. [Google Scholar] [CrossRef] [Green Version]
- Iacopetta, D.; Madeo, M.; Tasco, G.; Carrisi, C.; Curcio, R.; Martello, E.; Casadio, R.; Capobianco, L.; Dolce, V. A novel subfamily of mitochondrial dicarboxylate carriers from Drosophila melanogaster: Biochemical and computational studies. Biochim. Biophys. Acta 2011, 1807, 251–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavero, S.; Vozza, A.; del Arco, A.; Palmieri, L.; Villa, A.; Blanco, E.; Runswick, M.J.; Walker, J.E.; Cerdan, S.; Palmieri, F.; et al. Identification and metabolic role of the mitochondrial aspartate-glutamate transporter in Saccharomyces cerevisiae. Mol. Microbiol. 2003, 50, 1257–1269. [Google Scholar] [CrossRef]
- Palmieri, L.; Vozza, A.; Agrimi, G.; De Marco, V.; Runswick, M.J.; Palmieri, F.; Walker, J.E. Identification of the yeast mitochondrial transporter for oxaloacetate and sulfate. J. Biol. Chem. 1999, 274, 22184–22190. [Google Scholar] [CrossRef] [Green Version]
- Porcelli, V.; Vozza, A.; Calcagnile, V.; Gorgoglione, R.; Arrigoni, R.; Fontanesi, F.; Marobbio, C.M.T.; Castegna, A.; Palmieri, F.; Palmieri, L. Molecular identification and functional characterization of a novel glutamate transporter in yeast and plant mitochondria. Biochim. Biophys. Acta Bioenerg. 2018, 1859, 1249–1258. [Google Scholar] [CrossRef]
- Fedorenko, A.; Fedorenko, A.; Lishko, P.V.; Lishko, P.V.; Kirichok, Y.; Kirichok, Y. Mechanism of fatty-acid-dependent UCP1 uncoupling in brown fat mitochondria. Cell 2012, 151, 400–413. [Google Scholar] [CrossRef] [Green Version]
- Raho, S.; Capobianco, L.; Malivindi, R.; Vozza, A.; Piazzolla, C.; De Leonardis, F.; Gorgoglione, R.; Scarcia, P.; Pezzuto, F.; Agrimi, G.; et al. KRAS-regulated glutamine metabolism requires UCP2-mediated aspartate transport to support pancreatic cancer growth. Nat. Metab. 2020, 2, 1373–1381. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, F. The mitochondrial transporter family (SLC25): Physiological and pathological implications. Pflug. Arch. 2004, 447, 689–709. [Google Scholar] [CrossRef] [PubMed]
- Riley, C.L.; Dao, C.; Kenaston, M.A.; Muto, L.; Kohno, S.; Nowinski, S.M.; Solmonson, A.D.; Pfeiffer, M.; Sack, M.N.; Lu, Z.; et al. The complementary and divergent roles of uncoupling proteins 1 and 3 in thermoregulation. J. Physiol. 2016, 594, 7455–7464. [Google Scholar] [CrossRef]
- Iacopetta, D.; Carrisi, C.; De Filippis, G.; Calcagnile, V.M.; Cappello, A.R.; Chimento, A.; Curcio, R.; Santoro, A.; Vozza, A.; Dolce, V.; et al. The biochemical properties of the mitochondrial thiamine pyrophosphate carrier from Drosophila melanogaster. FEBS J. 2010, 277, 1172–1181. [Google Scholar] [CrossRef] [PubMed]
- Gorgoglione, R.; Impedovo, V.; Riley, C.L.; Fratantonio, D.; Tiziani, S.; Palmieri, L.; Dolce, V.; Fiermonte, G. Glutamine-Derived Aspartate Biosynthesis in Cancer Cells: Role of Mitochondrial Transporters and New Therapeutic Perspectives. Cancers 2022, 14, 245. [Google Scholar] [CrossRef]
- Son, J.; Lyssiotis, C.A.; Ying, H.; Wang, X.; Hua, S.; Ligorio, M.; Perera, R.M.; Ferrone, C.R.; Mullarky, E.; Shyh-Chang, N.; et al. Glutamine supports pancreatic cancer growth through a KRAS-regulated metabolic pathway. Nature 2013, 496, 101–105. [Google Scholar] [CrossRef]
- Piskur, J.; Kolbak, D.; Sondergaard, L.; Pedersen, M.B. The dominant mutation Suppressor of black indicates that de novo pyrimidine biosynthesis is involved in the Drosophila tan pigmentation pathway. Mol. Gen. Genet. 1993, 241, 335–340. [Google Scholar] [CrossRef]
- Davidson, J.N.; Kern, C.B. Revision in sequence of CAD aspartate transcarbamylase domain of Drosophila. J. Mol. Biol. 1994, 243, 364–366. [Google Scholar] [CrossRef]
- Borycz, J.; Borycz, J.A.; Edwards, T.N.; Boulianne, G.L.; Meinertzhagen, I.A. The metabolism of histamine in the Drosophila optic lobe involves an ommatidial pathway: Beta-alanine recycles through the retina. J. Exp. Biol. 2012, 215, 1399–1411. [Google Scholar] [CrossRef] [Green Version]
- Izore, T.; Tailhades, J.; Hansen, M.H.; Kaczmarski, J.A.; Jackson, C.J.; Cryle, M.J. Drosophila melanogaster nonribosomal peptide synthetase Ebony encodes an atypical condensation domain. Proc. Natl. Acad. Sci. USA 2019, 116, 2913–2918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, C.A.; Krantz, D.E. Drosophila melanogaster as a genetic model system to study neurotransmitter transporters. Neurochem. Int. 2014, 73, 71–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suh, J.; Jackson, F.R. Drosophila ebony activity is required in glia for the circadian regulation of locomotor activity. Neuron 2007, 55, 435–447. [Google Scholar] [CrossRef] [Green Version]
- Wright, T.R. The genetics of biogenic amine metabolism, sclerotization, and melanization in Drosophila melanogaster. Adv. Genet. 1987, 24, 127–222. [Google Scholar] [CrossRef]
- Richardson, G.; Ding, H.; Rocheleau, T.; Mayhew, G.; Reddy, E.; Han, Q.; Christensen, B.M.; Li, J. An examination of aspartate decarboxylase and glutamate decarboxylase activity in mosquitoes. Mol. Biol. Rep. 2010, 37, 3199–3205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, F.R.; Haydon, P.G. Glial cell regulation of neurotransmission and behavior in Drosophila. Neuron Glia Biol. 2008, 4, 11–17. [Google Scholar] [CrossRef]
- Gavin, B.A.; Arruda, S.E.; Dolph, P.J. The role of carcinine in signaling at the Drosophila photoreceptor synapse. PLoS Genet. 2007, 3, e206. [Google Scholar] [CrossRef]
- Halassa, M.M.; Fellin, T.; Haydon, P.G. The tripartite synapse: Roles for gliotransmission in health and disease. Trends Mol. Med. 2007, 13, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Newby, L.M.; Jackson, F.R. Drosophila ebony mutants have altered circadian activity rhythms but normal eclosion rhythms. J. Neurogenet 1991, 7, 85–101. [Google Scholar] [CrossRef]
- Profilo, E.; Pena-Altamira, L.E.; Corricelli, M.; Castegna, A.; Danese, A.; Agrimi, G.; Petralla, S.; Giannuzzi, G.; Porcelli, V.; Sbano, L.; et al. Down-regulation of the mitochondrial aspartate-glutamate carrier isoform 1 AGC1 inhibits proliferation and N-acetylaspartate synthesis in Neuro2A cells. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 1422–1435. [Google Scholar] [CrossRef] [PubMed]
- Wibom, R.; Lasorsa, F.M.; Tohonen, V.; Barbaro, M.; Sterky, F.H.; Kucinski, T.; Naess, K.; Jonsson, M.; Pierri, C.L.; Palmieri, F.; et al. AGC1 deficiency associated with global cerebral hypomyelination. N. Engl. J. Med. 2009, 361, 489–495. [Google Scholar] [CrossRef] [Green Version]
- Levin, B.E.; Katzen, H.L.; Maudsley, A.; Post, J.; Myerson, C.; Govind, V.; Nahab, F.; Scanlon, B.; Mittel, A. Whole-brain proton MR spectroscopic imaging in Parkinson’s disease. J. Neuroimaging 2014, 24, 39–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klietz, M.; Bronzlik, P.; Nosel, P.; Wegner, F.; Dressler, D.W.; Dadak, M.; Maudsley, A.A.; Sheriff, S.; Lanfermann, H.; Ding, X.Q. Altered Neurometabolic Profile in Early Parkinson’s Disease: A Study With Short Echo-Time Whole Brain MR Spectroscopic Imaging. Front. Neurol. 2019, 10, 777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hattingen, E.; Magerkurth, J.; Pilatus, U.; Mozer, A.; Seifried, C.; Steinmetz, H.; Zanella, F.; Hilker, R. Phosphorus and proton magnetic resonance spectroscopy demonstrates mitochondrial dysfunction in early and advanced Parkinson’s disease. Brain 2009, 132, 3285–3297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henchcliffe, C.; Shungu, D.C.; Mao, X.; Huang, C.; Nirenberg, M.J.; Jenkins, B.G.; Beal, M.F. Multinuclear magnetic resonance spectroscopy for in vivo assessment of mitochondrial dysfunction in Parkinson’s disease. Ann. N. Y. Acad. Sci. 2008, 1147, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Mazuel, L.; Chassain, C.; Jean, B.; Pereira, B.; Cladiere, A.; Speziale, C.; Durif, F. Proton MR Spectroscopy for Diagnosis and Evaluation of Treatment Efficacy in Parkinson Disease. Radiology 2016, 278, 505–513. [Google Scholar] [CrossRef]
- Griffith, H.R.; den Hollander, J.A.; Okonkwo, O.C.; O’Brien, T.; Watts, R.L.; Marson, D.C. Brain N-acetylaspartate is reduced in Parkinson disease with dementia. Alzheimer Dis. Assoc. Disord. 2008, 22, 54–60. [Google Scholar] [CrossRef]
- Tahay, G.; Wiame, E.; Tyteca, D.; Courtoy, P.J.; Van Schaftingen, E. Determinants of the enzymatic activity and the subcellular localization of aspartate N-acetyltransferase. Biochem. J. 2012, 441, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.H.; Chakraborty, G.; Ledeen, R.W.; Yahya, D.; Wu, G. N-Acetylaspartate synthase is bimodally expressed in microsomes and mitochondria of brain. Brain Res. Mol. Brain Res. 2004, 122, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Wilinski, D.; Winzeler, J.; Duren, W.; Persons, J.L.; Holme, K.J.; Mosquera, J.; Khabiri, M.; Kinchen, J.M.; Freddolino, P.L.; Karnovsky, A.; et al. Rapid metabolic shifts occur during the transition between hunger and satiety in Drosophila melanogaster. Nat. Commun. 2019, 10, 4052. [Google Scholar] [CrossRef] [Green Version]
- Curcio, R.; Aiello, D.; Vozza, A.; Muto, L.; Martello, E.; Cappello, A.R.; Capobianco, L.; Fiermonte, G.; Siciliano, C.; Napoli, A.; et al. Cloning, Purification, and Characterization of the Catalytic C-Terminal Domain of the Human 3-Hydroxy-3-methyl glutaryl-CoA Reductase: An Effective, Fast, and Easy Method for Testing Hypocholesterolemic Compounds. Mol. Biotechnol. 2020, 62, 119–131. [Google Scholar] [CrossRef]
- Dandurand, J.; Ostuni, A.; Armentano, M.F.; Crudele, M.A.; Dolce, V.; Marra, F.; Samouillan, V.; Bisaccia, F. ICalorimetry and FTIR reveal the ability of URG7 protein to modify the aggregation state of both cell lysate and amylogenic α-synuclein. AIMS Biophys. 2020, 7, 189–203. [Google Scholar] [CrossRef]
- Santoro, A.; Cappello, A.R.; Madeo, M.; Martello, E.; Iacopetta, D.; Dolce, V. Interaction of fosfomycin with the glycerol 3-phosphate transporter of Escherichia coli. Biochim. Biophys. Acta 2011, 1810, 1323–1329. [Google Scholar] [CrossRef]
- Carrisi, C.; Antonucci, D.; Lunetti, P.; Migoni, D.; Girelli, C.R.; Dolce, V.; Fanizzi, F.P.; Benedetti, M.; Capobianco, L. Transport of platinum bonded nucleotides into proteoliposomes, mediated by Drosophila melanogaster thiamine pyrophosphate carrier protein (DmTpc1). J. Inorg. Biochem. 2014, 130, 28–31. [Google Scholar] [CrossRef] [PubMed]
- Torraco, A.; Ardissone, A.; Invernizzi, F.; Rizza, T.; Fiermonte, G.; Niceta, M.; Zanetti, N.; Martinelli, D.; Vozza, A.; Verrigni, D.; et al. Novel mutations in IBA57 are associated with leukodystrophy and variable clinical phenotypes. J. Neurol. 2017, 264, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Lunetti, P.; Cappello, A.R.; Marsano, R.M.; Pierri, C.L.; Carrisi, C.; Martello, E.; Caggese, C.; Dolce, V.; Capobianco, L. Mitochondrial glutamate carriers from Drosophila melanogaster: Biochemical, evolutionary and modeling studies. Biochim. Biophys. Acta 2013, 1827, 1245–1255. [Google Scholar] [CrossRef] [Green Version]
- Lunetti, P.; Romano, A.; Carrisi, C.; Antonucci, D.; Verri, T.; De Benedetto, G.E.; Dolce, V.; Fanizzi, F.P.; Benedetti, M.; Capobianco, L. Platinated Nucleotides are Substrates for the Human Mitochondrial Deoxynucleotide Carrier (DNC) and DNA Polymerase Upsilon: Relevance for the Development of New Platinum-Based Drugs. Chemistryselect 2016, 1, 4633–4637. [Google Scholar] [CrossRef]
- Ito, H.; Fukuda, Y.; Murata, K.; Kimura, A. Transformation of intact yeast cells treated with alkali cations. J. Bacteriol. 1983, 153, 163–168. [Google Scholar] [CrossRef] [Green Version]
- Madeo, M.; Carrisi, C.; Iacopetta, D.; Capobianco, L.; Cappello, A.R.; Bucci, C.; Palmieri, F.; Mazzeo, G.; Montalto, A.; Dolce, V. Abundant expression and purification of biologically active mitochondrial citrate carrier in baculovirus-infected insect cells. J. Bioenerg. Biomembr. 2009, 41, 289–297. [Google Scholar] [CrossRef]
- Li, Y.; Cappello, A.R.; Muto, L.; Martello, E.; Madeo, M.; Curcio, R.; Lunetti, P.; Raho, S.; Zaffino, F.; Frattaruolo, L.; et al. Functional characterization of the partially purified Sac1p independent adenine nucleotide transport system (ANTS) from yeast endoplasmic reticulum. J. Biochem. 2018, 164, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Zara, V.; Dietmeier, K.; Palmisano, A.; Vozza, A.; Rassow, J.; Palmieri, F.; Pfanner, N. Yeast mitochondria lacking the phosphate carrier/p32 are blocked in phosphate transport but can import preproteins after regeneration of a membrane potential. Mol. Cell Biol. 1996, 16, 6524–6531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonofiglio, D.; Santoro, A.; Martello, E.; Vizza, D.; Rovito, D.; Cappello, A.R.; Barone, I.; Giordano, C.; Panza, S.; Catalano, S.; et al. Mechanisms of divergent effects of activated peroxisome proliferator-activated receptor-gamma on mitochondrial citrate carrier expression in 3T3-L1 fibroblasts and mature adipocytes. Biochim. Biophys. Acta 2013, 1831, 1027–1036. [Google Scholar] [CrossRef]
- De Marcos Lousa, C.; Trezeguet, V.; Dianoux, A.C.; Brandolin, G.; Lauquin, G.J. The human mitochondrial ADP/ATP carriers: Kinetic properties and biogenesis of wild-type and mutant proteins in the yeast S. cerevisiae. Biochemistry 2002, 41, 14412–14420. [Google Scholar] [CrossRef] [PubMed]
- Iacopetta, D.; Lappano, R.; Cappello, A.R.; Madeo, M.; De Francesco, E.M.; Santoro, A.; Curcio, R.; Capobianco, L.; Pezzi, V.; Maggiolini, M.; et al. SLC37A1 gene expression is up-regulated by epidermal growth factor in breast cancer cells. Breast Cancer Res. Treat. 2010, 122, 755–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lunetti, P.; Gorgoglione, R.; Curcio, R.; Marra, F.; Pignataro, A.; Vozza, A.; Riley, C.L.; Capobianco, L.; Palmieri, L.; Dolce, V.; et al. Drosophila melanogaster Uncoupling Protein-4A (UCP4A) Catalyzes a Unidirectional Transport of Aspartate. Int. J. Mol. Sci. 2022, 23, 1020. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031020
Lunetti P, Gorgoglione R, Curcio R, Marra F, Pignataro A, Vozza A, Riley CL, Capobianco L, Palmieri L, Dolce V, et al. Drosophila melanogaster Uncoupling Protein-4A (UCP4A) Catalyzes a Unidirectional Transport of Aspartate. International Journal of Molecular Sciences. 2022; 23(3):1020. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031020
Chicago/Turabian StyleLunetti, Paola, Ruggiero Gorgoglione, Rosita Curcio, Federica Marra, Antonella Pignataro, Angelo Vozza, Christopher L. Riley, Loredana Capobianco, Luigi Palmieri, Vincenza Dolce, and et al. 2022. "Drosophila melanogaster Uncoupling Protein-4A (UCP4A) Catalyzes a Unidirectional Transport of Aspartate" International Journal of Molecular Sciences 23, no. 3: 1020. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031020