
Thyroid Hormone Receptor α Controls the Hind Limb Metamorphosis by Regulating Cell Proliferation and Wnt Signaling Pathways in Xenopus tropicalis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
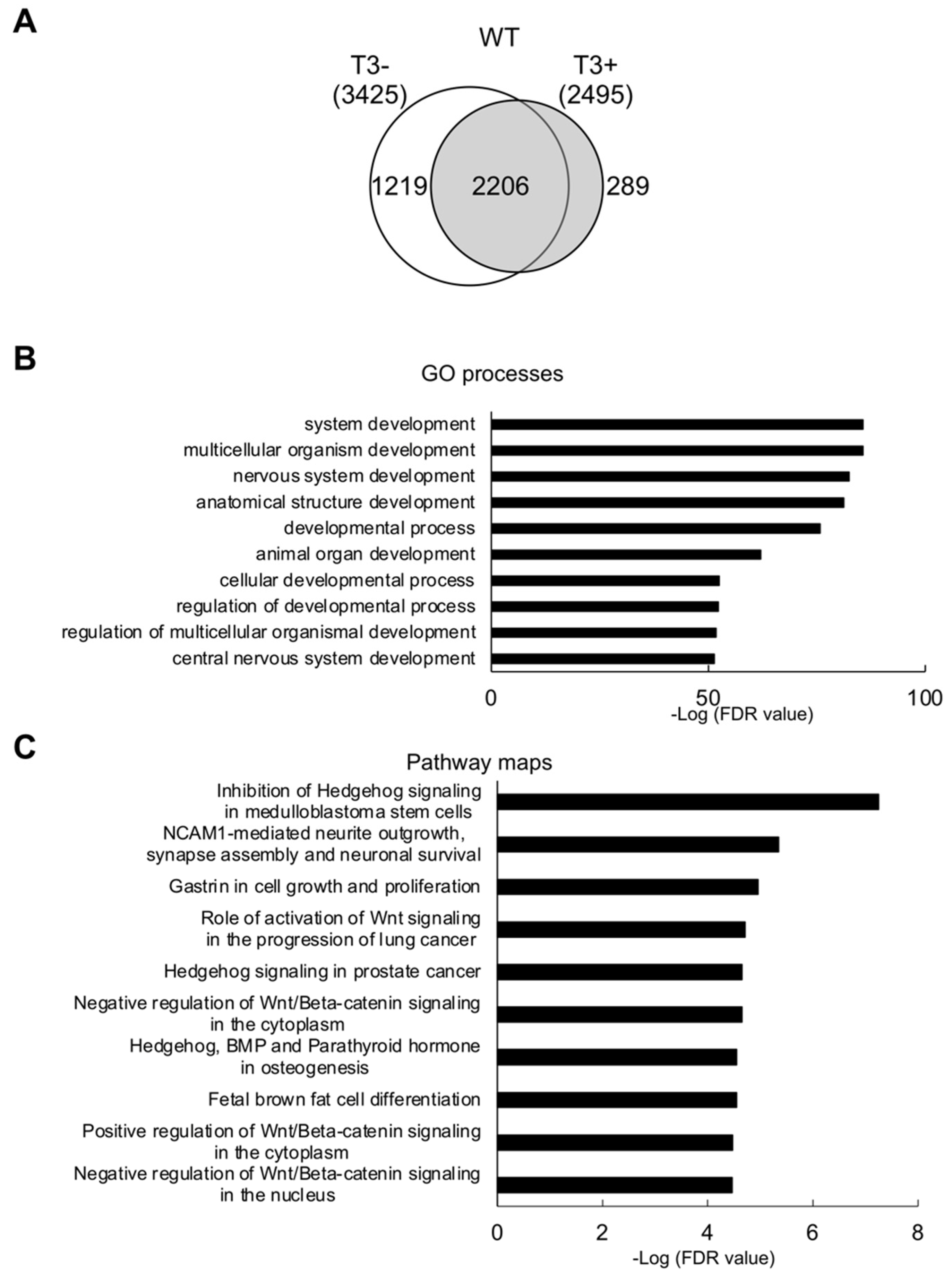
2.1. Direct Target Genes of T3 in Wild Type Hind Limb at the Onset of the Metamorphosis
2.2. Direct Target Genes of TRα in Hind Limb at the Onset of the Metamorphosis
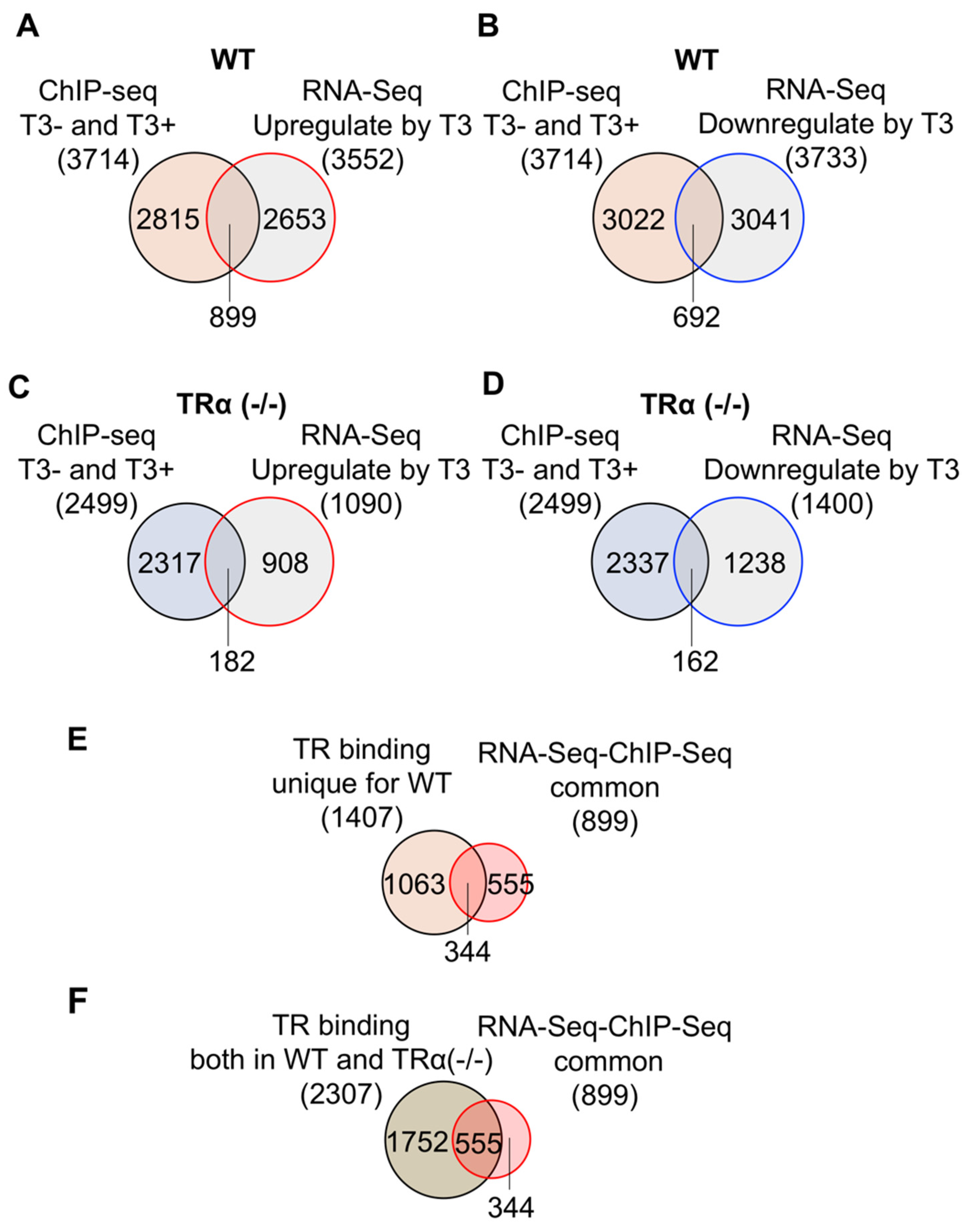
2.3. Gene Regulation by T3 in Wild Type and TRα (-/-) Hind limb
2.4. TRα Knockout Leads to Derepression of T3 Response Genes and Precocious Activation of Hind Limb Development Program
2.5. TRα Knockout Reduces the Number of TR-Bound Genes Regulated by T3 in the Hind Limb
2.6. Coordinate TR-Binding and T3-Regulation of Genes in Cell Cycle and Wnt/β-Catenin Signaling Pathways by TRα during Hind Limb Development
3. Discussion
3.1. TRα Is Critical for Both the Binding of Many Target Genes by TR and Ensuring Sufficient Levels of TR Binding at Target Genes for Their Regulation by T3 during Limb Development
3.2. TRα Regulates Pathways Such as Cell Cycle and Wnt/β-Catenin Signaling to Control the Timing and Rate of Limb Development
4. Materials and Methods
4.1. Animals
4.2. Chromatin Immunoprecipitation-Sequencing (ChIP-Seq) and ChIP-PCR Analysis
4.3. ChIP-Seq Data Processing
4.4. Quantitative Real-Time PCR
4.5. RNA-Sequencing (RNA-Seq) Analysis
4.6. GO and Pathway Analysis
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lazar, M.A. Thyroid hormone receptors: Multiple forms, multiple possibilities. Endocr. Rev. 1993, 14, 184–193. [Google Scholar] [CrossRef]
- Yen, P.M.; Heart, B. Physiological and molecular basis of Thyroid hormone action. Physiol. Rev. 2001, 81, 1097–1142. [Google Scholar] [CrossRef] [Green Version]
- Tata, J.R. Gene expression during metamorphosis: An ideal model for post-embryonic development. Bioessays 1993, 15, 239–248. [Google Scholar] [CrossRef]
- Shi, Y.-B. Amphibian Metamorphosis: From Morphology to Molecular Biology; John Wiley and Sons Ltd.: New York, NY, USA, 1999. [Google Scholar]
- DeGroot, L.J. The Story of Iodine Deficiency: An International Challenge in Nutrition. JAMA J. Am. Med. Assoc. 1990, 264, 2684. [Google Scholar] [CrossRef]
- Querido, A. Endemic cretinism—A continuous personal educational experience during 10 years. Postgrad. Med. J. 1975, 51, 591–599. [Google Scholar] [CrossRef] [Green Version]
- Laudet, V.; Gronemeyer, H. The Nuclear Receptor FactsBook; Gulf Professional Publishing: Houston, TX, USA, 2002; ISBN 9780124377356. [Google Scholar]
- Shi, Y.-B.; Matsuura, K.; Fujimoto, K.; Wen, L.; Fu, L. Thyroid hormone receptor actions on transcription in amphibia: The roles of histone modification and chromatin disruption. Cell Biosci. 2012, 2, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, L.J.; Baniahmad, A. Co-repressors 2000. FASEB J. 2000, 14, 1876–1888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, P.L.; Shi, Y.-B. N-CoR-HDAC Corepressor Complexes: Roles in Transcriptional Regulation by Nuclear Hormone Receptors. In Protein Complexes that Modify Chromatin; Springer Science & Business Media: Berlin, Germany, 2003; pp. 237–268. [Google Scholar]
- Zhang, J.; Lazar, M.A. The Mechanism of Action of Thyroid Hormones. Annu. Rev. Physiol. 2000, 62, 439–466. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.L.; Sachs, L.M.; Rouse, N.; Wade, P.A.; Shi, Y.-B. Multiple N-CoR Complexes Contain Distinct Histone Deacetylases. J. Biol. Chem. 2001, 276, 8807–8811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mckenna, N.J.; O’malley, B.W. Nuclear Receptors, Coregulators, Ligands, and Selective Receptor Modulators. Ann. N. Y. Acad. Sci. 2006, 949, 3–5. [Google Scholar] [CrossRef]
- Wong, J.; Shi, Y.B.; Wolffe, A.P. A role for nucleosome assembly in both silencing and activation of the Xenopus TRβA gene by the thyroid hormone receptor. Genes Dev. 1995, 9, 2696–2711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, R.M. The steroid and thyroid hormone receptor superfamily. Science 1988, 240, 889–895. [Google Scholar] [CrossRef]
- Tsai, M.; O’Malley, B.W. Molecular Mechanisms of Action of Steroid/Thyroid Receptor Superfamily Members. Annu. Rev. Biochem. 1994, 63, 451–486. [Google Scholar] [CrossRef] [PubMed]
- Sirakov, M.; Plateroti, M. The thyroid hormones and their nuclear receptors in the gut: From developmental biology to cancer. Biochim. Biophys. Acta-Mol. Basis Dis. 2011, 1812, 938–946. [Google Scholar] [CrossRef]
- Flamant, F.; Samarut, J. Thyroid hormone receptors: Lessons from knockout and knock-in mutant mice. Trends Endocrinol. Metab. 2003, 14, 85–90. [Google Scholar] [CrossRef]
- Jones, I.; Srinivas, M.; Ng, L.; Forrest, D. The Thyroid Hormone Receptor β Gene: Structure and Functions in the Brain and Sensory Systems. Thyroid 2003, 13, 1057–1068. [Google Scholar] [CrossRef]
- Hadj-Sahraoui, N.; Seugnet, I.; Ghorbel, M.T.; Demeneix, B. Hypothyroidism prolongs mitotic activity in the post-natal mouse brain. Neurosci. Lett. 2000, 280, 79–82. [Google Scholar] [CrossRef]
- White, P.; Burton, K.A.; Fowden, A.L.; Dauncey, M.J. Developmental expression analysis of thyroid hormone receptor isoforms reveals new insights into their essential functions in cardiac and skeletal muscles. FASEB J. 2001, 15, 1367–1376. [Google Scholar] [CrossRef]
- Shibata, Y.; Wen, L.; Okada, M.; Shi, Y.-B. Organ-Specific Requirements for Thyroid Hormone Receptor Ensure Temporal Coordination of Tissue-Specific Transformations and Completion of Xenopus Metamorphosis. Thyroid 2020, 30, 300–313. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Shibata, Y.; Su, D.; Fu, L.; Luu, N.; Shi, Y.-B. Thyroid Hormone Receptor α Controls Developmental Timing and Regulates the Rate and Coordination of Tissue-Specific Metamorphosis in Xenopus tropicalis. Endocrinology 2017, 158, 1985–1998. [Google Scholar] [CrossRef] [PubMed]
- Shibata, Y.; Tanizaki, Y.; Shi, Y.-B. Thyroid hormone receptor beta is critical for intestinal remodeling during Xenopus tropicalis metamorphosis. Cell Biosci. 2020, 10, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakajima, K.; Tazawa, I.; Yaoita, Y. Thyroid Hormone Receptor α– and β–Knockout Xenopus tropicalis Tadpoles Reveal Subtype-Specific Roles During Development. Endocrinology 2018, 159, 733–743. [Google Scholar] [CrossRef]
- Sakane, Y.; Iida, M.; Hasebe, T.; Fujii, S.; Buchholz, D.R.; Ishizuya-Oka, A.; Yamamoto, T.; Suzuki, K.T. Functional analysis of thyroid hormone receptor beta in Xenopus tropicalis founders using CRISPR-Cas. Biol. Open 2018, 7, bio030338. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.-B. Life without thyroid hormone receptor. Endocrinology 2021, 162, bqab028. [Google Scholar] [CrossRef]
- Shi, Y.; Wong, J.; Puzianowska-Kuznicka, M.; Stolow, M.A. Tadpole competence and tissue-specific temporal regulation of amphibian metamorphosis: Roles of thyroid hormone and its receptors. BioEssays 1996, 18, 391–399. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geetha-Loganathan, P.; Nimmagadda, S.; Scaal, M. Wnt signaling in limb organogenesis. Organogenesis 2008, 4, 109–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.; Ishizuya-Oka, A.; Buchholz, D.R. Growth, Development, and Intestinal Remodeling Occurs in the Absence of Thyroid Hormone Receptor α in Tadpoles of Xenopus tropicalis. Endocrinology 2017, 158, 1623–1633. [Google Scholar] [CrossRef]
- Tanizaki, Y.; Shibata, Y.; Zhang, H.; Shi, Y.-B. Analysis of Thyroid Hormone Receptor α-Knockout Tadpoles Reveals That the Activation of Cell Cycle Program Is Involved in Thyroid Hormone-Induced Larval Epithelial Cell Death and Adult Intestinal Stem Cell Development During Xenopus tropicalis Metamorphos. Thyroid 2021, 31, 128–142. [Google Scholar] [CrossRef] [PubMed]
- Tanizaki, Y.; Zhang, H.; Shibata, Y.; Shi, Y.-B. Thyroid hormone receptor α controls larval intestinal epithelial cell death by regulating the CDK1 pathway Commun Biol. in press.
- Lawrence, I.G.; Jamshed, R.T.; Atkinson, B.G. (Eds.) Metamorphosis: Postembryonic Reprogramming of Gene Expression in Amphibian and Insect Cells; Academic Press: Cambridge, MA, USA, 1996; Volume 103. [Google Scholar]
- Ito, M.; Roeder, R.G. The TRAP/SMCC/Mediator complex and thyroid hormone receptor function. Trends Endocrinol. Metab. 2001, 12, 127–134. [Google Scholar] [CrossRef]
- Rachez, C.; Freedman, L.P. Mechanisms of gene regulation by vitamin D3 receptor: A network of coactivator interactions. Gene 2000, 246, 9–21. [Google Scholar] [CrossRef]
- Jepsen, K.; Rosenfeld, M.G. Biological roles and mechanistic actions of corepressor complexes. J. Cell Sci. 2002, 115, 689–698. [Google Scholar] [CrossRef]
- Rachez, C.; Freedman, L.P. Mediator complexes and transcription. Curr. Opin. Cell Biol. 2001, 13, 274–280. [Google Scholar] [CrossRef]
- Hu, X.; Lazar, M.A. Transcriptional Repression by Nuclear Hormone Receptors. Trends Endocrinol. Metab. 2000, 11, 6–10. [Google Scholar] [CrossRef]
- Brown, D.D.; Cai, L.; Das, B.; Marsh-Armstrong, N.; Schreiber, A.M.; Juste, R. Thyroid hormone controls multiple independent programs required for limb development in Xenopus laevis metamorphosis. Proc. Natl. Acad. Sci. USA 2005, 102, 12455–12458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soshnikova, N.; Zechner, D.; Huelsken, J.; Mishina, Y.; Behringer, R.R.; Taketo, M.M.; Crenshaw, E.B.; Birchmeier, W. Genetic interaction between Wnt/β-catenin and BMP receptor signaling during formation of the AER and the dorsal-ventral axis in the limb. Genes Dev. 2003, 17, 1963–1968. [Google Scholar] [CrossRef] [Green Version]
- Petit, F.; Sears, K.E.; Ahituv, N. Limb development: A paradigm of gene regulation. Nat. Rev. Genet. 2017, 18, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Nieuwkoop, P.F.J. Normal Table of Xenopus Laevis (Daudin): A Systematical & Chronological Survey of the Development from the Fertilized Egg till the End of Metamorphosis; Garland Science: New York, NY, USA, 1994; ISBN 9780815318965. [Google Scholar]
- Tanizaki, Y.; Bao, L.; Shi, B.; Shi, Y.-B. A Role of Endogenous Histone Acetyltransferase Steroid Hormone Receptor Coactivator 3 in Thyroid Hormone Signaling During Xenopus Intestinal Metamorphosis. Thyroid 2021, 31, 692–702. [Google Scholar] [CrossRef]
- Wang, X.; Matsuda, H.; Shi, Y.B. Developmental regulation and function of thyroid hormone receptors and 9-cis retinoic acid receptors during Xenopus tropicalis metamorphosis. Endocrinology 2008, 149, 5610–5618. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.B.; Liang, V.C.T.C.-T. Cloning and characterization of the ribosomal protein L8 gene from Xenopus laevis. Biochim. Biophys. Acta-Gene Struct. Expr. 1994, 1217, 227–228. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanizaki, Y.; Shibata, Y.; Zhang, H.; Shi, Y.-B. Thyroid Hormone Receptor α Controls the Hind Limb Metamorphosis by Regulating Cell Proliferation and Wnt Signaling Pathways in Xenopus tropicalis. Int. J. Mol. Sci. 2022, 23, 1223. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031223
Tanizaki Y, Shibata Y, Zhang H, Shi Y-B. Thyroid Hormone Receptor α Controls the Hind Limb Metamorphosis by Regulating Cell Proliferation and Wnt Signaling Pathways in Xenopus tropicalis. International Journal of Molecular Sciences. 2022; 23(3):1223. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031223
Chicago/Turabian StyleTanizaki, Yuta, Yuki Shibata, Hongen Zhang, and Yun-Bo Shi. 2022. "Thyroid Hormone Receptor α Controls the Hind Limb Metamorphosis by Regulating Cell Proliferation and Wnt Signaling Pathways in Xenopus tropicalis" International Journal of Molecular Sciences 23, no. 3: 1223. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031223