
Anti-Inflammatory Effect of an O-2-Substituted (1-3)-β-D-Glucan Produced by Pediococcus parvulus 2.6 in a Caco-2 PMA-THP-1 Co-Culture Model

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effects of (1-3)-β-D-Glucans on Caco-2 Cells Viability
2.2. Effects of (1-3)-β-D-Glucans on the Viability of PMA-THP-1 Cells
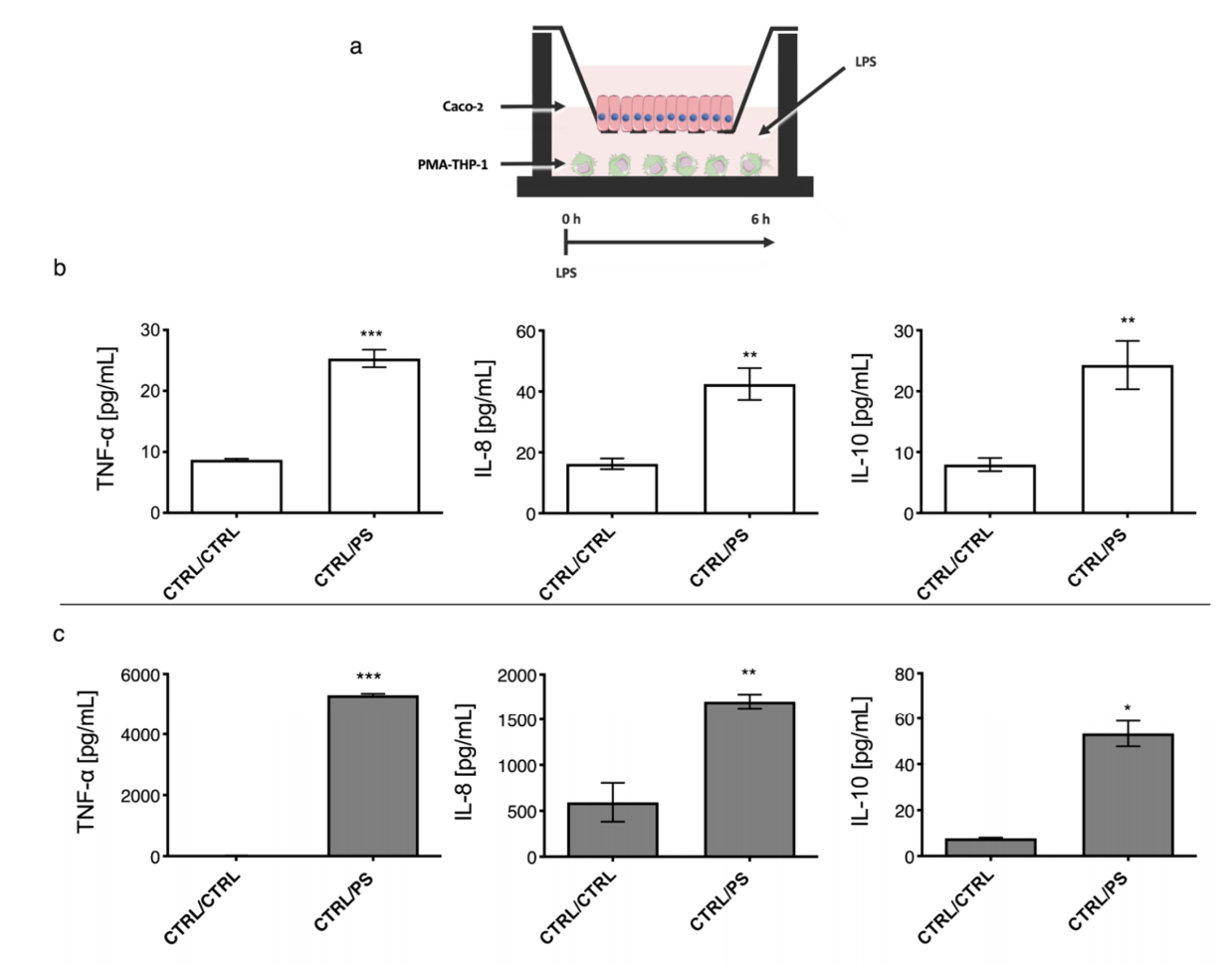
2.3. Establishment of a Co-Culture Model with Caco-2 and PMA-THP-1 Cells under LPS Stimulation
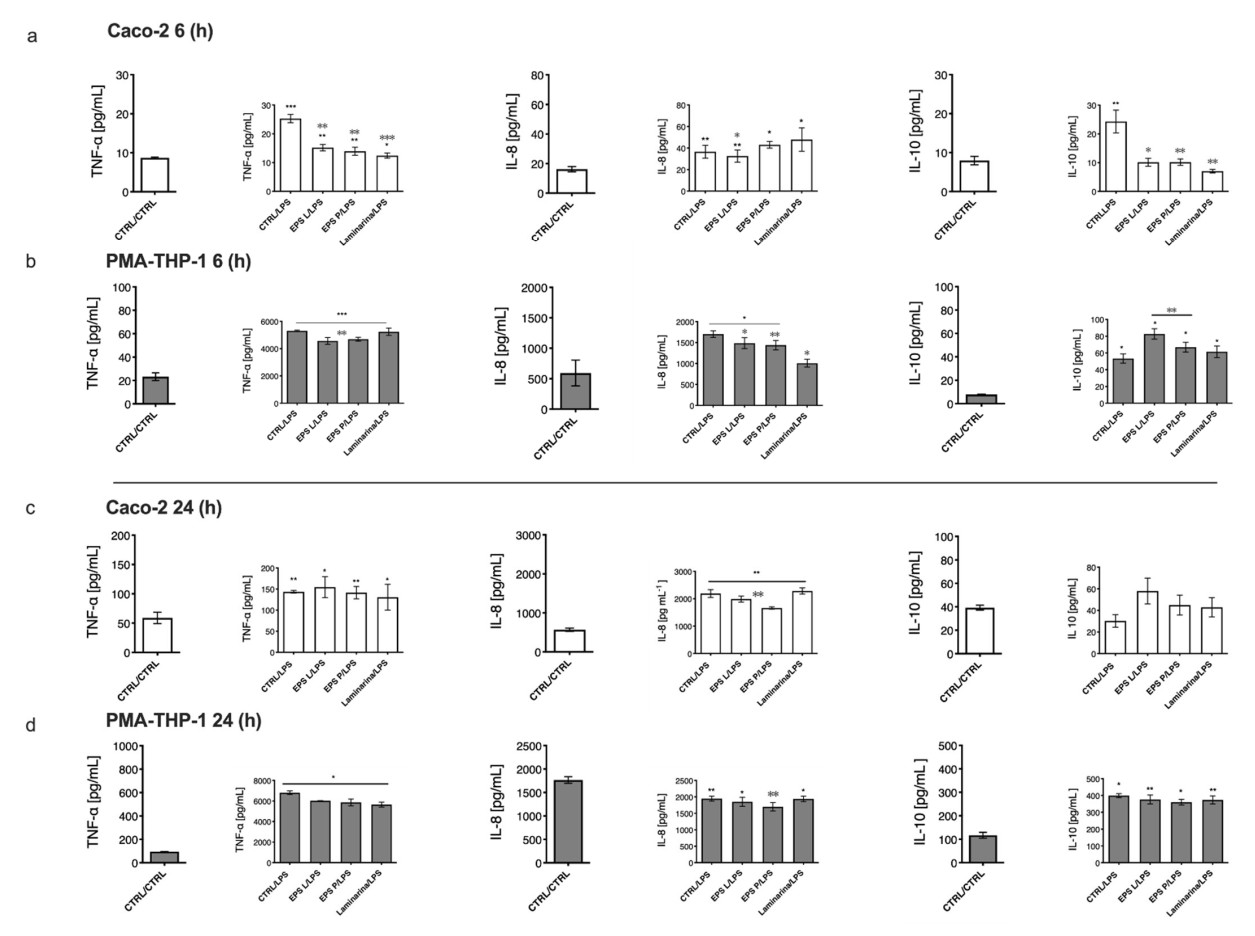
2.4. Immunomodulation of O-2-Substituted (1-3)-β-D-Glucan Produced by P. parvulus 2.6 on Cytokine Production in the Co-Cultured Model of Gut Mucosa
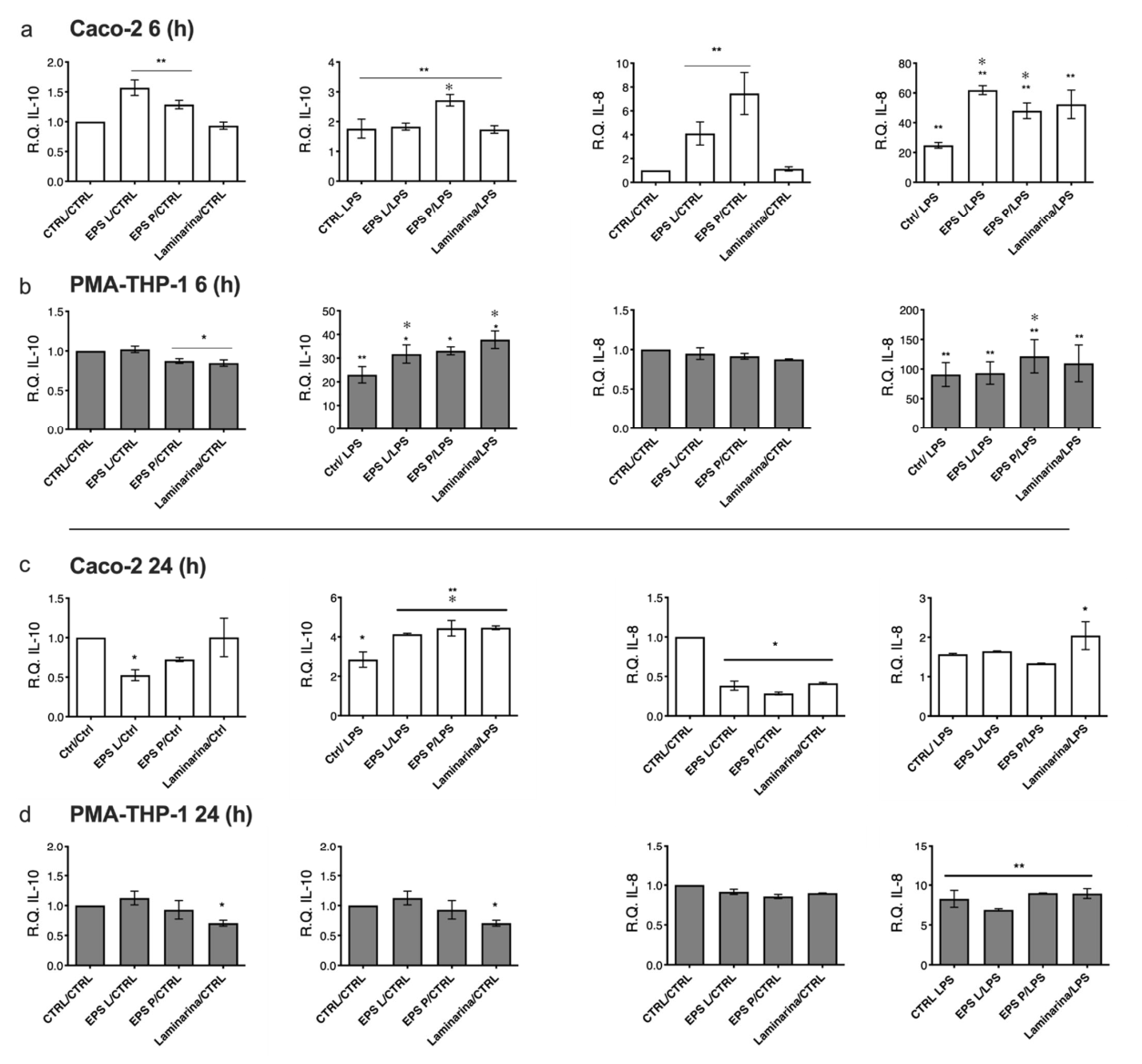
2.5. Effect of O-2-Substituted (1-3)-β-D-Glucan Produced by P. parvulus 2.6 on Gene Expression Profiles
3. Discussion
4. Materials and Methods
4.1. Cell Cultures
4.2. EPS P and EPS L Production and Purification
4.3. Cytotoxic Assay with XTT
4.4. Co-Cultured In Vitro Gut Model of Caco-2 and PMA-THP-1
4.5. Determination of Cytokine Levels
4.6. Total RNA Extraction and Analysis
4.7. Synthesis of cDNA from Total RNA Preparations
4.8. Analysis of Gene Expression by Quantitative PCR
4.9. Statistical Analyses
- Treatment vs. untreated control to assess the possible effects of the EPS and laminarin in a basal state vs. untreated control. Significant values were represented as p*.
- Treatment vs. inflammation control: to assess possible inflammation-modulating effects of EPSs and laminarin vs. inflammatory state (LPS treatment). Significant values were represented as p✽.
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef] [PubMed]
- Guinane, C.M.; Cotter, P.D. Role of the gut microbiota in health and chronic gastrointestinal disease: Understanding a hidden metabolic organ. Ther. Adv. Gastroenterol. 2013, 6, 295–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Deferme, S.; Annaert, P.; Augustijns, P. In vitro screening models to assess intestinal drug absorption and tetabolism. In Drug Absorption Studies; Ehrhardt, C., Kim, K.-J., Eds.; Springer: New York, NY, USA, 2008; Volume 7, pp. 182–215. [Google Scholar]
- Lesuffleur, T.; Porchet, N.; Aubert, J.P.; Swallow, D.; Gum, J.R.; Kim, Y.S.; Real, F.X.; Zweibaum, A. Differential expression of the human mucin genes MUC1 to MUC5 in relation to growth and differentiation of different mucus-secreting HT-29 cell subpopulations. J. Cell Sci. 1993, 106, 771–783. [Google Scholar] [CrossRef] [PubMed]
- Kauffman, A.L.; Gyurdieva, A.V.; Mabus, J.R.; Ferguson, C.; Yan, Z.; Hornby, P.J. Alternative functional in vitro models of human intestinal epithelia. Front. Pharmacol. 2013, 4, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Creff, J.; Malaquin, L.; Besson, A. In vitro models of intestinal epithelium: Toward bioengineered systems. J. Tissue Eng. 2021, 12, 2041731420985202. [Google Scholar] [CrossRef]
- Meunier, V.; Bourrié, M.; Berger, Y.; Fabre, G. The human intestinal epithelial cell line Caco-2; pharmacological and pharmacokinetic applications. Cell Biol. Toxicol. 1995, 11, 187–194. [Google Scholar] [CrossRef]
- Phillips, T.E.; Huet, C.; Bilbo, P.R.; Podolsky, D.K.; Louvard, D.; Neutra, M.R. Human intestinal goblet cells in monolayer culture: Characterization of a mucus-secreting subclone derived from the HT29 colon adenocarcinoma cell line. Gastroenterology 1988, 94, 1390–1403. [Google Scholar] [CrossRef]
- Schoultz, I.; Keita, Å.V. The Intestinal Barrier and Current Techniques for the Assessment of Gut Permeability. Cells 2020, 9, 1909. [Google Scholar] [CrossRef]
- Fernández de Palencia, P.; Werning, M.L.; Sierra-Filardi, E.; Dueñas-Chasco, M.; Irastorza, A.; Corbí, A.L.; López, P. Probiotic properties of the 2-substituted (1-3)-β-D-glucan-producing bacterium Pediococcus parvulus 2.6. Appl. Environ. Microbiol. 2009, 75, 4887–4891. [Google Scholar] [CrossRef] [Green Version]
- Garai-Ibabe, G.; Dueñas, M.A.T.; Irastorza, A.; Sierra-Filardi, E.; Werning, M.L.; López, P.; Corbí, A.L.; Fernández de Palencia, P. Naturally occurring 2-substituted (1,3)-beta-D-glucan producing Lactobacillus suebicus and Pediococcus parvulus strains with potential utility in the production of functional foods. Bioresour. Technol. 2010, 101, 9254–9263. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, H.C.; de Sousa Melo, D.; Ramos, C.L.; Dias, D.R.; Schwan, R.F. Probiotic Properties of Lactobacilli and Their Ability to Inhibit the Adhesion of Enteropathogenic Bacteria to Caco-2 and HT-29 Cells. Probiotics Antimicrob. Proteins 2021, 13, 102–112. [Google Scholar] [CrossRef]
- de los Reyes-Gavilán, C.G.; Suárez, A.; Fernández-García, M.; Margolles, A.; Gueimonde, M.; Ruas-Madiedo, P. Adhesion of bile-adapted Bifidobacterium strains to the HT29-MTX cell line is modified after sequential gastrointestinal challenge simulated in vitro using human gastric and duodenal juices. Res. Microbiol. 2011, 162, 514–519. [Google Scholar] [CrossRef] [Green Version]
- López, P.; González-Rodríguez, I.; Sánchez, B.; Ruas-Madiedo, P.; Suárez, A.; Margolles, A.; Gueimonde, M. Interaction of Bifidobacterium bifidum LMG13195 with HT29 cells influences regulatory-T-cell-associated chemokine receptor expression. Appl. Environ. Microbiol. 2012, 78, 2850–2857. [Google Scholar] [CrossRef] [Green Version]
- Payne, A.N.; Zihler, A.; Chassard, C.; Lacroix, C. Advances and perspectives in in vitro human gut fermentation modeling. Trends Biotechnol. 2012, 30, 17–25. [Google Scholar] [CrossRef]
- Tanoue, T.; Nishitani, Y.; Kanazawa, K.; Hashimoto, T.; Mizuno, M. In vitro model to estimate gut inflammation using co-cultured Caco-2 and RAW264.7 cells. Biochem. Biophys. Res. Res Commun. 2008, 374, 565–569. [Google Scholar] [CrossRef] [Green Version]
- Takisawa, R.; Nishitani, Y.; Mizuno, M.; Osawa, R. Anti-Inflammatory effect of Bifidobacterium longum on macrophage-like THP-1 cells via epithelial cell Caco-2. Biosci. Microbiota Food Health 2009, 28, 45–48. [Google Scholar] [CrossRef] [Green Version]
- Kordulewska, N.K.; Topa, J.; Tańska, M.; Cieślińska, A.; Fiedorowicz, E.; Savelkoul, H.F.J.; Jarmołowska, B. Modulatory Effects of Osthole on Lipopolysaccharides-Induced Inflammation in Caco-2 Cell Monolayer and Co-Cultures with THP-1 and THP-1-Derived Macrophages. Nutrients 2021, 13, 123. [Google Scholar] [CrossRef]
- Dertli, E.; Mayer, M.J.; Narbad, A. Impact of the exopolysaccharide layer on biofilms, adhesion and resistance to stress in Lactobacillus johnsonii FI9785. BMC Microbiol. 2015, 15, 8. [Google Scholar] [CrossRef] [Green Version]
- Lebeer, S.; Verhoeven, T.L.; Francius, G.; Schoofs, G.; Lambrichts, I.; Dufrene, Y.; Vanderleyden, J.; De Keersmaecker, S.C. Identification of a gene cluster for the biosynthesis of a long, galactose-rich exopolysaccharide in Lactobacillus rhamnosus GG and functional Analysis of the priming glycosyltransferase. Appl. Environ. Microbiol. 2009, 75, 3554–3563. [Google Scholar] [CrossRef] [Green Version]
- Lebeer, S.; Claes, I.; Tytgat, H.L.; Verhoeven, T.L.; Marien, E.; von Ossowski, I.; Reunanen, J.; Palva, A.; Vos, W.M.; Keersmaecker, S.C.; et al. Functional analysis of Lactobacillus rhamnosus GG pili in relation to adhesion and immunomodulatory interactions with intestinal epithelial cells. Appl. Environ. Microbiol. 2012, 78, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Nácher-Vázquez, M.; Iturria, I.; Zarour, K.; Mohedano, M.L.; Aznar, R.; Pardo, M.; López, P. Dextran production by Lactobacillus sakei MN1 coincides with reduced autoagglutination, biofilm formation and epithelial cell adhesion. Carbohydr. Polym. 2017, 168, 22–31. [Google Scholar] [CrossRef]
- Zarour, K.; Llamas, M.G.; Prieto, A.; Rúas-Madiedo, P.; Dueñas, M.T.; de Palencia, P.F.; Aznar, R.; Kihal, M.; López, P. Rheology and bioactivity of high molecular weight dextrans synthesised by lactic acid bacteria. Carbohydr. Polym. 2017, 174, 646–657. [Google Scholar] [CrossRef]
- Besrour-Aouam, N.; Fhoula, I.; Hernández-Alcántara, A.M.; Mohedano, M.L.; Najjari, A.; Prieto, A.; Ruas-Madiedo, P.; López, P.; Ouzari, H.-I. The role of dextran production in the metabolic context of Leuconostoc and Weissella Tunisian strains. Carbohydr. Polym. 2021, 253, 117254. [Google Scholar] [CrossRef]
- Notararigo, S.; de Las Casas-Engel, M.; Fernández de Palencia, P.; Corbí, A.L.; López, P. Immunomodulation of human macrophages and myeloid cells by 2-substituted (1-3)-beta-D-glucan from P. parvulus 2.6. Carbohydr. Polym. 2014, 112, 109–113. [Google Scholar] [CrossRef] [Green Version]
- Notararigo, S.; Varela, E.; Otal, A.; Cristobo, I.; Antolín, M.; Guarner, F.; Prieto, A.; López, P. Evaluation of an O-2-Substituted (1-3)-β-D-Glucan, Produced by Pediococcus parvulus 2.6, in ex vivo Models of Crohn’s Disease. Front. Microbiol. 2021, 12, 621280. [Google Scholar] [CrossRef]
- Pérez-Ramos, A.; Mohedano, M.L.; Pardo, M.Á.; López, P. β-Glucan-Producing Pediococcus parvulus 2.6: Test of Probiotic and Immunomodulatory Properties in Zebrafish Models. Front. Microbiol. 2018, 9, 1684. [Google Scholar] [CrossRef]
- Russo, P.; López, P.; Capozzi, V.; de Palencia, P.F.; Dueñas, M.T.; Spano, G.; Fiocco, D. Beta-glucans improve growth, viability and colonization of probiotic microorganisms. Int. J. Mol. Sci. 2012, 13, 6026–6039. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Ramos, A.; Mohedano, M.L.; López, P.; Spano, G.; Fiocco, D.; Russo, P.; Capozzi, V. In Situ β-Glucan Fortification of Cereal-Based Matrices by Pediococcus parvulus 2.6: Technological Aspects and Prebiotic Potential. Int. J. Mol. Sci. 2017, 18, 1588. [Google Scholar] [CrossRef] [Green Version]
- Lindström, C.; Holst, O.; Nilsson, L.; Oste, R.; Andersson, K.E. Effects of Pediococcus parvulus 2.6 and its exopolysaccharide on plasma cholesterol levels and inflammatory markers in mice. AMB Express 2012, 2, 66. [Google Scholar] [CrossRef] [Green Version]
- Lindström, C.; Xu, J.; Öste, R.; Holst, O. Oral administration of live exopolysaccharide-producing Pediococcus parvulus, but not purified exopolysaccharide, suppressed Enterobacteriaceae without affecting bacterial diversity in ceca of mice. Appl. Environ. Microbiol. 2013, 79, 5030–5037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- London, L.E.; Kumar, A.H.; Wall, R.; Casey, P.G.; O’Sullivan, O.; Shanahan, F.; Hill, C.; Cotter, P.D.; Fitzgerald, G.F.; Ross, R.P.; et al. Exopolysaccharide-producing probiotic Lactobacilli reduce serum cholesterol and modify enteric microbiota in ApoE-deficient mice. J. Nutr. 2014, 144, 1956–1962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mårtensson, O.; Biörklund, M.; Lambo, A.M. Fermented, ropy, oat-based products reduce cholesterol levels and stimulate the bifidobacteria flora in humans. Nutr. Res. 2005, 25, 429–442. [Google Scholar] [CrossRef]
- Yang, Y.; Zhao, Y.; Yu, A.; Sun, D.; Yu, L.X. Chapter 12—Oral Drug Absorption: Evaluation and Prediction. In Developing Solid Oral Dosage Forms, 2nd ed.; Qiu, Y., Chen, Y., Zhang, G.G.Z., Yu, L., Mantri, R.V., Eds.; Academic Press: Boston, MA, USA, 2017; pp. 331–354. [Google Scholar]
- Sonnier, D.; Bailey, S.; Schuster, R.; Lentsch, A.; Pritts, T. TNF-α induces vectorial secretion of IL-8 in Caco-2 cells. J. Gastrointest. Surg. 2010, 14, 1592–1599. [Google Scholar] [CrossRef] [Green Version]
- Rossi, O.; Karczewski, J.; Stolte, E.H.; Brummer, R.J.M.; van Nieuwenhoven, M.A.; Meijerink, M.; van Neerven, J.R.J.; van Ijzendoorn, S.C.D.; van Baarlen, P.; Wells, J.M. Vectorial secretion of interleukin-8 mediates autocrine signalling in intestinal epithelial cells via apically located CXCR1. BMC Res. Notes 2013, 6, 431. [Google Scholar] [CrossRef] [Green Version]
- Claud, E.C.; Savidge, T.; Walker, W.A. Modulation of Human Intestinal Epithelial Cell IL-8 Secretion by Human Milk Factors. Pediatr. Res. 2003, 53, 419–425. [Google Scholar] [CrossRef] [Green Version]
- Abreu, M.T.; Vora, P.; Faure, E.; Thomas, L.S.; Arnold, E.T.; Arditi, M. Decreased expression of Toll-like receptor-4 and MD-2 correlates with intestinal epithelial cell protection against dysregulated proinflammatory gene expression in response to bacterial lipopolysaccharide. J. Immunol. 2001, 167, 1609–1616. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Al-Sadi, R.; Said, H.M.; Ma, T.Y. Lipopolysaccharide causes an increase in intestinal tight junction permeability in vitro and in vivo by inducing enterocyte membrane expression and localization of TLR-4 and CD14. Am. J. Pathol. 2013, 182, 375–387. [Google Scholar] [CrossRef] [Green Version]
- Stephens, M.; Liao, S.; von der Weid, P.-Y. Ultra-purification of Lipopolysaccharides reveals species-specific signalling bias of TLR4: Importance in macrophage function. Sci. Rep. 2021, 11, 1335. [Google Scholar] [CrossRef]
- Oosenbrug, T.; van de Graaff, M.J.; Haks, M.C.; van Kasteren, S.; Ressing, M.E. An alternative model for type I interferon induction downstream of human TLR2. J. Biol. Chem. 2020, 295, 14325–14342. [Google Scholar] [CrossRef]
- Rattigan, R.; O’Doherty, J.V.; Vigors, S.; Ryan, M.T.; Sebastiano, R.S.; Callanan, J.J.; Thornton, K.; Rajauria, G.; Margassery, L.M.; Dobson, A.D.W.; et al. The Effects of the Marine-Derived Polysaccharides Laminarin and Chitosan on Aspects of Colonic Health in Pigs Challenged with Dextran Sodium Sulphate. Mar. Drugs 2020, 18, 262. [Google Scholar] [CrossRef]
- Vigors, S.; O’Doherty, J.V.; Rattigan, R.; McDonnell, M.J.; Rajauria, G.; Sweeney, T. Effect of a Laminarin Rich Macroalgal Extract on the Caecal and Colonic Microbiota in the Post-Weaned Pig. Mar. Drugs 2020, 18, 157. [Google Scholar] [CrossRef] [Green Version]
- Ryan, M.T.; Collins, C.B.; O’Doherty, J.V.; Sweeney, T. Effects of dietary β-glucans supplementation on cytokine expression in porcine liver. J. Anim. Sci. 2012, 90, 40–42. [Google Scholar] [CrossRef]
- Werning, M.L.; Pérez-Rámos, A.; Fernández de Palencia, P.; Mohedano, M.L.; Dueñas, M.T.; Prieto, A.; López, P. A specific immunological method to detect and quantify bacterial 2-substituted (1,3)-beta-D-glucan. Carbohydr. Polym. 2014, 113, 39–45. [Google Scholar] [CrossRef]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in Inflammatory Disease. Int. J. Mol. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef] [Green Version]
- Van De Walle, J.; Hendrickx, A.; Romier, B.; Larondelle, Y.; Schneider, Y.-J. Inflammatory parameters in Caco-2 cells: Effect of stimuli nature, concentration, combination and cell differentiation. Toxicol. Vitr. 2010, 24, 1441–1449. [Google Scholar] [CrossRef]
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef]
- Dueñas-Chasco, M.; Rodriguez-Carvajal, M.A.; Tejero-Mateo, P.; Franco-Rodriguez, G.; Espartero, J.L.; Irastorza-Iribas, A.; Gil-Serrano, A.M. Structural analysis of the exopolysaccharide produced by Pediococcus damnosus 2.6. Carbohyd. Res. 1997, 303, 453–458. [Google Scholar] [CrossRef]
- Werning, M.L.; Corrales, M.A.; Prieto, A.; Fernández de Palencia, P.; Navas, J.; López, P. Heterologous expression of a position 2-substituted (1-3)-β-D-glucan in Lactococcus lactis. Appl. Environ. Microbiol. 2008, 74, 5259–5262. [Google Scholar] [CrossRef] [Green Version]
- Notararigo, S.; Nácher-Vázquez, M.; Ibarburu, I.; Werning, M.L.; Fernández de Palencia, P.; Dueñas, M.T.; Aznar, R.; López, P.; Prieto, A. Comparative analysis of production and purification of homo- and hetero-polysaccharides produced by lactic acid bacteria. Carbohyd. Polym. 2013, 93, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Zarour, K.; Prieto, A.; Pérez-Ramos, A.; Kihal, M.; López, P. Analysis of technological and probiotic properties of Algerian L. mesenteroides strains isolated from dairy and non-dairy products. J. Funct Foods 2018, 49, 351–361. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Notararigo, S.; Varela, E.; Otal, A.; Antolín, M.; Guarner, F.; López, P. Anti-Inflammatory Effect of an O-2-Substituted (1-3)-β-D-Glucan Produced by Pediococcus parvulus 2.6 in a Caco-2 PMA-THP-1 Co-Culture Model. Int. J. Mol. Sci. 2022, 23, 1527. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031527
Notararigo S, Varela E, Otal A, Antolín M, Guarner F, López P. Anti-Inflammatory Effect of an O-2-Substituted (1-3)-β-D-Glucan Produced by Pediococcus parvulus 2.6 in a Caco-2 PMA-THP-1 Co-Culture Model. International Journal of Molecular Sciences. 2022; 23(3):1527. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031527
Chicago/Turabian StyleNotararigo, Sara, Encarnación Varela, Anna Otal, María Antolín, Francisco Guarner, and Paloma López. 2022. "Anti-Inflammatory Effect of an O-2-Substituted (1-3)-β-D-Glucan Produced by Pediococcus parvulus 2.6 in a Caco-2 PMA-THP-1 Co-Culture Model" International Journal of Molecular Sciences 23, no. 3: 1527. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031527