
Molecular Mechanisms of Malignant Transformation of Oral Submucous Fibrosis by Different Betel Quid Constituents—Does Fibroblast Senescence Play a Role?

 ,
,
Abstract
:1. Introduction
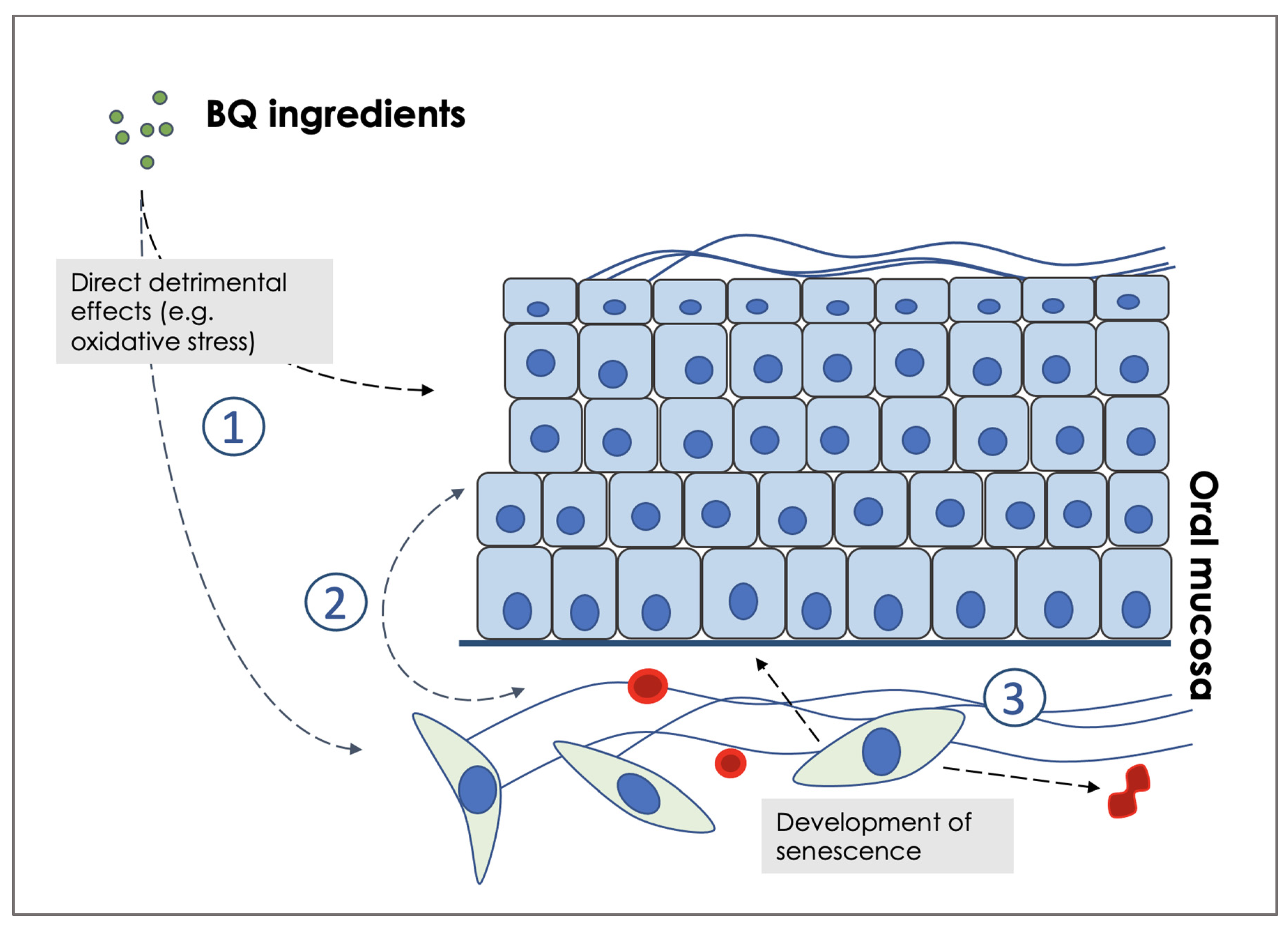
2. Why It Is Important to Understand the Effects of Individual BQ Ingredients
3. Chemical Constituents of Betel Quids
3.1. Alkaloid: Arecoline
3.2. Alkaloid: Guavacine
3.3. Alkaloid: Guvacoline and Arecaidine
3.4. New Alkaloids and Additives
3.5. Polyphenols
3.6. Other Chemical Consituents
4. Mechanisms of Malignant Transformation: Role of BQ Components
4.1. Areca Nut Alkaloids
4.2. Slaked Lime
4.3. Polyphenols and Other Compounds with Anti-Oxidant Properties
4.3.1. Catechins
4.3.2. Chavibetol
4.3.3. Flavonoids
4.3.4. Tannins
5. Fibroblast Senescence as a Possible Pathogenic Mechanism of OSMF and Progression to Malignancy
5.1. Cellular Senescence Is Associated with OSMF
5.2. Fibroblast Senescence Promotes Oxidative Stress and Oral Cancer Progression
5.3. A Pathogenic Model for OSMF and Possible Therapeutic Strategies
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- International Agency for Research on Cancer (IARC). Betel-quid and areca-nut chewing and some areca-nut derived nitrosamines. IARC Monogr. Eval. Carcinog. Risks Hum. 2004, 85, 1. [Google Scholar]
- Mehrtash, H.; Duncan, K.; Parascandola, M.; David, A.; Gritz, E.R.; Gupta, P.C.; Mehrotra, R.; Nordin, A.S.A.; Pearlman, P.C.; Warnakulasuriya, S.; et al. Defining a global research and policy agenda for betel quid and areca nut. Lancet Oncol. 2017, 18, e767–e775. [Google Scholar] [CrossRef]
- Wang, Y.-Y.; Tail, Y.-H.; Wang, W.-C.; Chen, C.-Y.; Kao, Y.-H.; Chen, Y.-K.; Chen, C.-H. Malignant transformation in 5071 southern Taiwanese patients with potentially malignant oral mucosal disorders. BMC Oral Health 2014, 14, 99. [Google Scholar] [CrossRef] [PubMed]
- Shih, Y.-H.; Wang, T.-H.; Shieh, T.-M.; Tseng, Y.-H. Oral Submucous Fibrosis: A Review on Etiopathogenesis, Diagnosis, and Therapy. Int. J. Mol. Sci. 2019, 20, 2940. [Google Scholar] [CrossRef] [Green Version]
- Bari, S.; Metgud, R.; Vyas, Z.; Tak, A. An update on studies on etiological factors, disease progression, and malignant transformation in oral submucous fibrosis. J. Cancer Res. Ther. 2017, 13, 399–405. [Google Scholar]
- Gupta, S.; Jawanda, M.K. Oral submucous fibrosis: An overview of a challenging entity. Indian J. Dermatol. Venereol. Leprol. 2021, 87, 768–777. [Google Scholar] [CrossRef]
- Yang, M.-S.; Chung, T.-C.; Yang, M.-J.; Hsu, T.-Y.; Ko, Y.-C. Betel quid chewing and risk of adverse birth outcomes among aborigines in eastern taiwan. J. Toxicol. Environ. Health Part A 2001, 64, 465–472. [Google Scholar] [CrossRef]
- Sari, E.F.; Prayogo, G.P.; Loo, Y.T.; Zhang, P.; McCullough, M.J.; Cirillo, N. Distinct phenolic, alkaloid and antioxidant profile in betel quids from four regions of Indonesia. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Jayalakshmi, A.; Mathew, G. The Areca Nut Palm; Central Plantation Crops Research Institute: Kerala, India, 1982; pp. 225–244.
- Zaidi, J.H.; Arif, M.; Fatima, I.; Qureshi, I.H. Radiochemical neutron activation analysis for trace elements of basic ingredients of pan. J. Radioanal. Nucl. Chem. 2002, 253, 459–464. [Google Scholar] [CrossRef]
- Bissa, S.; Songara, D.; Bohra, A. Traditions in oral hygiene: Chewing of betel (Piper betle L.) leaves. Curr. Sci. 2007, 92, 26–28. [Google Scholar]
- Burton-Bradley, B.G. Arecaidinism: Betel Chewing in Transcultural Perspective. Can. J. Psychiatry 1979, 24, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Tham, J.; Sem, G.; Sit, E.; Tai, M.C.-T. A Scientific and Socioeconomic Review of Betel Nut Use in Taiwan with Bioethical Reflections. Asian Bioeth. Rev. 2017, 9, 401–414. [Google Scholar] [CrossRef]
- Sharma, D.C. Betel quid and areca nut are carcinogenic without tobacco. Lancet Oncol. 2003, 4, 587. [Google Scholar] [CrossRef]
- Winstock, A. Areca nut-abuse liability, dependence and public health. Addict. Biol. 2002, 7, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.S.; Bawany, F.I.; Ahmed, M.U.; Hussain, M.; Khan, A.; Lashari, M.N. Betel Nut Usage Is a Major Risk Factor for Coronary Artery Disease. Glob. J. Health Sci. 2013, 6, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Yu, S.-Y.; Lv, Z.-T.; Yao, Y. Betel nut chewing and the risk of chronic kidney disease: Evidence from a meta-analysis. Int. Urol. Nephrol. 2018, 50, 1097–1104. [Google Scholar] [CrossRef]
- Tsai, J.-F.; Jeng, J.-E.; Chuang, L.-Y.; Ho, M.-S.; Ko, Y.-C.; Lin, Z.-Y.; Hsieh, M.-Y.; Chen, S.-C.; Chuang, W.-L.; Wang, L.-Y.; et al. Habitual Betel Quid Chewing and Risk for Hepatocellular Carcinoma Complicating Cirrhosis. Medicine 2004, 83, 176–187. [Google Scholar] [CrossRef]
- Arakeri, G.; Rai, K.K.; Hunasgi, S.; Merkx, M.A.; Gao, S.; Brennan, P.A. Oral submucous fibrosis: An update on current theories of pathogenesis. J. Oral Pathol. Med. 2017, 46, 406–412. [Google Scholar] [CrossRef]
- Trivedy, C.R.; Craig, G.; Warnakulasuriya, S. The oral health consequences of chewing areca nut. Addict. Biol. 2002, 7, 115–125. [Google Scholar] [CrossRef]
- Chang, M.-C.; Chan, C.-P.; Chen, Y.-J.; Hsien, H.-C.; Chang, Y.-C.; Yeung, S.-Y.; Jeng, P.-Y.; Cheng, R.-H.; Hahn, L.-J.; Jeng, J.-H. Areca nut components stimulate ADAM17, IL-1α, PGE2 and 8-isoprostane production in oral keratinocyte: Role of reactive oxygen species, EGF and JAK signaling. Oncotarget 2016, 7, 16879–16894. [Google Scholar] [CrossRef] [Green Version]
- Franke, A.A.; Lai, J.F.; Kawamoto, C.T.; Pokhrel, P.; A Herzog, T. University of Hawai’i Cancer Center connection: Areca (betel) nut consumption: An underappreciated cause of cancer. Hawai’i J. Med. Public Health A J. Asia Pac. Med. Public Health 2014, 73, 400–403. [Google Scholar]
- Sari, E.F.; McCullough, M.; Cirillo, N. Oral pre-malignant and malignant lesion detection among indonesian; the prevalence and risk factors. Head Neck 2017, 39, E58. [Google Scholar]
- Jain, V.; Garg, A.; Parascandola, M.; Chaturvedi, P.; Khariwala, S.S.; Stepanov, I. Analysis of Alkaloids in Areca Nut-Containing Products by Liquid Chromatography–Tandem Mass Spectrometry. J. Agric. Food Chem. 2017, 65, 1977–1983. [Google Scholar] [CrossRef] [PubMed]
- Franke, A.A.; Mendez, A.J.; Lai, J.F.; Arat-Cabading, C.; Li, X.; Custer, L.J. Composition of betel specific chemicals in saliva during betel chewing for the identification of biomarkers. Food Chem. Toxicol. 2015, 80, 241–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, M.; Yuan, H.; Daniyal, M.; Yu, H.; Xie, Q.; Liu, Y.; Li, B.; Jian, Y.; Peng, C.; Tan, D.; et al. Two new alkaloids isolated from traditional Chinese medicine Binglang the fruit of Areca catechu. Fitoterapia 2019, 138, 104276. [Google Scholar] [CrossRef]
- Srimany, A.; George, C.; Naik, H.R.; Pinto, D.G.; Chandrakumar, N.; Pradeep, T. Developmental patterning and segregation of alkaloids in areca nut (seed of Areca catechu) revealed by magnetic resonance and mass spectrometry imaging. Phytochemistry 2016, 125, 35–42. [Google Scholar] [CrossRef]
- Shirname, L.P.; Menon, M.M.; Nair, J.; Bhide, S.V. Correlation of mutagenicity and tumorigenicity of betel quid and its ingredients. Nutr. Cancer 1983, 5, 87–91. [Google Scholar] [CrossRef]
- Kadi, A.; Attwa, M.W.; Rahman, A.M. A Preliminary Study of Arecoline and Guvacoline Presence in the Saliva of a “Betel-Quid” Chewer Using Liquid-Chromatography Ion Trap Mass Spectrometry. Eur. J. Mass Spectrom. 2013, 19, 391–397. [Google Scholar] [CrossRef]
- Tang, S.-N.; Zhang, J.; Liu, D.; Liu, Z.-W.; Zhang, X.-Q.; Ye, W.-C. Three new areca alkaloids from the nuts of Areca catechu. J. Asian Nat. Prod. Res. 2017, 19, 1155–1159. [Google Scholar] [CrossRef]
- Lord, G.A.; Lim, C.K.; Warnakulasuriya, S.; Peters, T.J. Chemical and analytical aspects of areca nut. Addict. Biol. 2002, 7, 99–102. [Google Scholar] [CrossRef]
- Sur, T.K.; Adhikari, A.; Hazra, A.K. Detection of arecoline by simple high-performance thin-layer chromatographic method in Indian nontobacco pan masala. J. Adv. Pharm. Technol. Res. 2015, 6, 195–199. [Google Scholar] [CrossRef]
- Nieschulz, O. On the pharmacology of the active substances of betel. 1. Central effect of arecoline. Arzneimittelforschung 1967, 17, 1292–1297. [Google Scholar]
- Sharma, M.; Shetty, S.S.; Radhakrishnan, R. Oral Submucous Fibrosis as an Overhealing Wound: Implications in Malignant Transformation. Recent Patents Anti-Cancer Drug Discov. 2018, 13, 272–291. [Google Scholar] [CrossRef] [PubMed]
- Thangjam, G.S.; Kondaiah, P. Regulation of oxidative-stress responsive genes by arecoline in human keratinocytes. J. Periodontal Res. 2009, 44, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.-P.; Wu, K.-J.; Chen, H.-M.; Deng, Y.-T. Arecoline activates latent transforming growth factor β1 via mitochondrial reactive oxygen species in buccal fibroblasts: Suppression by epigallocatechin-3-gallate. J. Formos. Med. Assoc. 2018, 117, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Bale, R.; Kattappagari, K.K.; Vidya, D.; Vuddandi, S.; Gummalla, C.; Baddam, V.R.R. Oral submucous fibrosis: A quantitative assessment of serum malondialdehyde, superoxide dismutase and correlation with clinical staging. J. Oral Maxillofac. Pathol. 2017, 21, 41–45. [Google Scholar] [CrossRef] [Green Version]
- Radhakrishnan, R.; Shetty, S.S.; Sharma, M.; Kabekkodu, S.P.; Kumar, N.A.; Satyamoorthy, K. Understanding the molecular mechanism associated with reversal of oral submucous fibrosis targeting hydroxylysine aldehyde-derived collagen cross-links. J. Carcinog. 2021, 20, 9. [Google Scholar] [CrossRef]
- Jayasooriya, P.R.; Jayasinghe, K.A.N.; Tilakaratne, W.M. Relationship between thickness of fibrosis and epithelial dysplasia in oral submucous fibrosis. J. Investig. Clin. Dent. 2011, 2, 171–175. [Google Scholar] [CrossRef]
- Jeng, J.; Chang, M.; Hahn, L. Role of areca nut in betel quid-associated chemical carcinogenesis: Current awareness and future perspectives. Oral Oncol. 2001, 37, 477–492. [Google Scholar] [CrossRef]
- Campisi, J.; Andersen, J.K.; Kapahi, P.; Melov, S. Cellular senescence: A link between cancer and age-related degenerative disease? Semin. Cancer Biol. 2011, 21, 354–359. [Google Scholar] [CrossRef] [Green Version]
- Tilakaratne, W.M.; Iqbal, Z.; Teh, M.-T.; Ariyawardana, A.; Pitiyage, G.; Cruchley, A.; Stewart, J.E.; Hagi-Pavli, E.; Lalli, A.; Waseem, A.; et al. Upregulation of HIF-1α in malignant transformation of oral submucous fibrosis. J. Oral Pathol. Med. 2008, 37, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Nair, J.; Ohshima, H.; Friesen, M.; Croisy, A.; Bhide, S.; Bartsch, H. Tobacco-specific and betel nut-specific N-nitroso compounds: Occurrence in saliva and urine of betel quid chewers and formation in vitro by nitrosation of betel quid. Carcinogenesis 1985, 6, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.; Muthumala, M.; Matsuoka, H.; Uehara, O.; Kuramitsu, Y.; Chiba, I.; Abiko, Y. How Each Component of Betel Quid Is Involved in Oral Carcinogenesis: Mutual Interactions and Synergistic Effects with Other Carcinogens—A Review Article. Curr. Oncol. Rep. 2019, 21, 53. [Google Scholar] [CrossRef] [PubMed]
- Niaz, K.; Maqbool, F.; Khan, F.; Bahadar, H.; Hassan, F.I.; Abdollahi, M. Smokeless tobacco (paan and gutkha) consumption, prevalence, and contribution to oral cancer. Epidemiol. Health 2017, 39, e2017009. [Google Scholar] [CrossRef]
- Sharan, R.N.; Mehrotra, R.; Choudhury, Y.; Asotra, K. Association of Betel Nut with Carcinogenesis: Revisit with a Clinical Perspective. PLoS ONE 2012, 7, e42759. [Google Scholar] [CrossRef] [Green Version]
- Harvey, W.; Scutt, A.; Meghji, S.; Canniff, J. Stimulation of human buccal mucosa fibroblasts in vitro by betel-nut alkaloids. Arch. Oral Biol. 1986, 31, 45–49. [Google Scholar] [CrossRef]
- Krishnakumar, K.; Ramadoss, R.; Krishnan, R.; Sukhija, H. In vitro Quantification of Collagen and Snail1 Gene Expression in Experimentally Induced Fibrosis by Arecoline and Commercial Smokeless Tobacco Products. Asian Pac. J. Cancer Prev. 2020, 21, 1143–1148. [Google Scholar] [CrossRef] [Green Version]
- Xia, L.; Tian-You, L.; Yi-Jun, G.; Dong-sheng, T.; Wen-Hui, L. Arecoline and oral keratinocytes may affect the collagen metabolism of fibroblasts. J. Oral Pathol. Med. 2009, 38, 422–426. [Google Scholar] [CrossRef]
- Jeng, J.-H.; Lan, W.-H.; Hahn, L.-J.; Hsieh, C.-C.; Kuo, M.Y.-P. Inhibition of the migration, attachment, spreading, growth and collagen synthesis of human gingival fibroblasts by arecoline, a major areca alkaloid, in vitro. J. Oral Pathol. Med. 1996, 25, 371–375. [Google Scholar] [CrossRef]
- Nair, U.; Bartsch, H.; Nair, J. Alert for an epidemic of oral cancer due to use of the betel quid substitutes gutkha and pan masala: A review of agents and causative mechanisms. Mutagenesis 2004, 19, 251–262. [Google Scholar] [CrossRef] [Green Version]
- Sundqvist, K.; Liu, Y.; Nair, J.; Bartsch, H.; Arvidson, K.; Grafström, R.C. Cytotoxic and genotoxic effects of areca nut-related compounds in cultured human buccal epithelial cells. Cancer Res. 1989, 49, 5294–5298. [Google Scholar] [PubMed]
- Arakeri, G.; Brennan, P.A. Oral submucous fibrosis: An overview of the aetiology, pathogenesis, classification, and principles of management. Br. J. Oral Maxillofac. Surg. 2013, 51, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.C.; Ho, Y.S.; Lee, P.H.; Chan, C.P.; Lee, J.J.; Hahn, L.J.; Wang, Y.J.; Jeng, J.H. Areca nut extract and arecoline induced the cell cycle arrest but not apoptosis of cultured oral KB epithelial cells: Association of glutathione, reactive oxygen species and mitochondrial membrane potential. Carcinogenesis 2001, 22, 1527–1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, L.; Jian, X.; Guo, F.; Lian, Z.; Jiang, C.; Yin, P.; Min, A.-J.; Huang, L. miR-203 inhibits arecoline-induced epithelial-mesenchymal transition by regulating secreted frizzled-related protein 4 and transmembrane-4 L six family member 1 in oral submucous fibrosis. Oncol. Rep. 2015, 33, 2753–2760. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.-H.; Wang, Y.-P.; Chang, J.; Pan, Y.-H.; Chang, M.-C.; Jeng, J.-H. Genetic Susceptibility and Protein Expression of Extracellular Matrix Turnover-Related Genes in Oral Submucous Fibrosis. Int. J. Mol. Sci. 2020, 21, 8104. [Google Scholar] [CrossRef]
- Thomas, S.; MacLennan, R. Slaked lime and betel nut cancer in Papua New Guinea. Lancet 1992, 340, 577–578. [Google Scholar] [CrossRef]
- Sarode, S.C.; Mahuli, A.; Sarode, G.S.; Mahuli, S. Why only areca nut chewing cannot cause oral submucous fibrosis? Med. Hypotheses 2013, 81, 47–49. [Google Scholar] [CrossRef]
- Brew, K.; Nagase, H. The tissue inhibitors of metalloproteinases (TIMPs): An ancient family with structural and functional diversity. Biochim. Biophys. Acta BBA Mol. Cell Res. 2010, 1803, 55–71. [Google Scholar] [CrossRef] [Green Version]
- Sharan, R.N.; Choudhury, Y. Betel Nut and Susceptibility to Cancer. In Environmental Factors, Genes, and the Development of Human Cancers; Roy, D., Dorak, M.T., Eds.; Springer: New York, NY, USA, 2021; pp. 401–428. [Google Scholar]
- Chen, P.-H.; Mahmood, Q.; Mariottini, G.L.; Chiang, T.-A.; Lee, K.-W. Adverse Health Effects of Betel Quid and the Risk of Oral and Pharyngeal Cancers. BioMed Res. Int. 2017, 2017, 3904098. [Google Scholar] [CrossRef] [Green Version]
- Alexander, A.J.; Ramani, P.; Sherlin, H.J.; Gheena, S. Quantitative analysis of copper levels in areca nut plantation area—A role in increasing prevalence of oral submucous fibrosis: An In vitro study. Indian J. Dent. Res. 2019, 30, 261–266. [Google Scholar] [CrossRef]
- León-González, A.J.; Auger, C.; Schini-Kerth, V.B. Pro-oxidant activity of polyphenols and its implication on cancer chemoprevention and chemotherapy. Biochem. Pharmacol. 2015, 98, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Petti, S.; Scully, C. Polyphenols, oral health and disease: A review. J. Dent. 2009, 37, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Briguglio, G.; Costa, C.; Pollicino, M.; Giambò, F.; Catania, S.; Fenga, C. Polyphenols in cancer prevention: New insights (Review). Int. J. Funct. Nutr. 2020, 1, 9. [Google Scholar] [CrossRef]
- Babich, H.; Schuck, A.G.; Weisburg, J.H.; Zuckerbraun, H.L. Research Strategies in the Study of the Pro-Oxidant Nature of Polyphenol Nutraceuticals. J. Toxicol. 2011, 2011, 467305. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.C.; Yang, S.F.; Peng, C.Y.; Chou, M.Y.; Chang, Y.C. Epigallocatechin-3-gallate inhibits the invasion of humanoral cancer cells and decreases the productions of matrixmetalloproteinases and urokinase-plasminogen activator. J. Oral Pathol. Med. 2007, 36, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Khafif, A.; Schantz, S.P.; Al-Rawi, M.; Edelstein, D.; Sacks, P.G. Green tea regulates cell cycle progression in oralleukoplakia. Head Neck 1998, 20, 528–534. [Google Scholar] [CrossRef]
- Belobrov, S.; Seers, C.; Reynolds, E.; Cirillo, N.; McCullough, M. Functional and molecular effects of a green tea constituent on oral cancer cells. J. Oral Pathol. Med. 2019, 48, 604–610. [Google Scholar] [CrossRef]
- Rekha, V.; Kollipara, M.; Gupta, B.; Bharath, Y.; Pulicherla, K.K. A review on Piper betle L.: Nature’s promising medicinal reservoir. Am. J. Ethnomed. 2014, 1, 276–289. [Google Scholar]
- Mula, S.; Banerjee, D.; Patro, B.S.; Bhattacharya, S.; Barik, A.; Bandyopadhyay, S.K.; Chattopadhyay, S. Inhibitory property of the Piper betel phenolics against photosensitization-induced biological damages. Bioorg. Med. Chem. 2008, 16, 2932–2938. [Google Scholar] [CrossRef]
- Nagori, K.; Singh, M.K.; Alexander, A.; Kumar, T.; Dewangan, D.; Badwaik, H.; Tripathi, D. Piper betle L.: A review on its ethnobotany, phytochemistry, pharmacological profile and profiling by new hyphenated technique DART-MS (Direct Analysis in Real Time Mass Spectrometry). J. Pharm. Res. 2011, 4, 2991–2997. [Google Scholar]
- Pradhan, D.; Suri, K.; Pradhan, D.; Biswasroy, P. Golden heart of the nature: Piper betle L. J. Pharm. Phytochem. 2013, 1, 147–167. [Google Scholar]
- Rossi, M.; Garavello, W.; Talamini, R.; Negri, E.; Bosetti, C.; Maso, L.D.; Lagiou, P.; Tavani, A.; Polesel, J.; Barzan, L.; et al. Flavonoids and the Risk of Oral and Pharyngeal Cancer: A Case-Control Study from Italy. Cancer Epidemiol. Biomark. Prev. 2007, 16, 1621–1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iriti, M.; Varoni, E.M. Chemopreventive Potential of Flavonoids in Oral Squamous Cell Carcinoma in Human Studies. Nutrients 2013, 5, 2564–2576. [Google Scholar] [CrossRef]
- Johnson, T.L.; Lai, M.B.; Lai, J.C.K.; Bhushan, A. Inhibition of Cell Proliferation and MAP Kinase and Akt Pathways in Oral Squamous Cell Carcinoma by Genistein and Biochanin A. Evid.-Based Complement. Altern. Med. 2010, 7, 351–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silvan, S.; Manoharan, S.; Baskaran, N.; Anusuya, C.; Karthikeyan, S.; Prabhakar, M.M. Chemopreventive potential of apigenin in 7,12-dimethylbenz(a)anthracene induced experimental oral carcinogenesis. Eur. J. Pharmacol. 2011, 670, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Devi, P.U.; Ganasoundari, A. Modulation of glutathione and antioxidant enzymes by Ocimum sanctum and its role in protection against radiation injury. Indian J. Exp. Boil. 1999, 37, 262–268. [Google Scholar]
- Reshma, K.; Ashalatha, V.R.; Dinesh, M.; Vasudevan, D. Effect of ocimum flavonoids as a radioprotector on the erythrocyte antioxidants in oral cancer. Indian J. Clin. Biochem. 2005, 20, 160–164. [Google Scholar] [CrossRef] [Green Version]
- Morton, J.F. Further Associations of Plant Tannins and Human Cancer. Q. J. Crude Drug Res. 1972, 12, 1829–1841. [Google Scholar] [CrossRef]
- Panigrahi, G.; Rao, A. Study of the genotoxicity of the total aqueous extract of betel nut and its tannin. Carcinogenesis 1986, 7, 37–39. [Google Scholar] [CrossRef]
- Rajasekar, N.; Sivanantham, A.; Ravikumar, V.; Rajasekaran, S. An overview on the role of plant-derived tannins for the treatment of lung cancer. Phytochemistry 2021, 188, 112799. [Google Scholar] [CrossRef]
- Baer-Dubowska, W.; Szaefer, H.; Majchrzak-Celińska, A.; Krajka-Kuźniak, V. Tannic Acid: Specific Form of Tannins in Cancer Chemoprevention and Therapy-Old and New Applications. Curr. Pharmacol. Rep. 2020, 6, 28–37. [Google Scholar] [CrossRef] [Green Version]
- Ta, L.T.; Nguyen, T.T.K.; Yoo, H. Tannic acid-induced apoptosis in FaDu hypopharyngeal squamous cell carcinoma. Int. J. Oral Biol. 2019, 44, 43–49. [Google Scholar] [CrossRef]
- Darvin, P.; Baeg, S.J.; Joung, Y.H.; Sp, N.; Kang, D.Y.; Byun, H.J.; Park, J.U.; Yang, Y.M. Tannic acid inhibits the Jak2/STAT3 pathway and induces G1/S arrest and mitochondrial apoptosis in YD-38 gingival cancer cells. Int. J. Oncol. 2015, 47, 1111–1120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chattopadhyay, A.; Ray, J.G. Molecular Pathology of Malignant Transformation of Oral Submucous Fibrosis. J. Environ. Pathol. Toxicol. Oncol. 2016, 35, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Xing, G.; Qi, X.; Feng, C.; Liu, M.; Gong, L.; Luan, Y.; Ren, J. Assessment of the mutagenic potential of arecoline in gpt delta transgenic mice. Mutat. Res. Toxicol. Environ. Mutagen. 2012, 748, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Lai, K.-C.; Lee, T.-C. Genetic damage in cultured human keratinocytes stressed by long-term exposure to areca nut extracts. Mutat. Res. Mol. Mech. Mutagen. 2006, 599, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, E.; Omobono, J.D., II; Guo, Z.; Hopkinson, S.; Lazar, A.J.; Brenn, T.; Jones, J.C.; Rheinwald, J.G. A 318 keratinocyte hypermotility/growth-arrest response involving laminin 5 and p16INK4A activated in 319 wound healing and senescence. Am. J. Pathol. 2006, 168, 1821–1837. [Google Scholar] [CrossRef] [Green Version]
- Sharma, M.; Fonseca, F.P.; Hunter, K.D.; Radhakrishnan, R. Loss of oral mucosal stem cell markers in oral submucous fibrosis and their reactivation in malignant transformation. Int. J. Oral Sci. 2020, 12, 1–10. [Google Scholar] [CrossRef]
- Sharma, M.; Hunter, K.D.; Fonseca, F.P.; Radhakrishnan, R. Emerging role of cellular senescence in the pathogenesis of oral submucous fibrosis and its malignant transformation. Head Neck 2021, 43, 3153–3164. [Google Scholar] [CrossRef]
- Rehman, A.; Ali, S.; Lone, M.A.; Atif, M.; Hassona, Y.; Prime, S.S.; Pitiyage, G.N.; James, E.; Parkinson, E.K. Areca nut alkaloids induce irreparable DNA damage and senescence in fibroblasts and may create a favourable environment for tumour progression. J. Oral Pathol. Med. 2015, 45, 365–372. [Google Scholar] [CrossRef]
- Pitiyage, G.N.; Slijepcevic, P.; Gabrani, A.; Chianea, Y.G.; Lim, K.P.; Prime, S.S.; Tilakaratne, W.M.; Fortune, F.; Parkinson, E.K. Senescent mesenchymal cells accumulate in human fibrosis by a telomere-independent mechanism and ameliorate fibrosis through matrix metalloproteinases. J. Pathol. 2011, 223, 604–617. [Google Scholar] [CrossRef] [PubMed]
- Prime, S.S.; Cirillo, N.; Hassona, Y.; Lambert, D.W.; Paterson, I.C.; Mellone, M.; Thomas, G.J.; James, E.N.L.; Parkinson, E.K. Fibroblast activation and senescence in oral cancer. J. Oral Pathol. Med. 2016, 46, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.P.; Cirillo, N.; Hassona, Y.; Wei, W.; Thurlow, J.K.; Cheong, S.C.; Pitiyage, G.; Parkinson, E.K.; Prime, S.S. Fibroblast gene expression profile reflects the stage of tumour progression in oral squamous cell carcinoma. J. Pathol. 2011, 223, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Hassona, Y.; Cirillo, N.; Lim, K.P.; Herman, A.; Mellone, M.; Thomas, G.J.; Pitiyage, G.N.; Parkinson, E.; Prime, S.S. Progression of genotype-specific oral cancer leads to senescence of cancer-associated fibroblasts and is mediated by oxidative stress and TGF-β. Carcinogenesis 2013, 34, 1286–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassona, Y.; Cirillo, N.; Heesom, K.J.; Parkinson, E.K.; Prime, S.S. Senescent cancer-associated fibroblasts secrete active MMP-2 that promotes keratinocyte dis-cohesion and invasion. Br. J. Cancer 2014, 111, 1230–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cirillo, N.; Hassona, Y.; Celentano, A.; Lim, K.; Manchella, S.; Parkinson, E.; Prime, S. Cancer-associated fibroblasts regulate keratinocyte cell–cell adhesion via TGF-β-dependent pathways in genotype-specific oral cancer. Carcinogenesis 2017, 38, 76–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shetty, S.S.; Sharma, M.; Fonseca, F.P.; Jayaram, P.; Tanwar, A.S.; Kabekkodu, S.P.; Satyamoorthy, K.; Radhakrishnan, R. Signaling pathways promoting epithelial mesenchymal transition in oral submucous fibrosis and oral squamous cell carcinoma. Jpn. Dent. Sci. Rev. 2020, 56, 97–108. [Google Scholar] [CrossRef]
- Holst, C.R.; Nuovo, G.J.; Esteller, M.; Chew, K.; Baylin, S.B.; Herman, J.G.; Tlsty, T.D. Methylation of p16(INK4a) promoters occurs in vivo in histologically normal human mammary epithelia. Cancer Res. 2003, 63, 1596–1601. [Google Scholar]
- Freund, A.; Patil, C.K.; Campisi, J. p38MAPK is a novel DNA damage response-independent regulator of the senescence-associated secretory phenotype. EMBO J. 2011, 30, 1536–1548. [Google Scholar] [CrossRef] [Green Version]
- Song, S.; Tchkonia, T.; Jiang, J.; Kirkland, J.L.; Sun, Y. Targeting Senescent Cells for a Healthier Aging: Challenges and Opportunities. Adv. Sci. 2020, 7, 2002611. [Google Scholar] [CrossRef]
- Nambiar, A.; Justice, J.; Pascual, R.; Hashmi, S.; Tchkonia, T.; Lebrasseur, N.; Kirkland, J.; Musi, N.; Kritchevsky, S. B17. Senolytics in Idiopathic Pulmonary Fibrosis: Preliminary Results from a First-in-Human, Open-Label, Pilot Study. Adv. Ild Ther. 2019, 40, 554–563. [Google Scholar] [CrossRef]
- Ingle, E. Turmeric in the management of oral submucous fibrosis—A systematic review and meta-analysis. Int. J. Health Sci. 2020, 14, 41–46. [Google Scholar]
- Kashyap, R.R.; Kashyap, R.S. Herbal derivatives in the management of mouth opening in oral submucous fibrosis—A network meta-analysis. Oral Dis. 2021, 27, 1606–1615. [Google Scholar] [CrossRef] [PubMed]
- Chuang, S.-L.; Wang, C.-P.; Chen, M.-K.; Su, W.W.-Y.; Su, C.-W.; Chen, S.L.-S.; Chiu, S.Y.-H.; Fann, J.C.-Y.; Yen, A.M.-F. Malignant transformation to oral cancer by subtype of oral potentially malignant disorder: A prospective cohort study of Taiwanese nationwide oral cancer screening program. Oral Oncol. 2018, 87, 58–63. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Jain et al., 2017 [24] | Franke et al., 2015 [25] | Cao et al., 2019 [26] | Srimany et al., 2016 [27] | Shirname et al., 1983 [28] | Kadi et al., 2013 [29] | Tang et al., 2017 [30] | Lord et al., 2002 [31] | Adhikari et al., 2015 [32] | |
|---|---|---|---|---|---|---|---|---|---|
| Arecoline | |||||||||
| Arecaidine | |||||||||
| Guvacoline | |||||||||
| Guvacine | |||||||||
| Isoguvacine | |||||||||
| Acatechu A | |||||||||
| Acatechu B | |||||||||
| Homoarecoline | |||||||||
| Arecatemine A | |||||||||
| Arecatemine B | |||||||||
| Arecatemine C | |||||||||
| Flavonoids | |||||||||
| Tannins | |||||||||
| Carbohydrates | |||||||||
| Fatty acids | |||||||||
| Chavibetol |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, P.; Chua, N.Q.E.; Dang, S.; Davis, A.; Chong, K.W.; Prime, S.S.; Cirillo, N. Molecular Mechanisms of Malignant Transformation of Oral Submucous Fibrosis by Different Betel Quid Constituents—Does Fibroblast Senescence Play a Role? Int. J. Mol. Sci. 2022, 23, 1637. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031637
Zhang P, Chua NQE, Dang S, Davis A, Chong KW, Prime SS, Cirillo N. Molecular Mechanisms of Malignant Transformation of Oral Submucous Fibrosis by Different Betel Quid Constituents—Does Fibroblast Senescence Play a Role? International Journal of Molecular Sciences. 2022; 23(3):1637. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031637
Chicago/Turabian StyleZhang, Pangzhen, Nathaniel Quan En Chua, Simon Dang, Ashleigh Davis, Kah Wee Chong, Stephen S. Prime, and Nicola Cirillo. 2022. "Molecular Mechanisms of Malignant Transformation of Oral Submucous Fibrosis by Different Betel Quid Constituents—Does Fibroblast Senescence Play a Role?" International Journal of Molecular Sciences 23, no. 3: 1637. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031637