
Genome-Wide Analysis of BpYABs and Function Identification Involving in the Leaf and Silique Development in Transgenic Arabidopsis

Abstract
:1. Introduction
2. Results
2.1. Identification and Phylogenetic Analysis of the YABBY Gene Family in Paper Mulberry
2.2. Sequence Analysis and Structural Characterization of BpYAB Members
2.3. Analysis of Cis-Acting Elements in the Promoters of BpYAB Genes
2.4. The Conserved Motif Identification of BpYAB Family
2.5. Transactivation Activity and Subcellular Localization of the BpYAB Family
2.6. Expression Patterns of BpYAB Genes in Different Tissues
2.7. BpYAB3 and BpYAB5 Do Not Show a Polarity Distribution in the Leaf
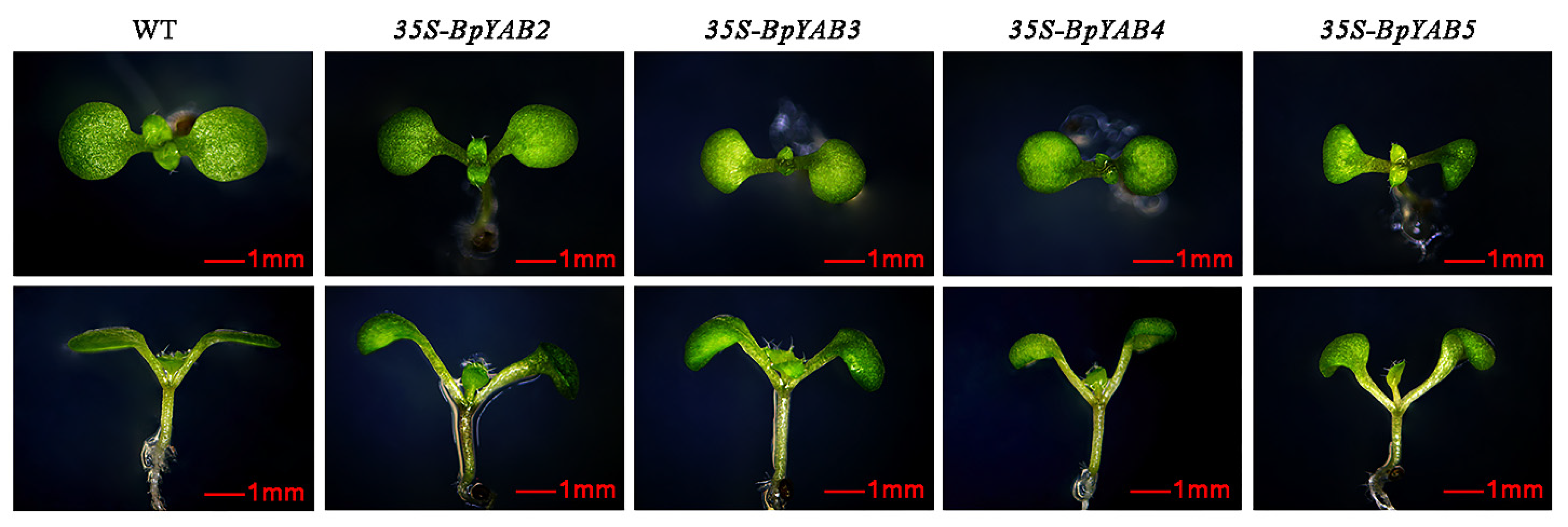
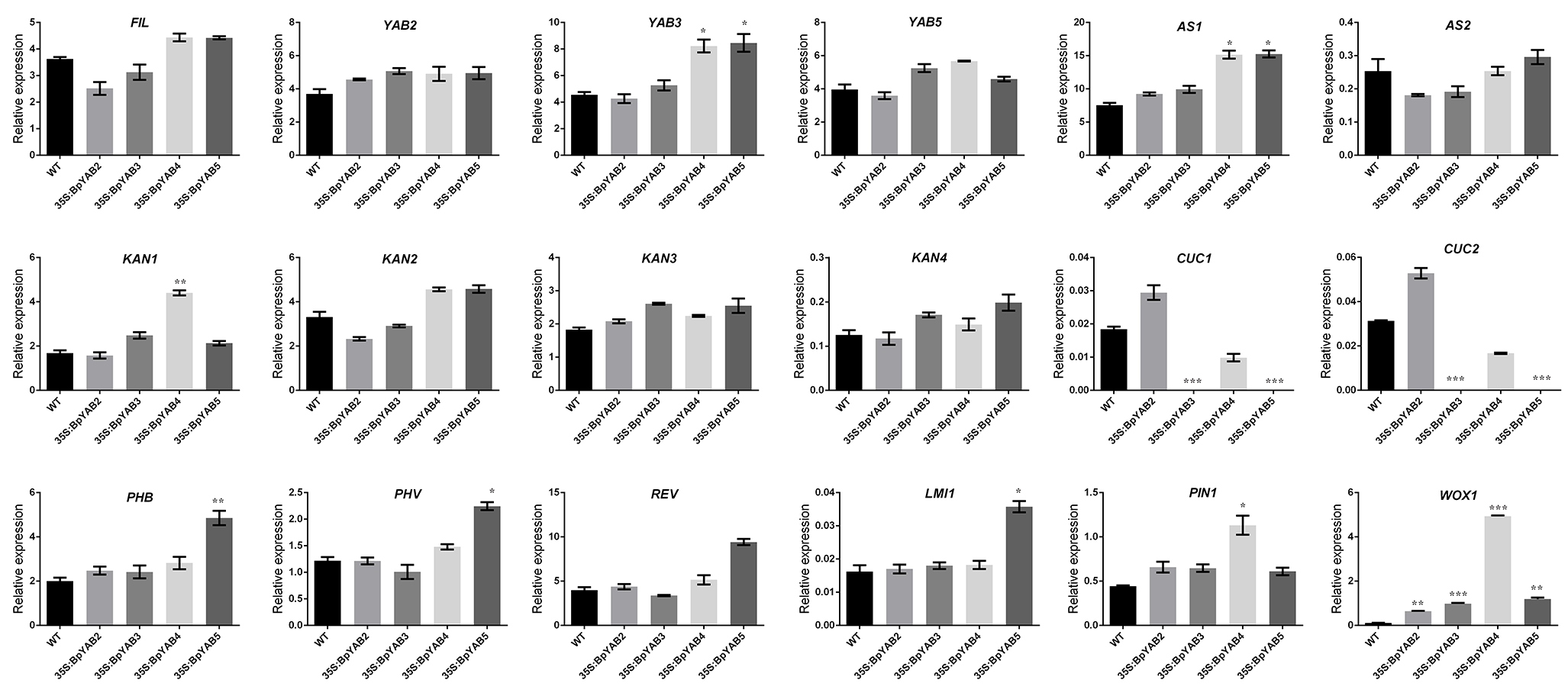
2.8. Ectopic Expression of BpYABs Affects Arabidopsis Leaf Development
2.9. Ectopic Expression of BpYABs Affects Arabidopsis Silique Length and Seed Size
3. Discussion
3.1. The Sequence Structures of YABBY Transcription Factors in Paper Mulberry Are Conserved
3.2. Three BpYAB Proteins Are Typical Transcription Factors
3.3. YAB Family Underwent Expansion in the FIL-like Clade
3.4. The Function of BpYAB Genes Was Related to the Evolutionary Relationship
3.5. BpYAB Did Not Show the Adaxial–Abaxial Polarity Distribution in Lamina
3.6. BpYABs Involved in Leaf and Silique Development
4. Materials and Methods
4.1. Identification and Gene Sequence Analysis of the BpYAB in Paper Mulberry
4.2. Phylogenetic Analysis and Conserved Motif Identification of BpYAB Proteins
4.3. Subcellular Localization and Transactivation Activity Assay of BpYAB Proteins
4.4. Plant Materials for qRT-PCR and RNA-Seq Data Analysis
4.5. In Situ Hybridization of BpYAB3 and BpYAB5
4.6. Agrobacterium-Mediated Transformation of Arabidopsis
4.7. The Morphological Observation of Transgenic Arabidopsis
4.8. The Statistical Analysis of Leaf Gene Expression in Arabidopsis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bowman, J.L. The YABBY gene family and abaxial cell fate. Curr. Opin. Plant Biol. 2000, 3, 17–22. [Google Scholar] [CrossRef]
- Gross, T.; Broholm, S.; Becker, A. CRABS CLAW acts as a bifunctional transcription factor in flower development. Front. Plant Sci. 2018, 9, 835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toriba, T.; Harada, K.; Takamura, A.; Nakamura, H.; Ichikawa, H.; Suzaki, T.; Hirano, H.Y. Molecular characterization the YABBY gene family in Oryza sativa and expression analysis of OsYABBY1. Mol. Genet. Genom. 2007, 277, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.; Hur, J.; Kim, S.J.; Han, M.J.; Kim, S.R. An GEctopic expression of OsYAB1 causes extra stamens and carpels in rice. Plant Mol. Biol. 2004, 56, 133–143. [Google Scholar] [CrossRef]
- Dai, M.; Zhao, Y.; Ma, Q.; Hu, Y.; Hedden, P.; Zhang, Q.; Zhou, D.X. The rice YABBY1 gene is involved in the feedback regulation of gibberellin metabolism. Plant Physiol. 2007, 144, 121–133. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.L.; Xu, Y.Y.; Xu, Z.H.; Chong, K. A rice YABBY gene, OsYABBY4, preferentially expresses in developing vascular tissue. Dev. Genes Evol. 2007, 217, 629–637. [Google Scholar] [CrossRef]
- Lv, S.; Wu, W.; Wang, M.; Meyer, R.S.; Ndjiondjop, M.N.; Tan, L.; Zhou, H.; Zhang, J.; Fu, Y.; Cai, H.; et al. Genetic control of seed shattering during African rice domestication. Nat. Plants. 2018, 4, 331–337. [Google Scholar] [CrossRef]
- Xiang, J.; Liu, R.Q.; Li, T.M.; Han, L.J.; Zou, Y.; Xu, T.F.; Wei, J.Y.; Wang, Y.J.; Xu, Y. Isolation and characterization of two VpYABBY genes from wild Chinese Vitis pseudoreticulata. Protoplasma 2013, 250, 1315–1325. [Google Scholar] [CrossRef]
- Sun, X.; Guan, Y.; Hu, X. Isolation and characterization of IaYABBY2 gene from Incarvillea arguta. Plant Mol. Biol. Rep. 2014, 32, 1219–1227. [Google Scholar] [CrossRef]
- Zhang, X.L.; Yang, Z.P.; Zhang, J.; Zhang, L.G. Ectopic expression of BraYAB1-702, a member of YABBY gene family in Chinese cabbage, causes leaf curling, inhibition of development of shoot apical meristem and flowering stage delaying in Arabidopsis thaliana. Int. J. Mol. Sci. 2013, 14, 14872–14891. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Reddy, V.A.; Panicker, D.; Mao, H.Z.; Kumar, N.; Rajan, C.; Venkatesh, P.N.; Chua, N.H.; Sarojam, R. Metabolic engineering of terpene biosynthesis in plants using a trichome-specific transcription factor MsYABBY5 from spearmint (Mentha spicata). Plant Biotechnol. J. 2016, 14, 1619–1632. [Google Scholar] [CrossRef] [Green Version]
- Finet, C.; Floyd, S.K.; Conway, S.J.; Zhong, B.; Scutt, C.P.; Bowman, J.L. Evolution of the YABBY gene family in seed plants. Evol. Dev. 2016, 18, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Eckardt, N.A. YABBY genes and the development and origin of seed plant leaves. Plant Cell 2010, 22, 2103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowman, J.L.; Smyth, D.R. Crabs Claw, a gene that regulates carpel and nectary development in Arabidopsis, encodes a novel protein with zinc finger and helix-loop-helix domains. Development 1999, 126, 2387–2396. [Google Scholar] [CrossRef] [PubMed]
- Simon, M.K.; Skinner, D.J.; Gallagher, T.L.; Gasser, C.S. Integument development in Arabidopsis depends on interaction of YABBY protein INNER NO OUTER with coactivators and corepressors. Genetics 2017, 207, 1489–1500. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Atkinson, A.; Otsuga, D.; Christensen, T.; Reynolds, L.; Drews, G.N. The Arabidopsis Filamentous flower gene is required for flower formation. Development 1999, 126, 2715. [Google Scholar] [CrossRef] [PubMed]
- Bonaccorso, O.; Lee, J.E.; Puah, L.; Scutt, C.P.; Golz, J.F. Filamentous flower controls lateral organ development by acting as both an activator and a repressor. BMC Plant Biol. 2012, 12, 176. [Google Scholar] [CrossRef] [Green Version]
- Sarojam, R.; Sappl, P.G.; Goldshmidt, A.; Efroni, I.; Floyd, S.K.; Eshed, Y.; Bowman, J.L. Differentiating Arabidopsis shoots from leaves by combined YABBY activities. Plant Cell 2010, 22, 2113–2130. [Google Scholar] [CrossRef] [Green Version]
- Goldshmidt, A.; Alvarez, J.P.; Bowman, J.L.; Eshed, Y. Signals derived from YABBY gene activities in organ primordia regulate growth and partitioning of Arabidopsis shoot apical meristems. Plant Cell 2008, 20, 1217–1230. [Google Scholar] [CrossRef] [Green Version]
- Golz, J.F.; Roccaro, M.; Kuzoff, R.; Hudson, A. Graminifolia promotes growth and polarity of Antirrhinum leaves. Development 2004, 131, 3661–3670. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, W.; Toriba, T.; Hirano, H.Y. Three TOB1-related YABBY genes are required to maintain proper function of the spikelet and branch meristems in rice. New Phytol. 2017, 215, 825–839. [Google Scholar] [CrossRef] [Green Version]
- Ohmori, Y.; Toriba, T.; Nakamura, H.; Ichikawa, H.; Hirano, H.Y. Temporal and spatial regulation of Drooping leaf gene expression that promotes midrib formation in rice. Plant J. 2011, 65, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Juarez, M.T.; Twigg, R.W.; Timmermans, M.C. Specification of adaxial cell fate during maize leaf development. Development 2004, 131, 4533–4544. [Google Scholar] [CrossRef] [Green Version]
- Olsen, K.M. One gene’s shattering effects. Nat. Genet. 2012, 44, 616–617. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.J.; Shen, S.H. The paper mulberry: A novel model system for woody Chinese. Chinese Bull. Bot. 2018, 53, 372–381. [Google Scholar]
- Peng, X.; Liu, H.; Chen, P.; Tang, F.; Hu, Y.; Wang, F.; Pi, Z.; Zhao, M.; Chen, N.; Chen, H.; et al. A chromosome-scale genome assembly of paper mulberry (B. papyrifera) provides new insights into its forage and papermaking usage. Mol. Plant. 2019, 12, 661–677. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Wu, Q.; Teng, L.; Tang, F.; Pi, Z.; Shen, S. Transcriptional regulation of the paper mulberry under cold stress as revealed by a comprehensive analysis of transcription factors. BMC Plant Biol. 2015, 15, 108. [Google Scholar] [CrossRef] [Green Version]
- Tang, F.; Chen, N.; Zhao, M.; Wang, Y.; He, R.; Peng, X.; Shen, S. Identification and functional divergence analysis of WOX gene family in paper mulberry. Int. J. Mol. Sci. 2017, 18, 1782. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Liu, S.; Sun, T.; Liu, Z.; Tao, J.; Zhao, Y.; Wang, A. Bioinformatics analysis on YABBY gene family in tomato. J. Northeast Agric. Univ. 2017, 48, 11–19. [Google Scholar]
- Zhao, S.P.; Lu, D.; Yu, T.F.; Ji, Y.J.; Zheng, W.J.; Zhang, S.X.; Chai, S.C.; Chen, Z.Y.; Cui, X.Y. Genome-wide analysis of the YABBY family in soybean and functional identification of GmYABBY10 involvement in high salt and drought stresses. Plant Physiol. Bioch. 2017, 119, 132–146. [Google Scholar] [CrossRef]
- Yang, Z.; Gong, Q.; Wang, L.; Jin, Y.; Xi, J.; Li, Z.; Qin, W.; Yang, Z.; Lu, L.; Chen, Q.; et al. Genome-wide study of YABBY genes in upland cotton and their expression patterns under different stresses. Front. Genet. 2018, 9, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villanueva, J.M.; Broadhvest, J.; Hauser, B.A.; Meister, R.J.; Schneitz, K.; Gasser, C.S. INNER NO OUTER regulates abaxial-adaxial patterning in Arabidopsis ovules. Genes Dev. 1999, 13, 3160–3169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boter, M.; Golz, J.F.; Gimenez-Ibanez, S.; Fernandez-Barbero, G.; Franco-Zorrilla, J.M.; Solano, R. Filamentous flower is a direct target of JAZ3 and modulates responses to jasmonate. Plant Cell. 2015, 27, 3160–3174. [Google Scholar] [CrossRef] [Green Version]
- Bartholmes, C.; Hidalgo, O.; Gleissberg, S. Evolution of the YABBY gene family with emphasis on the basal eudicot Eschscholzia californica (Papaveraceae). Plant Biol. 2012, 14, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Eshed, Y.; Izhaki, A.; Baum, S.F.; Floyd, S.K.; Bowman, J.L. Asymmetric leaf development and blade expansion in Arabidopsis are mediated by KANADI and YABBY activities. Development 2004, 131, 2997–3006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kidner, C.A.; Timmermans, M.C.P. Signaling sides: Adaxial-abaxial patterning in leaves. Curr. Top. Dev. Biol. 2010, 91, 141–168. [Google Scholar] [PubMed]
- Sawa, S.; Watanabe, K.; Goto, K.; Kanaya, E.; Morita, E.H.; Okada, K. Filamentous flower, a meristem and organ identity gene of Arabidopsis, encodes a protein with a zinc finger and HMG-related domains. Genes Dev. 1999, 13, 1079–1088. [Google Scholar] [CrossRef] [Green Version]
- Dai, M.; Hu, Y.; Zhao, Y.; Liu, H.; Zhou, D.X. A Wuschel-like Homeobox gene represses a YABBY gene expression required for rice leaf development. Plant Physiol. 2007, 144, 380–390. [Google Scholar] [CrossRef] [Green Version]
- Xia, M.L.; Tang, D.Y.; Yang, Y.Z.; Li, Y.X.; Wang, W.W.; Lu, H.; Liu, X.M.; Lin, J.Z. Preliminary study on the rice OsYABBY6 gene involving in the regulation of leaf development. Life Sci. Res. 2017, 21, 23–30. [Google Scholar]
- Kamiuchi, Y.; Yamamoto, K.; Furutani, M.; Tasaka, M.; Aida, M. The CUC1 and CUC2 genes promote carpel margin meristem formation during Arabidopsis gynoecium development. Front Plant Sci. 2014, 5, 165. [Google Scholar] [CrossRef] [Green Version]
- Cucinotta, M.; Manrique, S.; Cuesta, C.; Benkova, E.; Novak, O.; Colombo, L. Cup-Shaped Cotyledon1 (CUC1) and CUC2 regulate cytokinin homeostasis to determine ovule number in Arabidopsis. J. Exp. Bot. 2018, 69, 5169–5176. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Xie, Y.; Ma, J.; Luo, X.; Nie, P.; Zuo, Z.; Lahrmann, U.; Zhao, Q.; Zheng, Y.; Zhao, Y.; et al. IBS: An illustrator for the presentation and visualization of biological sequences. Bioinformatics 2015, 31, 3359–3361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kouchi, H.; Hata, S. Isolation and characterization of novel nodulin cDNAs representing genes expressed at early stages of soybean nodule development. Mol. Genet. Genom. 1993, 238, 106–119. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene ID | Chr. | Genomic Location | Direction | AA | MW(Da) | pI |
|---|---|---|---|---|---|---|---|
| BpYAB1 | Bp0940004 | scaffold094 | 52,363–54,474 | - | 180 | 20,070 | 9.32 |
| BpYAB2 | Bp09g0299 | 9 | 4,247,279–4,253,760 | reverse | 190 | 21,027 | 8.69 |
| BpYAB3 | Bp06g0033 | 6 | 408,527–411,063 | reverse | 222 | 24,558 | 7.77 |
| BpYAB4 | Bp01g1037 | 1 | 3,368,630–3,370,433 | forward | 226 | 25,048 | 9.04 |
| BpYAB5 | Bp1460001 | scaffold146 | 32,046–36,003 | - | 194 | 21,896 | 8.77 |
| BpYAB6 | Bp07g0370 | 7 | 12,253,864–12,256,651 | forward | 179 | 20,222 | 4.97 |
| Cis-Element | Function | BpYAB1 | BpYAB2 | BpYAB3 | BpYAB4 | BpYAB5 | BpYAB6 | |
|---|---|---|---|---|---|---|---|---|
| Development | CCAAT-box | MYBHv1 binding site | ◆ | ◆ | ||||
| O2-site | zeatin metabolism regulation | ◆ | ◆ | ◆ | ◆ | |||
| RY-element | seed-specific regulation | ◆ | ||||||
| MSA-like | cell cycle regulation | ◆ | ||||||
| Hormone | ABRE | abscisic acid responsiveness | ◆ | ◆ | ◆ | |||
| AuxRR-core | auxin responsiveness | ◆ | ||||||
| P-box | gibberellin-responsive | ◆ | ||||||
| TATC-box | gibberellin-responsiveness | ◆ | ◆ | ◆ | ||||
| TCA-element | salicylic acid responsiveness | ◆ | ◆ | |||||
| TGA-element | auxin-responsive | ◆ | ||||||
| Environment | ARE | the anaerobic induction | ◆ | ◆ | ◆ | ◆ | ◆ | |
| LTR | low-temperature responsiveness | ◆ | ||||||
| MBS | drought-inducibility | ◆ | ◆ | |||||
| WUN-motif | wound-responsive | ◆ | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, F.; Zhang, D.; Chen, N.; Peng, X.; Shen, S. Genome-Wide Analysis of BpYABs and Function Identification Involving in the Leaf and Silique Development in Transgenic Arabidopsis. Int. J. Mol. Sci. 2022, 23, 1670. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031670
Tang F, Zhang D, Chen N, Peng X, Shen S. Genome-Wide Analysis of BpYABs and Function Identification Involving in the Leaf and Silique Development in Transgenic Arabidopsis. International Journal of Molecular Sciences. 2022; 23(3):1670. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031670
Chicago/Turabian StyleTang, Feng, Dan Zhang, Naizhi Chen, Xianjun Peng, and Shihua Shen. 2022. "Genome-Wide Analysis of BpYABs and Function Identification Involving in the Leaf and Silique Development in Transgenic Arabidopsis" International Journal of Molecular Sciences 23, no. 3: 1670. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031670