
Immunofluorescence and High-Resolution Microscopy Reveal New Insights in Human Globozoospermia

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
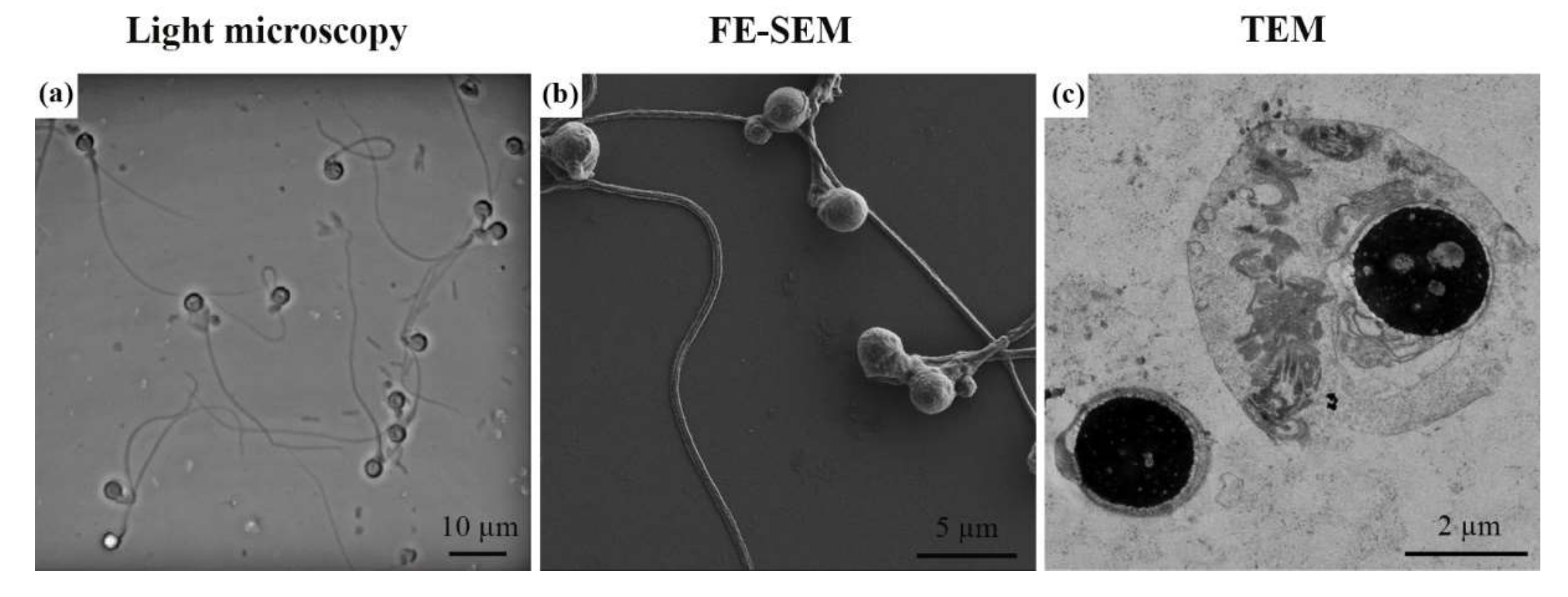
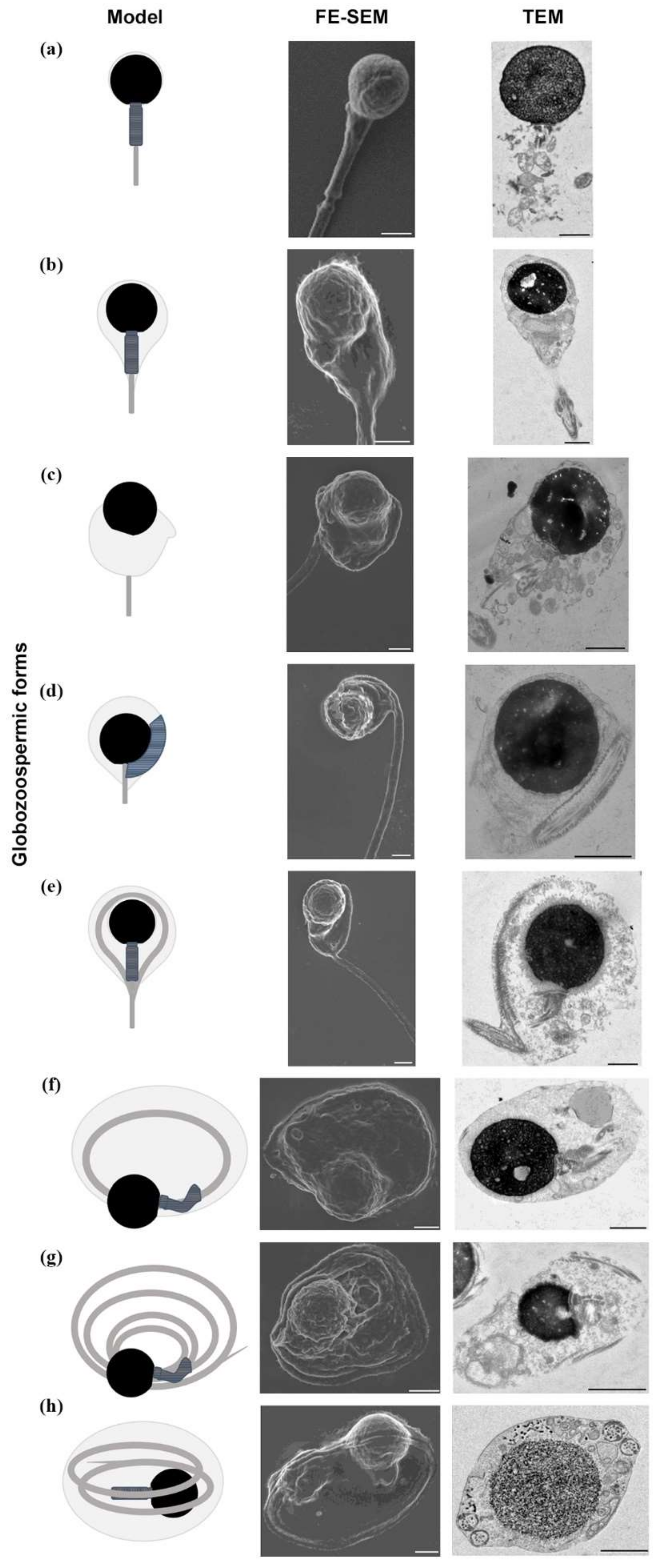
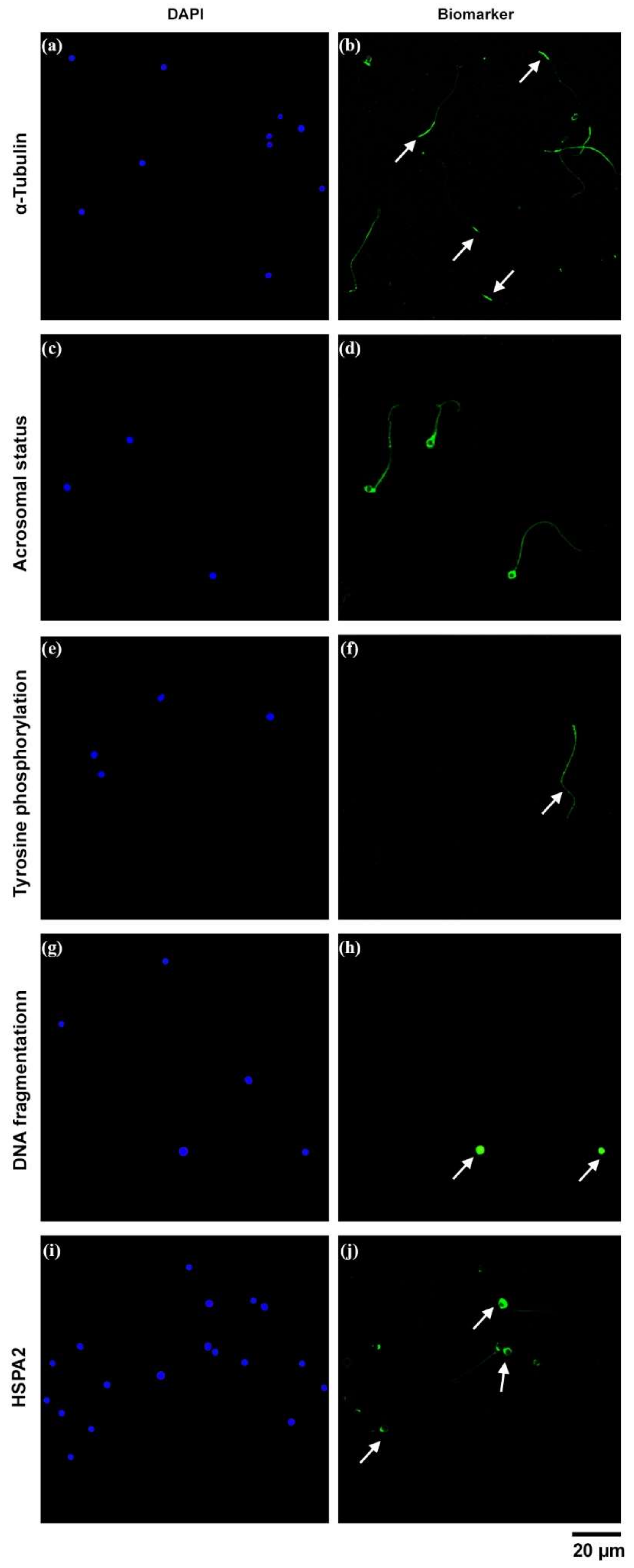
2. Results
3. Discussion
4. Materials and Methods
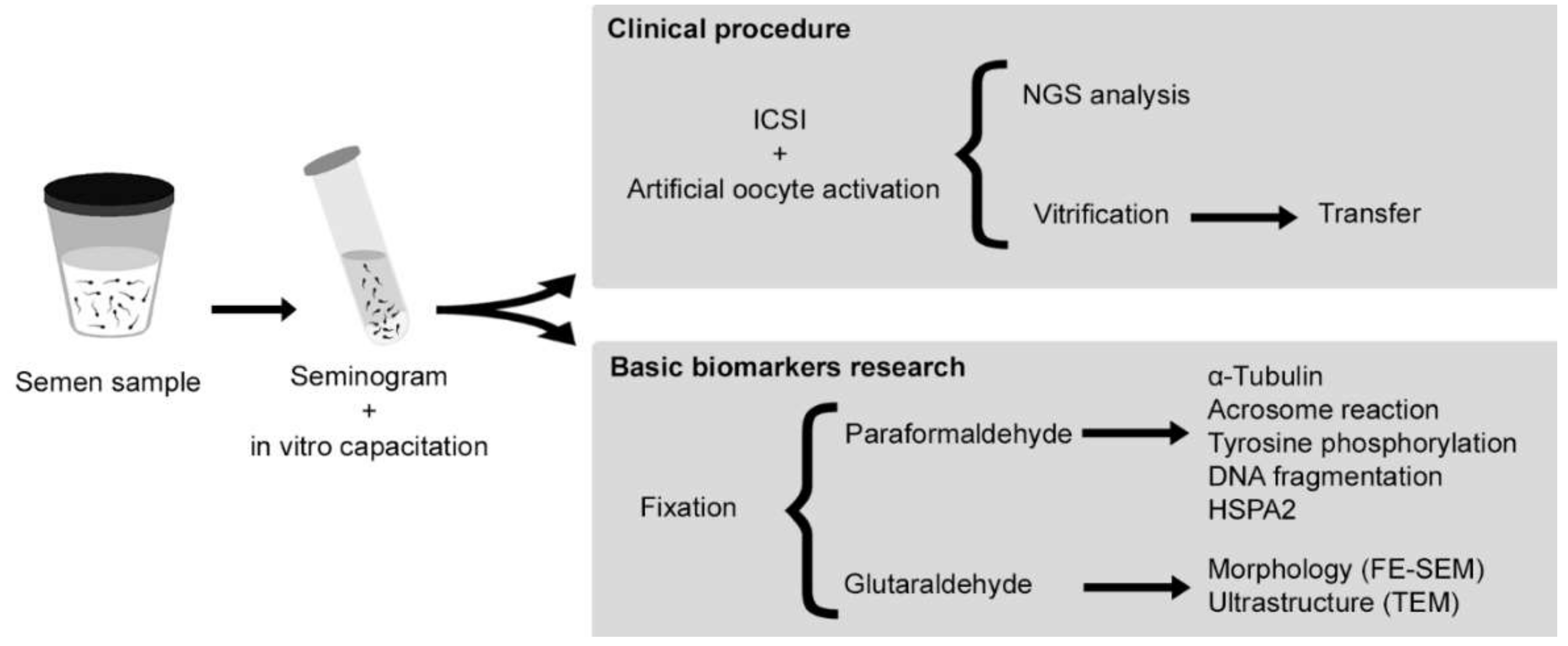
4.1. Experimental Design
4.2. Patient and Assisted Reproductive Technologies (ART)
4.3. Fixation
4.4. Morphological and Ultrastructural Characterization
4.5. Functional and Molecular Aspects
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sen, C.G.S.; Holstein, A.F.; Schirren, C. Über Die Morphogenese Rundköpfiger Spermatozoen Des Menschen. Andrologia 1971, 3, 117–125. [Google Scholar] [CrossRef]
- Fesahat, F.; Henkel, R.; Agarwal, A. Globozoospermia Syndrome: An Update. Andrologia 2020, 52, e13459. [Google Scholar] [CrossRef] [PubMed]
- Dam, A.H.D.M.; Feenstra, I.; Westphal, J.R.; Ramos, L.; van Golde, R.J.T.; Kremer, J.A.M. Globozoospermia Revisited. Hum. Reprod. Update 2007, 13, 63–75. [Google Scholar] [CrossRef] [Green Version]
- Singh, G. Ultrastructural Features of Round-Headed Human Spermatozoa. Int. J. Fertil. 1992, 37, 99–102. [Google Scholar] [PubMed]
- Han, F.; Liu, C.; Zhang, L.; Chen, M.; Zhou, Y.; Qin, Y.; Wang, Y.; Chen, M.; Duo, S.; Cui, X.; et al. Globozoospermia and Lack of Acrosome Formation in GM130-Deficient Mice. Cell Death Dis. 2018, 8, e2532. [Google Scholar] [CrossRef] [PubMed]
- Baccetti, B.; Collodel, G.; Piomboni, P. Apoptosis in Human Ejaculated Sperm Cells (Notulae Seminologicae 9). J. Submicrosc. Cytol. Pathol. 1996, 28, 587–596. [Google Scholar] [PubMed]
- Vicari, E.; Perdichizzi, A.; De Palma, A.; Burrello, N.; D’Agata, R.; Calogero, A.E. Globozoospermia Is Associated with Chromatin Structure Abnormalities: Case Report. Hum. Reprod. 2002, 17, 2128–2133. [Google Scholar] [CrossRef] [Green Version]
- Wollina, U.; Schreiber, G.; Hipler, C. Lectin-Binding Sites in Testis of Men with Acrosomeless Round-Headed Spermatozoa. Andrologia 1989, 21, 271–274. [Google Scholar] [CrossRef]
- Sha, Y.-W.; Mei, L.-B.; Zheng, L.-K.; Tian, R.-H.; Ding, L.; Ji, Z.-Y.; Zhang, Q.; Li, P. Globozoospermic Infertility Associated with Balanced DPY19L2 Translocation/Gene Deletion at the Chromosomal Breakpoint. Asian J. Androl. 2018, 20, 210–211. [Google Scholar] [CrossRef] [PubMed]
- Vozdova, M.; Rybar, R.; Kloudova, S.; Prinosilova, P.; Texl, P.; Rubes, J. Total Globozoospermia Associated with Increased Frequency of Immature Spermatozoa with Chromatin Defects and Aneuploidy: A Case Report. Andrologia 2014, 46, 831–836. [Google Scholar] [CrossRef]
- Eskandari, N.; Tavalaee, M.; Zohrabi, D.; Nasr-Esfahani, M.H. Association between Total Globozoospermia and Sperm Chromatin Defects. Andrologia 2018, 50, e12843. [Google Scholar] [CrossRef]
- Wolenski, J.S.; Hart, N.H. Scanning Electron Microscope Studies of Sperm Incorporation into the Zebrafish (Brachydanio) Egg. J. Exp. Zool. 1987, 243, 259–273. [Google Scholar] [CrossRef] [PubMed]
- Rubino, P.; Viganò, P.; Luddi, A.; Piomboni, P. The ICSI Procedure from Past to Future: A Systematic Review of the More Controversial Aspects. Hum. Reprod. Update 2016, 22, 194–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawe, V.Y.; Galaverna, G.D.; Acosta, A.A.; Olmedo, S.B.; Chemes, H.E. Incidence of Tail Structure Distortions Associated with Dysplasia of the Fibrous Sheath in Human Spermatozoa. Hum. Reprod. 2001, 16, 879–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motiei, M.; Tavalaee, M.; Rabiei, F.; Hajihosseini, R.; Nasr-Esfahani, M.-H. Evaluation of HSPA2 in Fertile and Infertile Individuals. Andrologia 2013, 45, 66–72. [Google Scholar] [CrossRef]
- Buffone, M.G.; Doncel, G.F.; Marín Briggiler, C.I.; Vazquez-Levin, M.H.; Calamera, J.C. Human Sperm Subpopulations: Relationship between Functional Quality and Protein Tyrosine Phosphorylation. Hum. Reprod. (Oxf. Engl.) 2004, 19, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Dam, A.H.; Ramos, L.; Dijkman, H.B.; Woestenenk, R.; Robben, H.; van den Hoven, L.; Kremer, J.A. Morphology of Partial Globozoospermia. J. Androl. 2011, 32, 199–206. [Google Scholar] [CrossRef]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen; WHO: Geneve, Switzerland, 2010. [Google Scholar]
- Karaca, N.; Akpak, Y.K.; Oral, S.; Durmus, T.; Yilmaz, R. A Successful Healthy Childbirth in a Case of Total Globozoospermia with Oocyte Activation by Calcium Ionophore. J. Reprod. Infertil. 2015, 16, 116–120. [Google Scholar]
- Sermondade, N.; Hafhouf, E.; Dupont, C.; Bechoua, S.; Palacios, C.; Eustache, F.; Poncelet, C.; Benzacken, B.; Lévy, R.; Sifer, C. Successful Childbirth after Intracytoplasmic Morphologically Selected Sperm Injection without Assisted Oocyte Activation in a Patient with Globozoospermia. Hum. Reprod. (Oxf. Engl.) 2011, 26, 2944–2949. [Google Scholar] [CrossRef] [Green Version]
- Ghasemzadeh, J.; Talebi, A.R.; Khalili, M.A.; Fesahat, F.; Halvaei, I.; Nabi, A.; Ashourzadeh, S. Sperm Parameters, Protamine Deficiency, and Apoptosis in Total Globozoospermia. Iran. J. Reprod. Med. 2015, 13, 495–502. [Google Scholar]
- Hosseinifar, H.; Yazdanikhah, S.; Modarresi, T.; Totonchi, M.; Sadighi Gilani, M.A.; Sabbaghian, M. Correlation between Sperm DNA Fragmentation Index and CMA3 Positive Spermatozoa in Globozoospermic Patients. Andrology 2015, 3, 526–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talebi, A.R.; Ghasemzadeh, J.; Khalili, M.A.; Halvaei, I.; Fesahat, F. Sperm Chromatin Quality and DNA Integrity in Partial versus Total Globozoospermia. Andrologia 2018, 50, e12823. [Google Scholar] [CrossRef]
- Tavalaee, M.; Nomikos, M.; Lai, F.A.; Nasr-Esfahani, M.H. Expression of Sperm PLCζ and Clinical Outcomes of ICSI-AOA in Men Affected by Globozoospermia Due to DPY19L2 Deletion. Reprod. Biomed. Online 2018, 36, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Moretti, E.; Collodel, G.; Salvatici, M.C.; Belmonte, G.; Signorini, C. New Insights into Sperm with Total Globozoospermia: Increased Fatty Acid Oxidation and Centrin1 Alteration. Syst. Biol. Reprod. Med. 2019, 65, 390–399. [Google Scholar] [CrossRef]
- Ricci, G.; Andolfi, L.; Zabucchi, G.; Luppi, S.; Boscolo, R.; Martinelli, M.; Zweyer, M.; Trevisan, E. Ultrastructural Morphology of Sperm from Human Globozoospermia. BioMed Res. Int. 2015, 2015, 798754. [Google Scholar] [CrossRef] [Green Version]
- Yao, R.; Ito, C.; Natsume, Y.; Sugitani, Y.; Yamanaka, H.; Kuretake, S.; Yanagida, K.; Sato, A.; Toshimori, K.; Noda, T. Lack of Acrosome Formation in Mice Lacking a Golgi Protein, GOPC. Proc. Nat. Acad. Sci. USA 2002, 99, 11211–11216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francou, M.M.; Ten, J.; Bernabeu, R.; De Juan, J. Capacitation and Acrosome Reaction Changes α-Tubulin Immunodistribution in Human Spermatozoa. Reprod. BioMed. Online 2014, 28, 246–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sáez-Espinosa, P.; Ferrández-Rives, M.; Huerta-Retamal, N.; Robles-Gómez, L.; Aizpurua, J.; Romero, A.; Gómez-Torres, M.J. Proper Cytoskeleton α-Tubulin Distribution Is Concomitant to Tyrosine Phosphorylation during In Vitro Capacitation and Acrosomal Reaction in Human Spermatozoa. Cytoskeleton 2020, 77, 333–341. [Google Scholar] [CrossRef]
- Salvolini, E.; Buldreghini, E.; Lucarini, G.; Vignini, A.; Lenzi, A.; Di Primio, R.; Balercia, G. Involvement of Sperm Plasma Membrane and Cytoskeletal Proteins in Human Male Infertility. Fertil. Steril. 2013, 99, 697–704. [Google Scholar] [CrossRef]
- Carrell, D.T.; Emery, B.R.; Liu, L. Characterization of Aneuploidy Rates, Protamine Levels, Ultrastructure, and Functional Ability of Round-Headed Sperm from Two Siblings and Implications for Intracytoplasmic Sperm Injection. Fertil. Steril. 1999, 71, 511–516. [Google Scholar] [CrossRef]
- Deemeh, M.R.; Tavalaee, M.; Shahnaz, R.; Nasr-Esfahani, M.H. Evaluation of Protamine Deficiency and DNA Fragmentation in Two Globozoospermia Patients Undergoing ICSI. Int. J. Fertil. Steril. 2007, 1, 85–88. [Google Scholar] [CrossRef]
- Tavalaee, M.; Nasr-Esfahani, M.H. Expression Profile of PLCζ, PAWP, and TR-KIT in Association with Fertilization Potential, Embryo Development, and Pregnancy Outcomes in Globozoospermic Candidates for Intra-Cytoplasmic Sperm Injection and Artificial Oocyte Activation. Andrology 2016, 4, 850–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrin, A.; Coat, C.; Nguyen, M.H.; Talagas, M.; Morel, F.; Amice, J.; De Braekeleer, M. Molecular Cytogenetic and Genetic Aspects of Globozoospermia: A Review. Andrologia 2013, 45, 1–9. [Google Scholar] [CrossRef]
- Larson, K.L.; Brannian, J.D.; Singh, N.P.; Burbach, J.A.; Jost, L.K.; Hansen, K.P.; Kreger, D.O.; Evenson, D.P. Chromatin Structure in Globozoospermia: A Case Report. J. Androl. 2001, 22, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Perrin, A.; Louanjli, N.; Ziane, Y.; Louanjli, T.; Le Roy, C.; Gueganic, N.; Amice, V.; De Braekeleer, M.; Morel, F. Study of Aneuploidy and DNA Fragmentation in Gametes of Patients with Severe Teratozoospermia. Reprod. Biomed. Online 2011, 22, 148–154. [Google Scholar] [CrossRef] [Green Version]
- Brahem, S.; Mehdi, M.; Elghezal, H.; Saad, A. Analysis of Sperm Aneuploidies and DNA Fragmentation in Patients with Globozoospermia or with Abnormal Acrosomes. Urology 2011, 77, 1343–1348. [Google Scholar] [CrossRef]
- Haghighat, S.; Tavalaee, M.; Kouhkan, A.; Zakeri, Z.; Noureddini, M.; Shahverdi, A.H.; Nasr Esfahani, M.H. Reduction of Truncated Kit Expression in Men with Abnormal Semen Parameters, Globozoospermia and History of Low or Fertilization Failure. Cell J. 2019, 21, 314–321. [Google Scholar] [CrossRef]
- De Braekeleer, M.; Nguyen, M.H.; Morel, F.; Perrin, A. Genetic Aspects of Monomorphic Teratozoospermia: A Review. J. Assist. Reprod. Genet. 2015, 32, 615–623. [Google Scholar] [CrossRef] [Green Version]
- Huerta-Retamal, N.; Sáez-Espinosa, P.; Robles-Gómez, L.; Avilés, M.; Romero, A.; Aizpurua, J.; Gómez-Torres, M.J. Human Sperm Chaperone HSPA2 Distribution during in Vitro Capacitation. J. Reprod. Immunol. 2020, 143, 103246. [Google Scholar] [CrossRef]
- Cayli, S.; Sakkas, D.; Vigue, L.; Demir, R.; Huszar, G. Cellular Maturity and Apoptosis in Human Sperm: Creatine Kinase, Caspase-3 and Bcl-XL Levels in Mature and Diminished Maturity Sperm. Mol. Hum. Reprod. 2004, 10, 365–372. [Google Scholar] [CrossRef] [Green Version]
- Kovanci, E.; Kovacs, T.; Moretti, E.; Vigue, L.; Bray-Ward, P.; Ward, D.C.; Huszar, G. FISH Assessment of Aneuploidy Frequencies in Mature and Immature Human Spermatozoa Classified by the Absence or Presence of Cytoplasmic Retention. Hum. Reprod. 2001, 16, 1209–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huszar, G.; Ozenci, C.C.; Cayli, S.; Zavaczki, Z.; Hansch, E.; Vigue, L. Hyaluronic Acid Binding by Human Sperm Indicates Cellular Maturity, Viability, and Unreacted Acrosomal Status. Fertil. Steril. 2003, 79 (Suppl 3), 1616–1624. [Google Scholar] [CrossRef]
- Harbuz, R.; Zouari, R.; Pierre, V.; Ben Khelifa, M.; Kharouf, M.; Coutton, C.; Merdassi, G.; Abada, F.; Escoffier, J.; Nikas, Y.; et al. A Recurrent Deletion of DPY19L2 Causes Infertility in Man by Blocking Sperm Head Elongation and Acrosome Formation. Am. J. Hum. Genet. 2011, 88, 351–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, Y.-L.; Zhu, F.-X.; Yan, J.; Chen, L.; Tang, W.-H.; Xiao, S.; Mo, W.-K.; Zhang, Z.-G.; He, X.-J.; Qiao, J.; et al. Novel DPY19L2 Variants in Globozoospermic Patients and the Overcoming This Male Infertility. Asian J. Androl. 2019, 21, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Stone, S.; O’Mahony, F.; Khalaf, Y.; Taylor, A.; Braude, P. A Normal Livebirth after Intracytoplasmic Sperm Injection for Globozoospermia without Assisted Oocyte Activation: Case Report. Hum. Reprod. (Oxf. Engl.) 2000, 15, 139–141. [Google Scholar] [CrossRef]
- Chianese, C.; Fino, M.G.; Riera Escamilla, A.; López Rodrigo, O.; Vinci, S.; Guarducci, E.; Daguin, F.; Muratori, M.; Tamburrino, L.; Lo Giacco, D.; et al. Comprehensive Investigation in Patients Affected by Sperm Macrocephaly and Globozoospermia. Andrology 2015, 3, 203–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyono, K.; Nakajo, Y.; Nishinaka, C.; Hattori, H.; Kyoya, T.; Ishikawa, T.; Abe, H.; Araki, Y. A Birth from the Transfer of a Single Vitrified-Warmed Blastocyst Using Intracytoplasmic Sperm Injection with Calcium Ionophore Oocyte Activation in a Globozoospermic Patient. Fertil. Steril. 2009, 91, 931.e7–931.e11. [Google Scholar] [CrossRef] [PubMed]
- Heindryckx, B.; Van der Elst, J.; De Sutter, P.; Dhont, M. Treatment Option for Sperm- or Oocyte-Related Fertilization Failure: Assisted Oocyte Activation Following Diagnostic Heterologous ICSI. Hum. Reprod. 2005, 20, 2237–2241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egashira, A.; Murakami, M.; Haigo, K.; Horiuchi, T.; Kuramoto, T. A Successful Pregnancy and Live Birth after Intracytoplasmic Sperm Injection with Globozoospermic Sperm and Electrical Oocyte Activation. Fertil. Steril. 2009, 92, 2037.e5–2037.e9. [Google Scholar] [CrossRef]
- Lanzendorf, S.; Maloney, M.; Ackerman, S.; Acosta, A.; Hodgen, G. Fertilizing Potential of Acrosome-Defective Sperm Following Microsurgical Injection into Eggs. Gamete Res. 1988, 19, 329–337. [Google Scholar] [CrossRef]
- Virant-Klun, I.; Tomazevic, T.; Meden-Vrtovec, H. Sperm Single-Stranded DNA, Detected by Acridine Orange Staining, Reduces Fertilization and Quality of ICSI-Derived Embryos. J. Assist. Reprod. Genet. 2002, 19, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Frydman, N.; Prisant, N.; Hesters, L.; Frydman, R.; Tachdjian, G.; Cohen-Bacrie, P.; Fanchin, R. Adequate Ovarian Follicular Status Does Not Prevent the Decrease in Pregnancy Rates Associated with High Sperm DNA Fragmentation. Fertil. Steril. 2008, 89, 92–97. [Google Scholar] [CrossRef]
- Zhang, Z.-Q.; Long, S.-G.; Huang, Z.-H.; Xin, C.-L.; Wu, Q.-F. Different Outcomes after Intracytoplasmic Sperm Injection without Oocyte Activation in Two Patients with Different Types of Globozoospermia. Andrologia 2016, 48, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Borges, E.J.; de Almeida Ferreira Braga, D.P.; de Sousa Bonetti, T.C.; Iaconelli, A.J.; Franco, J.G.J. Artificial Oocyte Activation Using Calcium Ionophore in ICSI Cycles with Spermatozoa from Different Sources. Reprod. Biomed. Online 2009, 18, 45–52. [Google Scholar] [CrossRef]
- Gómez-Torres, M.J.; Robles-Gómez, L.; Huerta-Retamal, N.; Sáez-Espinosa, P.; Avilés, M.; Aizpurua, J.; Romero, A. FE-SEM Characterization of α-Mannose Density and Surface Mapping Changes in Human Sperm Head During In Vitro Capacitation. Microsc. Microanal. 2020, 26, 1220–1225. [Google Scholar] [CrossRef] [PubMed]
- Sáez-Espinosa, P.; Huerta-Retamal, N.; Robles-Gómez, L.; Avilés, M.; Aizpurua, J.; Velasco, I.; Romero, A.; Gómez-Torres, M.J. Influence of In Vitro Capacitation Time on Structural and Functional Human Sperm Parameters. Asian J. Androl. 2020, 22, 447–453. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sáez-Espinosa, P.; Robles-Gómez, L.; Ortega-López, L.; Aizpurua, J.; Gómez-Torres, M.J. Immunofluorescence and High-Resolution Microscopy Reveal New Insights in Human Globozoospermia. Int. J. Mol. Sci. 2022, 23, 1729. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031729
Sáez-Espinosa P, Robles-Gómez L, Ortega-López L, Aizpurua J, Gómez-Torres MJ. Immunofluorescence and High-Resolution Microscopy Reveal New Insights in Human Globozoospermia. International Journal of Molecular Sciences. 2022; 23(3):1729. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031729
Chicago/Turabian StyleSáez-Espinosa, Paula, Laura Robles-Gómez, Leonor Ortega-López, Jon Aizpurua, and María José Gómez-Torres. 2022. "Immunofluorescence and High-Resolution Microscopy Reveal New Insights in Human Globozoospermia" International Journal of Molecular Sciences 23, no. 3: 1729. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031729