
Engineering Chimeras by Fusing Plant Receptor-like Kinase EMS1 and BRI1 Reveals the Two Receptors’ Structural Specificity and Molecular Mechanisms

 , and
, and 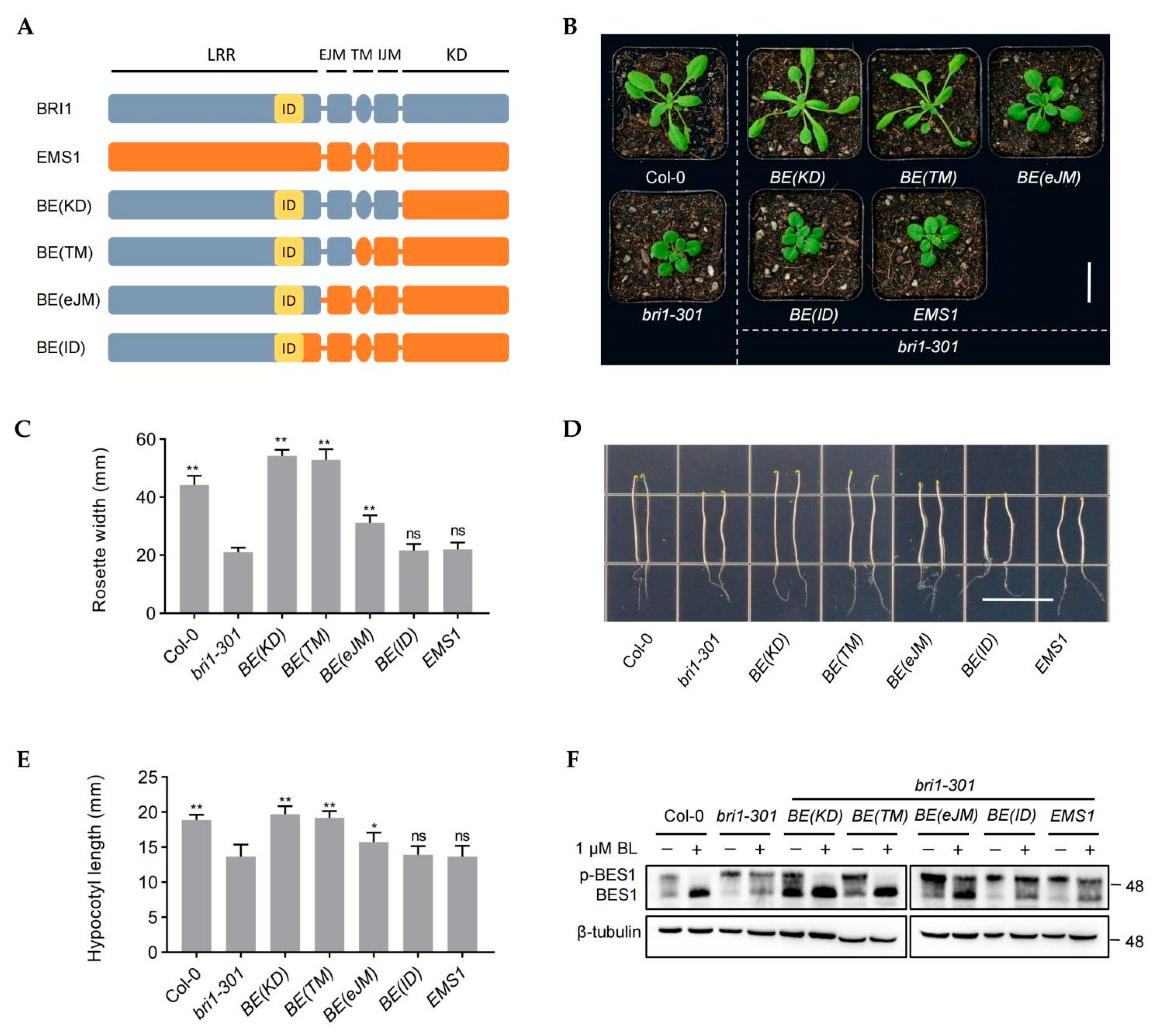{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Transmembrane, Intracellular Juxtamembrane, and Kinase Domains, but Not the Extracellular Jux-Tamembrane Domain, of EMS1 Can Completely Replace the Corresponding BRI1 Domain
2.2. The LRR14-eJM Domain Confers the Functional Divergence between BRI1 and EMS1
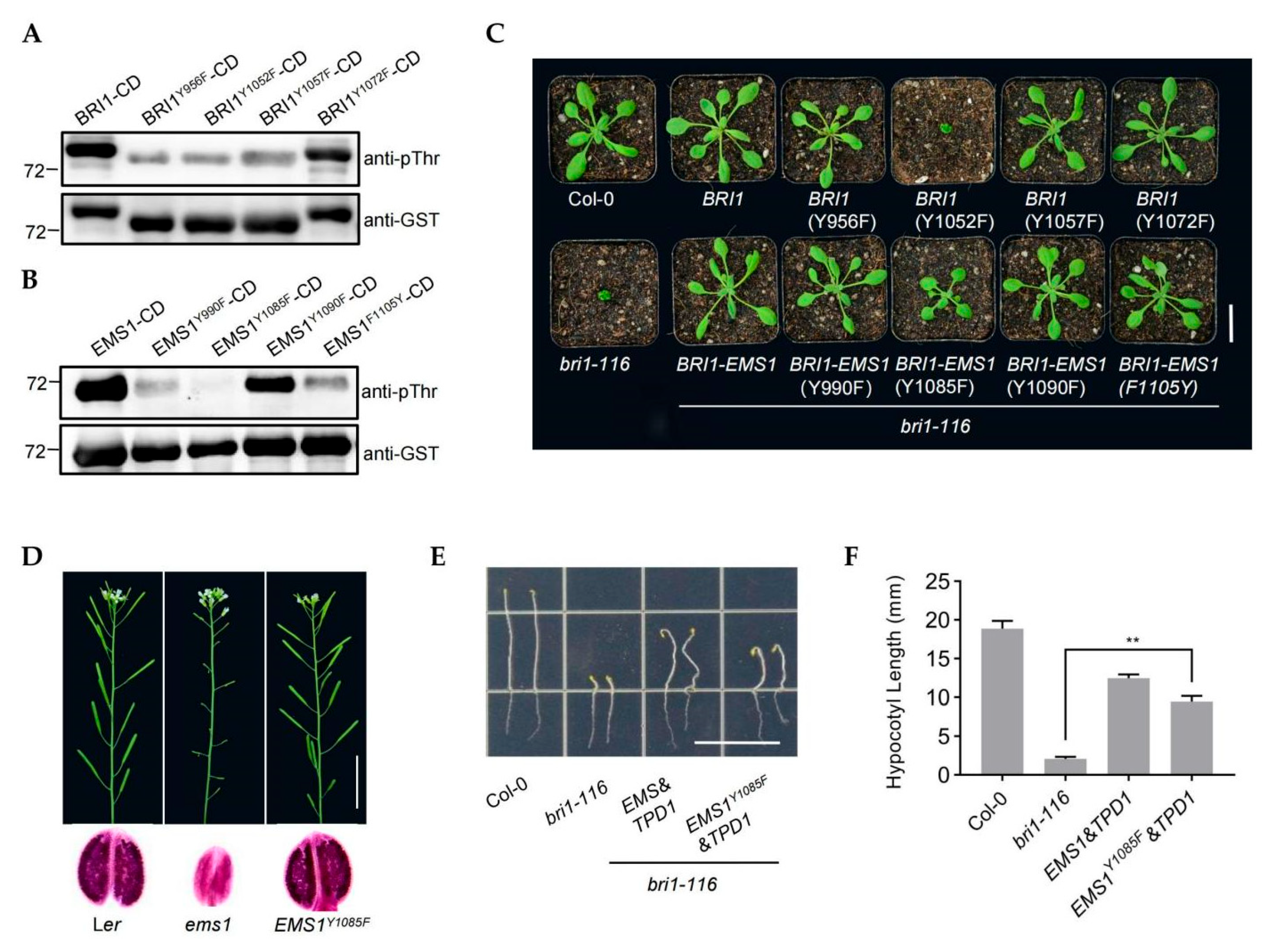
2.3. The Kinase Activity of EMS1 Is Weaker than That of BRI1
2.4. The Tyrosine (Tyr) Residues Y1052 Is Essential for BRI1 but the Corresponding Residue Y1085 Is Not Essential for EMS1
2.5. EMS1 and BRI1 Share the Substrate BSKs
3. Discussion
4. Methods and Materials
4.1. Plant Materials and Growth Conditions
4.2. Constructs and Transgenic Plants Generation
4.3. Protein Extraction and Immunoblot Analysis
4.4. Pollen Staining
4.5. RT and qRT-PCR Analysis
4.6. Bimolecular Fluorescence Complementation Assay
4.7. Coimmunoprecipitation
4.8. In Vitro Kinase Assays
4.9. Pull-Down
4.10. GUS Staining
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shiu, S.H.; Bleecker, A.B. Plant Receptor-Like Kinase Gene Family: Diversity, Function, and Signaling. Sci. STKE 2001, 113, re22. [Google Scholar] [CrossRef] [PubMed]
- Manhães, A.; Ortiz-Morea, F.; He, P.; Shan, L. Plant Plasma Membrane-Resident Receptors: Surveillance for Infections and Coordination for Growth and Development. J. Integr. Plant Biol. 2020, 63, 79–101. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chory, J. A Putative Leucine-Rich Repeat Receptor Kinase Involved in Brassinosteroid Signal Transduction. Cell 1997, 90, 929–938. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, T.; Caño-Delgado, A.; Seto, H.; Hiranuma, S.; Fujioka, S.; Yoshida, S.; Chory, J. Binding of Brassinosteroids to the Extracellular Domain of Plant Receptor Kinase BRI1. Nature 2005, 433, 167–171. [Google Scholar] [CrossRef]
- Kim, T.-W.; Guan, S.; Sun, Y.; Deng, Z.; Tang, W.; Shang, J.-X.; Sun, Y.; Burlingame, A.; Wang, Z.-Y. Brassinosteroid Signal Transduction from Cell-Surface Receptor Kinases to Nuclear Transcription Factors. Nat. Cell Biol. 2009, 11, 1254–1260. [Google Scholar] [CrossRef]
- Wang, Z.-Y.; Nakano, T.; Gendron, J.; He, J.; Chen, M.; Vafeados, D.; Yang, Y.; Fujioka, S.; Yoshida, S.; Asami, T.; et al. Nuclear-Localized BZR1 Mediates Brassinosteroid-Induced Growth and Feedback Suppression of Brassinosteroid Biosynthesis. Dev. Cell 2002, 2, 505–513. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Wang, Z.-Y.; Mora-Garcia, S.; Li, J.; Yoshida, S.; Asami, T.; Chory, J. BES1 Accumulates in the Nucleus in Response to Brassinosteroids to Regulate Gene Expression and Promote Stem Elongation. Cell 2002, 109, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.-W.; Wang, Z.-Y. Brassinosteroid Signal Transduction from Receptor Kinases to Transcription Factors. Annu. Rev. Plant Biol. 2010, 61, 681–704. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Vafeados, D.; Tao, Y.; Yoshida, S.; Asami, T.; Chory, J. A New Class of Transcription Factors Mediates Brassinosteroid-Regulated Gene Expression in Arabidopsis. Cell 2005, 120, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Lv, M.; Li, J. Molecular Mechanisms of Brassinosteroid-Mediated Responses to Changing Environments in Arabidopsis. Int. J. Mol. Sci. 2020, 21, 2737. [Google Scholar] [CrossRef] [Green Version]
- Wolf, S. Deviating from the Beaten Track: New Twists in Brassinosteroid Receptor Function. Int. J. Mol. Sci. 2020, 21, 1561. [Google Scholar] [CrossRef] [Green Version]
- Jia, G.; Liu, X.; Owen, H.; Zhao, D. Signaling of Cell Fate Determination by the TPD1 Small Protein and EMS1 Receptor Kinase. Proc. Natl. Acad. Sci. USA 2008, 105, 2220–2225. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Xie, L.-F.; Mao, H.-Z.; Puah, C.; Yang, W.; Jiang, L.; Sundaresan, V.; Ye, D. Tapetum Determinant1 Is Required for Cell Specialization in the Arabidopsis Anther. Plant Cell 2004, 15, 2792–2804. [Google Scholar] [CrossRef] [Green Version]
- Zheng, B.; Bai, Q.; Wu, L.; Liu, H.; Liu, Y.; Xu, W.; Li, G.; Ren, H.; She, X.; Wu, G. EMS1 and BRI1 Control Separate Biological Processes via Extracellular Domain Diversity and Intracellular Domain Conservation. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Lv, M.; Wang, Y.; Wang, P.-A.; Cui, Y.; Li, M.; Wang, R.; Gou, X.; Li, J. BES1 Is Activated by EMS1-TPD1-SERK1/2-Mediated Signaling to Control Tapetum Development in Arabidopsis Thaliana. Nat. Commun. 2019, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.-Z.; Wang, G.-F.; Speal, B.; Ma, H. The Excess Microsporocytes 1 Encodes a Putative Leucine-Rich Repeat Receptor Kinase that Controls Somatic and Reproductive Cell Fates in the Arabidopsis Anther. Genes Dev. 2002, 16, 2021–2031. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Wang, Y.; Huang, J.; Ahsan, N.; Biener, G.; Paprocki, J.; Thelen, J.; Raicu, V.; Zhao, D. Two SERK Receptor-Like Kinases Interact with the Receptor-Like Kinase EMS1 in Anther Cell Fate Determination. Plant Physiol. 2016, 173, 326–337. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wen, J.; Lease, K.; Doke, J.; Tax, F.; Walker, J. BAK1, an Arabidopsis LRR Receptor-Like Protein Kinase, Interacts with BRI1 and Modulates Brassinosteroid Signaling. Cell 2002, 110, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Nam, K.; Li, J. BRI1/BAK1, a Receptor Kinase Pair Mediating Brassinosteroid Signaling. Cell 2002, 110, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Kota, U.; He, K.; Blackburn, R.; Li, J.; Goshe, M.; Huber, S.; Clouse, S. Sequential Transphosphorylation of the BRI1/BAK1 Receptor Kinase Complex Impacts Early Events in Brassinosteroid Signaling. Dev. Cell 2008, 15, 220–235. [Google Scholar] [CrossRef] [Green Version]
- Noguchi, T.; Fujioka, S.; Choe, S.; Takatsuto, S.; Yoshida, S.; Yuan, H.; Feldmann, K.; Tax, F. Brassinosteroid-Insensitive Dwarf Mutants of Arabidopsis Accumulate Brassinosteroids. Plant Physiol. 1999, 121, 743–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; He, Y. Roles of Brassinosteroids in Plant Reproduction. Int. J. Mol. Sci. 2020, 21, 872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hothorn, M.; Belkhadir, Y.; Dreux, M.; Dabi, T.; Noel, J.; Wilson, I.; Chory, J. Structural Basis of Steroid Hormone Perception by the Receptor Kinase BRI1. Nature 2011, 474, 467–471. [Google Scholar] [CrossRef] [Green Version]
- She, J.; Zhifu, H.; Kim, T.-W.; Wang, J.; Cheng, W.; Chang, J.; Shi, S.; Wang, J.; Yang, M.; Wang, Z.-Y.; et al. Structure Insight into Brassinosteroid Perception by BRI1. Nature 2011, 474, 472–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vert, G.; Nemhauser, J.; Geldner, N.; Hong, F.; Chory, J. Molecular Mechanisms of Steroid Hormone Signaling in Plants. Annu. Rev. Cell Dev. Biol. 2005, 21, 177–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedrichsen, D. Brassinosteroid-Insensitive-1 Is a Ubiquitously Expressed Leucine-Rich Repeat Receptor Serine/Threonine Kinase. Plant Physiol. 2000, 123, 1247–1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, C.; Yan, K.; Han, J.-T.; Tao, L.; Lv, M.; Shi, T.; He, Y.-X.; Wierzba, M.; Tax, F.; Li, J. Scanning for New BRI1 Receptor Mutations via TILLING Analysis. Plant Physiol. 2017, 174, 1881–1896. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, X.; Meisenhelder, J.; Hunter, T.; Yoshida, S.; Asami, T.; Chory, J. Autoregulation and Homodimerization Are Involved in the Activation of the Plant Steroid Receptor BRI1. Dev. Cell 2005, 8, 855–865. [Google Scholar] [CrossRef] [Green Version]
- Walker, J. Structure and Function of the Receptor-Like Protein Kinases of Higher Plants. Plant Mol. Biol. 1995, 26, 1599–1609. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, S.; Gan, S.; Wang, X.; Liu, J.; Wang, X. Role of Specific Phosphorylation Sites of Arabidopsis Brassinosteroid-Insensitive 1 Receptor Kinase in Plant Growth and Development. J. Plant Growth Regul. 2016, 35, 755–769. [Google Scholar] [CrossRef]
- Oh, M.-H.; Ray, W.K.; Huber, S.C.; Asara, J.M.; Gage, D.A.; Clouse, S.D. Recombinant Brassinosteroid Insensitive 1 Receptor-Like Kinase Autophosphorylates on Serine and Threonine Residues and Phosphorylates a Conserved Peptide Motif in Vitro. Plant Physiol. 2000, 124, 751–766. [Google Scholar] [CrossRef] [Green Version]
- Oh, M.-H.; Wang, X.; Kota, U.; Goshe, M.; Clouse, S.; Huber, S. Tyrosine Phosphorylation of the BRI1 Receptor Kinase Emerges as a Component of Brassinosteroid Signaling in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 658–663. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Goshe, M.B.; Soderblom, E.J.; Phinney, B.S.; Kuchar, J.A.; Li, J.; Asami, T.; Yoshida, S.; Huber, S.C.; Clouse, S.D. Identification and Functional Analysis of in Vivo Phosphorylation Sites of the Arabidopsis BRASSINOSTEROID-INSENSITIVE1 Receptor Kinase. Plant Cell 2005, 17, 1685–1703. [Google Scholar] [CrossRef] [Green Version]
- Oh, M.-H.; Clouse, S.; Huber, S. Tyrosine Phosphorylation in Brassinosteroid Signaling. Plant Signal. Behav. 2009, 4, 1182–1185. [Google Scholar] [CrossRef] [Green Version]
- Caño-Delgado, A.; Yin, Y.; Yu, C.; Vafeados, D.; Mora-García, S.; Cheng, J.-C.; Nam, K.; Li, J.; Chory, J. BRL1 and BRL3 are Novel Brassinosteroid Receptors that Function in Vascular Differentiation in Arabidopsis. Development 2004, 131, 5341–5351. [Google Scholar] [CrossRef] [Green Version]
- Zhou, A.; Huachun, W.; Walker, J.; Li, J. BRL1, a Leucine-Rich Repeat Receptor-Like Protein Kinase, Is Functionally Redundant with BRI1 in Regulating Arabidopsis Brassinosteroid Signaling. Plant J. Cell Mol. Biol. 2004, 40, 399–409. [Google Scholar] [CrossRef]
- Furumizu, C.; Sawa, S. Insight into Early Diversification of Leucine-Rich Repeat Receptor-Like Kinases Provided by the Sequenced Moss and Hornwort Genomes. Plant Mol. Biol. 2021, 107, 1–17. [Google Scholar] [CrossRef]
- Ferreira-Guerra, M.; Marques, M.; Mora-García, S.; Caño-Delgado, A. Delving into the Evolutionary Origin of Steroid Sensing in Plants. Curr. Opin. Plant Biol. 2020, 57, 87–95. [Google Scholar] [CrossRef]
- Wang, H.; Mao, H. On the Origin and Evolution of Plant Brassinosteroid Receptor Kinases. J. Mol. Evol. 2013, 78, 118–129. [Google Scholar] [CrossRef]
- Tanaka, K.; Asami, T.; Yoshida, S.; Nakamura, Y.; Matsuo, T.; Okamoto, S. Brassinosteroid Homeostasis in Arabidopsis Is Ensured by Feedback Expressions of Multiple Genes Involved in Its Metabolism. Plant Physiol. 2005, 138, 1117–1125. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Zhang, T.; Linstroth, L.; Tillman, Z.; Otegui, M.; Owen, H.; Zhao, D. Control of Anther Cell Differentiation by the Small Protein Ligand TPD1 and Its Receptor EMS1 in Arabidopsis. PLoS Genet. 2016, 12, e1006147. [Google Scholar] [CrossRef] [Green Version]
- Sreeramulu, S.; Mostizky, Y.; Sukumaran, S.; Shani, E.; Nahum, H.; Salomon, D.; Hayun, L.; Gruetter, C.; Rauh, D.; Ori, N.; et al. BSKs Are Partially Redundant Positive Regulators of Brassinosteroid Signaling in Arabidopsis. Plant J. Cell Mol. Biol. 2013, 74, 905–919. [Google Scholar] [CrossRef]
- Tang, W.; Kim, T.-W.; Oses-Prieto, J.; Sun, Y.; Deng, Z.; Zhu, S.; Wang, R.; Burlingame, A.; Wang, Z.-Y. BSKs Mediate Signal Transduction from the Receptor Kinase BRI1 in Arabidopsis. Science 2008, 321, 557–560. [Google Scholar] [CrossRef] [Green Version]
- Jose, J.; Ghantasala, S.; Roy Choudhury, S. Arabidopsis Transmembrane Receptor-Like Kinases (RLKs): A Bridge between Extracellular Signal and Intracellular Regulatory Machinery. Int. J. Mol. Sci. 2020, 21, 4000. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.; Zhang, D. The Role of Receptor-Like Kinases in Regulating Plant Male Reproduction. Plant Reprod. 2018, 31, 77–87. [Google Scholar] [CrossRef]
- Ou, Y.; Kui, H.; Li, J. Receptor-Like Kinases in Root Development: Current Progress and Future Directions. Mol. Plant 2020, 14, 166–185. [Google Scholar] [CrossRef] [PubMed]
- Tsuwamoto, R.; Fukuoka, H.; Takahata, Y. GASSHO1 and GASSHO2 Encoding a Putative Leucine-Rich Repeat Transmembrane-Type Receptor Kinase Are Essential for the Normal Development of the Epidermal Surface in Arabidopsis Embryos. Plant J. Cell Mol. Biol. 2008, 54, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Ali, K.; Li, W.; Qin, Y.; Wang, S.; Feng, L.; Wei, Q.; Bai, Q.; Zheng, B.; Li, G.; Ren, H.; et al. Kinase Function of Brassinosteroid Receptor Specified by Two Allosterically Regulated Subdomains. Front. Plant Sci. 2022, 12, 1–13. [Google Scholar] [CrossRef]
- Kim, S.; Park, J.; Jeon, B.; Hwang, G.; Kang, N.; We, Y.; Park, W.-Y.; Oh, E.; Kim, J. Chemical Control of Receptor Kinase Signaling by Rapamycin-Induced Dimerization. Mol. Plant 2021, 14, 1379–1390. [Google Scholar] [CrossRef]
- He, Z.; Wang, Z.Y.; Li, J.; Zhu, Q.; Lamb, C.; Ronald, P.; Chory, J. Perception of Brassinosteroids by the Extracellular Domain of the Receptor Kinase BRI1. Science 2000, 288, 2360–2363. [Google Scholar] [CrossRef] [Green Version]
- Brutus, A.; Sicilia, F.; Macone, A.; Cervone, F.; De Lorenzo, G. A Domain Swap Approach Reveals a Role of the Plant Wall-Associated Kinase 1 (WAK1) as a Receptor of Oligogalacturonides. Proc. Natl. Acad. Sci. USA 2010, 107, 9452–9457. [Google Scholar] [CrossRef] [Green Version]
- Hohmann, U.; Ramakrishna, P.; Wang, K.; Lorenzo-Orts, L.; Nicolet, J.; Henschen, A.; Barberon, M.; Bayer, M.; Hothorn, M. Constitutive Activation of Leucine-Rich Repeat Receptor Kinase Signaling Pathways by BAK1-Interacting Receptor-like Kinase 3 Chimera. Plant Cell 2020, 32, 3311–3323. [Google Scholar] [CrossRef]
- Wang, X.; Chory, J. Brassinosteroids Regulate Dissociation of BKI1, A Negative Regulator of BRI1 Signaling, from the Plasma Membrane. Science 2006, 313, 1118–1122. [Google Scholar] [CrossRef]
- Jaillais, Y.; Hothorn, M.; Belkhadir, Y.; Dabi, T.; Nimchuk, Z.; Meyerowitz, E.; Chory, J. Tyrosine Phosphorylation Controls Brassinosteroid Receptor Activation by Triggering Membrane Release of Its Kinase Inhibitor. Genes Dev. 2011, 25, 232–237. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Li, Z.; Biener, G.; Xiong, E.; Malik, S.; Eaton, N.; Zhao, C.; Raicu, V.; Kong, H.; Zhao, D. Carbonic Anhydrases Function in Anther Cell Differentiation Downstream of the Receptor-Like Kinase EMS1. Plant Cell 2017, 29, 1335–1356. [Google Scholar] [CrossRef] [Green Version]
- Gou, X.; Li, J. Paired Receptor and Coreceptor Kinases Perceive Extracellular Signals to Control Plant Development. Plant Physiol. 2020, 182, 1667–1681. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, Q.; Li, C.; Wu, L.; Liu, H.; Ren, H.; Li, G.; Wang, Q.; Wu, G.; Zheng, B. Engineering Chimeras by Fusing Plant Receptor-like Kinase EMS1 and BRI1 Reveals the Two Receptors’ Structural Specificity and Molecular Mechanisms. Int. J. Mol. Sci. 2022, 23, 2155. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23042155
Bai Q, Li C, Wu L, Liu H, Ren H, Li G, Wang Q, Wu G, Zheng B. Engineering Chimeras by Fusing Plant Receptor-like Kinase EMS1 and BRI1 Reveals the Two Receptors’ Structural Specificity and Molecular Mechanisms. International Journal of Molecular Sciences. 2022; 23(4):2155. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23042155
Chicago/Turabian StyleBai, Qunwei, Chenxi Li, Lei Wu, Huan Liu, Hongyan Ren, Guishuang Li, Qiuling Wang, Guang Wu, and Bowen Zheng. 2022. "Engineering Chimeras by Fusing Plant Receptor-like Kinase EMS1 and BRI1 Reveals the Two Receptors’ Structural Specificity and Molecular Mechanisms" International Journal of Molecular Sciences 23, no. 4: 2155. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23042155