
Salivary Glands after Prolonged Aluminum Exposure: Proteomic Approach Underlying Biochemical and Morphological Impairments in Rats

 , , , and
, , , and 
Abstract
:1. Introduction
2. Results
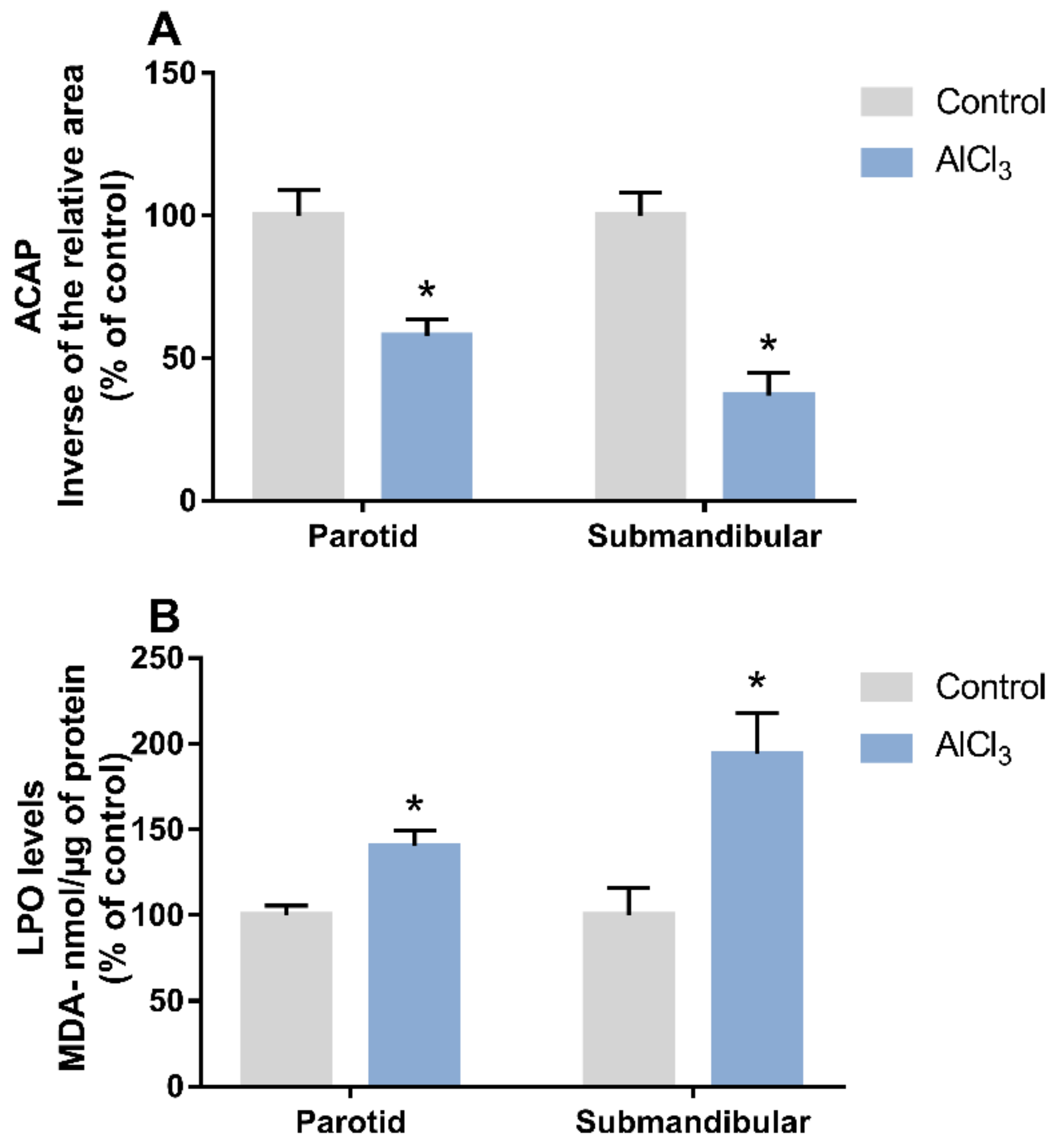
2.1. Al Triggered Biochemical Changes Related to Redox Homeostasis in the Salivary Glands of Rats
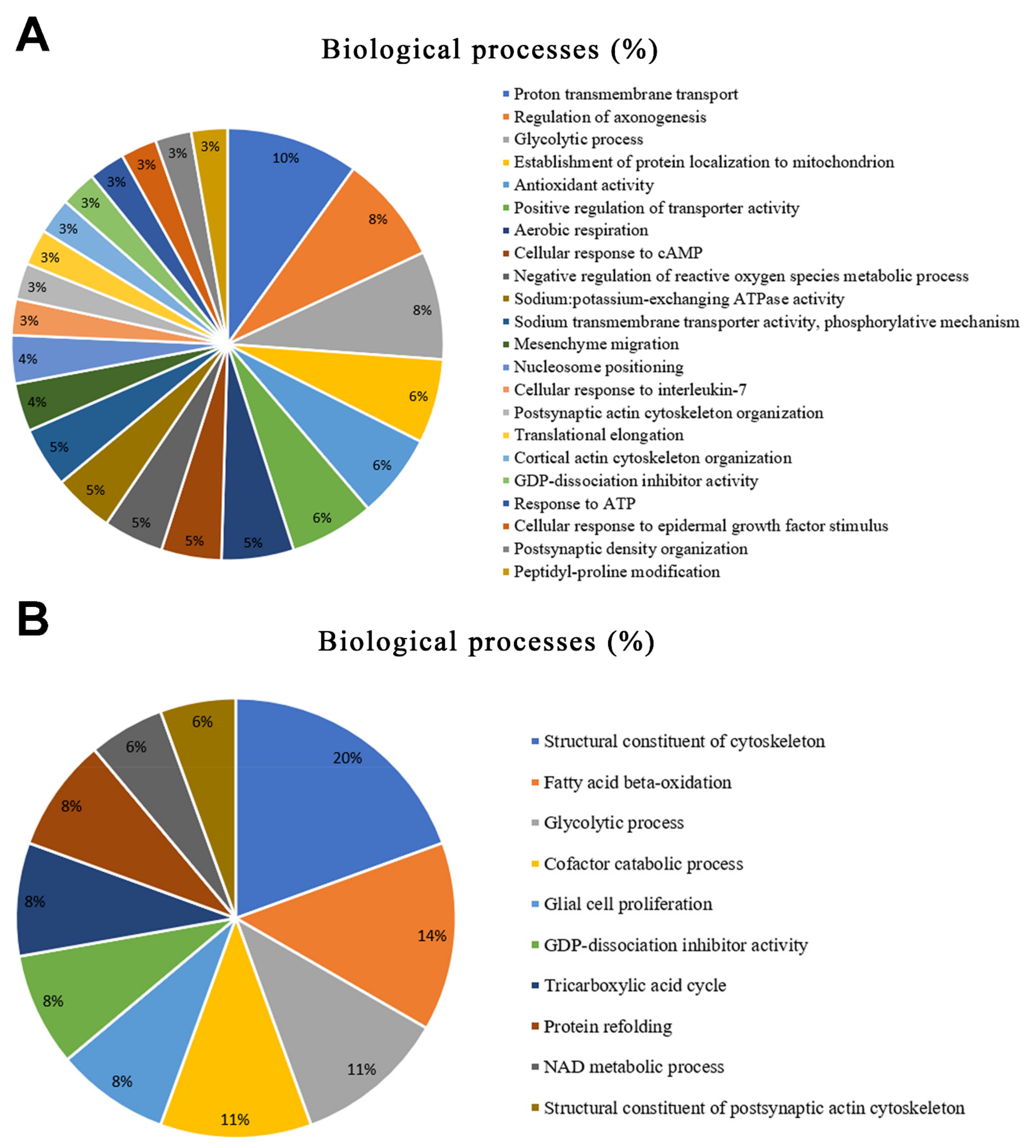
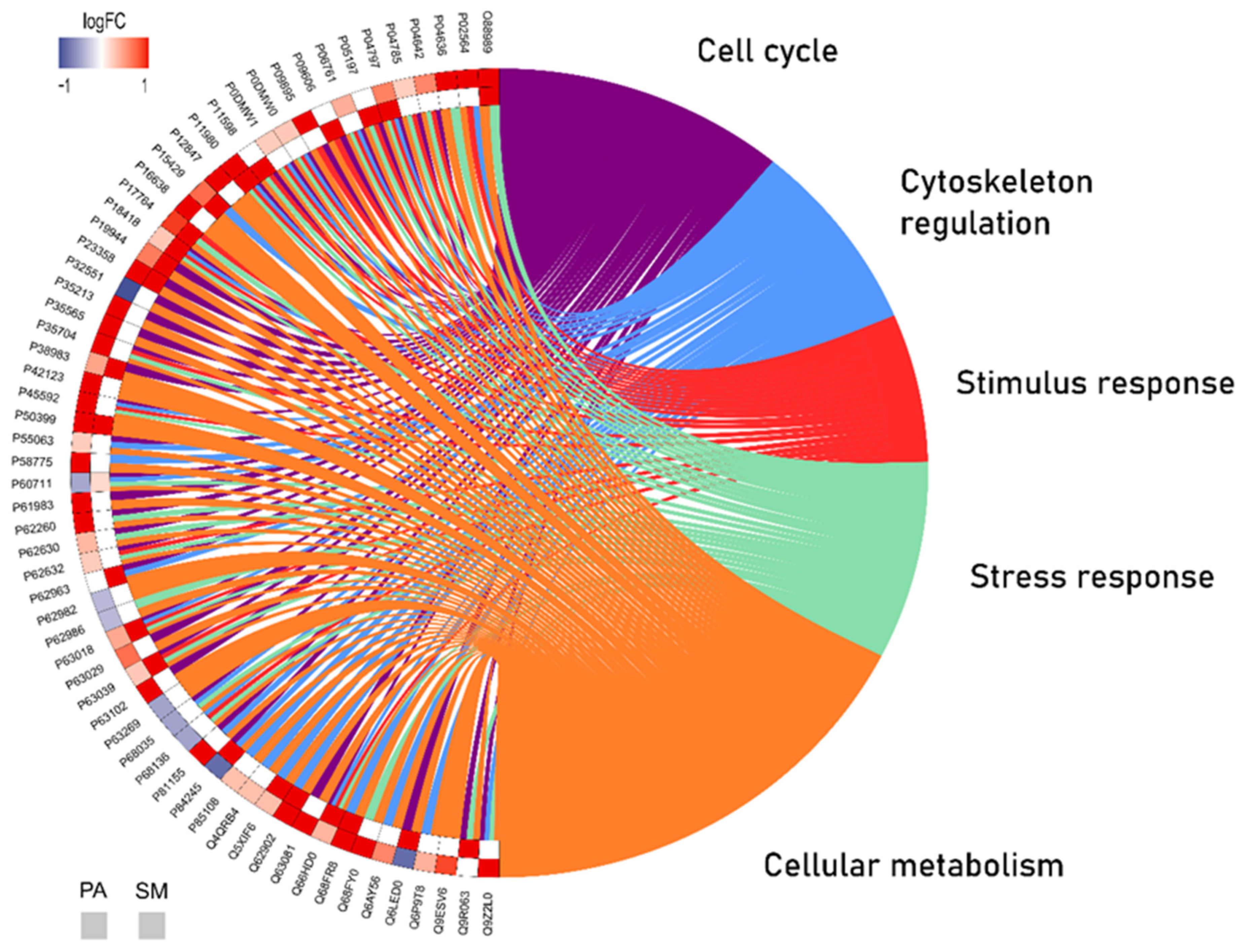
2.2. Long-Term AlCl3 Exposure Promoted Significant Changes in the Global Proteomic Profile of Major Salivary Glands
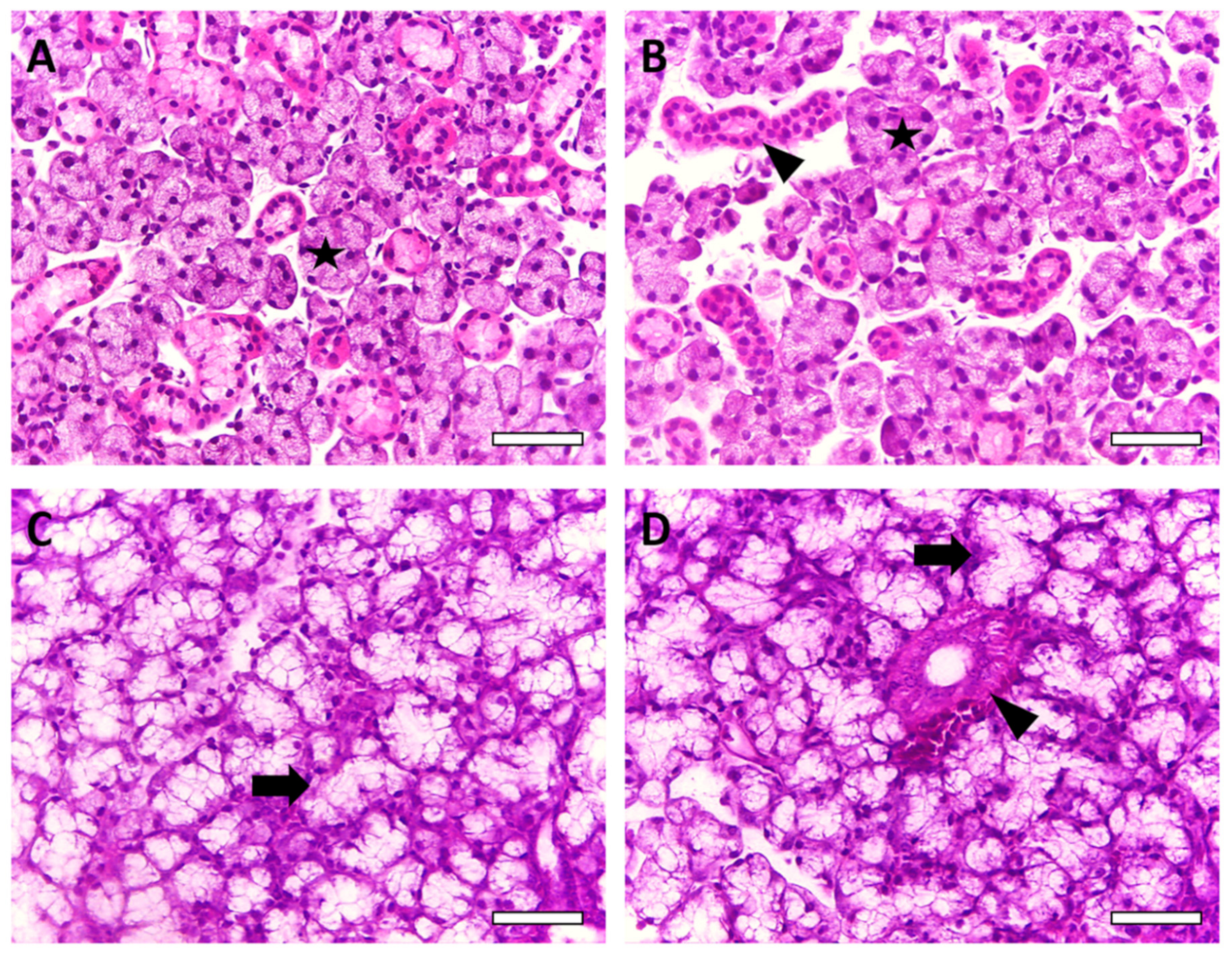
2.3. Salivary Gland Morphology Was Affected by Chronic AlCl3 Exposure
3. Discussion
4. Materials and Methods
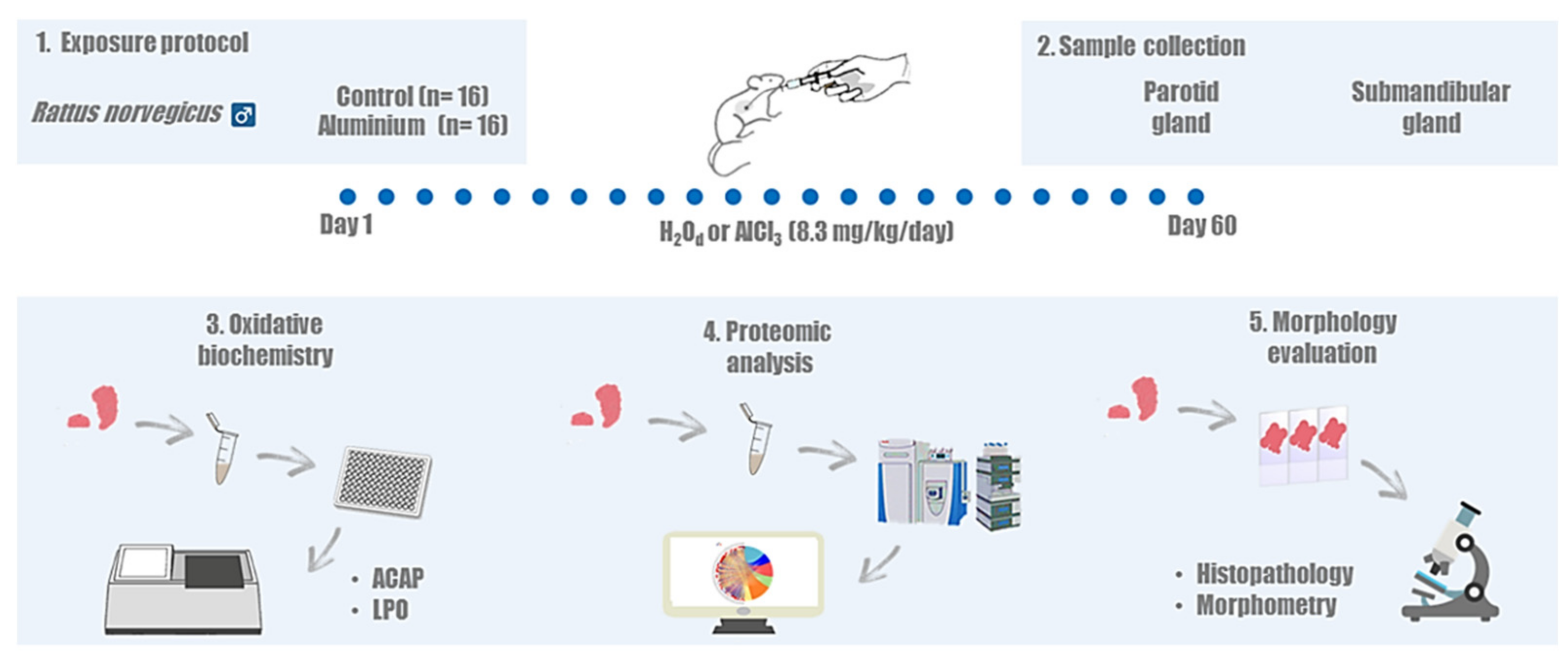
4.1. Ethics Statement and Experimental Groups
4.2. Aluminum Exposure Protocol
4.3. Sample Collection
4.4. Biochemical Assays
4.4.1. Antioxidant Capacity against Peroxyl Radicals Assay
4.4.2. Lipid Peroxidation Assay
4.5. Proteomic Analysis
4.5.1. Protein Extraction
4.5.2. Mass Spectrometry Analysis
4.5.3. Over-Representation Analysis
4.6. Perfusion and Histological Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carson, B.L. Aluminum compounds. Review of toxicological literature. In Abridged Final Report, Integrated Laboratory Systems; Research Triangle Park: North Carolina, NC, USA, 2000; p. 84. [Google Scholar]
- Associação Brasileira do Alumínio (ABAL). Fundamentos e Aplicações do Alumínio; Associação Brasileira do Alumínio: São Paulo, Brazil, 2007. [Google Scholar]
- Monteiro, M.A. Meio século de mineração industrial na Amazônia e suas implicações para o desenvolvimento regional. Estud. Avançados 2008, 19, 187–207. [Google Scholar] [CrossRef] [Green Version]
- De Meyer, C.M.C.; Rodríguez, J.M.; Carpio, E.A.; García, P.A.; Stengel, C.; Berg, M. Arsenic, manganese and aluminum contamination in groundwater resources of Western Amazonia (Peru). Sci. Total Environ. 2017, 607–608, 1437–1450. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (ESFA). Dietary exposure to aluminium-containing food additives. EFSA Supporting Publ. 2013, 10, 411E. [Google Scholar]
- Lin, Y.-J.; Shih, Y.-J.; Chen, C.-H.; Fang, C.-T. Aluminum salts as an adjuvant for pre-pandemic influenza vaccines: A meta-analysis. Sci. Rep. 2018, 8, 1–7. [Google Scholar] [CrossRef]
- Yokel, R.A.; Florence, R.L. Aluminum bioavailability from tea infusion. Food Chem. Toxicol. 2008, 46, 3659–3663. [Google Scholar] [CrossRef] [Green Version]
- Poole, R.L.; Hintz, S.R.; Mackenzie, N.I.; Kerner, J.A., Jr. Aluminum exposure from pediatric parenteral nutrition: Meeting the new FDA regulation. J. Parenter. Enteral Nutr. 2008, 32, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Yokel, R.A.; Mcnamara, P. Aluminium toxicokinetics: An updated minireview. J. Pharmacol. Toxicol. 2001, 88, 159–167. [Google Scholar] [CrossRef]
- Fernandes, R.M.; Nascimento, P.C.; Martins, M.K.; Bragança Aragão, W.A.; Rivera, L.F.; Bittencourt, L.O.; Cartágenes, S.C.; Crespo-Lopez, M.E.; Maia, C.; Lima, R. Evaluation of Cerebellar Function and Integrity of Adult Rats After Long-Term Exposure to Aluminum at Equivalent Urban Region Consumption Concentrations. Biol. Trace Elem. Res. 2020, 199, 1–12. [Google Scholar] [CrossRef]
- Yu, L.; Zhai, Q.; Yin, R.; Li, P.; Tian, F.; Liu, X.; Zhao, J.; Gong, J.; Zhang, H.; Chen, W. Lactobacillus plantarum CCFM639 Alleviate Trace Element Imbalance-Related Oxidative Stress in Liver and Kidney of Chronic Aluminum Exposure Mice. Biol. Trace Elem. Res. 2017, 176, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Costa, N.M.M.; Correa, R.S.; Júnior, I.S.M.; Figueiredo, A.J.R.; Vilhena, K.F.B.; Farias-Junior, P.M.A.; Teixeira, F.B.; Ferreira, N.M.M.; Pereira-Júnior, J.B.; Dantas, K.D.G.F. Physical, Chemical, and Immunohistochemical Investigation of the Damage to Salivary Glands in a Model of Intoxication with Aluminium Citrate. Int. J. Environ. Res. 2014, 11, 12429–12440. [Google Scholar] [CrossRef] [PubMed]
- Miletich, I. Introduction to salivary glands: Structure, function and embryonic development. Front. Oral Biol. 2010, 14, 1–20. [Google Scholar]
- Plemons, J.M.; Al-Hashimi, I.; Marek, C.L. American Dental Association Council on Scientific Affairs. Managing xerostomia and salivary gland hypofunction: Executive summary of a report from the American Dental Association Council on Scientific Affairs. J. Am. Dent. Assoc. 2014, 145, 867–873. [Google Scholar] [CrossRef]
- Walsh, L.J. Dry mouth: A clinical problem for children and young adults. Int. Dent. SA 2010, 9, 48–58. [Google Scholar]
- Aragão, W.A.B.; Costa, M.; Fagundes, N.; Silva, M.; Alves, S.; Pinheiro, J.; Amado, L.; Crespo, M.; Maia, C.; Lima, R. Chronic exposure to inorganic mercury induces biochemical and morphological changes in the salivary glands of rats. Metallomics 2017, 9, 1271–1278. [Google Scholar] [CrossRef]
- Bittencourt, L.O.; Dionizio, A.; Nascimento, P.C.; Puty, B.; Leao, L.K.R.; Luz, D.A.; Silva, M.C.F.; Amado, L.L.; Leite, A.; Buzalaf, M.R. Proteomic approach underlying the hippocampal neurodegeneration caused by low doses of methylmercury after long-term exposure in adult rats. Metallomics 2019, 11, 390–403. [Google Scholar] [CrossRef]
- Farias-Junior, P.M.A.; Teixeira, F.B.; Fagundes, N.C.; Miranda, G.F.; Bittencourt, L.O.; de Oliveira, R.O.; Silva, M.C.; Sagica, S.; de Oliveira, E.; Crespo-López, M.E.; et al. Chronic intoxication by methylmercury leads to oxidative damage and cell death in salivary glands of rats. Metallomics 2017, 9, 1778–1785. [Google Scholar] [CrossRef] [PubMed]
- Lima, L.A.; Bittencourt, L.; Puty, B.; Fernandes, R.; Nascimento, P.; Silva, M.C.; Alves-Junior, S.; Pinheiro, J.; Lima, R. Methylmercury intoxication promotes metallothionein response and cell damage in salivary glands of rats. Biol. Trace Elem. Res. 2018, 185, 135–142. [Google Scholar] [CrossRef]
- Nascimento, P.C.; Ferreira, M.K.; Balbinot, K.M.; Alves-Junior, S.M.; Pinheiro, J.V.; Silveira, F.M.; Martins, M.D.; Lopez, E.M.; Lima, R.R. Methylmercury-Induced Toxicopathologic Findings in Salivary Glands of Offspring Rats After Gestational and Lactational Exposure. Biol. Trace Elem. Res. 2020, 199, 1–9. [Google Scholar] [CrossRef]
- Oliveira, L.G.; Aragão, W.; Nascimento, P.; Bittencourt, L.; Oliveira, A.; Leão, L.; Alves-Júnior, S.; Pinheiro, J.; Crespo-Lopez, M.; Lima, L. Effects of lead exposure on salivary glands of rats: Insights into the oxidative biochemistry and glandular morphology. Environ. Sci. Pollut. Res. 2020, 28, 1–13. [Google Scholar]
- Souza-Monteiro, D.; Nunes, P.B.O.; Ferreira, R.O.; Eiró, L.G.; Bittencourt, L.O.; Chemelo, V.S.; Santos, S.M.; Souza-Rodrigues, R.D.; Monteiro, M.C.; Lima, R.R. Aluminum-Induced Toxicity in Salivary Glands of Mice After Long-term Exposure: Insights into the Redox State and Morphological Analyses. Biol. Trace Elem. Res. 2020, 198, 575–582. [Google Scholar] [CrossRef]
- Martinez, C.S.; Escobar, A.G.; Uranga-Ocio, J.A.; Pecanha, F.M.; Vassallo, D.V.; Exley, C.; Miguel, M.; Wiggers, G.A. Aluminum exposure for 60 days at human dietary levels impairs spermatogenesis and sperm quality in rats. Reprod. Toxicol. 2017, 73, 128–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joint FAO/WHO Expert Committee on Food Additives (WHO). Meeting, & World Health Organization. Evaluation of Certain Food Additives and Contaminants: Seventy-Third Report of the Joint FAO/WHO Expert Committee on Food Additives; World Health Organization: Geneva, Switzerland, 2011; p. 73. [Google Scholar]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2008, 22, 659–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza-Monteiro, D.; Ferreira, R.O.; Eiró, L.G. Long-term exposure to low doses of aluminum affects mineral content and microarchitecture of rats alveolar bone. Environ. Sci. Pollut. Res. 2021, 28, 45879–45890. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Wang, P.; Gao, X.; Shao, B.; Zhao, S.; Li, Y. Lycopene attenuates aluminum-induced hippocampal lesions by inhibiting oxidative stress-mediated inflammation and apoptosis in the rat. J. Inorg. Biochem. 2019, 193, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Denisov, A.B. Morphological and functional state of major salivary glands under conditions of aluminum chloride excess in drinking water. Bull. Exp. Biol. Med. 2009, 148, 940–942. [Google Scholar] [CrossRef] [PubMed]
- Exley, C. What is the risk of aluminium as a neurotoxin? Expert Rev. Neurother. 2014, 14, 589–591. [Google Scholar] [CrossRef]
- Ruipérez, F.; Mujika, J.; Ugalde, M.; Exley, C.; Lopez, X. Pro-oxidant activity of aluminum: Promoting the Fenton reaction by reducing Fe (III) to Fe (II). J. Inorg. Biochem. 2012, 117, 118–123. [Google Scholar] [CrossRef]
- Mujika, J.I.; Rezabal, E.; Mercero, J.; Ruipérez, F.; Costa, D.; Ugalde, J.; Lopez, X. Aluminium in biological environments: A computational approach. Comput. Struct. Biotechnol. J. 2014, 9, e201403002. [Google Scholar] [CrossRef] [Green Version]
- Samarghandian, S.; Azimi-Nezhad, M.; Farkhondeh, T.; Samini, F. Anti-oxidative effects of curcumin on immobilization-induced oxidative stress in rat brain, liver and kidney. Biomed. Pharmacother. 2017, 87, 223–229. [Google Scholar] [CrossRef]
- Fernandes, R.M.; Corrêa, M.; Aragão, W.; Nascimento, C.; Cartágenes, S.; Rodrigues, C.; Sarmiento, L.; Monteiro, M.; Maia, C.; Crespo-López, M.; et al. Preclinical evidences of aluminum-induced neurotoxicity in hippocampus and pre-frontal cortex of rats exposed to low doses. Ecotoxicol. Environ. Saf. 2020, 206, 111139. [Google Scholar] [CrossRef]
- Furnkranz, A.; Leitinger, N. Regulation of inflammatory responses by oxidized phospholipids: Structure-function relationships. Curr. Pharm. Des. 2004, 10, 915–921. [Google Scholar] [CrossRef]
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef]
- Othman, M.S.; Fareid, M.A.; Abdel Hameed, R.S.; Abdel Moneim, A.E. The Protective Effects of Melatonin on Aluminum-Induced Hepatotoxicity and Nephrotoxicity in Rats. Oxid. Med. Cell. Longev. 2020, 2020, 7375136. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.B.; do Nascimento, J.L.M.; Herculano, A.M.; Crespo-López, M.E. Molecular chaperones: Toward new therapeutic tools. Biomed. Pharmacother. 2011, 65, 239–243. [Google Scholar] [CrossRef]
- Bittencourt, L.O.; Puty, B. Oxidative biochemistry disbalance and changes on proteomic profile in salivary glands of rats induced by chronic exposure to methylmercury. Oxid. Med. Cell. Longev. 2017, 2017, 5653291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araujo, T.T.; Pereira, H. Changes in energy metabolism induced by fluoride: Insights from inside the mitochondria. Chemosphere 2019, 236, 124357. [Google Scholar] [CrossRef]
- Mazumder, S.; Pleasca, D. Caspase-3 activation is a critical determinant of genotoxic stress-induced apoptosis. Apoptosis Cancer. Hum. Press. 2008, 414, 13–21. [Google Scholar]
- Xu, F.; Liu, Y.; Zhao, H.; Yu, K.; Song, M.; Zhu, Y.; Li, Y. Aluminum chloride caused liver dysfunction and mitochondrial energy metabolism disorder in rat. J. Inorg. Biochem. 2017, 174, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Redpath, C.J.; Bou Khalil, M.; Drozdzal, G.; Radisic, M.; McBride, H.M. Mitochondrial hyperfusion during oxidative stress is coupled to a dysregulation in calcium handling within a C2C12 cell model. PLoS ONE 2013, 8, e69165. [Google Scholar]
- Binarová, P.; Tuszynski, J. Tubulin: Structure, Functions and Roles in Disease. Cells 2019, 8, 1294. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, S.G.; Bourgoin-Voillard, S.; Seve, M.; Rachidi, W. Tubulin Beta-3 Chain as a New Candidate Protein Biomarker of Human Skin Aging: A Preliminary Study. Oxid. Med. Cell. Longev. 2017, 2017, 5140360. [Google Scholar] [CrossRef] [PubMed]
- Dasika, S.K.; Vinnakota, K.C.; Beard, D.A. Determination of the catalytic mechanism for mitochondrial malate dehydrogenase. Biophys. J. 2015, 108, 408–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minárik, P.; Tomásková, N.; Kollárová, M.; Antalík, M. Malate dehydrogenases-structure and function. Gen. Physiol. Biophys. 2002, 21, 257–265. [Google Scholar] [PubMed]
- Witjes, L.; Van Troys, M.; Vandekerckhove, J.; Vandepoele, K.; Ampe, C. A new evolutionary model for the vertebrate actin family including two novel groups. Mol. Phylogenet. Evol. 2019, 141, 106632. [Google Scholar] [CrossRef] [PubMed]
- Arnoldi, R.; Chaponnier, C.; Gabbiani, G.; Hinz, B. Heterogeneity of Smooth Muscle; Elsevier: Amsterdam, The Netherlands, 2012; pp. 1183–1195. [Google Scholar]
- Hand, A.R. Salivary Glands, Anatomy and Histology; Elsevier: Amsterdam, The Netherlands, 2004. [Google Scholar]
- Ayuob, N.N.; Abdel-Tawab, H.S.; El-Mansy, A.A.; Ali, S.S. The protective role of musk on salivary glands of mice exposed to chronic unpredictable mild stress. J. Oral. Sci. 2019, 61, 95–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Z.; Rockey, D.C. Upregulation of the actin cytoskeleton via myocardin leads to increased expression of type 1 collagen. Lab. Investig. 2017, 97, 1412–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alali, Z.; Graham, A.; Swan, K.; Flyckt, R.; Falcone, T.; Cui, W. 60S acidic ribosomal protein P1 (RPLP1) is elevated in human endometriotic tissue and in a murine model of endometriosis and is essential for endometriotic epithelial cell survival in vitro. Mol. Hum. Reprod. 2020, 26, 53–64. [Google Scholar] [CrossRef]
- Sisto, M.; Lisi, S. The role of the epithelial-to-mesenchymal transition (EMT) in diseases of the salivary glands. Histochem. Cell Biol. 2018, 150, 133–147. [Google Scholar] [CrossRef]
- Takahashi-Horiuchi, Y.; Sugiyama, K.; Sakashita, H.; Amano, O. Expression of heat shock protein 27 with the transition from proliferation to differentiation of acinar precursor cell in regenerating submandibular gland of rats. Tohoku J. Exp. Med. 2008, 214, 221–230. [Google Scholar] [CrossRef] [Green Version]
- Porcheri, C.; Mitsiadis, T.A. Physiology, Pathology and Regeneration of Salivary Glands. Cells 2019, 8, 976. [Google Scholar] [CrossRef] [Green Version]
- Amano, O.; Mizobe, K. Anatomy and histology of rodent and human major salivary glands—Overview of the japan salivary gland society-sponsored workshop. Acta Histochem. Cytochem. 2012, 45, 241–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Council, N.R. Guide for the Care and Use of Laboratory Animals, 8th ed.; The National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. PLoS Biol. 2020, 18, e3000410. [Google Scholar]
- Amado, L.L.; Garcia, M.L.; Ramos, P.B.; Freitas, R.F.; Zafalon, B.; Ferreira, J.L.R.; Yunes, J.S.; Monserrat, J.M. A method to measure total antioxidant capacity against peroxyl radicals in aquatic organisms: Application to evaluate microcystins toxicity. Sci. Total Environ. 2009, 407, 2115–2123. [Google Scholar] [CrossRef]
- Esterbauer, H.; Cheeseman, K.H. Determination of aldehydic lipid peroxidation products: Malonaldehyde and 4-hydroxynonenal. Methods Enzymol. 1990, 186, 407–421. [Google Scholar] [PubMed]
- Dionizio, A.S.; Melo, C.G.S.; Arias-Sabino, I.T.; Ventura, T.M.S.; Leite, L.A.; Souza, S.R.G.; Santos, E.X.; Heubel, A.D.; Souza, J.G.; Perles, J.V.C.M.; et al. Chronic treatment with fluoride affects the jejunum: Insights from proteomics and enteric innervation analysis. Sci. Rep. 2018, 8, 3180. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Lima-Leite, A.; Lobo, J.G.V.M.; Pereira, H.A.B.S.; Fernandes, M.S.; Martini, T.; Zucki, F.; Sumida, D.H.; Rigalli, A.; Buzalaf, M.A.R. Proteomic analysis of gastrocnemius muscle in rats with streptozotocin-induced diabetes and chronically exposed to fluoride. PLoS ONE 2014, 9, e106646. [Google Scholar]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [Green Version]
- R Development Core Team. R: A Language and Environment for Statistical Computing. 2013. Available online: https://www.gbif.org/tool/81287/r-a-language-and-environment-for-statistical-computing (accessed on 11 November 2021).
- Alhamdoosh, M.; Ng, M.; Wilson, N.J.; Sheridan, J.M.; Huynh, H.; Wilson, M.J.; Ritchie, M.E. Combining multiple tools outperforms individual methods in gene set enrichment analyses. Bioinformatics 2017, 33, 414–424. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Benner, M.J.; Hancock, R.E.W. NetworkAnalyst-integrative approaches for protein–protein interaction network analysis and visual exploration. Nucleic Acids Res. 2014, 42, W167–W174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohl, L.; Merlo, C.; Carda, C.; Gomez de Ferrari, M.E.; Carranza, M. Morphometric analysis of the parotid gland affected by alcoholic sialosis. J. Oral. Pathol. Med. 2008, 37, 499–503. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measures | Parotid Glands | Submandibular Glands | ||||
|---|---|---|---|---|---|---|
| Control Group | Aluminum Group | p Value * | Control Group | Aluminum Group | p Value * | |
| Total parenchyma area (µm2) | 64,563 ± 2613 | 58,262 ± 1100 | 0.0397 | 69,726 ± 1713 | 54,543 ± 1564 | 0.0001 |
| Total stroma area (µm2) | 21,980 ± 634 | 28,803 ± 734.7 | 0.0011 | 4941 ± 1099 | 24,263 ± 1473 | <0.0001 |
| Total acinar area (µm2) | 62,676 ± 3062 | 55,020 ± 426.7 | 0.0326 | 66,660 ± 2079 | 50,606 ± 1911 | 0.0006 |
| Total ductal area (µm2) | 2325 ± 611.8 | 2159 ± 279.7 | 0.7852 | 3002 ± 285.9 | 4022 ± 255.2 | 0.0312 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Souza-Monteiro, D.; Guerra, M.C.d.S.; Bittencourt, L.O.; Aragão, W.A.B.; Dionizio, A.; Silveira, F.M.; Buzalaf, M.A.R.; Martins, M.D.; Crespo-Lopez, M.E.; Lima, R.R. Salivary Glands after Prolonged Aluminum Exposure: Proteomic Approach Underlying Biochemical and Morphological Impairments in Rats. Int. J. Mol. Sci. 2022, 23, 2251. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23042251
Souza-Monteiro D, Guerra MCdS, Bittencourt LO, Aragão WAB, Dionizio A, Silveira FM, Buzalaf MAR, Martins MD, Crespo-Lopez ME, Lima RR. Salivary Glands after Prolonged Aluminum Exposure: Proteomic Approach Underlying Biochemical and Morphological Impairments in Rats. International Journal of Molecular Sciences. 2022; 23(4):2251. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23042251
Chicago/Turabian StyleSouza-Monteiro, Deiweson, Márcia Cristina dos Santos Guerra, Leonardo Oliveira Bittencourt, Walessa Alana Bragança Aragão, Aline Dionizio, Felipe Martins Silveira, Marília Afonso Rebelo Buzalaf, Manoela Domingues Martins, Maria Elena Crespo-Lopez, and Rafael Rodrigues Lima. 2022. "Salivary Glands after Prolonged Aluminum Exposure: Proteomic Approach Underlying Biochemical and Morphological Impairments in Rats" International Journal of Molecular Sciences 23, no. 4: 2251. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23042251