
β-Catenin-Specific Inhibitor, iCRT14, Promotes BoHV-1 Infection-Induced DNA Damage in Human A549 Lung Adenocarcinoma Cells by Enhancing Viral Protein Expression


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
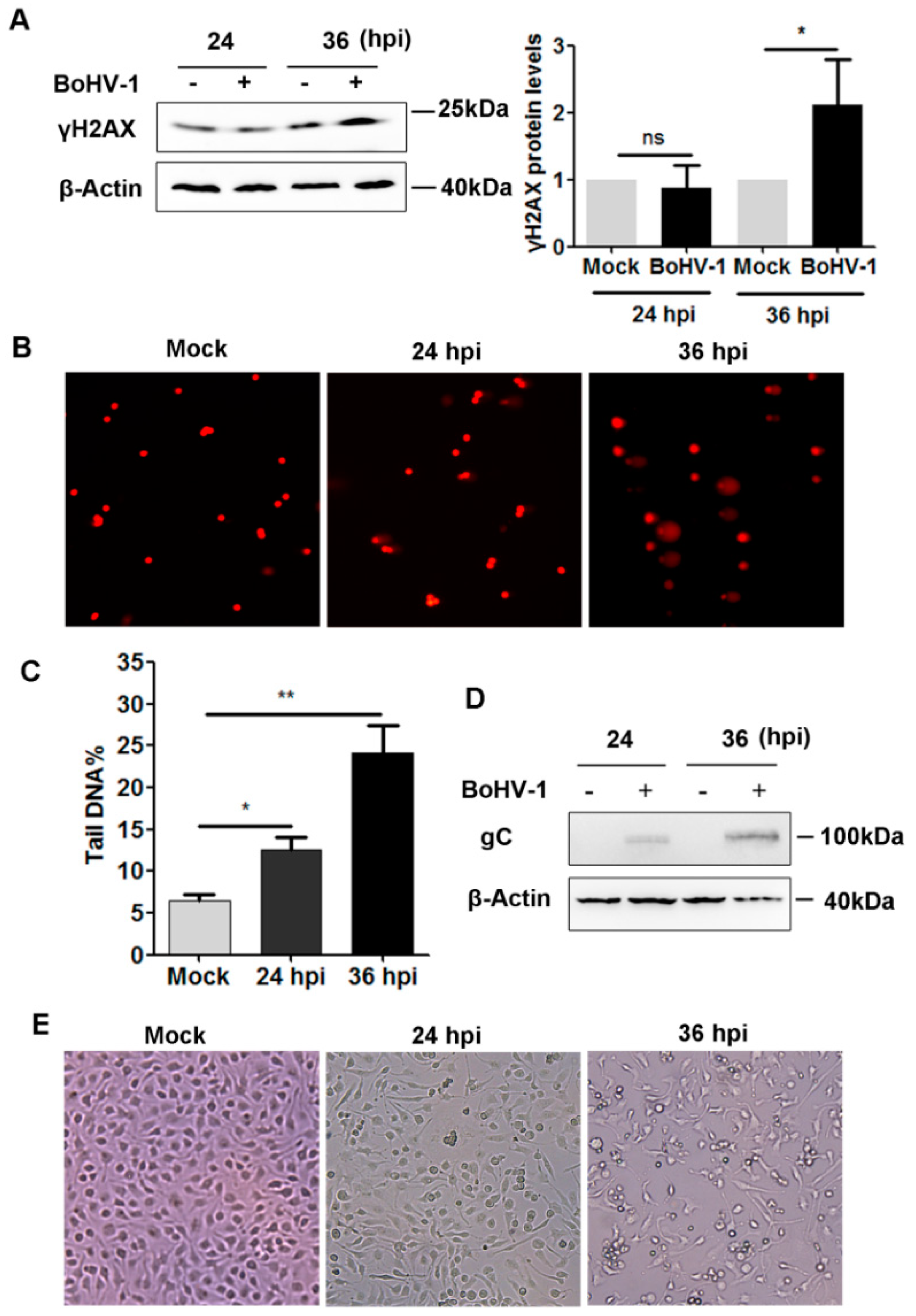
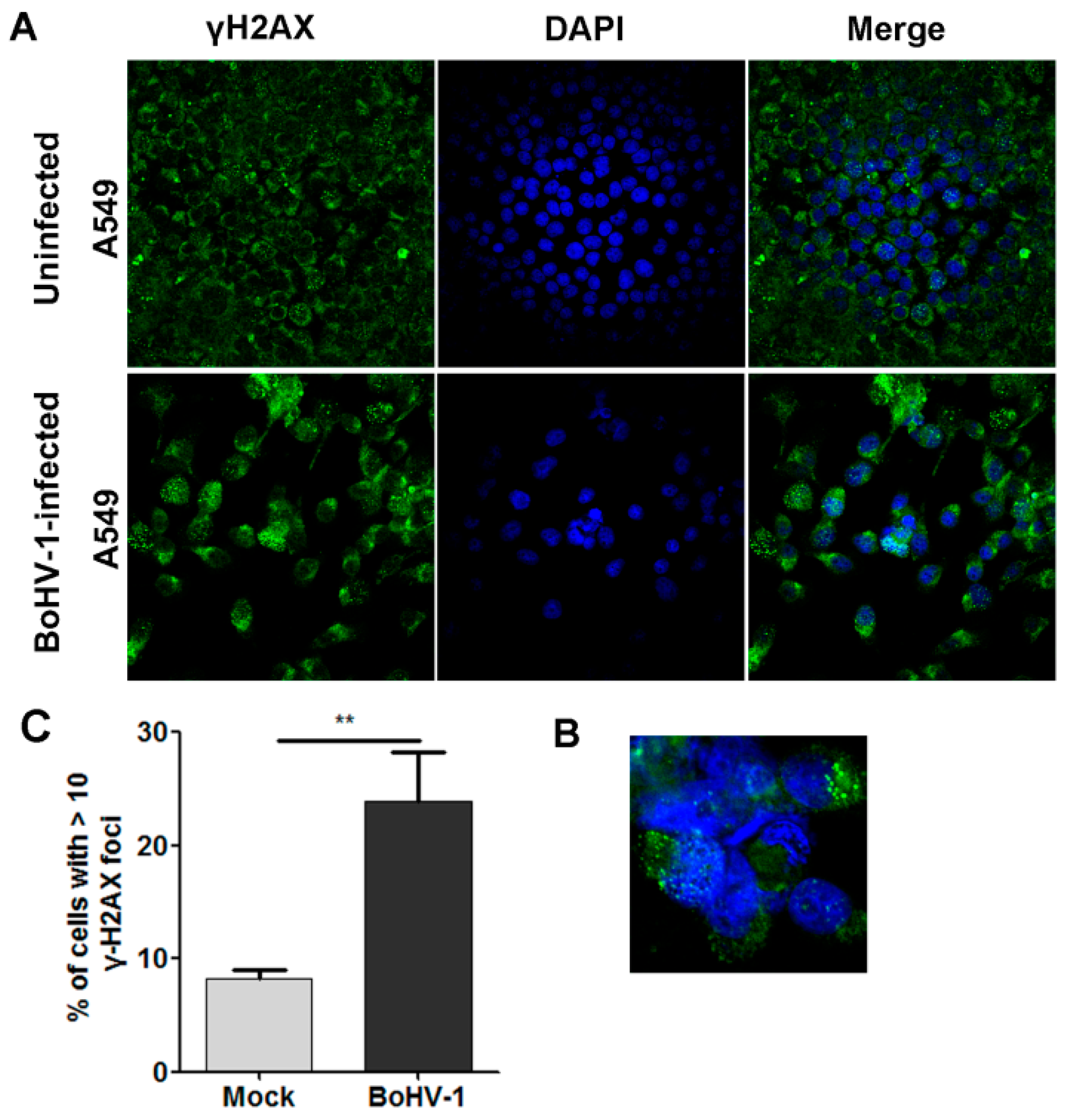
2.1. BoHV-1 Infection Induced γH2AX foci, a Hallmark of DNA Damage in A549 Cells
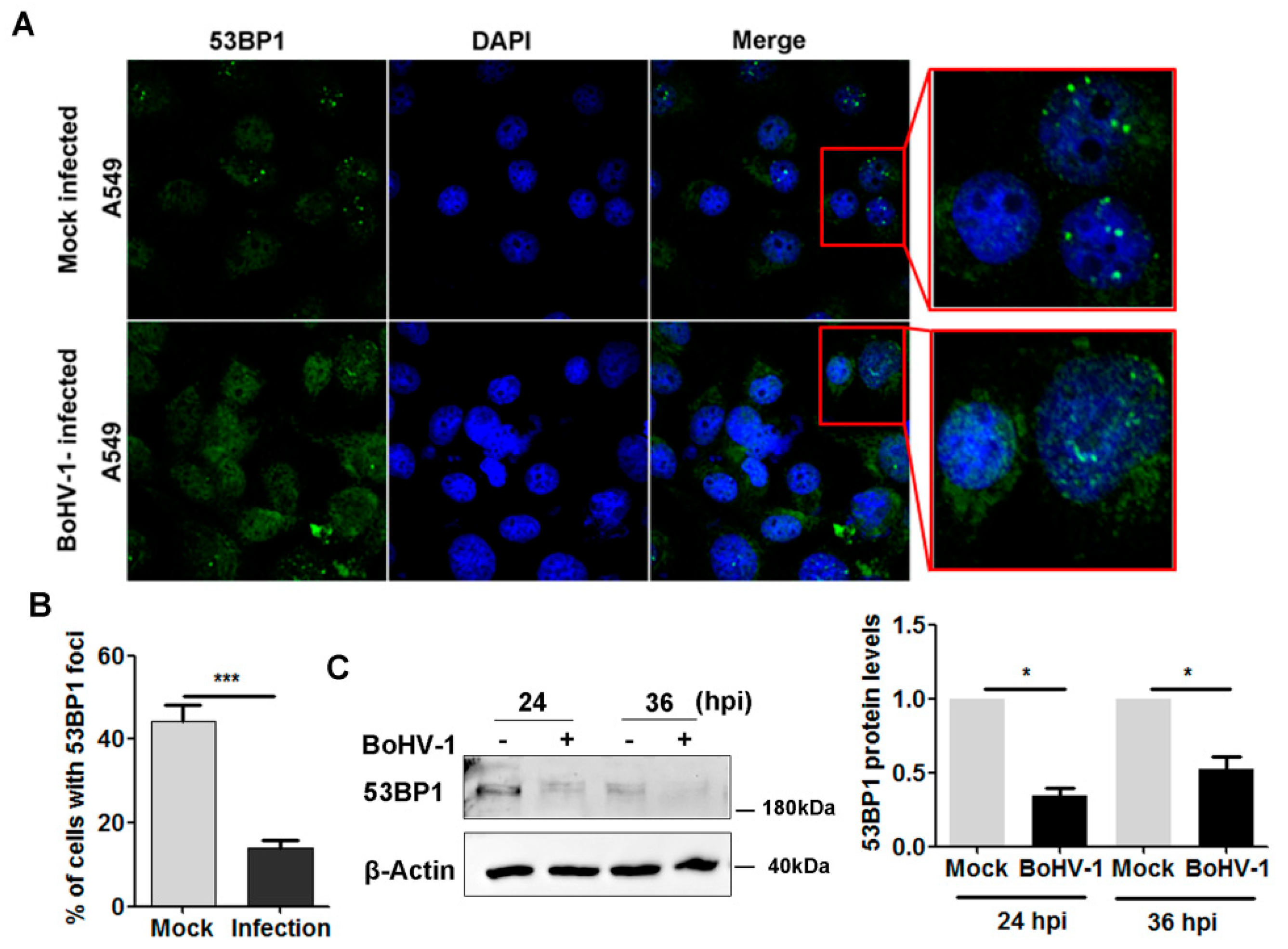
2.2. BoHV-1 Infection Leads to Inhibition of 53BP1 Signaling
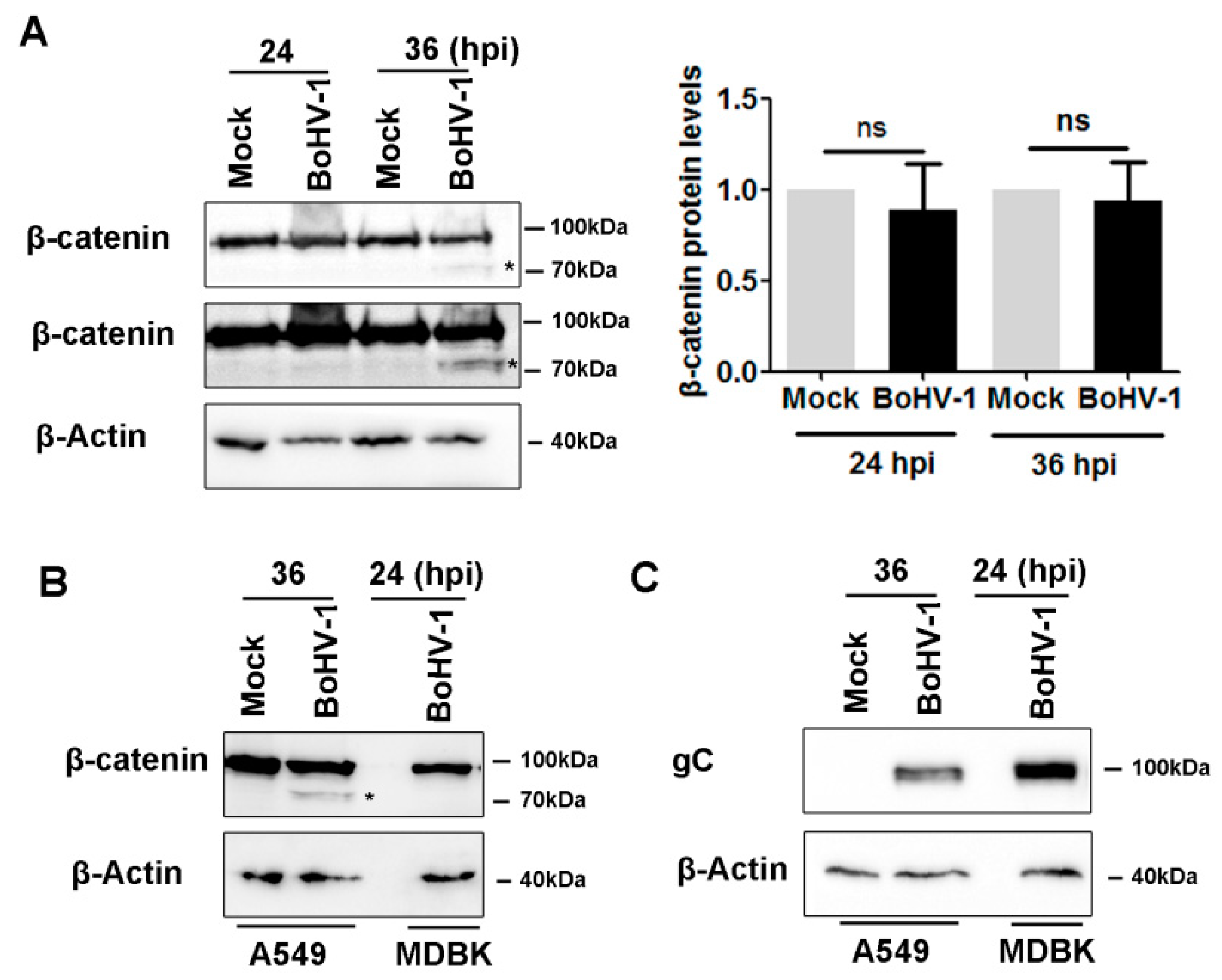
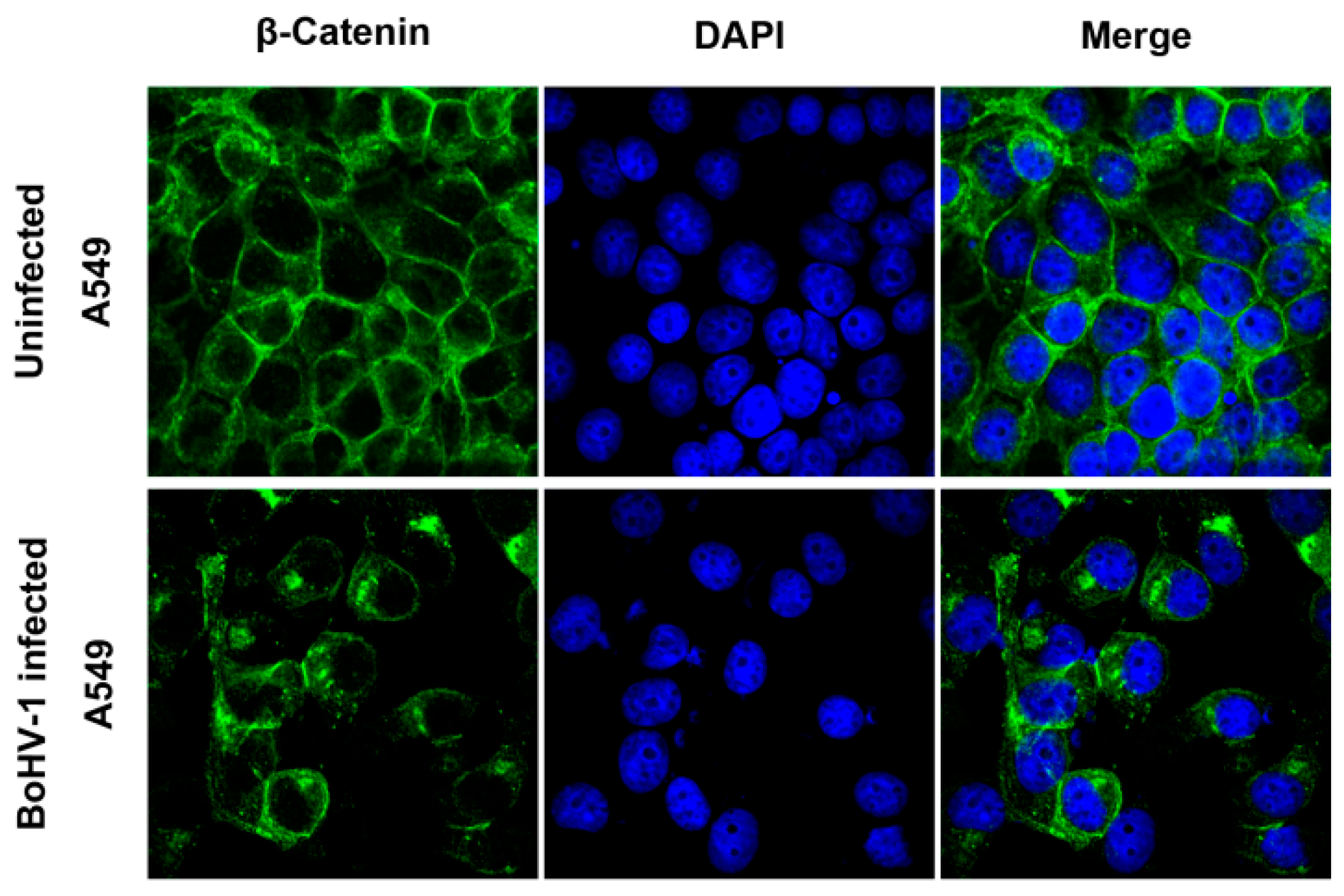
2.3. BoHV-1 Infection Alters β-Catenin Subcellular Localization
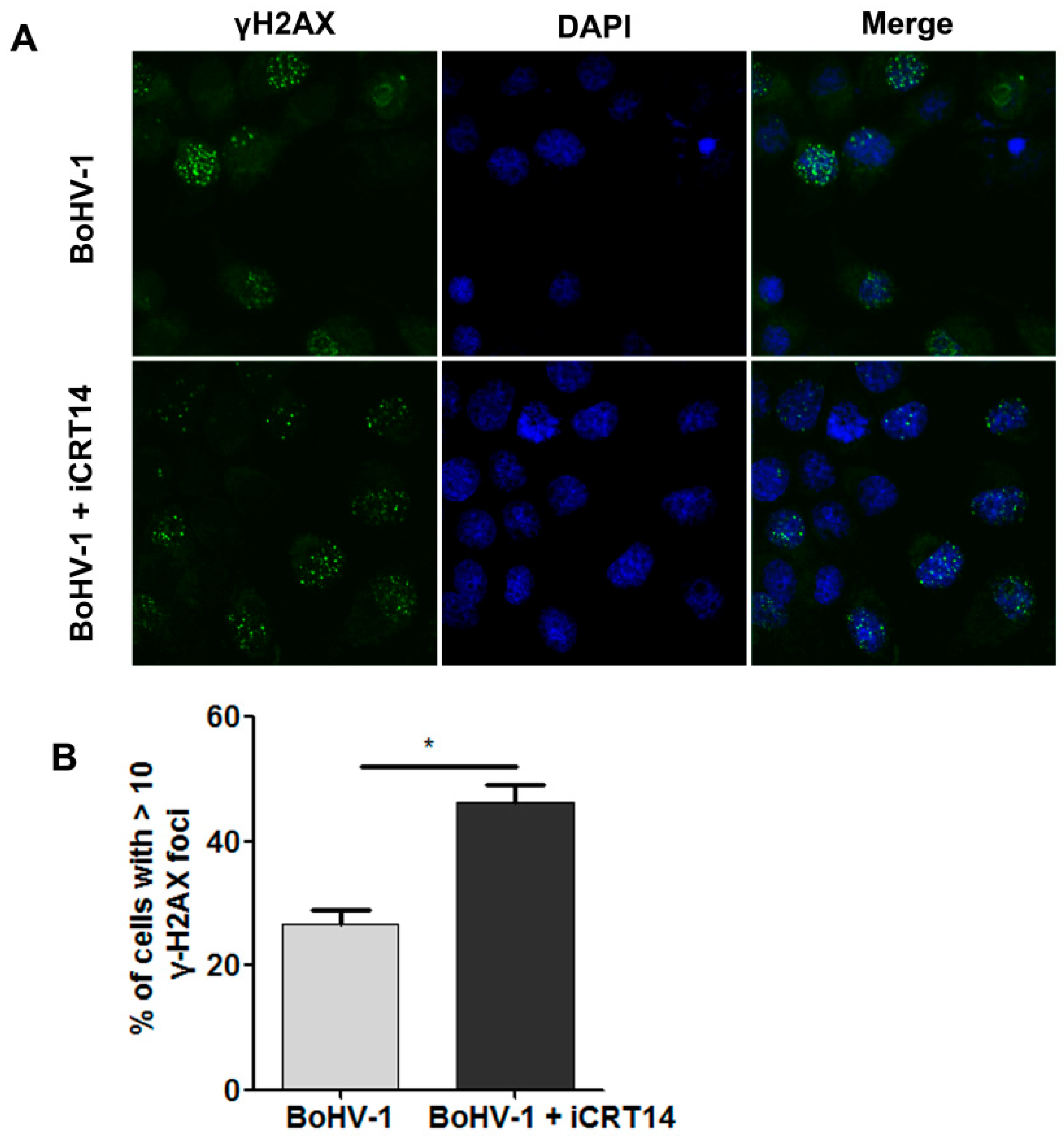
2.4. β-Catenin-Specific Inhibitor iCRT14 Promotes BoHV-1 Infection-Induced DNA Damage
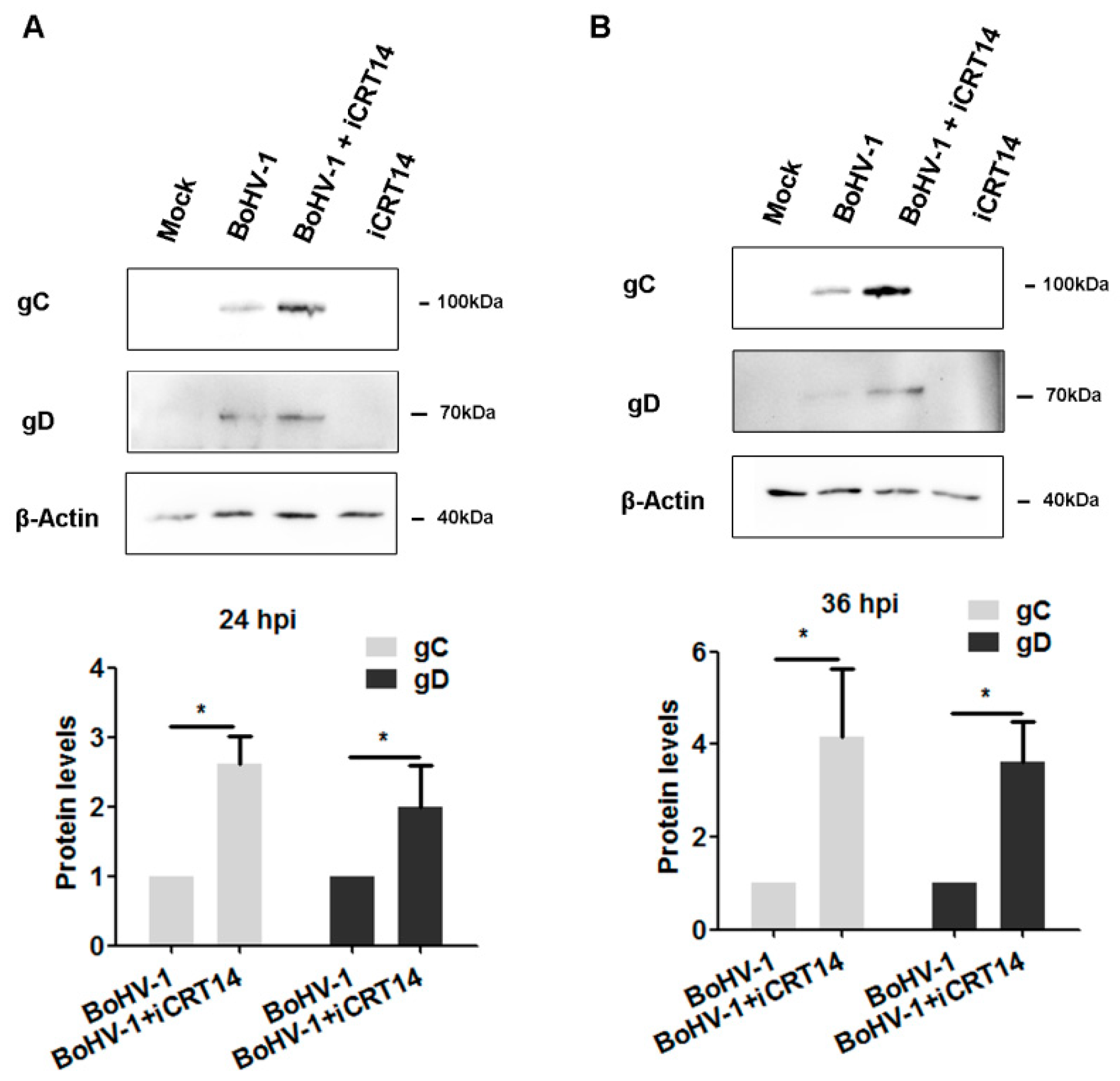
2.5. β-catenin-Specific Inhibitor iCRT14 Promotes Viral Gene Expression
3. Discussion
4. Materials and Methods
4.1. Cells and Virus
4.2. Antibodies
4.3. Western Blotting Analysis
4.4. Immunofluorescence Assay (IFA)
4.5. Comet Assay
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jones, C.; Chowdhury, S. A review of the biology of bovine herpesvirus type 1 (BHV-1), its role as a cofactor in the bovine respiratory disease complex and development of improved vaccines. Anim. Health Res. Rev. 2007, 8, 187–205. [Google Scholar] [CrossRef] [PubMed]
- Fiorito, F.; Irace, C.; Nocera, F.P.; Piccolo, M.; Ferraro, M.G.; Ciampaglia, R.; Tenore, G.C.; Santamaria, R.; De Martino, L. MG-132 interferes with iron cellular homeostasis and alters virulence of bovine herpesvirus 1. Res. Vet. Sci. 2021, 137, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, R.; Cuddington, B.; Mossman, K. Bovine herpesvirus type 1 as a novel oncolytic virus. Cancer Gene Ther. 2010, 17, 344–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuddington, B.P.; Mossman, K.L. Oncolytic bovine herpesvirus type 1 as a broad spectrum cancer therapeutic. Curr. Opin. Virol. 2015, 13, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Kafer, G.R.; Li, X.; Horii, T.; Suetake, I.; Tajima, S.; Hatada, I.; Carlton, P.M. 5-Hydroxymethylcytosine Marks Sites of DNA Damage and Promotes Genome Stability. Cell Rep. 2016, 14, 1283–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, L.W.; Dong, C.; Li, J.S.; Chen, J.; Yuan, J.S.; Huang, J.; Chan, K.M.; Yu, C.H.; Huen, M.S.Y. RNF169 limits 53BP1 deposition at DSBs to stimulate single-strand annealing repair. Proc. Natl. Acad. Sci. USA 2018, 115, Eb286–Eb295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhuppar, S.; Roy, S.; Mazumder, A. Gamma H2AX in the S Phase after UV Irradiation Corresponds to DNA Replication and Does Not Report on the Extent of DNA Damage. Mol. Cell Biol. 2020, 40, e00328-20. [Google Scholar] [CrossRef]
- Shang, Y.; Zhang, L.; Jiang, Y.T.; Li, Y.; Lu, P. Airborne quinones induce cytotoxicity and DNA damage in human lung epithelial A549 cells: The role of reactive oxygen species. Chemosphere 2014, 100, 42–49. [Google Scholar] [CrossRef]
- Qiu, W.C.; Ding, X.Y.; Li, S.T.; He, Y.M.; Zhu, L.Q. Oncolytic Bovine Herpesvirus 1 Inhibits Human Lung Adenocarcinoma A549 Cell Proliferation and Tumor Growth by Inducing DNA Damage. Int. J. Mol. Sci. 2021, 22, 8582. [Google Scholar] [CrossRef]
- Zhu, L.Q.; Fu, X.T.; Yuan, C.; Jiang, X.Y.; Zhang, G.P. Induction of Oxidative DNA Damage in Bovine Herpesvirus 1 Infected Bovine Kidney Cells (MDBK Cells) and Human Tumor Cells (A549 Cells and U2OS Cells). Viruses 2018, 10, 393. [Google Scholar] [CrossRef] [Green Version]
- Mirza-Aghazadeh-Attari, M.; Mohammadzadeh, A.; Yousefi, B.; Mihanfar, A.; Karimian, A.; Majidinia, M. 53BP1: A key player of DNA damage response with critical functions in cancer. DNA Repair 2019, 73, 110–119. [Google Scholar] [CrossRef]
- Fradet-Turcotte, A.; Canny, M.D.; Escribano-Diaz, C.; Orthwein, A.; Leung, C.C.Y.; Huang, H.; Landry, M.C.; Kitevski-LeBlanc, J.; Noordermeer, S.M.; Sicheri, F.; et al. 53BP1 is a reader of the DNA-damage-induced H2A Lys 15 ubiquitin mark. Nature 2013, 499, 50–54. [Google Scholar] [CrossRef] [Green Version]
- Drane, P.; Brault, M.E.; Cui, G.F.; Meghani, K.; Chaubey, S.; Detappe, A.; Parnandi, N.; He, Y.Z.; Zheng, X.F.; Botuyan, M.V.; et al. TIRR regulates 53BP1 by masking its histone methyl-lysine binding function. Nature 2017, 543, 211–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, I.M.; Minn, K.; Jorda, K.G.; Chen, J.J. Accumulation of checkpoint protein 53BP1 at DNA breaks involves its binding to phosphorylated histone H2AX. J. Biol. Chem. 2003, 278, 19579–19582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morin, P.J. Beta-catenin signaling and cancer. BioEssays News Rev. Mol. Cell. Dev. Biol. 1999, 21, 1021–1030. [Google Scholar] [CrossRef]
- Zhu, L.Q.; Thunuguntla, P.; Liu, Y.L.; Hancock, M.; Jones, C. The beta-catenin signaling pathway stimulates bovine herpesvirus 1 productive infection. Virology 2017, 500, 91–95. [Google Scholar] [CrossRef]
- Workman, A.; Zhu, L.Q.; Keel, B.N.; Smith, T.P.L.; Jones, C. The Wnt Signaling Pathway Is Differentially Expressed during the Bovine Herpesvirus 1 Latency-Reactivation Cycle: Evidence That Two Protein Kinases Associated with Neuronal Survival, Akt3 and BMPR2, Are Expressed at Higher Levels during Latency. J. Virol. 2018, 92, e01937-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karimaian, A.; Majidinia, M.; Baghi, H.B.; Yousefi, B. The crosstalk between Wnt/beta-catenin signaling pathway with DNA damage response and oxidative stress: Implications in cancer therapy. DNA Repair 2017, 51, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.Y.; Yi, J.; Tao, L.L.; Huang, G.C.; Chu, X.Y.; Song, H.Z.; Chen, L.B. Wnt signaling induces radioresistance through upregulating HMGB1 in esophageal squamous cell carcinoma. Cell Death Dis. 2018, 9, 433. [Google Scholar] [CrossRef] [PubMed]
- Spitzner, M.; Emons, G.; Schutz, K.B.; Wolff, H.A.; Rieken, S.; Ghadimi, B.M.; Schneider, G.; Grade, M. Inhibition of Wnt/beta-Catenin Signaling Sensitizes Esophageal Cancer Cells to Chemoradiotherapy. Int. J. Mol. Sci. 2021, 22, 10301. [Google Scholar] [CrossRef]
- Mah, L.J.; El-Osta, A.; Karagiannis, T.C. Gamma H2AX: A sensitive molecular marker of DNA damage and repair. Leukemia 2010, 24, 679–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panier, S.; Boulton, S.J. Double-strand break repair: 53BP1 comes into focus. Nat. Rev. Mol. Cell Biol. 2014, 15, 7–18. [Google Scholar] [CrossRef]
- Ji, Y.; Lv, J.; Sun, D.; Huang, Y.F. Therapeutic strategies targeting Wnt/beta-catenin signaling for colorectal cancer. Int. J. Mol. Med. 2022, 49, 1. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.Q.; Yuan, C.; Zhang, D.; Ma, Y.; Ding, X.Y.; Zhu, G.Q. BHV-1 induced oxidative stress contributes to mitochondrial dysfunction in MDBK cells. Vet. Res. 2016, 47, 47. [Google Scholar] [CrossRef] [PubMed]
- Sandri-Goldin, R.M. The many roles of the highly interactive HSV protein ICP27, a key regulator of infection. Future Microbiol. 2011, 6, 1261–1277. [Google Scholar] [CrossRef] [PubMed]
- Saira, K.; Zhou, Y.; Jones, C. The Infected Cell Protein 0 Encoded by Bovine Herpesvirus 1 (bICP0) Associates with Interferon Regulatory Factor 7 and Consequently Inhibits Beta Interferon Promoter Activity. J. Virol. 2009, 83, 3977–3981. [Google Scholar] [CrossRef] [Green Version]
- Qiu, W.C.; Chang, L.; He, Y.M.; Zhu, L.Q. The Role of Epidermal Growth Factor Receptor Signaling Pathway during Bovine Herpesvirus 1 Productive Infection in Cell Culture. Viruses 2020, 12, 927. [Google Scholar] [CrossRef] [PubMed]
- Shackelford, J.; Maier, C.; Pagano, J.S. Epstein-Barr virus activates beta-catenin in type III latently infected B lymphocyte lines: Association with deubiquitinating enzymes. Proc. Natl. Acad. Sci. USA 2003, 100, 15572–15576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabatkova, M.; Zapletal, O.; Tylichova, Z.; Neca, J.; Machala, M.; Milcova, A.; Topinka, J.; Kozubiik, A.; Vondracek, J. Inhibition of beta-catenin signalling promotes DNA damage elicited by benzo[a]pyrene in a model of human colon cancer cells via CYP1 deregulation. Mutagenesis 2015, 30, 565–576. [Google Scholar] [CrossRef] [Green Version]
- Hanon, E.; Keil, G.; van Drunen Littel-van den Hurk, S.; Griebel, P.; Vanderplasschen, A.; Rijsewijk, F.A.; Babiuk, L.; Pastoret, P.P. Bovine herpesvirus 1-induced apoptotic cell death: Role of glycoprotein D. Virology 1999, 257, 191–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geiser, V.; Rose, S.; Jones, C. Bovine herpesvirus type 1 induces cell death by a cell-type-dependent fashion. Microb. Pathog. 2008, 44, 459–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Yu, Y.; Jiang, X.; Yuan, W.; Zhu, G. First report of bovine herpesvirus 1 isolation from bull semen samples in China. Acta Virol. 2017, 61, 483–486. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, P.D.; Aich, P.; Manuja, A.; Hokamp, K.; Roche, F.M.; Brinkman, F.S.; Potter, A.; Babiuk, L.A.; Griebel, P.J. Effect of stress on viral-bacterial synergy in bovine respiratory disease: Novel mechanisms to regulate inflammation. Comp. Funct. Genom. 2005, 6, 244–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, X.; Yuan, W.; Yang, H.; Liu, C.; Li, S.; Zhu, L. β-Catenin-Specific Inhibitor, iCRT14, Promotes BoHV-1 Infection-Induced DNA Damage in Human A549 Lung Adenocarcinoma Cells by Enhancing Viral Protein Expression. Int. J. Mol. Sci. 2022, 23, 2328. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23042328
Ding X, Yuan W, Yang H, Liu C, Li S, Zhu L. β-Catenin-Specific Inhibitor, iCRT14, Promotes BoHV-1 Infection-Induced DNA Damage in Human A549 Lung Adenocarcinoma Cells by Enhancing Viral Protein Expression. International Journal of Molecular Sciences. 2022; 23(4):2328. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23042328
Chicago/Turabian StyleDing, Xiuyan, Weifeng Yuan, Hao Yang, Chang Liu, Shitao Li, and Liqian Zhu. 2022. "β-Catenin-Specific Inhibitor, iCRT14, Promotes BoHV-1 Infection-Induced DNA Damage in Human A549 Lung Adenocarcinoma Cells by Enhancing Viral Protein Expression" International Journal of Molecular Sciences 23, no. 4: 2328. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23042328