
Enzymes-Assisted Extraction of Plants for Sustainable and Functional Applications


Lithuanian Research Centre for Agriculture and Forestry, Institute of Horticulture, 54333 Babtai, Lithuania
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2022, 23(4), 2359; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23042359
Submission received: 12 January 2022
/
Revised: 9 February 2022
/
Accepted: 17 February 2022
/
Published: 21 February 2022
(This article belongs to the Special Issue Health Properties of Plant Bioactive Compounds: Immune, Antioxidant and Metabolic Effects)
Abstract
:The scientific community and industrial companies have discovered significant enzyme applications to plant material. This rise imparts to changing consumers’ demands while searching for ‘clean label’ food products, boosting the immune system, uprising resistance to bacterial and fungal diseases, and climate change challenges. First, enzymes were used for enhancing production yield with mild and not hazardous applications. However, enzyme specificity, activity, plant origin and characteristics, ratio, and extraction conditions differ depending on the goal. As a result, researchers have gained interest in enzymes’ ability to cleave specific bonds of macroelements and release bioactive compounds by enhancing value and creating novel derivatives in plant extracts. The extract is enriched with reducing sugars, phenolic content, and peptides by disrupting lignocellulose and releasing compounds from the cell wall and cytosolic. Nonetheless, depolymerizing carbohydrates and using specific enzymes form and release various saccharides lengths. The latest studies show that oligosaccharides released and formed by enzymes have a high potential to be slowly digestible starches (SDS) and possibly be labeled as prebiotics. Additionally, they excel in new technological, organoleptic, and physicochemical properties. Released novel derivatives and phenolic compounds have a significant role in human and animal health and gut-microbiota interactions, affecting many metabolic pathways. The latest studies have contributed to enzyme-modified extracts and products used for functional, fermented products development and sustainable processes: in particular, nanocellulose, nanocrystals, nanoparticles green synthesis with drug delivery, wound healing, and antimicrobial properties. Even so, enzymes’ incorporation into processes has limitations and is regulated by national and international levels.
1. Introduction
The demand for new and natural compounds, ‘clean label’ trend, rising drug resistance, holistic wellbeing approach of the post-pandemic period, and sustainable living has intensified the development of plant-derived compounds called biologically active components [1,2,3]. Biologically active substances bind by interaction or binding to specific receptors in stem cells, improving a particular physiological function of the body. Unfortunately, many such compounds are present in cytosolic cell spaces and plant cell walls [4]. Many extraction methods cannot achieve these compounds and thus obtain the highest components yields. That is why enzymes incorporation in various extractions is currently one of the few methods to provide this result. Enzymes with specific hydrolytic properties are used to degrade this matrix to gain access to biologically active components from cytosolic spaces and cell walls [5]. Nevertheless, the global market for industrial enzymes is expected to grow up to $9.2 billion by 2027 [6]. One of the advantages of the usage of enzymes is that they can be added to hydrophilic and multi-step lipophilic extractions, especially for by-product valorization [7]. For example, in Europe, grain, fruit, and vegetable food loss from post-harvest to distribution varies from 20, 41, and 46%, respectively [7,8]. However, enzyme-assisted incorporation increases phenolic content in lipophilic extracts, which is potently applicable for nutraceuticals or pharmaceuticals. However, limitations in the safety of by-products have risen, and greater attention is given to this topic [8]. In comparison, for hydrophilic extracts, enzymes efficiently increase the water-soluble content of novel derivatives applicable in food industries [9].
Due to the high demand for diverse health outcomes, functional food categories arise during the post-pandemic period, especially probiotic and prebiotic categories [10]. Incorporating these foods into human diets may reduce obesity. According to WHO, from 2018 to 2030, obese children will reach from 150 billion to 250 billion, respectively [11]. Gut microbiota modulates lipogenesis and cholesterol synthesis. Dysbiosis initiates higher absorption of sugars in the small intestines by modulating membrane transport [11,12]. Moreover, acetate, the metabolite made by the gut microbiome in the proper amount, can boost immune responses by promoting B10 cells, and in higher amounts can lead to adiposity [13,14,15]. These challenges invite scientists to search for sustainable and functional food development worldwide. One scope is enzymes, usually used for plant-based drinks production and syrups for saccharification, decreased viscosity, higher yield, and low toxicity in the food industry. However, lately, studies suggest that controlled enzyme-assisted extraction could lead to a higher and broader density of nutrients [16,17]. For example, dietary fibers with three or more monomeric units, phenolic compounds, and complexes can be suggested prebiotics and used for functional food development [18]. Chen et al. [19] investigated amylolytic and cellulolytic enzymes impact of releasing phenolic compounds and the correlation in solid-state fermentation with significant results for increased phenolic compounds content and antioxidant activities [19].
Because the plant material is complex, with varied compositions and matrices, enzymes are used in mixtures or cocktails. Besides releasing secondary metabolites and small peptides, they cleave long-chain molecules into shorter ones. Likewise, these substances are soluble in the solvent and can enhance organoleptic, technological, and functional properties. Moreover, enzymatic extraction methods are characterized by mild reaction conditions, substrate specificity, industrial applicability, and many other advantages [20]. These extracts may be used continuously in many fields and, surprisingly, in green synthesis development [16,21,22].
Green nanoparticle synthesis in aqueous plant extracts has increased over the last decade. Scientific discussion and research indicate the appropriate size of nanoparticles with high potential antimicrobial properties, involving the most common pathogenic bacteria like Escherichia coli, Staphylococcus aureus, and widely spread, highly resistant Candida albicans [23,24]. Studies identify that phenolic compounds and sugars play an essential capping and stabilizing role in green nanoparticles synthesis, and enzymes incorporation could increase the synthesized media yield with economically friendly conditions [25,26].
This review briefly suggests scientific approaches of commercially used hydrolases and carbohydrases for various plant materials to extract functional ingredients, products development, and possible applications.
2. Carbohydrases and Phenolic Compounds in Plants
In this review, carbohydrases and hydrolases get a more profound overview due to the plant cell wall mainly consisting of various carbohydrates, trapping active biological components. The cell contains various linear heterogeneous polymeric carbohydrates homologous to cellulose, such as xyloglucans and mannan, and hemicellulose is covalently linked to cellulose microfibrils and lignin to form complex structural branches. This multi-component structure in the plant cell wall is called lignocellulose. However, plant polymeric substances are usually categorized to waste. The global agricultural sector is estimated to produce 5 × 109 tons of plant-derived biomass each year, where the total amount of lignocellulosic waste is about 2 × 1011 tons per year [27,28]. The structure of lignocellulose gives the stability and resistance of the cell to the extraction of internal cellular components (Figure 1), where various enzymatic activities are required to degrade all the different forms of hemicellulose.
Enzymes are derived from bacteria, fungi, yeasts, archaea, animal organs, or plant extracts. However, microbial enzymes are more stable compared to ones having a plant or animal origin. Moreover, the production of the enzyme during microbial fermentation is cost-effective and easily adapted to modifications and high purity [31]. Carbohydrases can be categorized in starch-degrading enzymes: amylases and glucoamylases; and non-starch polysaccharides (NSP) catalyzing enzymes with cellulolytic, pectinolytic xylanolytic activities [31,32,33]. In general, NSP enzymes also can be named xylases, cellulases, and pectinases due to being composed of glycoside hydrolases, carbohydrate esterases, xylanases, etc. (Table 1).
NSP enzymes are preferred as a part of commercial enzyme mixture, thus ensuring complete lysis of cell walls while contributing to a cost-effective means [5,34]. Various fungi, including Trichoderma sp. and Aspergillus sp., produce carbohydrate-hydrolyzing enzymes. For many years, Trichoderma sp. has been extensively studied for high cellulase production [35]. However, most strains of Trichoderma are known to have low β-glucosidase activity, which causes cellobiose accumulation. Although much effort has been made to obtain T. reesei mutants by classical mutagenicity, such as RUT-C30, the relatively low activity of β-glucosidase remains one of the significant barriers to efficient cellulose hydrolysis [36]. Aspergillus sp. is important in xylanase production, and the latest studies showed UV-irradiated Aspergillus mutants for a higher yield of enzymes [37,38]. Endoxynalases, specifically endo-β-1,4-xynalases, are the most important, depolymerizing xylan polymer into small branches. Xylooligosaccharides later are converted to xylotriose, xylobiose, and xylose [33]. Another essential component of the cell wall is pectin. It is a polymer of α-D-galacturonate and L-rhamnose units linked to α-1,4 or 1,2 to form so-called pectic elbows. Pectin, associated with cellulose, imparts stiffness and cohesion to cell walls [5,34]. Galactose, mannose, fucose, arabinose, xylose, and L-methyl, O-acetyl groups all these components make four main regions of pectin structure: rhamnogalacturonan I (RG-I), rhamnogalacturonan II (RG-II), homogalacturonan (HG), and xylogalacturonan (XG), which are involved in reducing inflammatory processes in human [39]. Pectinolytic enzymes or pectinases were the first enzymes commercially available in wine and fruit juices, although the cell wall structure was only determined later [31]. They consist of three main classes: protopectinases, esterases, and depolymerases. Protopectinases occur naturally and are responsible for dissolving otherwise undissolved protopectins from immature fruit during maturation. Esterases or pectin methylesterases remove esterified units to remove methoxy esters. Depolymerases are represented by lyses and hydrolases that catalyze the fragmentation of glycosidic bonds. Today, pectinases are used in the fruit juice industry because of their effectiveness: higher juice yield, filterability, lower viscosity, and increased transparency. Pectinolytic enzymes have been found to be particularly effective in the extraction of polyphenols, particularly in the release of anthocyanidins from glycosides. Many pectinolytic mixtures sold today contain all three types of the above classes and a mixture with cellulases and hemicellulases to achieve an overall synergistic effect [5,40,41].
In general, biological raw material systems range from 5000 to 25,000 individual phytochemicals that can have biological activity. Biologically active substances are metabolites synthesized in plants that perform plant protection and other functions. There is growing evidence that biologically active substances can help maintain optimal health and reduce the risk of chronic diseases such as cancer, cardiovascular disease, stroke, and Alzheimer’s disease (AD) [42,43]. For example, quercetin found in pines, buckwheats, and many other plants has a high binding link through hydrogen bonds to the cyclin-dependent kinase 6 (CDK6) and inhibits its activity, which plays an essential role in the progression of different types of cancer [44]. However, phenolic compounds are usually soluble conjugates (glycosides) or insoluble forms (phenolic acids) covalently linked to carbohydrate radicals or cell wall structural components such as cellulose, hemicellulose, lignin, and pectin. In the insoluble form, the phenolic compounds are linked to the structural cell wall components by covalent bonds. Phenolic acids bind to lignin through the hydroxyl groups of their aromatic rings and various polysaccharides and other proteins through ester linkages. They perform a protective function. Significant interests are given to flavonoids which are found in the form of glycosides bound to sugar residues via –OH groups (O-glycosides) or via –C–C– bonds (C-glycosides) [44,45]. The latest article discovers the promising neuroprotective and memory-enhancer characteristics of flavonoid-rich food and plant extracts by increasing the functions of neurons and cell proliferation [46]. Flavonoids are involved in regulating kinase-signaling cascades, including PI3K/Akt, PKC, and MAPK pathways [46,47]. In the food industry, functional food components are used to replace preservatives due to their antioxidant and antimicrobial properties [48,49,50]. Unfortunately, solvent-based extraction of biologically active substances often suffers from low extraction efficiency, requires a long extraction time, and often leaves traces of organic solvents in the final product, which reduces the quality of the product [51,52]. Therefore, there is a need to develop optimized, step-by-step extraction methods for beneficial substances for each type of raw material. Therefore, detailed protocols are required to produce bioactive compounds, especially from plants where the cell wall may inhibit the extraction efficiency [7,49]. Enzymatic extraction of biologically active compounds from plants is a potential alternative to conventional solvent-based extraction methods. Enzymes are ideal catalysts for extracting, modifying, or synthesizing complex biologically active compounds of natural origin. Enzyme-assisted extraction is based on the inherent ability of enzymes to catalyze reactions with exceptional specificity and the ability to function under mild processing conditions in aqueous solutions [53]. However, parameters such as enzyme specificity, activity, botanical origin, pH, enzyme and substrate, liquid to solid ratios, temperature, and time are essential for obtaining the highest value results [54,55].
Carbohydrases-active enzymes database (CAZy; https://www.cazy.org/, accessed on 10 February 2022) classified five main classes: glycoside hydrolases, glycosyltransferases, polysaccharides lyases, carbohydrate esterases, and auxiliary activities [56]. A mixture of these enzymes has to be implemented into reactions on the results maintained to achieve. Ideal selected enzymes have high activity and regio-/stereo-selectivity [57]. Moreover, raw materials of plant origin consist of a complex system of various macro-/micro-components [29]. Plant origin, morphological part, cultivar, and growing conditions impact the storage of secondary metabolites. The approach of Woo et al. [58] of fifteen cultivars in oats showed the variation by cultivars and harvesting days of phenolic content from 3748.4 to 5700.0 mg/100 g [58]. Enzyme usage usually requires low temperatures, usually 50–60 °C; short extraction periods of up to several hours, waste-free production possibilities, reduced substrate specificity, allowing extraction of many bioactive compounds, otherwise, were made inaccessible. [45,59]. Examples of various plant material extraction are presented in Table 2. Additionally, the yield of the extract is often characterized by high quality and bioavailability. Gonzalez et al. [60] compared ultrasound-assisted extraction with enzyme-assisted extraction for anthocyanins from blackcurrants, where a significant difference was not evaluated. Major two Cyanidin 3–O–rutinoside (C3R) and Cyanidin 3–O–glucoside were extracted at similar quantities and accounted for up to 90%. C3R stimulates the mechanism of insulin secretion through INS-pancreatic β-cells by promoting calcium channels and activating the PLC-IP3 pathway [61].
Furthermore, the method also allows applying greener chemistry in the food industry and pharmaceutical companies to optimize purer ways to extract new compounds. Enzymes can disrupt specific bonds and interactions in cell walls and membranes, resulting in higher extraction yields of bioactive substances [40,67,68].
3. Enzymatically Treated Polysaccharides as Possible Functional Components
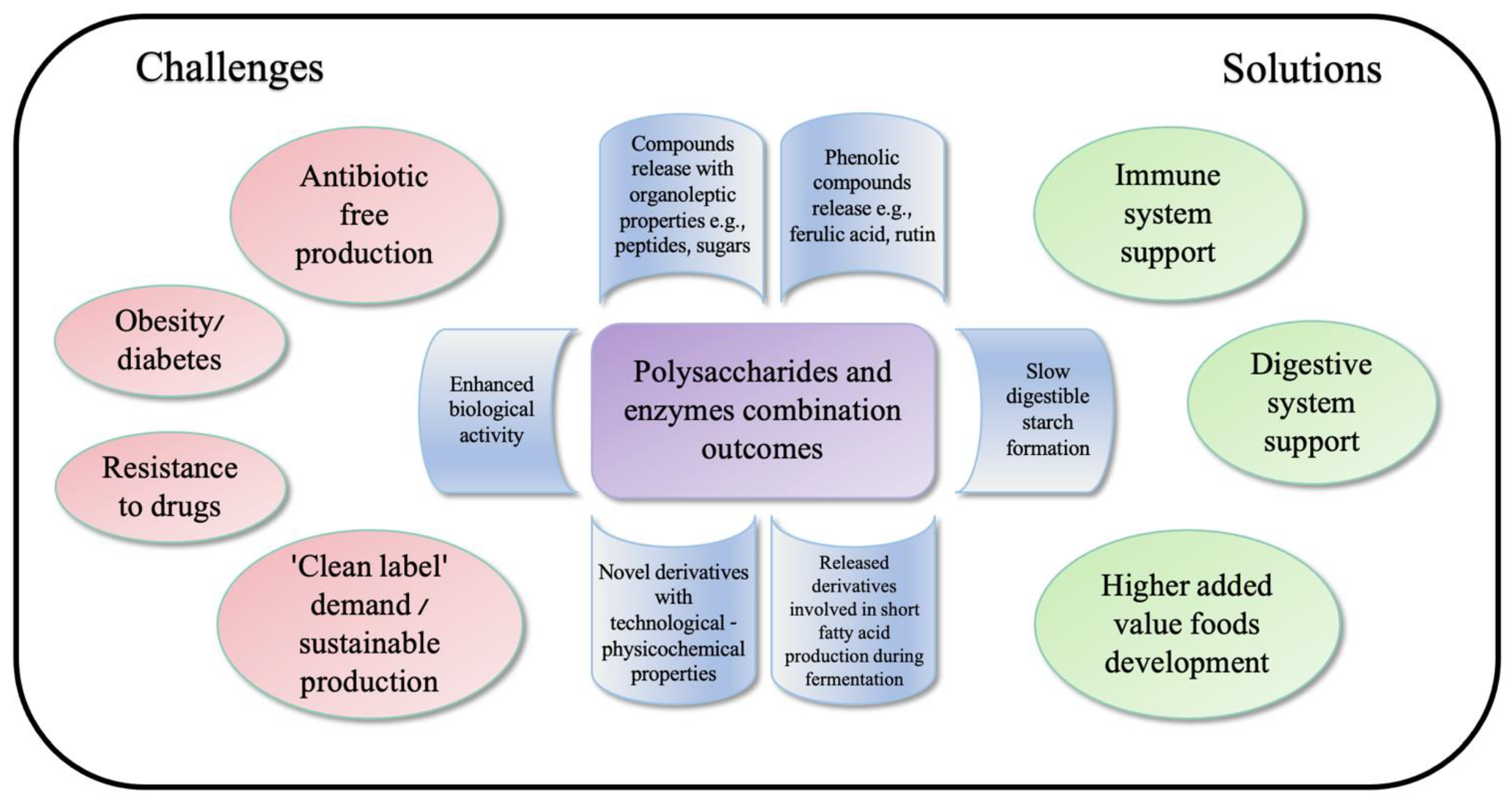
Lignocellulose and starch content differs depending on plant material. Grain materials usually consist of more starches because of the endosperm in the seed, whereas other plant parts, like brans, leaves, and others, consist of non-starch polysaccharides. Shah et al. [62] described three categories of starches: rapidly digestible starch (RDS), resistant starch (RS), and slowly digestible starch (SDS), which relate glucose release during digestion. Mainly, starches are found in cereals and pseudocereals where β-glucans are the most widely investigated non-starch polysaccharide and have enormous health-promoting properties [69]. However, β-glucans properties may vary depending on molar mass, which can be from 209 up to 2500 (kg mol−1) depending on the cultivar, variety, and the location of growth [70,71]. Nonetheless, β-glucans are found in fungi, yeasts, bacteria, and seaweed. The main difference is the ratio, length, and linkage between D-glucose of β-(1,3) and β-(1,4) for cereal grains β-(1,3) and β-(1,6) for fungi and yeasts [72]. For oats, novel oligosaccharides were produced by hydrolyzing β-glucans with dual enzyme-assisted hydrolysis using α-amylases and transglucosidase in result shorter α-(1,6)-branch linkage glucans, which stabilize the glucose release during the intestinal phase in vitro and may be prebiotic [73]. Another study in vitro also resulted in SDS increasing with corn starch using branching enzymes and maltogenic alpha-amylase [74]. Moreover, as a major non-starch polysaccharide in cereal grains, arabinoxylans are getting attention for their prebiotic properties, where enzymatically hydrolyzed fiber releases ferulic acid or combines with arabinose [75,76]. In general, dietary fibers, with more than three monomeric units known as prebiotics, are a growing trend for immune support through the microbiome and may prevent the uprising resistance to drugs for humans and animals [10]. Moreover, because antibiotic-free products are getting more attention from consumers, feed enhancement with dietary fibers may support the immune system of animals [19,75]. However, the clinical study by Wastyk et al. [77] for identifying humans‘ health marker variation while having the intervention of 10 weeks of high-fiber diet showed an increase in short-chain fatty acid (SCFAs), the density of microbes for higher protein content, and quantity but no diversity of microbiome [77]. It suggests that microbiota was insufficient to process increased fiber intake, even though glycan-degrading carbohydrate-active enzymes increased [77,78]. These studies implement the essential role of carbohydrate lengths for human diets. Animal studies with rats showed the xylan-oligosaccharides (XOS) as a potent dietary supplement of obesity prevalence. XOS promotes growth of Bifidobacterium strains involved in the development of obesity and insulin resistance [14,79]. In between, galacto-oligosaccharides (GOS) in studies with rats implement the potency of gut recoveries after alcohol withdrawal by a significant increase of butyric and propionic acid and the proliferation of Lactobacillus and Bifidobacterium strains [80]. Another animal study with GOS extracted from mulberry treated with β-mannanases showed that 200 mg/kg/day can initiate expression of crucial glycolysis enzymes such as GK, PK, and PCB and proteins p110 and Akt of key signaling intermediates, which results in the prevalence of diabetes and obesity [81]. A graphical scheme of summarized polysaccharides and enzymes combination results is presented in Figure 2.
4. Enzymes-Assisted Processes for Plant Materials
4.1. Bioactives Extraction from By-Products
In order to use the enzyme or their mixtures efficiently in extraction methods, it is essential to understand their catalytic mechanism of action and the optimal activity conditions for the recovery of individual biological raw materials and substances: e.g., a mixture of cellulose, pectin, and hemicellulose enzymes in a grapefruit peel during hydrolysis releases sugar into monomeric compounds that microorganisms can use later to produce ethanol and other fermentation products [82]. Another example is in tomatoes: lycopene is found mainly in the peel, giving it a red color. Carotenoids, especially lycopene, are one of the most potent antioxidants of plant origin, with a role in more than twenty different induced-signaling pathways and cell cycles described by Qi et al. [83]. According to scientific knowledge, lycopene is better absorbed from processed products than fresh tomatoes [84,85]. The digestive enzyme pancreatin is recommended before the solvent extraction of lycopene. Its use increased the yield of lycopene 2.5-fold compared to that obtained using the traditional extraction method [54]. Using Pectinex Ultra SP–L (P, pectinolytic enzyme), Celluclast (C, cellulolytic enzyme), and Viscozyme L commercial enzymes for solvent extractions, the yield of lycopene was increased as well as antioxidant activity compared with samples without enzyme mixtures [84]. Moreover, the scientific literature shows that in the stepwise extraction of buckwheat husk using xylanase commercial preparations, the yield of soluble fractions increased 4–5 times compared to the control extraction [7]. The use of enzymes reduces the amount of solvent required for extraction and increases the yield of extractable compounds. For example, tannase, pectase, cellulase, and hemicellulase are widely used in juices to increase product yield and improve quality [40,86]. However, when it is enzymatic extraction alone, compare with other extraction methods. Zheng et al. [87] described different extraction methods of palm kernel expeller. Dried extracts prepared with enzymes, hydrochloric acid, carboxymethylation, and hydroxipropylation were compared among chemical composition and physicochemical and functional properties. All the extract methods increased dietary content, phenolic content, and physicochemical properties. However, the goals of extract application have to be implemented because all extract methods were suitable, although enzyme assistance required fewer organic solvents, which implements in sustainability coverage [87].
Enzyme-assisted extraction (EAE) presents applicability to extract pectins from wastes and by–products by increasing plant cell wall permeability [88,89]. Enzymes are applicable to extract many phenolic compounds, including flavonoids and anthocyanidins [90]. Enzyme activity, treatment time, substrate ratio, and particle size are essential to get the highest efficiency during enzymatic treatment. Optimized conditions were discussed for pistachio green hull extraction by Yazdi et al. [84]. This research used cellulases, pectinases, tannases, and their mixtures for the extraction. Results indicated that all of the three enzymes at the same time used to extract phenolics were giving the highest score [91]. Moreover, by-products may be applied as enzymes production during solid-state fermentation (SSF). Some of the highest by-products of cacao are bean shell, brewers’ spent grain, and wheat bran, which have an estimated production of 140 million tons per year. These were implemented for SSF with Aspergillus awamori, Aspergillus niger, and Aspergillus oryzae, which produce feruloyl-esterases and amylases. These enzymes in bread enhanced ferulic acid quantity, total phenolic content, and antioxidant activity [92,93].
4.2. Plant-Based Drinks from Grains and Fermented Drinks Production
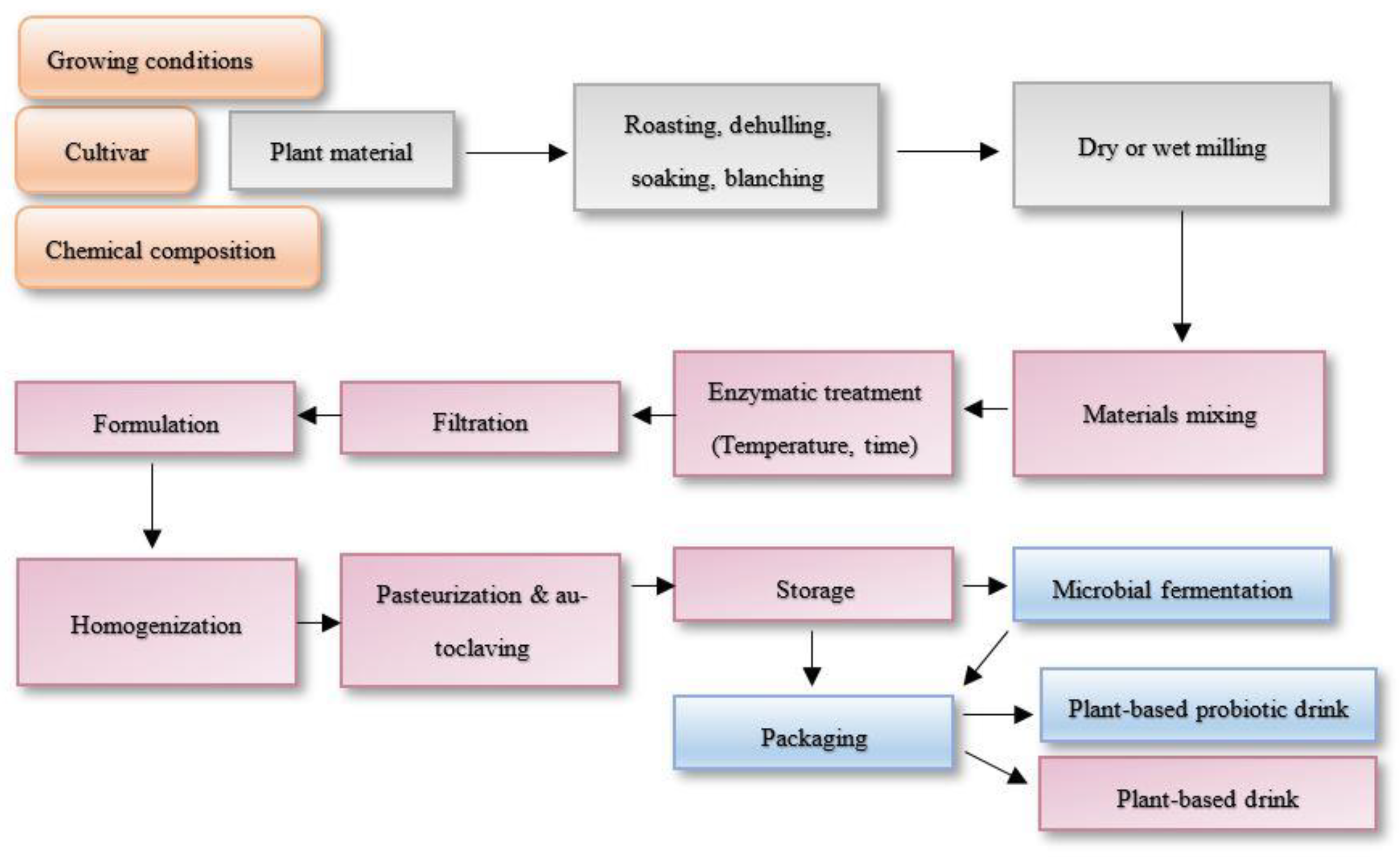
There is also no surprise that dairy milk substitute from grains production often requires enzymatic assistance for increasing extraction yields, proteins, and total solids content [22]. Annually, global plant-based dairy substitutes were marked to be grown by 10% and by 2019 had reached US 1.8 billion dollars [94]. Moreover, created derivatives and released sugars create sensory-acceptant organoleptic properties [16]. Amylolytic enzymes are required due to the amylose and amylolytic ratio of the starches, resulting in different rheological and textural properties. However, disrupting amylose and amylopectin molecules increases liquefying properties of grain beverages. Many starch-modifying enzymes including α-amylase (EC 3.2.1.1), β-amylase (EC 3.2.1.2), glucoamylase (EC 3.2.1.3), α-glucosidase (EC 3.2.1.20), pullulanase or amylopullulanase (EC 3.2.1.41), and cyclodextrin glycosyltransferase (EC 2.4.1.19) are used in industries that cleave the α-1,4- or α-1,6-glycosidic linkages, leading to the release of reducing sugars and oligosaccharides which are possibly prebiotic [32,95]. Interestingly, a quantitative prebiotic score of plant-based dairy substitutes can be identified, describing which prebiotics foster probiotic strains’ selective growth. Phenolic compounds, reduced sugars, and oligosaccharides are also great nutrients for bacterial growth, and the fermented food sector is rising due to their possible probiotic functions [66,96]. However, different plant materials are different sources for particular strains [76,97]. Enzymes are widely applicable and potent as precursors of nutrient production and initiate the growth of fermentation starter cultures: e.g., Lactobacillus strains L. reuteri L45, L. plantarum L47, and L. johnsonii L63 growth were initiated by cellulase- and pectinase-degraded rapeseed fibers [98]. Nissen et al. [99] identified prebiotic scores with different selectivity of hemp, soy, rice, and their mixtures, where hemp and soy drinks initiated the highest growth of L. plantarum 98b, and for growth B. bifidum B700795 mixture of hemp-rice and hemp-soy drinks, showed the potent scores [99,100]. Five main steps are incorporated in the flow chart for manufacturing enzymatically treated and by-the-step fermented plant-based drinks from plant materials: plant disruption, extraction, formulation, fermentation, and packaging [94]. However, it is also important not to forget the cultivar’s selectivity to get the highest value product [101]. Figure 3 represents the summarized flow chart of plant-based drinks production, where additional importance is given for plant material selection due to European Commission strategy incorporation ‘from farm to fork’, which is the heart of the European Green Deal priorities from 2019–2024. Continuing on, the flow chart represents the prebiotic plant-based drink production, which further on leads to probiotic drink production which can be mono- or multi-microbial [94]. Shori et al. [102] reviewed the plant-based dairy substitutes fermented with probiotic strains functionality, shelf-life expansions, and nutrition value enhancement [102].
Moreover, plant-based fermented products have properties against pathogenic bacteria, and pH is lower than regular plant-based drinks, affecting product stability [103]. However, another aspect of ingested probiotic food pathway and viability may be investigated. The texture and nutrient density and the complexity of the food matrix are responsible for releasing the nutrients delivering specific metabolites and strains to the intestines [104,105]. Moreover, the density and variety of live cultures may be the essential indicators for increasing the variety of gut microbiome [77].
4.3. Nanocrystals, Nanofibers, and Nanocellulose
The latest studies indicate that phenolic content increased in fermented products [96,106,107]. As a sidestream nanocellulosic material, it is usually produced by Komagataeibacater, Acetobacter, Gluconacetobacter strains which might be used as, e.g., wound healing biofilm [108]. Specific enzymes release, cleave, transport, and form derivatives from different plant origins by opening the ability to discover green synthesis applications for nanofibers, nanocrystals, and nanoparticles. Aqueous different plant extracts are the new scientific approach for synthesizing nanoparticles by changing environmentally disruptive chemical and physical methods. Enzyme-assistance by disrupting plant cell wall microfibrils and amorphous zones is visible through Transmission Electron Microscopy (TEM), Atomic Force Microscopy (AFM), or Scanning Electron Microscopy (SEM), which also implies an increase of extract yield [109,110]. Plant extracts contain high phenolic content and reducing sugars and reducing or stabilizing agents [26,111]. Additionally, as mentioned before, increasing drug resistance and nanofibers formation is getting attention for possible antibacterial and drug delivery properties and nanocellulose formation for biodegradability, non-toxicity, and potential physicochemical properties [112]. Yarbrough et at. [113] described cellulolytic enzymes performance of nanofibrils and nanocrystals formation by depolymerizing carbohydrates into smaller units [113]. Depending on the plant origin, charges and forces change using specific cellulolytic activity characterized enzymes [68].
Summarized application of enzymes-assisted extraction and their products is described in Figure 4.
5. Existing Rules on the Use of Enzymes and Their Products
At present, in the European Union, food enzymes usage is described under Regulation (EC) No 1332/2008 of the European Parliament and of the Council of 16 December 2008 on food enzymes. Enzymes are considered food additives because they are found in the final products only in inactivated form and in smaller amounts. The definition of food enzymes are described in Regulation (EC) No 1169/2011 [34]. Specifically, directive 2000/13/EC describes labeling requirements for food enzymes. However, some limitations occur while extracting by-products: first, the lack of specific regulations to assure the safety of valorized products. For human consumption, some substances of food by-products are categorized as food additives and described in EC Regulation No. 1881/2006. However, it is categorized as nutraceutical; these products are covered by food supplement Directive 200/46/EC. Moreover, the contamination of food by-products must be implemented, which is described in Council Regulation 315/93/EEC. Secondly, if by-products are enzymatically modified, EC regulation No 258/97 (1997) could apply to food and food ingredients with a new or intentionally modified primary molecule structure if not used before May 1997. This regulation ensures the safety assessment before entering the EU market. While entering the USA market, Food and Drug Administration (FDA) regulates food additives (Sec. 348), new dietary ingredients (Sec. 350b), pesticide chemical residues (Sec. 346a), and others through Federal Food, Drug, and Cosmetic Act (Chapter 9, Subchapter IV) [8]. Moreover, for worldwide fair food trade, international food standard CODEX Alimentarius, which is covered by the World Health Organization (WHO) and Food and Agriculture Organization of the United Nations (FAO), also describes food enzymes as food additives. Depending on how a product is made, the usage is characterized by enzyme name, the producent, and the highest dosage. In general, for the standard of food additives CODEX STAN 192-1995 for flours and starch products, two enzymes are described. In detail, for α-amylase and glucoamylase, whose producers are Bacillus subtilis (INS 1100(iii)) and Aspergillus oryzae var. (INS 1100(iii)), dosage is not identified. However, all food additives, including food enzymes such as xylanases, pectinases, and others, are collected in the list of CODEX Specification for Food Additives CXN 6–2019 [115]. However, for enzymes producers, the recommended purity specifications for food-grade enzymes are given by the Joint FAO/WHO Expert Committee on Food Additives (JECFA) and the Food Chemical Codex (FCC).
6. Conclusions
The use of enzymes in extracting biological raw material compounds is an up-and-coming area from small-scale, laboratory optimization studies to large-scale, industrial applications. It implies food processing, functional components, and medical devices development for high antioxidant, anti-inflammatory, and antimicrobial characteristics. However, success in this area requires interdisciplinary research from various life sciences disciplines. An important area of research is investigating the stability of enzymes and their interaction with other food and plant ingredients during processing and storage; repeatability is also questionable because the plant material differs from the origin, cultivars, and growing, harvesting, and storage conditions. Additionally, limitations occur in the form of worldwide regulations of enzymes usage and dosages due to the novel components that are produced during these processes. However, enzyme-assisted processes are reaching for more sustainable development of innovations in a broad spectrum of industries.
Author Contributions
Conceptualization, P.S.; software, P.S.; validation, P.S., J.V. and P.V.; formal analysis, P.S.; investigation, P.S.; resources, P.S.; data curation, P.S.; writing—original draft preparation, P.S.; writing—review and editing, P.S., J.V., P.V.; visualization, P.S.; supervision, J.V. and P.V.; project administration, J.V.; funding acquisition, P.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the article.
Acknowledgments
The authors wish to thank the Lithuanian Research Centre for Agriculture and Forestry for the support of this study. The work is partly attributed to the project “Innovative application of biologically active substances for the prevention of cardiovascular insufficiency and sarcopenia” (Nr. 01.2.1-LVPA-K-856-01-0065) under grant agreement with the Lithuanian Business Support Agency (LVPA).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Rose, N.; Reynolds, T.; Kolodinsky, J. P90 Consumer Use of Food Labels Increases as “Clean Label” Trend Continues. J. Nutr. Educ. Behav. 2020, 52, S58–S59. [Google Scholar] [CrossRef]
- Chawla, G.K.; Garg, K.; Kaur, K.; Chopra, V.; Suri, R. Pattern of drug resistance among patients of pulmonary tuberculosis. Indian J. Tuberc. 2021. [Google Scholar] [CrossRef]
- Grunert, K.G.; De Bauw, M.; Dean, M.; Lähteenmäki, L.; Maison, D.; Pennanen, K.; Sandell, M.A.; Stasiuk, K.; Stickel, L.; Tarrega, A.; et al. No lockdown in the kitchen: How the COVID-19 pandemic has affected food-related behaviours. Food Res. Int. 2021, 150, 110752. [Google Scholar] [CrossRef] [PubMed]
- Gligor, O.; Mocan, A.; Moldovan, C.; Locatelli, M.; Crișan, G.; Ferreira, I.C. Enzyme-assisted extractions of polyphenols—A comprehensive review. Trends Food Sci. Technol. 2019, 88, 302–315. [Google Scholar] [CrossRef]
- Noda-Garcia, L.; Liebermeister, W.; Tawfik, D.S. Metabolite–Enzyme Coevolution: From Single Enzymes to Metabolic Pathways and Networks. Annu. Rev. Biochem. 2018, 87, 187–216. [Google Scholar] [CrossRef] [PubMed]
- BlueWeave Consulting and Research Pvt Ltd. Global industrial enzymes market projected to reach worth $9.2 bn by 2027. Focus Catal. 2021, 2021, 2. [Google Scholar] [CrossRef]
- Mackėla, I.; Andriekus, T.; Venskutonis, P.R. Biorefining of buckwheat (Fagopyrum esculentum) hulls by using supercritical fluid, Soxhlet, pressurized liquid and enzyme-assisted extraction methods. J. Food Eng. 2017, 213, 38–46. [Google Scholar] [CrossRef]
- Socas-Rodríguez, B.; Álvarez-Rivera, G.; Valdés, A.; Ibáñez, E.; Cifuentes, A. Food by-products and food wastes: Are they safe enough for their valorization? Trends Food Sci. Technol. 2021, 114, 133–147. [Google Scholar] [CrossRef]
- Lao, Y.; Zhang, M.; Li, Z.; Bhandari, B. A novel combination of enzymatic hydrolysis and fermentation: Effects on the flavor and nutritional quality of fermented Cordyceps militaris beverage. LWT 2019, 120, 108934. [Google Scholar] [CrossRef]
- Cunningham, M.; Vinderola, G.; Charalampopoulos, D.; Lebeer, S.; Sanders, M.E.; Grimaldi, R. Applying probiotics and prebiotics in new delivery formats—Is the clinical evidence transferable? Trends Food Sci. Technol. 2021, 112, 495–506. [Google Scholar] [CrossRef]
- Włodarczyk, M.; Śliżewska, K. Obesity as the 21st Century’s major disease: The role of probiotics and prebiotics in prevention and treatment. Food Biosci. 2021, 42, 101115. [Google Scholar] [CrossRef]
- Alou, M.T.; Lagier, J.-C.; Raoult, D. Diet influence on the gut microbiota and dysbiosis related to nutritional disorders. Hum. Microbiome J. 2016, 1, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Janssen, A.W.F.; Kersten, S. The role of the gut microbiota in metabolic health. FASEB J. 2015, 29, 3111–3123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado, G.T.C.; Tamashiro, W.M.D.S.C. Role of prebiotics in regulation of microbiota and prevention of obesity. Food Res. Int. 2018, 113, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Daïen, C.I.; Tan, J.; Audo, R.; Mielle, J.; Quek, L.-E.; Krycer, J.R.; Angelatos, A.S.; Duraes, M.; Pinget, G.V.; Ni, D.; et al. Gut-derived acetate promotes B10 cells with antiinflammatory effects. JCI Insight 2021, 6, 144156. [Google Scholar] [CrossRef]
- Mäkinen, O.E.; Wanhalinna, V.; Zannini, E.; Arendt, E.K. Foods for Special Dietary Needs: Non-dairy Plant-based Milk Substitutes and Fermented Dairy-type Products. Crit. Rev. Food Sci. Nutr. 2016, 56, 339–349. [Google Scholar] [CrossRef]
- Penha, C.B.; Santos, V.D.P.; Speranza, P.; Kurozawa, L.E. Plant-based beverages: Ecofriendly technologies in the production process. Innov. Food Sci. Emerg. Technol. 2021, 72, 102760. [Google Scholar] [CrossRef]
- Cantu-Jungles, T.M.; Zhang, X.; Kazem, A.E.; Iacomini, M.; Hamaker, B.R.; Cordeiro, L.M. Microwave treatment enhances human gut microbiota fermentability of isolated insoluble dietary fibers. Food Res. Int. 2021, 143, 110293. [Google Scholar] [CrossRef]
- Zhang, Y.-G.; Kan, H.; Chen, S.-X.; Thakur, K.; Wang, S.; Zhang, J.-G.; Shang, Y.-F.; Wei, Z.-J. Comparison of phenolic compounds extracted from Diaphragma juglandis fructus, walnut pellicle, and flowers of Juglans regia using methanol, ultrasonic wave, and enzyme assisted-extraction. Food Chem. 2020, 321, 126672. [Google Scholar] [CrossRef]
- Puri, M.; Sharma, D.; Barrow, C.J. Enzyme-assisted extraction of bioactives from plants. Trends Biotechnol. 2012, 30, 37–44. [Google Scholar] [CrossRef]
- Islam, S.M.; Ju, L.-K. Enzymatic soybean flour processing: Modeling for insights into optimal carbohydrases composition and carbohydrate monomerization from complex biomass. Catal. Commun. 2021, 149, 106244. [Google Scholar] [CrossRef]
- Jeske, S.; Zannini, E.; Cronin, M.F.; Arendt, E.K. Impact of protease and amylase treatment on proteins and the product quality of a quinoa-based milk substitute. Food Funct. 2018, 9, 3500–3508. [Google Scholar] [CrossRef] [PubMed]
- Dhasarathan, P.; AlSalhi, M.S.; Devanesan, S.; Subbiah, J.; Ranjitsingh, A.; Binsalah, M.; Alfuraydi, A.A. Drug resistance in Candida albicans isolates and related changes in the structural domain of Mdr1 protein. J. Infect. Public Health 2021, 14, 1848–1853. [Google Scholar] [CrossRef] [PubMed]
- Balciunaitiene, A.; Viskelis, P.; Viskelis, J.; Streimikyte, P.; Liaudanskas, M.; Bartkiene, E.; Zavistanaviciute, P.; Zokaityte, E.; Starkute, V.; Ruzauskas, M.; et al. Green Synthesis of Silver Nanoparticles Using Extract of Artemisia absinthium L., Humulus lupulus L. and Thymus vulgaris L., Physico-Chemical Characterization, Antimicrobial and Antioxidant Activity. Process 2021, 9, 1304. [Google Scholar] [CrossRef]
- Chandhirasekar, K.; Thendralmanikandan, A.; Thangavelu, P.; Nguyen, B.-S.; Nguyen, T.-A.; Sivashanmugan, K.; Nareshkumar, A.; Nguyen, V.-H. Plant-extract-assisted green synthesis and its larvicidal activities of silver nanoparticles using leaf extract of Citrus medica, Tagetes lemmonii, and Tarenna asiatica. Mater. Lett. 2021, 287, 129265. [Google Scholar] [CrossRef]
- Jalab, J.; Abdelwahed, W.; Kitaz, A.; Al-Kayali, R. Green synthesis of silver nanoparticles using aqueous extract of Acacia cyanophylla and its antibacterial activity. Heliyon 2021, 7, 08033. [Google Scholar] [CrossRef]
- Naidu, D.; Hlangothi, S.P.; John, M.J. Bio-based products from xylan: A review. Carbohydr. Polym. 2018, 179, 28–41. [Google Scholar] [CrossRef]
- De Bhowmick, G.; Sarmah, A.K.; Sen, R. Lignocellulosic biorefinery as a model for sustainable development of biofuels and value added products. Bioresour. Technol. 2018, 247, 1144–1154. [Google Scholar] [CrossRef]
- Acosta-Estrada, B.A.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Bound phenolics in foods, a review. Food Chem. 2014, 152, 46–55. [Google Scholar] [CrossRef]
- Carpita, N.C.; McCann, M.C. Redesigning plant cell walls for the biomass-based bioeconomy. J. Biol. Chem. 2020, 295, 15144–15157. [Google Scholar] [CrossRef]
- Raveendran, S.; Parameswaran, B.; Ummalyma, S.B.; Abraham, A.; Mathew, A.K.; Madhavan, A.; Rebello, S.; Pandey, A. Applications of Microbial Enzymes in Food Industry. Food Technol. Biotechnol. 2018, 56, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Miao, M.; Jiang, B.; Jin, Z.; BeMiller, J.N. Microbial Starch-Converting Enzymes: Recent Insights and Perspectives. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1238–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puchart, V.; Šuchová, K.; Biely, P. Xylanases of glycoside hydrolase family 30—An overview. Biotechnol. Adv. 2021, 47, 107704. [Google Scholar] [CrossRef] [PubMed]
- Danalache, F.; Mata, P.; Alves, V.D.; Moldão-Martins, M. Chapter 10—Enzyme-Assisted Extraction of Fruit Juices. In Fruit Juices; Rajauria, G., Brijesh, K., Eds.; Academic Press: San Diego, CA, USA, 2018; pp. 183–200. ISBN 978-0-12-802230-6. [Google Scholar]
- Kawamori, M.; Morikawa, Y.; Takasawa, S. Induction and production of cellulases by l-sorbose in Trichoderma reesei. Appl. Microbiol. Biotechnol. 1986, 24, 449–453. [Google Scholar] [CrossRef]
- Juturu, V.; Wu, J.C. Microbial cellulases: Engineering, production and applications. Renew. Sustain. Energy Rev. 2014, 33, 188–203. [Google Scholar] [CrossRef]
- Pedersen, N.R.; Ravn, J.L.; Pettersson, D. A multienzyme NSP product solubilises and degrades NSP structures in canola and mediates protein solubilisation and degradation in vitro. Anim. Feed Sci. Technol. 2017, 234, 244–252. [Google Scholar] [CrossRef]
- Ire, F.S.; Chima, I.J.; Ezebuiro, V. Enhanced xylanase production from UV-mutated Aspergillus niger grown on corn cob and sawdust. Biocatal. Agric. Biotechnol. 2021, 31, 101869. [Google Scholar] [CrossRef]
- Tan, H.; Nie, S. Deciphering diet-gut microbiota-host interplay: Investigations of pectin. Trends Food Sci. Technol. 2020, 106, 171–181. [Google Scholar] [CrossRef]
- Muley, A.B.; Thorat, A.S.; Singhal, R.S.; Babu, K.H. A tri-enzyme co-immobilized magnetic complex: Process details, kinetics, thermodynamics and applications. Int. J. Biol. Macromol. 2018, 118, 1781–1795. [Google Scholar] [CrossRef]
- Ladole, M.R.; Pokale, P.B.; Varude, V.R.; Belokar, P.G.; Pandit, A.B. One pot clarification and debittering of grapefruit juice using co-immobilized enzymes@chitosanMNPs. Int. J. Biol. Macromol. 2021, 167, 1297–1307. [Google Scholar] [CrossRef]
- Biesalski, H.-K.; Dragsted, L.O.; Elmadfa, I.; Grossklaus, R.; Müller, M.; Schrenk, D.; Walter, P.; Weber, P. Bioactive compounds: Definition and assessment of activity. Nutrition 2009, 25, 1202–1205. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.A.; Iqbal, M.; Rasheed, R.; Hussain, I.; Riaz, M.; Arif, M.S. Chapter 8—Environmental Stress and Secondary Metabolites in Plants: An Overview. In Plant Metabolites and Regulation under Environmental Stress; Ahmad, P., Ahanger, M.A., Singh, V.P., Tripathi, D.K., Alam, P., Nasser, M., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 153–167. ISBN 978-0-12-812689-9. [Google Scholar]
- Shamsi, A.; Shahwan, M.; Khan, M.S.; Husain, F.M.; Alhumaydhi, F.A.; Aljohani, A.S.M.; Rehman, T.; Hassan, I.; Islam, A. Elucidating the Interaction of Human Ferritin with Quercetin and Naringenin: Implication of Natural Products in Neurodegenerative Diseases: Molecular Docking and Dynamics Simulation Insight. ACS Omega 2021, 6, 7922–7930. [Google Scholar] [CrossRef] [PubMed]
- Paz, A.; Outeiriño, D.; Guerra, N.P.; Domínguez, J.M. Enzymatic hydrolysis of brewer’s spent grain to obtain fermentable sugars. Bioresour. Technol. 2019, 275, 402–409. [Google Scholar] [CrossRef]
- Uddin, S.; Kabir, T.; Niaz, K.; Jeandet, P.; Clément, C.; Mathew, B.; Rauf, A.; Rengasamy, K.R.; Sobarzo-Sánchez, E.; Ashraf, G.M.; et al. Molecular Insight into the Therapeutic Promise of Flavonoids against Alzheimer’s Disease. Molecules 2020, 25, 1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, J.P.E. Flavonoids: Modulators of brain function? Br. J. Nutr. 2008, 99, ES60–ES77. [Google Scholar] [CrossRef] [Green Version]
- Gulpinar, A.R.; Orhan, I.E.; Kan, A.; Senol, F.S.; Celik, S.A.; Kartal, M. Estimation of in vitro neuroprotective properties and quantification of rutin and fatty acids in buckwheat (Fagopyrum esculentum Moench) cultivated in Turkey. Food Res. Int. 2012, 46, 536–543. [Google Scholar] [CrossRef]
- Urbanavičiūtė, I.; Liaudanskas, M.; Bobinas, Č.; Šarkinas, A.; Rezgienė, A.; Viskelis, P. Japanese Quince (Chaenomeles japonica) as a Potential Source of Phenols: Optimization of the Extraction Parameters and Assessment of Antiradical and Antimicrobial Activities. Foods 2020, 9, 1132. [Google Scholar] [CrossRef]
- Raudonė, L.; Liaudanskas, M.; Vilkickytė, G.; Kviklys, D.; Žvikas, V.; Viškelis, J.; Viškelis, P. Phenolic Profiles, Antioxidant Activity and Phenotypic Characterization of Lonicera caerulea L. Berries, Cultivated in Lithuania. Antioxidants 2021, 10, 115. [Google Scholar] [CrossRef]
- Yang, B.; Jiang, Y.; Shi, J.; Chen, F.; Ashraf, M. Extraction and pharmacological properties of bioactive compounds from longan (Dimocarpus longan Lour.) fruit—A review. Food Res. Int. 2011, 44, 1837–1842. [Google Scholar] [CrossRef]
- Teo, C.C.; Tan, S.N.; Yong, J.W.H.; Hew, C.S.; Ong, E.S. Pressurized hot water extraction (PHWE). J. Chromatogr. A 2010, 1217, 2484–2494. [Google Scholar] [CrossRef]
- Gardossi, L.; Poulsen, P.B.; Ballesteros, A.; Hult, K.; Švedas, V.K.; Vasić-Rački, Đ.; Carrea, G.; Magnusson, A.; Schmid, A.; Wohlgemuth, R.; et al. Guidelines for reporting of biocatalytic reactions. Trends Biotechnol. 2010, 28, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhou, X.; Zhang, J. Optimization of enzyme assisted extraction of polysaccharides from Astragalus membranaceus. Carbohydr. Polym. 2014, 111, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Zuorro, A.; Lavecchia, R.; González-Delgado, Á.D.; García-Martinez, J.B.; L’Abbate, P. Optimization of Enzyme-Assisted Extraction of Flavonoids from Corn Husks. Process 2019, 7, 804. [Google Scholar] [CrossRef] [Green Version]
- Drula, E.; Garron, M.-L.; Dogan, S.; Lombard, V.; Henrissat, B.; Terrapon, N. The carbohydrate-active enzyme database: Functions and literature. Nucleic Acids Res. 2021, 50, D571–D577. [Google Scholar] [CrossRef] [PubMed]
- Bulmer, G.S.; de Andrade, P.; Field, R.A.; van Munster, J.M. Recent advances in enzymatic synthesis of β-glucan and cellulose. Carbohydr. Res. 2021, 508, 108411. [Google Scholar] [CrossRef]
- Woo, S.-Y.; Yang, J.Y.; Lee, H.; Ahn, H.J.; Lee, Y.B.; Do, S.H.; Kim, J.Y.; Seo, W.D. Changes in metabolites with harvest times of seedlings of various Korean oat (Avena sativa L.) cultivars and their neuraminidase inhibitory effects. Food Chem. 2021, 373, 131429. [Google Scholar] [CrossRef]
- Cole, M.R.; Eggleston, G.; Gaines, D.K.; Heckemeyer, M. Development of an enzyme cocktail to bioconvert untapped starch in sweet sorghum processing by-products: Part I. Ind. Crop. Prod. 2019, 133, 142–150. [Google Scholar] [CrossRef]
- González, M.J.A.; Carrera, C.; Barbero, G.F.; Palma, M. A comparison study between ultrasound–assisted and enzyme–assisted extraction of anthocyanins from blackcurrant (Ribes nigrum L.). Food Chem. X 2021, 13, 100192. [Google Scholar] [CrossRef]
- Kongthitilerd, P.; Thilavech, T.; Marnpae, M.; Rong, W.; Yao, S.; Adisakwattana, S.; Cheng, H.; Suantawee, T. Cyanidin-3-rutinoside stimulated insulin secretion through activation of L-type voltage-dependent Ca2+ channels and the PLC-IP3 pathway in pancreatic β-cells. Biomed. Pharmacother. 2021, 146, 112494. [Google Scholar] [CrossRef]
- Song, L.-W.; Qi, J.-R.; Liao, J.-S.; Yang, X.-Q. Enzymatic and enzyme-physical modification of citrus fiber by xylanase and planetary ball milling treatment. Food Hydrocoll. 2021, 121, 107015. [Google Scholar] [CrossRef]
- Belmiro, R.H.; Oliveira, L.D.C.; Geraldi, M.V.; Junior, M.R.M.; Cristianini, M. Modification of coffee coproducts by-products by dynamic high pressure, acetylation and hydrolysis by cellulase: A potential functional and sustainable food ingredient. Innov. Food Sci. Emerg. Technol. 2021, 68, 102608. [Google Scholar] [CrossRef]
- Phirom-On, K.; Apiraksakorn, J. Development of cellulose-based prebiotic fiber from banana peel by enzymatic hydrolysis. Food Biosci. 2021, 41, 101083. [Google Scholar] [CrossRef]
- Ninga, K.A.; Desobgo, Z.S.C.; De, S.; Nso, E.J. Pectinase hydrolysis of guava pulp: Effect on the physicochemical characteristics of its juice. Heliyon 2021, 7, 08141. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Liu, Y.; Zeng, J.; Tian, X.; Bei, Q.; Wu, Z. Enhancing three phenolic fractions of oats (Avena sativa L.) and their antioxidant activities by solid-state fermentation with Monascus anka and Bacillus subtilis. J. Cereal Sci. 2020, 93, 102940. [Google Scholar] [CrossRef]
- Pinelo, M.; Arnous, A.; Meyer, A.S. Upgrading of grape skins: Significance of plant cell-wall structural components and extraction techniques for phenol release. Trends Food Sci. Technol. 2006, 17, 579–590. [Google Scholar] [CrossRef]
- Zhou, Z.; Ju, X.; Chen, J.; Wang, R.; Zhong, Y.; Li, L. Charge-oriented strategies of tunable substrate affinity based on cellulase and biomass for improving in situ saccharification: A review. Bioresour. Technol. 2021, 319, 124159. [Google Scholar] [CrossRef]
- HozoVá, B.; Kuniak, L.; Moravcikova, P.; Gajdosova, A. Determination of Water-Insoluble Beta-D-Glucan in the Whole-Grain Cereals and Pseudocereals. Czech J. Food Sci. 2007, 25, 316. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M. Cereal beta-glucans: An underutilized health endorsing food ingredient. Crit. Rev. Food Sci. Nutr. 2020, 1–20. [Google Scholar] [CrossRef]
- Nishantha, M.D.L.C.; Zhao, X.; Jeewani, D.C.; Bian, J.; Nie, X.; Weining, S. Direct comparison of β-glucan content in wild and cultivated barley. Int. J. Food Prop. 2018, 21, 2218–2228. [Google Scholar] [CrossRef] [Green Version]
- Du, B.; Meenu, M.; Liu, H.; Xu, B. A Concise Review on the Molecular Structure and Function Relationship of β-Glucan. Int. J. Mol. Sci. 2019, 20, 4032. [Google Scholar] [CrossRef] [Green Version]
- Shah, A.; Masoodi, F.; Gani, A.; Ashwar, B. Dual enzyme modified oat starch: Structural characterisation, rheological properties, and digestibility in simulated GI tract. Int. J. Biol. Macromol. 2018, 106, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Herburger, K.; Kirkensgaard, J.J.K.; Khakimov, B.; Hansen, A.R.; Blennow, A. Sequential maltogenic α-amylase and branching enzyme treatment to modify granular corn starch. Food Hydrocoll. 2021, 120, 106904. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, P.; Zhao, J. Ferulic acid mediates prebiotic responses of cereal-derived arabinoxylans on host health. Anim. Nutr. 2021. [Google Scholar] [CrossRef]
- Pereira, G.V.; Abdel-Hamid, A.M.; Dutta, S.; D’Alessandro-Gabazza, C.N.; Wefers, D.; Farris, J.A.; Bajaj, S.; Wawrzak, Z.; Atomi, H.; Mackie, R.I.; et al. Degradation of complex arabinoxylans by human colonic Bacteroidetes. Nat. Commun. 2021, 12, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Wastyk, H.C.; Fragiadakis, G.K.; Perelman, D.; Dahan, D.; Merrill, B.D.; Yu, F.B.; Topf, M.; Gonzalez, C.G.; Van Treuren, W.; Han, S.; et al. Gut-microbiota-targeted diets modulate human immune status. Cell 2021, 184, 4137–4153.e14. [Google Scholar] [CrossRef]
- Vangay, P.; Johnson, A.; Ward, T.L.; Al-Ghalith, G.A.; Shields-Cutler, R.R.; Hillmann, B.M.; Lucas, S.K.; Beura, L.K.; Thompson, E.A.; Till, L.M.; et al. US Immigration Westernizes the Human Gut Microbiome. Cell 2018, 175, 962–972.e10. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Abdo, A.A.A.; Kaddour, B.; Wu, Q.; Xin, L.; Li, X.; Fan, G.; Teng, C. Xylan-oligosaccharides ameliorate high fat diet induced obesity and glucose intolerance and modulate plasma lipid profile and gut microbiota in mice. J. Funct. Foods 2020, 64, 103622. [Google Scholar] [CrossRef]
- Yang, F.; Wei, J.-D.; Lu, Y.-F.; Sun, Y.-L.; Wang, Q.; Zhang, R.-L. Galacto-oligosaccharides modulate gut microbiota dysbiosis and intestinal permeability in rats with alcohol withdrawal syndrome. J. Funct. Foods 2019, 60, 103423. [Google Scholar] [CrossRef]
- Li, E.; Long, X.; Liao, S.; Pang, D.; Li, Q.; Zou, Y. Effect of mulberry galacto-oligosaccharide isolated from mulberry on glucose metabolism and gut microbiota in a type 2 diabetic mice. J. Funct. Foods 2021, 87, 104836. [Google Scholar] [CrossRef]
- Wilkins, M.R.; Widmer, W.W.; Grohmann, K.; Cameron, R. Hydrolysis of grapefruit peel waste with cellulase and pectinase enzymes. Bioresour. Technol. 2007, 98, 1596–1601. [Google Scholar] [CrossRef]
- Qi, W.J.; Sheng, W.S.; Peng, C.; Xiaodong, M.; Yao, T.Z. Investigating into anti-cancer potential of lycopene: Molecular targets. Biomed. Pharmacother. 2021, 138, 111546. [Google Scholar] [CrossRef] [PubMed]
- Catalkaya, G.; Kahveci, D. Optimization of enzyme assisted extraction of lycopene from industrial tomato waste. Sep. Purif. Technol. 2019, 219, 55–63. [Google Scholar] [CrossRef]
- Urbonaviciene, D.; Viskelis, P. The cis-lycopene isomers composition in supercritical CO2 extracted tomato by-products. LWT 2017, 85, 517–523. [Google Scholar] [CrossRef]
- Kundu, D.; Karmakar, S.; Banerjee, R. Simultaneous debittering and clarification of enzyme mediated mixed citrus juice production. Appl. Food Res. 2021, 2, 100031. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, Y.; Tian, H. Effects of carboxymethylation, acidic treatment, hydroxypropylation and heating combined with enzymatic hydrolysis on structural and physicochemical properties of palm kernel expeller dietary fiber. LWT 2020, 133, 109909. [Google Scholar] [CrossRef]
- Marić, M.; Grassino, A.N.; Zhu, Z.; Barba, F.J.; Brnčić, M.; Brnčić, S.R. An overview of the traditional and innovative approaches for pectin extraction from plant food wastes and by-products: Ultrasound-, microwaves-, and enzyme-assisted extraction. Trends Food Sci. Technol. 2018, 76, 28–37. [Google Scholar] [CrossRef]
- Yuliarti, O.; Goh, K.K.; Matia-Merino, L.; Mawson, J.; Brennan, C. Extraction and characterisation of pomace pectin from gold kiwifruit (Actinidia chinensis). Food Chem. 2015, 187, 290–296. [Google Scholar] [CrossRef]
- Rodríguez De Luna, S.L.; Ramírez-Garza, R.E.; Serna-Saldívar, S.O. Environmentally friendly methods for flavonoid extraction from plant material: Impact of their operating conditions on yield and antioxidant properties. Sci. World J. 2020, 2020, 6792069. [Google Scholar] [CrossRef]
- Yazdi, A.P.G.; Barzegar, M.; Sahari, M.A.; Gavlighi, H.A. Optimization of the enzyme-assisted aqueous extraction of phenolic compounds from pistachio green hull. Food Sci. Nutr. 2018, 7, 356–366. [Google Scholar] [CrossRef]
- Costa, R.D.S.; de Almeida, S.S.; Cavalcanti, E.D.C.; Freire, D.M.G.; Moura-Nunes, N.; Monteiro, M.; Perrone, D. Enzymes produced by solid state fermentation of agro-industrial by-products release ferulic acid in bioprocessed whole-wheat breads. Food Res. Int. 2021, 140, 109843. [Google Scholar] [CrossRef]
- Yin, Z.; Wu, W.; Sun, C.; Lei, Z.; Chen, H.; Liu, H.; Chen, W.; Ma, J.; Min, T.; Zhang, M.; et al. Comparison of releasing bound phenolic acids from wheat bran by fermentation of three Aspergillus species. Int. J. Food Sci. Technol. 2018, 53, 1120–1130. [Google Scholar] [CrossRef]
- Tangyu, M.; Muller, J.; Bolten, C.J.; Wittmann, C. Fermentation of plant-based milk alternatives for improved flavour and nutritional value. Appl. Microbiol. Biotechnol. 2019, 103, 9263–9275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rempel, A.; Machado, T.; Treichel, H.; Colla, E.; Margarites, A.C.; Colla, L. Saccharification of Spirulina platensis biomass using free and immobilized amylolytic enzymes. Bioresour. Technol. 2018, 263, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Silva, K.A.; Uekane, T.M.; de Miranda, J.F.; Ruiz, L.F.; da Motta, J.C.B.; Silva, C.B.; Pitangui, N.D.S.; Gonzalez, A.G.M.; Fernandes, F.F.; Lima, A.R. Kombucha beverage from non-conventional edible plant infusion and green tea: Characterization, toxicity, antioxidant activities and antimicrobial properties. Biocatal. Agric. Biotechnol. 2021, 34, 102032. [Google Scholar] [CrossRef]
- Bai, J.; Li, Y.; Zhang, W.; Fan, M.; Qian, H.; Zhang, H.; Qi, X.; Wang, L. Source of gut microbiota determines oat β-glucan degradation and short chain fatty acid-producing pathway. Food Biosci. 2021, 41, 101010. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, L.; Zhang, Z.; Ding, L.; Hang, S. Combination of fiber-degrading enzymatic hydrolysis and lactobacilli fermentation enhances utilization of fiber and protein in rapeseed meal as revealed in simulated pig digestion and fermentation in vitro. Anim. Feed Sci. Technol. 2021, 278, 115001. [Google Scholar] [CrossRef]
- Nissen, L.; di Carlo, E.; Gianotti, A. Prebiotic potential of hemp blended drinks fermented by probiotics. Food Res. Int. 2020, 131, 109029. [Google Scholar] [CrossRef]
- Huebner, J.; Wehling, R.; Parkhurst, A.; Hutkins, R. Effect of processing conditions on the prebiotic activity of commercial prebiotics. Int. Dairy J. 2008, 18, 287–293. [Google Scholar] [CrossRef]
- Du, J.; Pan, R.; Obadi, M.; Li, H.; Shao, F.; Sun, J.; Wang, Y.; Qi, Y.; Xu, B. In vitro starch digestibility of buckwheat cultivars in comparison to wheat: The key role of starch molecular structure. Food Chem. 2021, 368, 130806. [Google Scholar] [CrossRef]
- Shori, A.B.; Aljohani, G.S.; Al-Zahrani, A.J.; Al-Sulbi, O.S.; Baba, A.S. Viability of probiotics and antioxidant activity of cashew milk-based yogurt fermented with selected strains of probiotic Lactobacillus spp. LWT 2021, 153, 112482. [Google Scholar] [CrossRef]
- Panghal, A.; Janghu, S.; Virkar, K.; Gat, Y.; Kumar, V.; Chhikara, N. Potential non-dairy probiotic products—A healthy approach. Food Biosci. 2018, 21, 80–89. [Google Scholar] [CrossRef]
- Guimarães, J.T.; Balthazar, C.F.; Silva, R.; Rocha, R.S.; Graça, J.S.; A Esmerino, E.; Silva, M.C.; Sant’Ana, A.S.; Duarte, M.C.K.H.; Freitas, M.Q.; et al. Impact of probiotics and prebiotics on food texture. Curr. Opin. Food Sci. 2020, 33, 38–44. [Google Scholar] [CrossRef]
- Shori, A.B.; AL Zahrani, A.J. Non-dairy plant-based milk products as alternatives to conventional dairy products for delivering probiotics. Food Sci. Technol. 2021. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, W.; Wei, Z.; Yin, B.; Man, C.; Jiang, Y. Enhancement of functional characteristics of blueberry juice fermented by Lactobacillus plantarum. LWT 2021, 139, 110590. [Google Scholar] [CrossRef]
- Gao, B.; Wang, J.; Wang, Y.; Xu, Z.; Li, B.; Meng, X.; Sun, X.; Zhu, J. Influence of fermentation by lactic acid bacteria and in vitro digestion on the biotransformations of blueberry juice phenolics. Food Control 2021, 133, 108603. [Google Scholar] [CrossRef]
- Ahmed, J.; Gultekinoglu, M.; Edirisinghe, M. Bacterial cellulose micro-nano fibres for wound healing applications. Biotechnol. Adv. 2020, 41, 107549. [Google Scholar] [CrossRef]
- Rossi, B.R.; Pellegrini, V.O.; Cortez, A.A.; Chiromito, E.M.; Carvalho, A.J.; Pinto, L.O.; Rezende, C.A.; Mastelaro, V.R.; Polikarpov, I. Cellulose nanofibers production using a set of recombinant enzymes. Carbohydr. Polym. 2021, 256, 117510. [Google Scholar] [CrossRef]
- Meftahi, A.; Samyn, P.; Geravand, S.A.; Khajavi, R.; Alibkhshi, S.; Bechelany, M.; Barhoum, A. Nanocelluloses as skin biocompatible materials for skincare, cosmetics, and healthcare: Formulations, regulations, and emerging applications. Carbohydr. Polym. 2021, 278, 118956. [Google Scholar] [CrossRef]
- Ceylan, R.; Demirbas, A.; Ocsoy, I.; Aktumsek, A. Green synthesis of silver nanoparticles using aqueous extracts of three Sideritis species from Turkey and evaluations bioactivity potentials. Sustain. Chem. Pharm. 2021, 21, 100426. [Google Scholar] [CrossRef]
- Liu, S.; Qamar, S.A.; Qamar, M.; Basharat, K.; Bilal, M. Engineered nanocellulose-based hydrogels for smart drug delivery applications. Int. J. Biol. Macromol. 2021, 181, 275–290. [Google Scholar] [CrossRef]
- Yarbrough, J.M.; Zhang, R.; Mittal, A.; Wall, T.V.; Bomble, Y.J.; Decker, S.R.; Himmel, M.E.; Ciesielski, P.N. Multifunctional Cellulolytic Enzymes Outperform Processive Fungal Cellulases for Coproduction of Nanocellulose and Biofuels. ACS Nano 2017, 11, 3101–3109. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Chae, M.; Beyene, D.; Danumah, C.; Tosto, F.; Bressler, D.C. Co-Production of Cellulose Nanocrystals and Fermentable Sugars Assisted by Endoglucanase Treatment of Wood Pulp. Materials 2018, 11, 1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ladics, G.S.; Fan, L.; Sewalt, V.J.; Spök, A. Chapter Eight—Safety Assessment and Regulation of Food Enzymes. In Enzymes; Kermasha, S., Michael, N.A., Eds.; Academic Press: San Diego, CA, USA, 2021; pp. 203–258. ISBN 978-0-12-800217-9. [Google Scholar]
Figure 1.
Plant cell wall graphical scheme, describing cross-linked phenolic compounds, peptides, and polysaccharides network adapted from Acosta et al., 2014 and Carpita et al., 2020 [29,30]. A—cellulose from cellulose microfibrils, B—hemicelluloses consisting of xyloglucans, glucuronoarabinoxylan, (1–3) (1–4) β glucans and glucomannan. C—structural proteins, D—pectin consisting of homogalacturonan, xylogalacturonan, and rhamnogalacturonans I and II; E—phenolic compounds, F—lignin; G—xylan and mannan coating of cellulose microfibrils.
Figure 1.
Plant cell wall graphical scheme, describing cross-linked phenolic compounds, peptides, and polysaccharides network adapted from Acosta et al., 2014 and Carpita et al., 2020 [29,30]. A—cellulose from cellulose microfibrils, B—hemicelluloses consisting of xyloglucans, glucuronoarabinoxylan, (1–3) (1–4) β glucans and glucomannan. C—structural proteins, D—pectin consisting of homogalacturonan, xylogalacturonan, and rhamnogalacturonans I and II; E—phenolic compounds, F—lignin; G—xylan and mannan coating of cellulose microfibrils.

Figure 2.
Summary graphic of polysaccharides and enzyme combination outcomes connected to solutions for uprising challenges.
Figure 2.
Summary graphic of polysaccharides and enzyme combination outcomes connected to solutions for uprising challenges.

Figure 3.
Chart flow of plant-based prebiotic and probiotic drinks production.

Figure 4.
Enzyme-assisted extract or possible product applications, where A presents alternative dairy substitutes development [22], B—fermented beverages production [96], C—nanocrystals formation from enzymatically treated wood pulp [114]. A and B photographs were taken by the authors at the Lithuanian Research Centre for Agriculture and Forestry.
Figure 4.
Enzyme-assisted extract or possible product applications, where A presents alternative dairy substitutes development [22], B—fermented beverages production [96], C—nanocrystals formation from enzymatically treated wood pulp [114]. A and B photographs were taken by the authors at the Lithuanian Research Centre for Agriculture and Forestry.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Structure of commonly found carbohydrates in plant material and enzymes usage of their cleavage [30,31,32,33].
| Non-Starch Polysaccharides | |||
|---|---|---|---|
| Hemicellulose | Cellulose | Pectin | |
| Consist of | xyloglucans | cellulose nanofibrils: | homogalacturonan |
| glucuronoarabinoxylan | (a) xylan | rhamnogalacturonan I and II | |
| β–glucan | (b) mannan | xylogalacturonan | |
| glucomannan | |||
| Enzymes used in processes | Xylanases: | Cellulases: | Pectinases: |
| exoxylanases | endo–(1,4)–β–d–glucanase (EC 3.2.1.4) | polygalacturonases | |
| β–xylosidases, | exo–(1,4)–β–d–glucanase (EC 3.2.1.91) | pectin esterases | |
| xylan–1,4–β-xylosidase | β–glucosidases (EC 3.3.1.21) | pectate lyase | |
| endoxylanases | β–glucosidases (EC 3.3.1.21) | ||
| Starches | |||
| Consist of | amylose | ||
| amylopectin | |||
| Enzymes used in processes | α–Amylases (EC 3.2.1.1) | ||
| β–amylase (EC 3.2.1.2) | |||
| glucoamylase (EC 3.2.1.3) | |||
| α-glucosidase (EC 3.2.1.20) | |||
| pullulanase or amylopullulanase (EC 3.2.1.41) | |||
| cyclodextrin glycosyltransferase (EC 2.4.1.19) | |||
Table 2.
Examples of various substrates technological parameters for enzyme-assisted extraction.
| Enzymes | Producent | Substrate | Enzymes Quantity | Liquid to Substrate Ratio | pH | Temp. °C | Time | Ref. |
|---|---|---|---|---|---|---|---|---|
| Xylanase cocktail | A. niger | Citrus fiber | 0.45% | 1:20 | 4.5−6.5 | 50 | 120 min | Song et al. [62] |
| Cellulase | A. niger | Coffee by-products | 5−15 U | 1:25 | 5.0−6.0 | 50 | 30−20 min | Belmiro et al. [63] |
| Cellulase from Celluclast 1.5 L | T. reesei | Banana peel | 5 FPU/ml | 1:20 | 6.0−7.0 | 50 | 120 h | Phirom-on et al. [64] |
| Pectinase | A. niger | Guava pulp | 0.10% | 2:5 | 2.97−3.97 | 45 | 3–90 min | Ninga et al. [65] |
| Pectinase | A. niger | Blackcurrant | 108 U/g | 0.1:15 and 0.2:15 | 5−6 | 60 | 10−90 min | Gonzalez et al. [60] |
| Heat stable alpha-amylase | Bacillus sp. | Oat flours | 0.01% | 1:5 | 5.0−9.0 | 100 | 15−75 min | Chen et al. [66] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Streimikyte, P.; Viskelis, P.; Viskelis, J. Enzymes-Assisted Extraction of Plants for Sustainable and Functional Applications. Int. J. Mol. Sci. 2022, 23, 2359. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23042359
AMA Style
Streimikyte P, Viskelis P, Viskelis J. Enzymes-Assisted Extraction of Plants for Sustainable and Functional Applications. International Journal of Molecular Sciences. 2022; 23(4):2359. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23042359
Chicago/Turabian StyleStreimikyte, Paulina, Pranas Viskelis, and Jonas Viskelis. 2022. "Enzymes-Assisted Extraction of Plants for Sustainable and Functional Applications" International Journal of Molecular Sciences 23, no. 4: 2359. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23042359
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.