
Genetic Dissection of CRISPR-Cas9 Mediated Inheritance of Independently Targeted Alleles in Tobacco α-1,3-Fucosyltransferase 1 and β-1,2-Xylosyltransferase 1 Loci

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
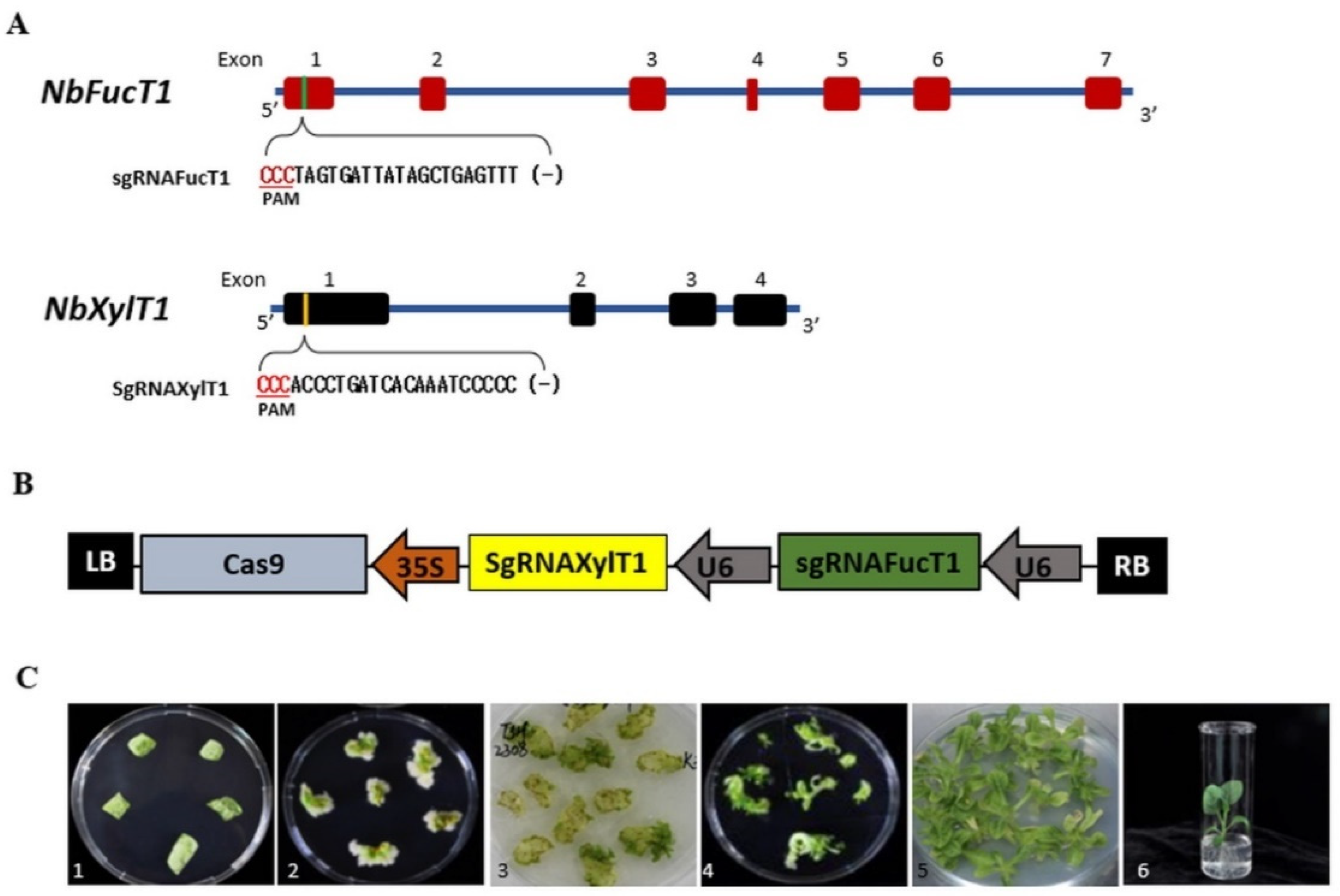
2.1. Generation of CRISPR/Cas9-Mediated Transgenic N. benthamiana
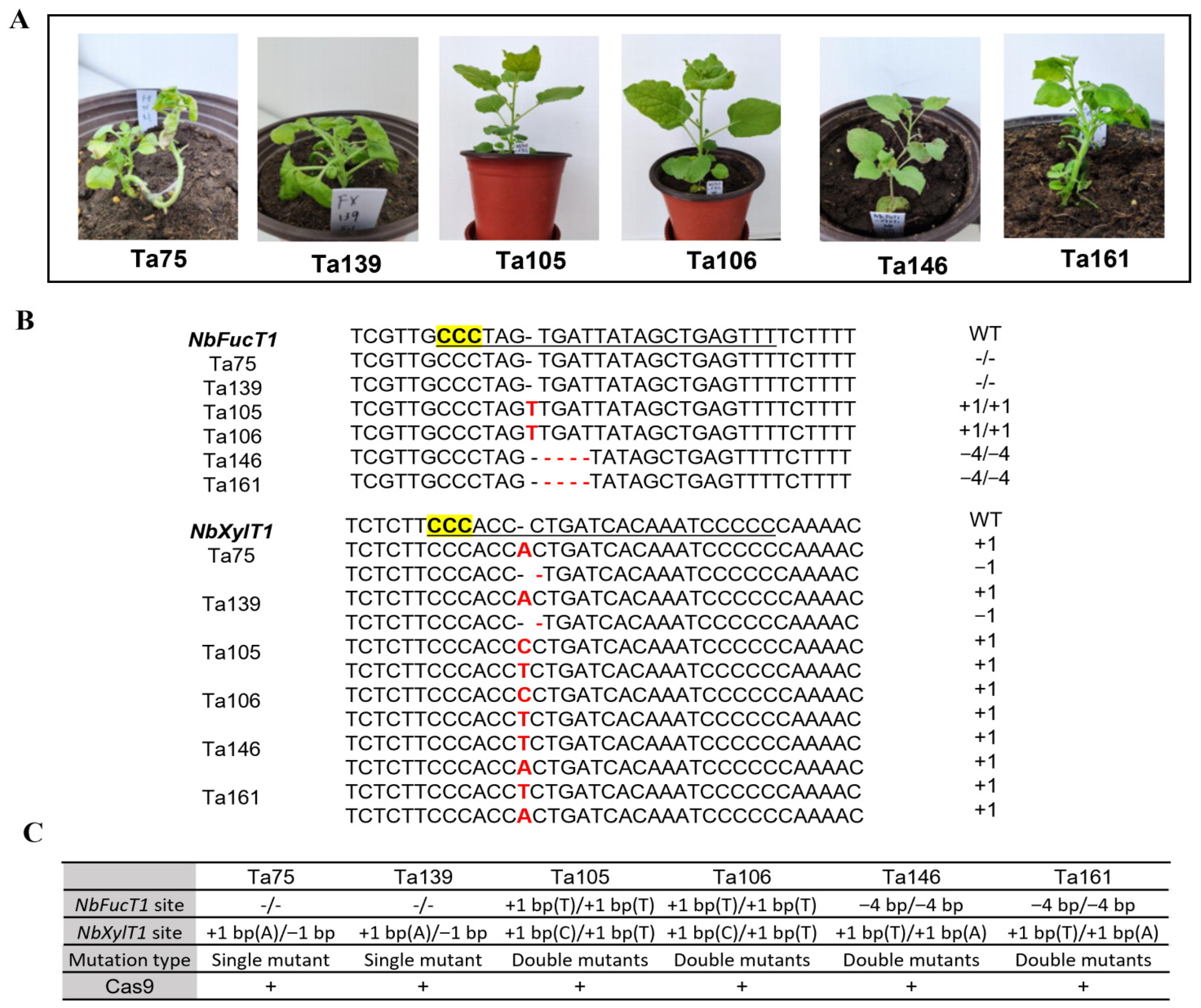
2.2. Identification of CRISPR-Cas9-Induced Mutations at Target Sites in T0 Plants
2.3. Inheritance of T0 Variant Target Mutations Evaluated in T1 Plants
2.4. Inheritance and Stability of T2 Plants Derived from Homozygotes
3. Discussion
4. Materials and Methods
4.1. Selection of sgRNA Target Sequences and Plasmid Construction
4.2. Agrobacterium-Mediated Transformation of N. benthamiana Plants
4.3. DNA Extraction and Identification of Induced Mutations
4.4. Analysis of the Presence of Cas9
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bhatta, B.P.; Malla, S. Improving Horticultural Crops via CRISPR/Cas9: Current Successes and Prospects. Plants 2020, 9, 1360. [Google Scholar] [CrossRef]
- Schaart, J.G.; van de Wiel, C.; Lotz, L.A.; Smulders, M.J. Opportunities for Products of New Plant Breeding Techniques. Trends Plant Sci. 2015, 21, 438–449. [Google Scholar] [CrossRef] [PubMed]
- Salgotra, R.K.; Stewart, C.N. Functional Markers for Precision Plant Breeding. Int. J. Mol. Sci. 2020, 21, 4792. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.; Gambhir, G.; Dass, A.; Tripathi, A.K.; Singh, A.; Jha, A.K.; Yadava, P.; Choudhary, M.; Rakshit, S. Genetically modified crops: Current status and future prospects. Planta 2020, 251, 1–27. [Google Scholar] [CrossRef]
- Ma, X.L.; Zhu, Q.L.; Chen, Y.L.; Liu, Y.G. CRISPR/Cas9 Platforms for Genome Editing in Plants: Developments and Applications. Mol. Plant 2016, 9, 961–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Massel, K.; Godwin, I.D.; Gao, C.X. Applications and potential of genome editing in crop improvement. Genome Biol. 2018, 19, 1–11. [Google Scholar] [CrossRef]
- Cermak, T.; Curtin, S.J.; Gil-Humanes, J.; Čegan, R.; Kono, T.J.Y.; Konečná, E.; Belanto, J.J.; Starker, C.G.; Mathre, J.W.; Greenstein, R.L.; et al. A Multipurpose Toolkit to Enable Advanced Genome Engineering in Plants. Plant Cell 2017, 29, 1196–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wada, N.; Ueta, R.; Osakabe, Y.; Osakabe, K. Precision genome editing in plants: State-of-the-art in CRISPR/Cas9-based genome engineering. BMC Plant Biol. 2020, 20, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Subburaj, S.; Tu, L.; Jin, Y.-T.; Bae, S.; Seo, P.J.; Jung, Y.J.; Lee, G.-J. Targeted genome editing, an alternative tool for trait improvement in horticultural crops. Hortic. Environ. Biotechnol. 2016, 57, 531–543. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Tu, L.; Subburaj, S.; Bae, S.; Lee, G.-J. Simultaneous targeting of duplicated genes in Petunia protoplasts for flower color modification via CRISPR-Cas9 ribonucleoproteins. Plant Cell Rep. 2020, 40, 1037–1045. [Google Scholar] [CrossRef]
- Michno, J.-M.; Virdi, K.; Stec, A.O.; Liu, J.; Wang, X.; Xiong, Y.; Stupar, R.M. Integration, abundance, and transmission of mutations and transgenes in a series of CRISPR/Cas9 soybean lines. BMC Biotechnol. 2020, 20, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, C.; Ye, L.; Qin, L.; Liu, X.; He, Y.; Wang, J.; Chen, L.; Lu, G. CRISPR/Cas9-mediated efficient and heritable targeted mutagenesis in tomato plants in the first and later generations. Sci. Rep. 2016, 6, 24765. [Google Scholar] [CrossRef]
- Feng, Z.; Mao, Y.; Xu, N.; Zhang, B.; Wei, P.; Yang, D.-L.; Wang, Z.; Zhang, Z.; Zheng, R.; Yang, L.; et al. Multigeneration analysis reveals the inheritance, specificity, and patterns of CRISPR/Cas-induced gene modifications in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 4632–4637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.; Yang, B.; Weeks, D.P. Efficient CRISPR/Cas9-Mediated Gene Editing in Arabidopsis thaliana and Inheritance of Modified Genes in the T2 and T3 Generations. PLoS ONE 2014, 9, e99225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giddings, G.; Brooks, D.; Carter, A. Transgenic plants as factories for biopharmaceuticals. Nat. Biotechnol. 2000, 18, 1151–1155. [Google Scholar] [CrossRef] [PubMed]
- Goodin, M.M.; Zaitlin, D.; Naidu, R.A.; Lommel, S.A. Nicotiana benthamiana: Its History and Future as a Model for Plant–Pathogen Interactions. Mol. Plant-Microbe Interact. 2014, 2015, 28–39. [Google Scholar] [CrossRef]
- Yao, J.; Weng, Y.; Dickey, A.; Wang, K.Y. Plants as Factories for Human Pharmaceuticals: Applications and Challenges. Int. J. Mol. Sci. 2015, 16, 28549–28565. [Google Scholar] [CrossRef]
- Huang, T.; Armstrong, B.; Schindele, P.; Puchta, H. Efficient gene targeting in Nicotiana tabacum using CRISPR/SaCas9 and temperature tolerant LbCas12a. Plant Biotechnol. J. 2021, 19, 1314–1324. [Google Scholar] [CrossRef]
- Jansing, J.; Sack, M.; Augustine, S.M.; Fischer, R.; Bortesi, L. CRISPR/Cas9-mediated knockout of six glycosyltransferase genes in Nicotiana benthamiana for the production of recombinant proteins lacking beta-1,2-xylose and core alpha-1,3-fucose. Plant Biotechnol. J. 2019, 17, 350–361. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Liu, X.; Fan, C.; Li, T.; Qin, H.; Li, X.; Chen, K.; Zheng, Y.; Chen, F.; Xu, Y. Enhancement of Tobacco (Nicotiana tabacum L.) Seed Lipid Content for Biodiesel Production by CRISPR-Cas9-Mediated Knockout of NtAn1. Front. Plant Sci. 2021, 11. [Google Scholar] [CrossRef]
- Niedbala, G.; Niazian, M.; Sabbatini, P. Modeling Agrobacterium-Mediated Gene Transformation of Tobacco (Nicotiana tab-acum)-A Model Plant for Gene Transformation Studies. Front. Plant Sci. 2021, 12, 695110. [Google Scholar] [CrossRef] [PubMed]
- Gebhardt, C. The historical role of species from the Solanaceae plant family in genetic research. Theor. Appl. Genet. 2016, 129, 2281–2294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Zhang, J.; Wei, P.; Zhang, B.; Gou, F.; Feng, Z.; Mao, Y.; Yang, L.; Zhang, H.; Xu, N.; et al. The CRISPR/Cas9 system produces specific and homozygous targeted gene editing in rice in one generation. Plant Biotechnol. J. 2014, 12, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.-F.; Li, H.; Qin, R.-Y.; Li, J.; Qiu, C.-H.; Yang, Y.-C.; Ma, H.; Li, L.; Wei, P.-C.; Yang, J.-B. Generation of inheritable and “transgene clean” targeted genome-modified rice in later generations using the CRISPR/Cas9 system. Sci. Rep. 2015, 5, 11491. [Google Scholar] [CrossRef] [Green Version]
- Ishizaki, T. CRISPR/Cas9 in rice can induce new mutations in later generations, leading to chimerism and unpredicted seg-regation of the targeted mutation. Mol. Breed. 2016, 36, 1–15. [Google Scholar] [CrossRef]
- Butler, N.M.; Atkins, P.A.; Voytas, D.F.; Douches, D.S. Generation and Inheritance of Targeted Mutations in Potato (Solanum tuberosum L.) Using the CRISPR/Cas System. PLoS ONE 2015, 10, e0144591. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.; Zhang, B.; Ding, W.; Liu, X.; Yang, D.-L.; Wei, P.; Cao, F.; Zhu, S.; Zhang, F.; Mao, Y.; et al. Efficient genome editing in plants using a CRISPR/Cas system. Cell Res. 2013, 23, 1229–1232. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Wang, G.; Ma, S.; Xie, X.; Wu, X.; Zhang, X.; Wu, Y.; Zhao, P.; Xia, Q. CRISPR/Cas9-mediated targeted mutagenesis in Nicotiana tabacum. Plant Mol. Biol. 2014, 87, 99–110. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, Q.; Zhu, Q.; Liu, W.; Chen, Y.; Qiu, R.; Wang, B.; Yang, Z.; Li, H.; Lin, Y.; et al. A Robust CRISPR/Cas9 System for Convenient, High-Efficiency Multiplex Genome Editing in Monocot and Dicot Plants. Mol. Plant 2015, 8, 1274–1284. [Google Scholar] [CrossRef]
- Zhang, N.; Roberts, H.M.; Van Eck, J.; Martin, G.B. Generation and Molecular Characterization of CRISPR/Cas9-Induced Mutations in 63 Immunity-Associated Genes in Tomato Reveals Specificity and a Range of Gene Modifications. Front. Plant Sci. 2020, 11, 10. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.W.; Kim, S.; Kim, Y.; Kweon, J.; Kim, H.S.; Bae, S.; Kim, J.-S. Analysis of off-target effects of CRISPR/Cas-derived RNA-guided endonucleases and nickases. Genome Res. 2013, 24, 132–141. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Cradick, T.; Brown, M.T.; Deshmukh, H.; Ranjan, P.; Sarode, N.; Wile, B.M.; Vertino, P.M.; Stewart, F.J.; Bao, G. CRISPR/Cas9 systems have off-target activity with insertions or deletions between target DNA and guide RNA sequences. Nucleic Acids Res. 2014, 42, 7473–7485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Modrzejewski, D.; Hartung, F.; Lehnert, H.; Sprink, T.; Kohl, C.; Keilwagen, J.; Wilhelm, R. Which Factors Affect the Occur-rence of Off-Target Effects Caused by the Use of CRISPR/Cas: A Systematic Review in Plants. Front. Plant Sci. 2020, 11, 574959. [Google Scholar] [CrossRef] [PubMed]
- Subburaj, S.; Chung, S.J.; Lee, C.; Ryu, S.M.; Kim, D.H.; Kim, J.S.; Bae, S.; Lee, G.J. Site-directed mutagenesis in Petunia x hybrida protoplast system using direct delivery of purified recombinant Cas9 ribonucleoproteins. Plant Cell Rep. 2016, 35, 1535–1544. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Target Gene | No. of Plants Examined | No. of Mutated Regenerants | Mutation Rate by Locus | ||||
|---|---|---|---|---|---|---|---|
| NbFucT1 | NbXylT1 | Both Loci | NbFucT1 | NbXylT1 | Both Loci | ||
| NbFucT1 | 248 | 56 | 62 | 51 | 22.5% | 25% | 20.56% |
| NbXylT1 | |||||||
| Target Gene: NbFucT1 | Target Gene: NbXylT1 | |||||||
|---|---|---|---|---|---|---|---|---|
| Mutant | Detected Mutations | Mutation Type | Total | Ratio | Detected Mutations | Mutation Type | Total | Ratio |
| Ta75 | No mutation | Wild type | 3 | 17.6% | + 1bp/−1 bp | Bi-allele | 16 | 94.% |
| Ta139 | No mutation | Wild type | +1 bp/−1 bp | Bi-allele | ||||
| Ta158 | No mutation | Wild type | +1 bp/+1 bp | Bi-allele | ||||
| Ta3 | −9 bp/−9 bp | Homozygote | 9 | 52.9% | −11 bp/+1 bp | Bi-allele | ||
| Ta12 | −9 bp/−9 bp | Homozygote | −11 bp/+1 bp | Bi-allele | ||||
| Ta18 | −9 bp/−9 bp | Homozygote | −11 bp/+1 bp | Bi-allele | ||||
| Ta27 | −9 bp/−9 bp | Homozygote | −11 bp/+1 bp | Bi-allele | ||||
| Ta105 | +1 bp/+1 bp | Homozygote | +1 bp/+1 bp | Bi-allele | ||||
| Ta106 | +1 bp/+1 bp | Homozygote | +1 bp/+1 bp | Bi-allele | ||||
| Ta146 | −4 bp/−4 bp | Homozygote | +1 bp/+1 bp | Bi-allele | ||||
| Ta161 | −4 bp/−4 bp | Homozygote | +1 bp/+1 bp | Bi-allele | ||||
| Ta122 | −113 bp/−113 bp | Homozygote | +1 bp/+1 bp | Bi-allele | ||||
| Ta117 | +1 bp/−9 bp | Bi-allele | 5 | 29.4% | +1 bp/−9 bp | Bi-allele | ||
| Ta133 | +1 bp/−9 bp | Bi-allele | +1 bp/−9 bp | Bi-allele | ||||
| Ta237 | +1 bp/−4 bp | Bi-allele | +1 bp/−9 bp | Bi-allele | ||||
| Ta248 | +1 bp/+1 bp | Bi-allele | +1 bp/+1 bp | Bi-allele | ||||
| Ta77 | +1 bp/−4 bp | Bi-allele | No Mutation | Wild type | 1 | 5.8% | ||
| Target Gene: NbFucT1 | ||||||
|---|---|---|---|---|---|---|
| T0 Generation | T1 Generation | |||||
| Plant ID | Zygosity (Detected Variation) | Plant ID | Zygosity | Detected Variation | No. of Plants | Ratio |
| Ta75 | Wild type (no mutation) | Bi-allele | 0 | 0.0% | ||
| Homozygote | 0 | 0.0% | ||||
| 75-2, 75-12, 75-14, 75-15 75-16, 75-18 | Chimera | +1 bp/−6 bp/−5 bp/−4 bp * WT/+1 bp/−6 bp/−5 bp/−4 bp * | 4 2 | 36.4% 18.2% | ||
| 75-3, 75-4, 75-11, 75-13, 75-17 | Wild type | No mutation | 5 | 45.5% | ||
| Ta139 | Wild type (no mutation) | Bi-allele | 0 | 0.0% | ||
| Homozygote | 0 | 0.0% | ||||
| 139-2, 139-16 139-1 | Chimera | +1 bp/−6 bp/WT/−5 bp/−4 bp * WT/+1 bp/−6 bp/−5 bp/−4 bp * | 2 1 | 25.0% 12.5% | ||
| 139-11, 139-12, 139-13, 139-14, 139-15 | Wild type | No mutation | 5 | 62.5% | ||
| Ta105 | Homozygote (+1 bp(T)/+1 bp(T)) | 105-4 | Bi-allele | +1 bp(G)/ −1 bp * | 1 | 10.0% |
| 105-2, 105-11, 105-12, 105-14, 105-15, 105-16, 105-17, 105-18 | Homozygote | +1 bp(T)/+1 bp(T) | 8 | 80.0% | ||
| 105-3 | +1 bp(G)/+1 bp(G) * | 1 | 10.0% | |||
| Chimera | 0 | 0.0% | ||||
| Wild type | No mutation | 0 | 0.0% | |||
| Ta106 | Homozygote (+1 bp(T)/+1 bp(T)) | Bi-allele | 0 | 0.0% | ||
| 106-2, 106-12, 106-13, 106-14, 106-15, 106-17, 106-18 | Homozygote | + 1bp(T)/+1 bp(T) | 7 | 87.5% | ||
| 106-1 | Chimera | −5 bp/−2 bp/−4 bp/+1 bp * | 1 | 12.5% | ||
| Wild type | No mutation | 0 | 0.0% | |||
| Ta146 | Homozygote (−4 bp/−4bp) | 146-16 | Bi-allele | −4 bp/+1 bp(T) * | 1 | 10.0% |
| 146-1, 146-2, 146-11, 146-12, 146-13, 146-17, 146-18 | Homozygote | −4 bp/−4 bp | 7 | 70.0% | ||
| 146-14 146-15 | Chimera | +1 bp(T)/−6 bp/−5 bp/−4 bp * −3 bp/+1 bp(T)/−6 bp/−5 bp * | 1 1 | 10.0% 10.0% | ||
| Wild type | No mutation | 0 | 0.0% | |||
| Ta161 | Homozygote (−4 bp/−4 bp) | Bi-allele | 0 | 0.0% | ||
| 161-1, 161-2, 161-11, 161-12, 161-13, 161-14, 161-15, 161-16, 161-18 | Homozygote | −4 bp/−4 bp | 9 | 90.0% | ||
| 161-17 | Chimera | +1 bp(T)/−6 bp/−5 bp/−4 bp * | 1 | 10.0% | ||
| Wild type | No mutation | 0 | 0.0% | |||
| Target Gene: NbXylT1 | ||||||
|---|---|---|---|---|---|---|
| T0 Generation | T1 Generation | |||||
| Plant ID | Zygosity (Detected Variation) | Plant ID | Zygosity | Detected Variation | No. of Plants | Ratio |
| Ta75 | Bi-allele (+1 bp(A)/−1 bp) | 75-11, 75-14, 75-15, 75-17 75-16 | Bi-allele | +1 bp(A)/−1 bp +1 bp(T)/−1 bp * | 4 1 | 36.4% 9.1% |
| 75-2, 75-12 75-3, 75-4, 75-13 | Homozygote | −1 bp/−1 bp +1 bp(A)/+1 bp(A) | 2 3 | 18.2% 27.3% | ||
| 75-18 | Chimera | +1 bp(T)/−1 bp/+1 bp(A) * | 1 | 9.1% | ||
| wild type | no mutation | 0 | 0.0% | |||
| Ta139 | Bi-allele (+1 bp(A)/−1 bp) | 139-1, 139-11, 139-12 139-15 | Bi-allele | +1 bp(A)/ −1 bp +1 bp(A)/+1 bp(T) * | 3 1 | 37.5% 12.5% |
| 139-13, 139-14, 139-16 | Homozygote | +1 bp(A)/+1 bp(A) | 3 | 37.5% | ||
| 139-2 | Chimera | +1 bp(A)/+1 bp(T)/ −1 bp * | 1 | 12.5% | ||
| Wild type | WT | 0 | 0.0% | |||
| Ta105 | Bi-allele (+1 bp(T)/+1 bp(C)) | 105-2, 105-11, 105-14, 105-15, 105-17 | Bi-allele | +1 bp(T)/+1 bp(C) | 5 | 50.0% |
| 105-12 105-16 105-3, 105-4 | Homozygote | +1 bp(T)/+1 bp(T) +1 bp(C)/+1 bp(C) +1 bp(A)/+1 bp(A) * | 1 1 2 | 10.0% 10.0% 20.0% | ||
| 105-18 | Chimera | +1 bp(T)/+1 bp(A)/−1 bp * | 1 | 10.0% | ||
| Wild type | WT | 0 | 0.0% | |||
| Ta106 | Bi-allele (+1 bp(T)/+1 bp(C)) | 106-1, 106-2, 106-13 | Bi-allele | +1 bp(T)/+1 bp(C) | 3 | 37.5% |
| 106-14, 106-15, 106-17 106-12 | Homozygote | +1 bp(T)/+1 bp(T) +1 bp(C)/+1 bp(C) | 3 1 | 37.5% 12.5% | ||
| 106-18 | Chimera | +1 bp(A)/+1 bp(T)/+1 bp(C) * | 1 | 12.5% | ||
| Wild type | WT | 0 | 0.0% | |||
| Ta146 | Bi-allele (+1 bp(A)/+1 bp(T)) | 146-1, 146-11, 146-12, 146-16, 146-17 146-15 | Bi-allele | +1 bp(A)/+1 bp(T) −1 bp/+1 bp(T) * | 5 1 | 50.0% 10.0% |
| 146-13 | Homozygote | +1 bp(A)/+1 bp(A) | 1 | 10.0% | ||
| 146-2, 146-14, 146-18 | Chimera | +1 bp(A)/+1 bp(T)/−1 bp * | 3 | 30.0% | ||
| Wild type | WT | 0 | 0.0% | |||
| Ta161 | Bi-allele (+1 bp(A)/+1 bp(T)) | 161-11, 161-13, 161-15 | Bi-allele | +1 bp(A)/+1 bp(T) | 3 | 30.0% |
| 161-1, 161-2, 161-12, 161-16, 161-17 161-14 | Homozygote | +1 bp(T)/+1 bp(T) +1 bp(A)/+1 bp(A) | 5 1 | 50.0% 10.0% | ||
| 161-18 | Chimera | +1 bp(A)/+1 bp(T)/−1 bp * | 1 | 10.0% | ||
| Wild type | WT | 0 | 0.0% | |||
| Generation | NbFucT1 | NbXylT1 | ||||
|---|---|---|---|---|---|---|
| Plant ID | cas9 | Mutation Detected | Mutation Type | Mutation Detected | Mutation Type | |
| T0 plant | Ta161 | + | +1 bp(T)/+1 bp (A) | Bi-allelic | −4 bp/−4 bp | Homozygous |
| T1 plant | Ta161-1 | + | +1 bp(T)/+1 bp (T) | Homozygous | −4 bp/−4 bp | Homozygous |
| T2 plant | Ta161-1-1 | + | +1 bp(T)/+1 bp (T) | Homozygous | −4 bp/−4 bp | Homozygous |
| Ta161-1-2 | − | +1 bp(T)/+1 bp (T) | Homozygous | −4 bp/−4 bp | Homozygous | |
| Ta161-1-3 | − | +1 bp(T)/+1 bp (T) | Homozygous | −4 bp/−4 bp | Homozygous | |
| Ta161-1-4 | + | +1 bp(T)/+1 bp (T) | Homozygous | −4 bp/−4 bp | Homozygous | |
| Ta161-1-5 | + | +1 bp(T)/+1 bp (T) | Homozygous | −4 bp/−4 bp | Homozygous | |
| Ta161-1-6 | − | +1 bp(T)/+1 bp (T) | Homozygous | −4 bp/−4 bp | Homozygous | |
| Ta161-1-7 | − | +1 bp(T)/+1 bp (T) | Homozygous | −4 bp/−4 bp | Homozygous | |
| Ta161-1-8 | + | +1 bp(T)/+1 bp (T) | Homozygous | −4 bp/−4 bp | Homozygous | |
| Ta161-1-9 | − | +1 bp(T)/+1 bp (T) | Homozygous | −4 bp/−4 bp | Homozygous | |
| Ta161-1-10 | + | +1 bp(T)/+1 bp (T) | Homozygous | −4 bp/−4 bp | Homozygous | |
| Ta161-1-11 | + | +1 bp(T)/+1 bp (T) | Homozygous | −4 bp/−4 bp | Homozygous | |
| Ta161-1-12 | + | +1 bp(T)/+1 bp (T) | Homozygous | −4 bp/−4 bp | Homozygous | |
| Ta161-1-13 | + | +1 bp(T)/+1 bp (T) | Homozygous | −4 bp/−4 bp | Homozygous | |
| Ta161-1-14 | − | +1 bp(T)/+1 bp (T) | Homozygous | −4 bp/−4 bp | Homozygous | |
| Ta161-1-15 | − | +1 bp(T)/+1 bp (T) | Homozygous | −4 bp/−4 bp | Homozygous | |
| Ta161-1-16 | + | +1 bp(T)/+1 bp (T) | Homozygous | −4 bp/−4 bp | Homozygous | |
| Ta161-1-17 | − | +1 bp(T)/+1 bp (T) | Homozygous | −4 bp/−4 bp | Homozygous | |
| Ta161-1-18 | − | +1 bp(T)/+1 bp (T) | Homozygous | −4 bp/−4 bp | Homozygous | |
| Ta161-1-19 | + | +1 bp(T)/+1 bp (T) | Homozygous | −4 bp/−4 bp | Homozygous | |
| Ta161-1-20 | − | +1 bp(T)/+1 bp (T) | Homozygous | −4 bp/−4 bp | Homozygous | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, H.; Ahn, J.-Y.; Yan, F.; Ran, Y.; Koo, O.; Lee, G.-J. Genetic Dissection of CRISPR-Cas9 Mediated Inheritance of Independently Targeted Alleles in Tobacco α-1,3-Fucosyltransferase 1 and β-1,2-Xylosyltransferase 1 Loci. Int. J. Mol. Sci. 2022, 23, 2450. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052450
Song H, Ahn J-Y, Yan F, Ran Y, Koo O, Lee G-J. Genetic Dissection of CRISPR-Cas9 Mediated Inheritance of Independently Targeted Alleles in Tobacco α-1,3-Fucosyltransferase 1 and β-1,2-Xylosyltransferase 1 Loci. International Journal of Molecular Sciences. 2022; 23(5):2450. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052450
Chicago/Turabian StyleSong, Hayoung, Ju-Young Ahn, Fanzhuang Yan, Yidong Ran, Okjae Koo, and Geung-Joo Lee. 2022. "Genetic Dissection of CRISPR-Cas9 Mediated Inheritance of Independently Targeted Alleles in Tobacco α-1,3-Fucosyltransferase 1 and β-1,2-Xylosyltransferase 1 Loci" International Journal of Molecular Sciences 23, no. 5: 2450. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052450