
Function and Regulation of microRNA171 in Plant Stem Cell Homeostasis and Developmental Programing


1
Department of Botany and Plant Pathology, Purdue University, West Lafayette, IN 47907, USA
2
Purdue Center for Plant Biology, Purdue University, West Lafayette, IN 47907, USA
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2022, 23(5), 2544; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052544
Submission received: 27 January 2022
/
Revised: 21 February 2022
/
Accepted: 22 February 2022
/
Published: 25 February 2022
(This article belongs to the Special Issue New Horizons in Plant Cell Signaling)
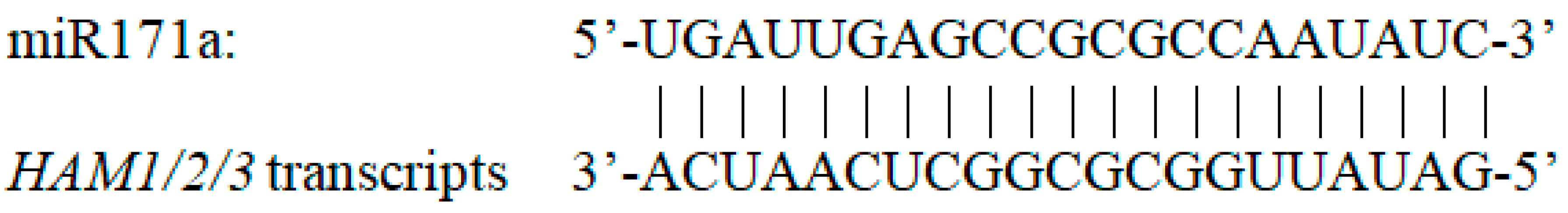{kind=link}
{kind=link}
{kind=link}
Abstract
:MicroRNA171 (miR171), a group of 21-nucleotide single-strand small RNAs, is one ancient and conserved microRNA family in land plants. This review focuses on the recent progress in understanding the role of miR171 in plant stem cell homeostasis and developmental patterning, and the regulation of miR171 by developmental cues and environmental signals. Specifically, miR171 regulates shoot meristem activity and phase transition through repressing the HAIRY MERISTEM (HAM) family genes. In the model species Arabidopsis, miR171 serves as a short-range mobile signal, which initiates in the epidermal layer of shoot meristems and moves downwards within a limited distance, to pattern the apical-basal polarity of gene expression and drive stem cell dynamics. miR171 levels are regulated by light and various abiotic stresses, suggesting miR171 may serve as a linkage between environmental factors and cell fate decisions. Furthermore, miR171 family members also demonstrate both conserved and lineage-specific functions in land plants, which are summarized and discussed here.
1. Introduction
MicroRNAs (miRNAs) are 20–24 ribonucleotide single-strand non-coding small RNA molecules, which regulate gene expression post-transcriptionally [1,2]. In animals, plants and fungi, miRNAs recognize their target messenger RNAs (mRNAs) through complementary base pairing, leading to the repression of gene expression at the transcriptional and post-transcriptional level [1,2,3]. The firstly identified endogenous miRNAs are lin-4 and let-7, which control the timing of larval development in the nematode Caenorhabditis elegans [4,5]. In plants, a considerable number of studies have demonstrated that miRNAs play vital roles in various processes, including developmental programming and responses to both biotic and abiotic stresses [1,2,3,6,7]. miRNAs share features with the small interfering RNAs (siRNAs) of the RNA interference (RNAi pathway) [8]; however, one of the differences between these two types of small RNAs is that miRNAs are endogenous products, while siRNAs are primarily exogenous in their origin, such as the virus and transgenes [8].
Plant miRNAs not only serve as local signals, but also can function non-cell-autonomously [9]. Mobile miRNAs travel from one cell to another, likely via plasmodesmata in plants [10,11,12]. Several miRNA species move over a long distance within a plant through vascular tissues or even between the plants and their interacting organisms, such as parasitic plants and pathogens [12,13,14,15,16,17]. Recent studies demonstrated that several miRNA species travel via the environment from one plant to its neighboring plant and induce gene silencing [17,18]. These results suggest that exogenous miRNAs, potentially as signaling molecules, are able to post-transcriptionally regulate expression of endogenous genes in plants [17,18]. In the model species of flowering plants, Arabidopsis, the cell-to-cell movement of miRNAs relays key positional signals during plant morphogenesis [19,20,21,22,23,24,25,26,27]. For example, the concentration gradient of the miR165/166 family members specifies the adaxial–abaxial polarity in developing leaves and determines the central stele fate in roots [20,22,23,28,29]; a recent study demonstrated that two related receptor-like kinases, BARELY ANY MERISTEM (BAM) 1 and BAM2, are required for the movement of miR165/166 in roots [29]. In Arabidopsis shoot apical meristems (SAMs), the movement of miR394 from the epidermis to inner cell layers is essential to maintain stem cell competency [24,30]. In addition, the cell-to-cell movement of miRNAs seems to be directional, suggesting a yet-undefined mechanism that gates their movement [31]. In this review, we summarize the recent progress in understanding the function and regulation of microRNA170/171 (hereafter referred to as miR171), a group of conserved miRNA species widely present in land plants. We discuss how they act as a short-range mobile signal in patterning gene expression in shoot meristems and how they are regulated in response to environmental signals and developmental cues.
2. miRNA171 and Its Targets
In general, plant MIRNA genes are transcribed into relatively long primary transcripts (pri-miRNAs), which are subsequently processed into shorter pre-miRNAs [32]. pre-miRNAs fold into characteristic stem-loop hairpin structures, which are cleaved by DICER-LIKE (DCL) proteins, forming 21- to 24-nt miRNA/miRNA* duplexes. The antisense miRNA* is then degraded and the mature miRNA is embedded into an RNA-induced silencing complex (RISC) that contains the ARGONAUTE (AGO) proteins [33,34,35]. miR171 represents a small group of the 21-nuleotide single-strand micro RNAs [1,6,7,36,37]. In Arabidopsis, the total miR171 species are from the products of four genes located at different regions of the genome, including MICRORNA171A (MIR171A), MICRORNA171B (MIR171B), MICRORNA171C (MIR171C) and MICRORNA170 (MIR170). The MIR170 and MIR171A genes were initially identified in 2002 [1,6,36,37] and the MIR171B and MIR171C genes and their products were identified two years later [7]. The sequence of mature miR171a (UGAUUGAGCCGCGCCAAUAUC) is perfectly complementary to its target mRNAs (Figure 1). In contrast, miR171b and miR171c have the identical miRNA sequences (UUGAGCCGUGCCAAUAUCACG) [7], which are slightly different with that of miR171a. In addition, the mature miR170 (UGAUUGAGCCGUGUCAAUAUC) differs from miR171a at the 12th and 14th positions, with two base pair mismatches to their targets [36]. Arabidopsis miR171 specifically targets and represses three HAIRY MERISTEM (HAM) family genes [1,6,7,36,37], which are also named as LOST MERISTEM (LOM) and SCARECROW-LIKE (SCL) genes in different plant species [6,38,39,40]. HAM family genes encode the GRAS (GAI, RGA and SCR) domain transcription factors [41,42,43], which belong to the GRAS superfamily [41,44]. There are four HAM family genes (HAM1-4) in Arabidopsis [41,45]. Among them, HAM1, HAM2 and HAM3 contain the conserved miR171-binding sequence, 5′-GATATTGGCGCGGCTCAATCA-3′, within their coding sequences [41,45] and their transcripts are specifically targeted by miR171 (Figure 1) [36,37,38]. In contrast, Arabidopsis HAM4 (AtHAM4) is classified into a different group and HAM4 lacks a conserved miR171 target site [41,45]. HAM family genes regulate multiple growth and developmental processes in Arabidopsis [38,39,40,41,42,43,45,46,47,48,49], including the control of stem cell identity and homeostasis in shoot apical meristems and axillary meristems. Shoot meristems harbor a small group of undifferentiated stem cells, maintaining a balance between the self-renewal of stem cells and cell differentiation into lateral organs. In shoot meristems, the miR171-regulated HAM1 and HAM2 play crucial and shared function in control of the WUSCHEL (WUS)-CLAVATA3 (CLV3) feedback loop and drive the stem cell homeostasis [41,42,45,46,47,48,49,50,51]. A recent work demonstrated that HAM3, the other HAM member regulated by miR171 in Arabidopsis, plays a dispensable or minor role in determining the apical-basal patterning of shoot meristems, but likely shares redundant functions with HAM1 and HAM2 in control of other aspects of shoot and root development [42,48].
Several miR171 species also target the NSP2 (NODULATION SIGNALING PATHWAY 2) family members in Medicago truncatula. As an example, Medicago miR171h (Mtr-miR171h, 5′-CGAGCCGAAUCAAUAUCACUC-3′) specifically recognizes and cleaves the transcript of MtNSP2. MtNSP2 also encodes a GRAS domain protein, which functions as a transcriptional activator and plays a key role in both arbuscular mycorrhizal colonization and nodulation signaling pathways [52,53,54,55]. The NSP2 family is sister to the HAM family, and they both belong to the GRAS superfamily. Although NSP2 orthologs are widely distributed in land plants and the cleavage of NSP2 family members by miR171 is also found in other plant species, including Lotus japonicus [56,57,58], a phylogenetic study suggests that the miR171h-NSP2 regulatory module is only conserved in the angiosperms that enable arbuscular mycorrhizal symbiosis [53].
The SU(VAR)3-9 HOMOLOG8 (SUVH8)/SET DOMAIN GROUP 21 gene is regulated by the MIR171/170 products through a different mechanism [59]. During microRNA biogenesis, usually the guide strand (named as miRNA) is retained and the passenger strand (named as miRNA*) is degraded, subsequent to association of the miRNA/miRNA* duplex with RISCs [32]. Under certain circumstances, the miRNA* is retained as the guide strand [60]. In Arabidopsis, miR170* and miR171a* trigger the tissue-specific silencing of SUVH8 [59,61]. miR171a* was preferentially loaded into AGO1 and SUVH8 mRNA degradation products were identified corresponding to the miR171a*-mediated cleavage [59]. The plants that specifically altered the miR171a*-SUVH8 balance (through overexpressing miR171a* target mimic or expressing the miR171* resistant SUVH8) showed dwarfed stature, long and curved leaves and reduced fertility [59]. The miR171a* target site in SUVH8 is only found in the Arabidopsis lineage, seems not to be conserved across species [59]. The molecular mechanism underlying the selection and regulation of miR171*/miR170* as the guide strand in Arabidopsis under different physiological and environmental contexts deserves further studies [62,63].
3. Function of miR171
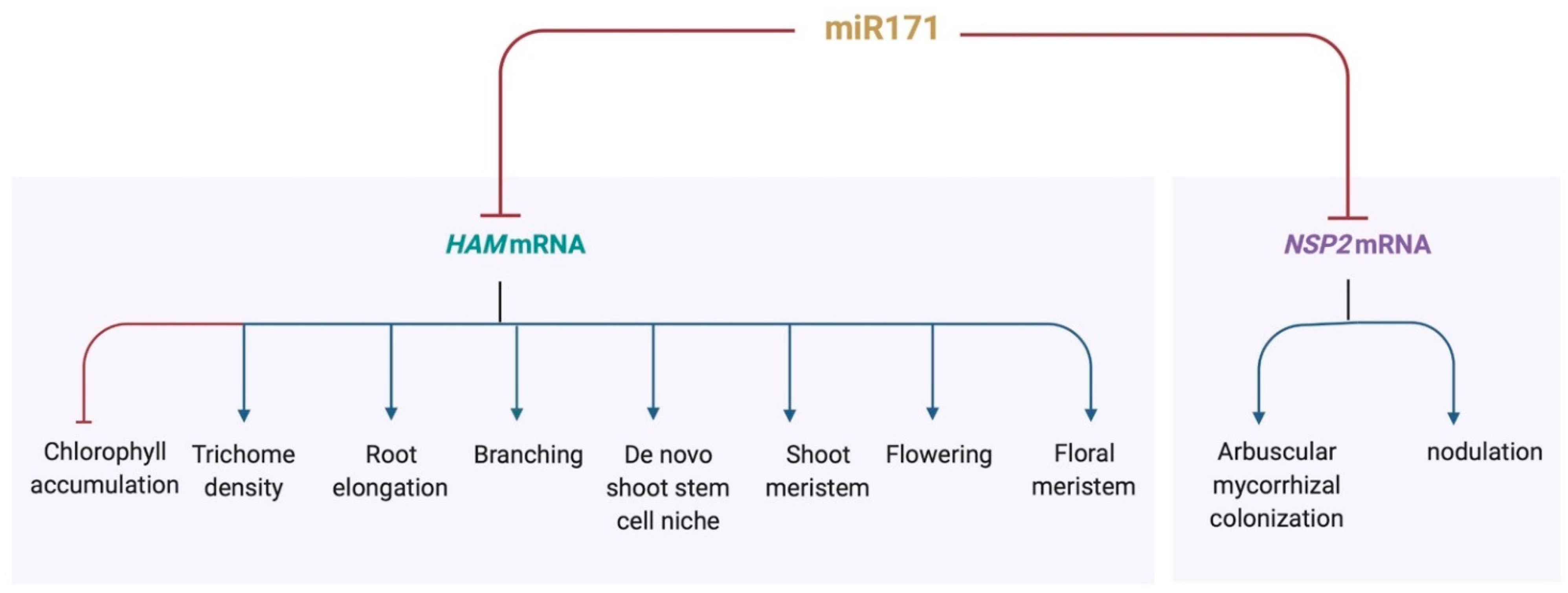
In Arabidopsis, miR171 plays an essential role during shoot meristem development. The overexpression of miR171 in Arabidopsis results in the defects in shoot stem cell initiation and maintenance, with the mis-regulated CLV3 expression pattern, disorganized meristem structure, and reduced shoot branches [38,42,47,48,49], which are comparable to the phenotypes of the ham1ham2ham3 triple or ham1ham2 double loss of function mutants [38,41,42,46,47,48]. A 3D computational model predicts that the partial loss of function of HAM genes induced by miR171 overproduction is sufficient to disturb the apical-basal pattern of the CLV3 expression domain in Arabidopsis SAMs [47]. This simulation has been validated by RNA in situ hybridization assays in the 35S::MIR171 plants, showing the CLV3 mRNA pattern with the greatly reduced (but detectable) expression in the first two upper layers and the increased level in the deep cell layers of the SAMs [47]. Furthermore, in 35S::MIR171 transgenic plants, the CLV3 expression in developing axillary meristems is always confined into deep cell layers, demonstrating the role of miR171 in both established and de novo stem cell niches [48]. Besides the shoot meristem defects, the miR171 overexpression plants also display the altered shoot architecture, leaf shape, leaf color and chlorophyll accumulation, flower structure, trichome morphology and primary root elongation, suggesting that miR171 functions a regulatory hub in control of diverse developmental processes (Figure 2) [38,39,40,41]. Among them, the miR171-HAM pathway regulates phase transition and trichome distribution, likely through inhibiting the activity of miR156-targeted SQUAMOSA PROMOTER BINDING PROTEIN LIKE (SPL) genes [39], the conserved and key regulator of phase transition [64,65,66]. In addition, the key role of miR171 in plant development is revealed by the silent mutations in the miR171 binding sites of the HAM genes, which lead to the miR171 uncleavable (insensitive) targets. These mutated HAM genes produces mRNAs that cannot be recognized by miR171 but in which the amino acid sequences of encoded proteins are unchanged [38,67]. When the miR171 insensitive HAM1 is specifically expressed in the epidermal layer, it leads to the reduced CLV3 expression in the epidermis and abnormal meristem development [46]. The accumulation of the miR171 insensitive HAM genes also results in developmental defects in cotyledons, young seedlings, leaves and shoot branching [38,67]. All these results suggest that it is crucial to spatially confine the HAM expression domain by miR171 in Arabidopsis.
The role of miR171 in regulating meristem identity seems to be highly conserved across flowering plants. As a result, the miR171-HAM regulatory module controls important agronomic traits in crops. For example, overexpression of Sly-miR171 in tomato leads to disturbed shoot development [56,68,69], which is similar to phenotypes of the ham loss-of-function mutants in petunia [43] and in pepper [70]. Utilizing a two component system, overexpression of either Sly-MIR171a or Sly-MIR171b leads to enlarged meristems and growth arrest after producing few leaves in tomato [56]. In monocots, the overexpression of osa-miR171c in rice (Oryza sativa) leads to the enlarged shoot apex and the misregulation of key meristem regulators, including the rice orthologs of CLV1, CLV3 and WOX4 [71]. The overexpression of miR171 in barley (Hordeum vulgare) results in similar phenotypes, including the reduced shoot branching [72]. In addition, miR171 participates in developmental phase transition in monocots (including barley and rice), likely through the conserved miR156-SPL signaling circuit [71,72].
miR171h negatively regulates arbuscular mycorrhizal and nodule symbiosis by targeting MtNSP2 in Medicago [52,53,54,73] (Figure 2). Fungal colonization and nodulation numbers was reduced in the roots of miR171h overexpression plants, similar to nsp2 loss of function mutants [53,74,75]. In addition, the miR171h-NSP2 module regulates root endosymbiosis likely through the phytohormone cytokinin responsive signaling [54]. Interestingly, Medicago miR171b (Mtr-miR171b) has evolved a target site mismatch at the identified cleavage site, which protects its target gene MtHAM1 from the cleavage by other miR171 family members. As a result, arbuscular mycorrhizal symbiosis is stimulated [73]. This regulation involving Mtr-miR171b seems to be specific in mycotrophic species [73]. In rice roots, a tissue-specific transposon transcribes competing endogenous RNAs, which also act as target mimics and interrupt the function of miR171, protecting the targets—OsHAM genes from the degradation [76]. In the future, it will be interesting to explore whether similar mechanisms exist in other miRNA families and in other biological processes.
The function of miR171 in flowering plants also has been uncovered by expressing the miRNA target MIMICs (MIMs) or short tandem target MIMICs (STTMs), which are uncleavable targets for sequestering miRNAs into non-productive interactions [77,78]. The 35S:MIM171a and 35S:MIM170 transgenic Arabidopsis plants showed similar phenotypes, with round leaves in pale green, defective anthesis and reduced fertility [79]. On the contrary, the 35S:MIM171b and 35S:MIM171c plants are comparable to the wild type control, which is likely due to the genetic redundancy and partial subfunctionalization among members of the MIR171/170 family. The STTM transgenic line silencing miR171 also lead to the developmental defects in the monocot rice, including the semi-dwarf stature, enclosed panicles, and drooping flag leaves [80]. Overexpression of STTM171 in tomato results in irregular compound leaf morphology, increased branch number and male sterility, demonstrating the role of sly-miR171 in regulating axillary meristem activity, compound leaf development and anther development [81].
4. Expression Patterns and Regulation of miR171
The miR171 precursors and mature miR171 species are widely accumulated in different tissues of Arabidopsis, including cotyledons, hypocotyls, leaves, flowers and roots [1,36,38,59,82]. Confocal imaging of the fluorescent transcriptional reporters demonstrated that the MIR171A gene is highly and specifically expressed in the epidermal layer of embryos, vegetative SAMs, young leaves and stems, inflorescence SAMs and floral meristems [67,82,83]. In addition, the promoter activities of the other three MIR170/171 family genes (MIR171B, MIR171C and MIR170) are also specifically turned on in the epidermis of shoot meristems at both vegetative and reproductive stages, though the expression levels of the MIR171C and MIR170 genes are weaker than that of MIR171A and MIR171B [82]. These findings suggest that the epidermis is the major synthesis site for miR171 in Arabidopsis shoots. On the contrary, these MIR171 genes are not expressed in the root epidermis [82], suggesting MIR171 expression patterns in shoots and roots are controlled by different mechanisms.
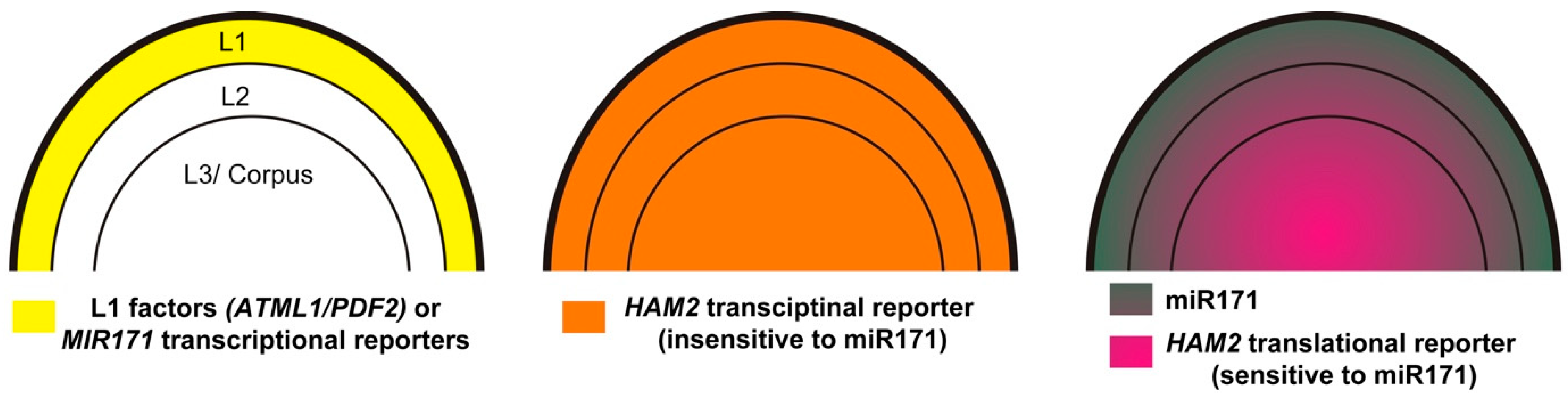
The miRNA in situ hybridization has shown that the localization of mature miR171 is not only highly accumulated in the epidermal layer but also detectable in the second and third cell layers of the SAM [82]. In addition, the comparison between the expression patterns of the miR171-sensitive and miR171-insensitive HAM reporters in the SAMs serves as a direct readout of the miR171 localization (Figure 3) [82]. For example, a miR171-insensitive HAM2 transcriptional fluorescent reporter is ubiquitously expressed in all the layers of Arabidopsis SAMs [82]. In contrast, the miR171-sensitive HAM2 translational fluorescent reporter is highly expressed in deep cell layers but repressed in the upper layers, forming a concentration gradient along the apical-basal axis of SAMs (Figure 3). This concentration gradient is complementary to the pattern of mature miR171 in the SAMs [82] and crucial for the apical-basal patterning of SAMs [47] (Figure 3). All these results suggest that miR171 is a short-range mobile signal, moving downwards across one to two cell layers and shaping the apical-basal patterns of HAM proteins in Arabidopsis SAMs [67,82]. The highly restricted movement of miR171 within Arabidopsis SAMs is consistent with the findings that the cell-to-cell movement of microRNAs in general is highly controlled and limited in Arabidopsis SAMs [31,82]. It will be interesting to explore how the movement of miR171 is governed in the future studies. In addition, although the function of miR171 family in control of root endosymbiosis is well studied as described above, whether miR171 moves within roots and whether miR171 serves as a short-range mobile signal in this process needs further studies.
The epidermis-specific expression of Arabidopsis MIR171 genes is directly determined by two closely related homeodomain leucine zipper class IV (HD-ZIP IV) transcription factors—ARABIDOPSIS MERISTEM LAYER 1 (ATML1) and PROTODERMAL FACTOR 2 (PDF2) [82]. ATML1 and PDF2 function as key regulators in the epidermal specification pathway and they both are specifically expressed in the epidermal layer of SAMs and floral meristems [84,85,86,87,88]. Both the yeast one hybrid and gel mobility shift assays demonstrated that ATML1 and PDF2 proteins directly bind to the promoters of MIR171 genes. The conserved L1-box motifs in the promoters of MIR171 genes, which serve as consensus binding sites for ATML1/PDF2, mediate the physical interactions between the MIR171 promoter DNAs and ATML1/PDF2 proteins. These L1-box motifs are also required for the activation of MIR171 genes by ATML1, because MIR171 genes fail to express in the L1 layer once these cis-elements are mutated [82]. In addition, compared to that in wild type, expression of the MIR171 reporters is absent or greatly reduced in the atml1 pdf2 double mutants, demonstrating that ATML1/PDF2 are required for the MIR171 expression in Arabidopsis (Figure 3) [82]. Using ATML1/PDF2 as a functional input, a 3D computational model was established to simulate and quantitatively evaluate the function of a L1(ATML1/PDF2)-miR171-HAM signaling cascade in Arabidopsis SAMs. Both in silico prediction and in vivo experimental results demonstrated that the transient activation of ATML1 in the SAMs rapidly induce the MIR171 promoter activities, subsequently leading to the reduction of HAM expression [82], suggesting an essential role of the L1-miR171-HAM signaling cascade in Arabidopsis SAMs.
The expression of miR171 is regulated by multiple environmental signals [7,89,90]. Determined by the RNA gel blot, the miR171 level increases during the light period of the daytime and decreases in the dark period, and this oscillatory pattern of miR171 accumulation is governed by light instead of the circadian clock [89]. The light-induced miR171 expression might have a connection with the findings that the miR171-HAM pathway affects chlorophyll biosynthesis under light [40]. Interestingly, osa-miR171 species in rice are also induced by light [71]. Their accumulation peak in the early morning and the expression of four rice HAM homologs show the complementary fluctuations [71]. Besides light, miR171 responds to stresses in different plant species [90,91,92,93]. Through comprehensive microarray expression analyses in Arabidopsis, it was found that several miR171 species (miR171b and miR171c) are induced by abiotic stresses, including high salinity, drought and low temperature [90]. Small RNA profiles generated by next-generation sequencing in potato demonstrated that the potato miR171a, miR171b and miR171c are also drought-induced [91]. In addition, Mtr-miR171h expression is induced in mycorrhizal roots and root nodules in Medicago [52,75]. It is also induced by high phosphate nutrition and repressed by phosphate starvation [52,75], suggesting a potential linkage between the nutritional status and endosymbiosis of plants [52,53,54,55]. All these findings lead to a hypothesis that miR171 likely serves as a signaling bridge linking environmental factors to plant growth and development. miR171 also demonstrates high expression in undifferentiated callus from a number of species, including rice, Japanese larch (Larix kaempferi), orange (Citrus sinensis L. Osb.) and lilies (Lilium pumilum) [94,95,96,97,98,99,100]. It will be interesting to determine whether miR171 involves callus formation, regeneration or somatic embryogenesis in the future.
The miR171 abundance is also positively regulated by a small peptide generated from its own pre-miRNA, suggesting an additional layer of regulation of miR171 [101,102]. In Arabidopsis, Medicago and grape, the primary transcripts of miRNAs encode small regulatory peptides called the miRNA-encoded peptides (miPEPs), which specifically activate the transcription of their own miRNAs [101,102]. An exogenous miPEP also sufficiently activates the production of its related miRNA in plants, suggesting potential agronomical applications. This regulation seems to be conserved in several different miRNA families. As an example, the Medicago miR171b produces miPEP171b, which in turn enhances the production of miR171b and plays a role in arbuscular mycorrhizal symbiosis [73]. It will be interesting to explore the molecular basis by which miPEPs positively regulate miRNAs.
5. Evolution of miR171 in Land Plants
The miR171 family is one of the ancient MIRNA families that have been identified in the common ancestor of all embryophytes [103]. miR171 is broadly present across land plants, from bryophytes to angiosperms (flowering plants) [103,104]. Consistently, their targets, the HAM family genes, are also widely present in land plants and they likely originated prior to the divergence of bryophytes [41,45]. The miR171 binding site, 5′-GATATTGGCGCGGCTCAATCA- 3′, seems to be an ancestral trait for the HAM family [45]. This site is highly conserved in almost all the non-angiosperm HAM members examined to date, including the HAM homologs identified in bryophytes, lycophytes, ferns and gymnosperms [41,45,105]. In angiosperms, the HAM family expanded and diversified into two distinct groups: Type I and Type II [41,45,106]. The miR171-binding sequence is retained in most Type II HAM members but lost in the majority of Type I HAM members [41,45]. Confocal imaging results demonstrated that the conserved miR171-binding sites in the HAM homologs from different species are recognized by miR171 in Arabidopsis SAMs, leading to the cleavage similar to endogenous Arabidopsis HAM2 transcripts [45]. Thus, the conserved miR171-binding sequence in these species suggests strong selective pressure in retaining this regulation and limiting HAM expression. Furthermore, the specific repression of HAM expression by miR171 is likely crucial for the conserved functions of the HAM family [45], which needs future studies. Recently, it was found that in the moss P. patens, the miR171-regulated PpHAM/PpGRAS12 plays a role in meristem formation [107] and the PpHAM/PpGRAS12 overexpression induces the enlarged and multiple apical meristems. The role of miR171 is also suggested in the liverwort Marchantia polymorpha, based on the RNA-seq and degradome analyses [108,109].
The number of MIR170/171 family genes demonstrated dramatic variations among different species (as per miRBase) [110]. The miR171 precursors also display high diversifications and members of the MIR170/171 family likely evolved in a lineage-specific manner [110,111,112,113]. In addition, the promoters of MIR170/171 family genes have undergone most sequence diversification within species, suggesting that different members may have evolved specialized expression patterns to differentially regulate or fine tune target genes [110]. In contrast, the mature miR171 sequences are highly conserved in a number of flowering plants. In four plant species, including Arabidopsis, poplar (P. trichocarpa), rice and grape (V. vinifera), 21 out of 34 miR171 sequences are nearly identical, with only different nucleotides at one or two positions [110]. In addition, a separate study of the miR171 phylogeny in several fruit species leads to a similar conclusion [113]. Nevertheless, despite strong conservation of mature miR171 sequences in general, a small proportion of miR171 species have critical sequence variations and target non-HAM genes as described above [52,56,110,114].
6. Future Directions and Perspectives
In Arabidopsis shoot apical meristems, miR171 only moves across one or two cell layers, leading to the question whether the mobility of miRNA is a characteristic of the specific miRNA or of the specific cell type. It will be interesting to explore whether the cell-to-cell movement of miR171 is also highly restricted in leaves, roots and stems, and whether its movement in these tissues involves key developmental decision and cellular patterning as well. The level of miR171 is directly controlled by the epidermal specification pathway (the ATML1 and PDF2 transcription factors) [82], and it is also induced by environmental signals, such as light and abiotic stresses [7,89,90]. It will be very informative to determine molecular mechanisms by which miR171 links environmental factors to plant development. Furthermore, miR171 is widely present in land plants, however, its function and regulation in non-seed plants is largely unknown. The future studies of miR171 in seed-free vascular plants, such as in the fern Ceratopteris richardii [44,115,116], will provide more insights into the evolution of this key miRNA family in land plants.
Author Contributions
H.H. and Y.Z. conceived the directions; H.H. and Y.Z. wrote and revised the manuscript; H.H. prepared the figures. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Purdue Start-up package, and funds from the Purdue Center for Plant Biology and NSF IOS-1931114 to Y.Z.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors apologize to the colleagues whose work was not cited here due to the space limitations.
Conflicts of Interest
Authors declare no conflict of interest.
References
- Reinhart, B.J.; Weinstein, E.G.; Rhoades, M.W.; Bartel, B.; Bartel, D.P. MicroRNAs in plants. Genes Dev. 2002, 16, 1616–1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Mei, J.; Ren, G. Plant microRNAs: Biogenesis, homeostasis, and degradation. Front. Plant Sci. 2019, 10, 360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.-K. Reconstituting plant miRNA biogenesis. Proc. Natl. Acad. Sci. USA 2008, 105, 9851–9852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Reinhart, B.J.; Slack, F.J.; Basson, M.; Pasquinelli, A.E.; Bettinger, J.C.; Rougvie, A.E.; Horvitz, H.R.; Ruvkun, G. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature 2000, 403, 901–906. [Google Scholar] [CrossRef]
- Llave, C.; Kasschau, K.D.; Rector, M.A.; Carrington, J.C. Endogenous and silencing-associated small RNAs in plants. Plant Cell 2002, 14, 1605–1619. [Google Scholar] [CrossRef] [Green Version]
- Sunkar, R.; Zhu, J.-K. Novel and stress-regulated microRNAs and other small RNAs from Arabidopsis. Plant Cell 2004, 16, 2001–2019. [Google Scholar] [CrossRef] [Green Version]
- Carthew, R.W.; Sontheimer, E.J. Origins and mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Rechavi, O. Plant and animal small RNA communications between cells and organisms. Nat. Rev. Mol. Cell Biol. 2021, 1–19. [Google Scholar] [CrossRef]
- Vatén, A.; Dettmer, J.; Wu, S.; Stierhof, Y.-D.; Miyashima, S.; Yadav, S.R.; Roberts, C.J.; Campilho, A.; Bulone, V.; Lichtenberger, R. Callose biosynthesis regulates symplastic trafficking during root development. Dev. Cell 2011, 21, 1144–1155. [Google Scholar] [CrossRef] [Green Version]
- Molnar, A.; Melnyk, C.; Baulcombe, D.C. Silencing signals in plants: A long journey for small RNAs. Genome Biol. 2011, 12, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ham, B.-K.; Lucas, W.J. Phloem-mobile RNAs as systemic signaling agents. Annu. Rev. Plant Biol. 2017, 68, 173–195. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, X.; Xu, W.; Liu, T.; Cai, C.; Chen, L.; Clark, C.B.; Ma, J. Unidirectional movement of small RNAs from shoots to roots in interspecific heterografts. Nat. Plants 2021, 7, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Shahid, S.; Kim, G.; Johnson, N.R.; Wafula, E.; Wang, F.; Coruh, C.; Bernal-Galeano, V.; Phifer, T.; Depamphilis, C.W.; Westwood, J.H. MicroRNAs from the parasitic plant Cuscuta campestris target host messenger RNAs. Nature 2018, 553, 82–85. [Google Scholar] [CrossRef]
- Zhang, T.; Zhao, Y.-L.; Zhao, J.-H.; Wang, S.; Jin, Y.; Chen, Z.-Q.; Fang, Y.-Y.; Hua, C.-L.; Ding, S.-W.; Guo, H.-S. Cotton plants export microRNAs to inhibit virulence gene expression in a fungal pathogen. Nat. Plants 2016, 2, 16153. [Google Scholar] [CrossRef]
- Lewsey, M.G.; Hardcastle, T.J.; Melnyk, C.W.; Molnar, A.; Valli, A.; Urich, M.A.; Nery, J.R.; Baulcombe, D.C.; Ecker, J.R. Mobile small RNAs regulate genome-wide DNA methylation. Proc. Natl. Acad. Sci. USA 2016, 113, E801–E810. [Google Scholar] [CrossRef] [Green Version]
- Brosnan, C.A.; Mitter, N. miRNA communication on another level. Nat. Plants 2021, 7, 1328–1329. [Google Scholar] [CrossRef]
- Betti, F.; Ladera-Carmona, M.J.; Weits, D.A.; Ferri, G.; Iacopino, S.; Novi, G.; Svezia, B.; Kunkowska, A.B.; Santaniello, A.; Piaggesi, A. Exogenous miRNAs induce post-transcriptional gene silencing in plants. Nat. Plants 2021, 7, 1379–1388. [Google Scholar] [CrossRef]
- Emery, J.F.; Floyd, S.K.; Alvarez, J.; Eshed, Y.; Hawker, N.P.; Izhaki, A.; Baum, S.F.; Bowman, J.L. Radial patterning of Arabidopsis shoots by class III HD-ZIP and KANADI genes. Curr. Biol. 2003, 13, 1768–1774. [Google Scholar] [CrossRef] [Green Version]
- Juarez, M.T.; Kui, J.S.; Thomas, J.; Heller, B.A.; Timmermans, M.C. microRNA-mediated repression of rolled leaf1 specifies maize leaf polarity. Nature 2004, 428, 84–88. [Google Scholar] [CrossRef]
- McConnell, J.R.; Emery, J.; Eshed, Y.; Bao, N.; Bowman, J.; Barton, M.K. Role of PHABULOSA and PHAVOLUTA in determining radial patterning in shoots. Nature 2001, 411, 709–713. [Google Scholar] [CrossRef]
- Carlsbecker, A.; Lee, J.-Y.; Roberts, C.J.; Dettmer, J.; Lehesranta, S.; Zhou, J.; Lindgren, O.; Moreno-Risueno, M.A.; Vatén, A.; Thitamadee, S. Cell signalling by microRNA165/6 directs gene dose-dependent root cell fate. Nature 2010, 465, 316–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyashima, S.; Koi, S.; Hashimoto, T.; Nakajima, K. Non-cell-autonomous microRNA165 acts in a dose-dependent manner to regulate multiple differentiation status in the Arabidopsis root. Development 2011, 138, 2303–2313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knauer, S.; Holt, A.L.; Rubio-Somoza, I.; Tucker, E.J.; Hinze, A.; Pisch, M.; Javelle, M.; Timmermans, M.C.; Tucker, M.R.; Laux, T. A protodermal miR394 signal defines a region of stem cell competence in the Arabidopsis shoot meristem. Dev. Cell 2013, 24, 125–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brosnan, C.A.; Sarazin, A.; Lim, P.; Bologna, N.G.; Hirsch-Hoffmann, M.; Voinnet, O. Genome-scale, single-cell-type resolution of micro RNA activities within a whole plant organ. EMBO J. 2019, 38, e100754. [Google Scholar] [CrossRef] [PubMed]
- Klesen, S.; Hill, K.; Timmermans, M.C. Small RNAs as plant morphogens. Curr. Top. Dev. Biol. 2020, 137, 455–480. [Google Scholar]
- Maizel, A.; Markmann, K.; Timmermans, M.; Wachter, A. To move or not to move: Roles and specificity of plant RNA mobility. Curr. Opin. Plant Biol. 2020, 57, 52–60. [Google Scholar] [CrossRef]
- Tatematsu, K.; Toyokura, K.; Miyashima, S.; Nakajima, K.; Okada, K. A molecular mechanism that confines the activity pattern of miR165 in A rabidopsis leaf primordia. Plant J. 2015, 82, 596–608. [Google Scholar] [CrossRef]
- Fan, P.; Aguilar, E.; Bradai, M.; Xue, H.; Wang, H.; Rosas-Diaz, T.; Tang, W.; Wolf, S.; Zhang, H.; Xu, L.; et al. The receptor-like kinases BAM1 and BAM2 are required for root xylem patterning. Proc. Natl. Acad. Sci. USA 2021, 118, e2022547118. [Google Scholar] [CrossRef]
- Kitagawa, M.; Jackson, D. Plasmodesmata-mediated cell-to-cell communication in the shoot apical meristem: How stem cells talk. Plants 2017, 6, 12. [Google Scholar] [CrossRef]
- Skopelitis, D.S.; Hill, K.; Klesen, S.; Marco, C.F.; von Born, P.; Chitwood, D.H.; Timmermans, M.C. Gating of miRNA movement at defined cell-cell interfaces governs their impact as positional signals. Nat. Commun. 2018, 9, 3107. [Google Scholar] [CrossRef]
- Voinnet, O. Origin, biogenesis, and activity of plant microRNAs. Cell 2009, 136, 669–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumberger, N.; Baulcombe, D. Arabidopsis ARGONAUTE1 is an RNA Slicer that selectively recruits microRNAs and short interfering RNAs. Proc. Natl. Acad. Sci. USA 2005, 102, 11928–11933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papp, I.; Mette, M.F.; Aufsatz, W.; Daxinger, L.; Schauer, S.E.; Ray, A.; Van Der Winden, J.; Matzke, M.; Matzke, A.J. Evidence for nuclear processing of plant micro RNA and short interfering RNA precursors. Plant Physiol. 2003, 132, 1382–1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, I.R.; Zhang, X.; Lu, C.; Johnson, L.; Meyers, B.C.; Green, P.J.; Jacobsen, S.E. Dissecting Arabidopsis thaliana DICER function in small RNA processing, gene silencing and DNA methylation patterning. Nat. Genet. 2006, 38, 721–725. [Google Scholar] [CrossRef]
- Llave, C.; Xie, Z.; Kasschau, K.D.; Carrington, J.C. Cleavage of Scarecrow-like mRNA targets directed by a class of Arabidopsis miRNA. Science 2002, 297, 2053–2056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhoades, M.W.; Reinhart, B.J.; Lim, L.P.; Burge, C.B.; Bartel, B.; Bartel, D.P. Prediction of plant microRNA targets. Cell 2002, 110, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Mai, Y.-X.; Zhang, Y.-C.; Luo, Q.; Yang, H.-Q. MicroRNA171c-targeted SCL6-II, SCL6-III, and SCL6-IV genes regulate shoot branching in Arabidopsis. Mol. Plant 2010, 3, 794–806. [Google Scholar] [CrossRef] [Green Version]
- Xue, X.-Y.; Zhao, B.; Chao, L.-M.; Chen, D.-Y.; Cui, W.-R.; Mao, Y.-B.; Wang, L.-J.; Chen, X.-Y. Interaction between two timing microRNAs controls trichome distribution in Arabidopsis. PLoS Genet. 2014, 10, e1004266. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Hu, X.; Cai, W.; Huang, W.; Zhou, X.; Luo, Q.; Yang, H.; Wang, J.; Huang, J. Arabidopsis miR171-targeted scarecrow-like proteins bind to GT cis-elements and mediate gibberellin-regulated chlorophyll biosynthesis under light conditions. PLoS Genet. 2014, 10, e1004519. [Google Scholar] [CrossRef] [Green Version]
- Engstrom, E.M.; Andersen, C.M.; Gumulak-Smith, J.; Hu, J.; Orlova, E.; Sozzani, R.; Bowman, J.L. Arabidopsis homologs of the petunia hairy meristem gene are required for maintenance of shoot and root indeterminacy. Plant Physiol. 2011, 155, 735–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulze, S.; Schäfer, B.N.; Parizotto, E.A.; Voinnet, O.; Theres, K. LOST MERISTEMS genes regulate cell differentiation of central zone descendants in Arabidopsis shoot meristems. Plant J. 2010, 64, 668–678. [Google Scholar] [CrossRef] [PubMed]
- Stuurman, J.; Jäggi, F.; Kuhlemeier, C. Shoot meristem maintenance is controlled by a GRAS-gene mediated signal from differentiating cells. Genes Dev. 2002, 16, 2213–2218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geng, Y.; Cai, C.; McAdam, S.A.; Banks, J.A.; Wisecaver, J.H.; Zhou, Y. A de novo transcriptome assembly of Ceratopteris richardii provides insights into the evolutionary dynamics of complex gene families in land plants. Genome Biol. Evol. 2021, 13, evab042. [Google Scholar] [CrossRef]
- Geng, Y.; Guo, L.; Han, H.; Liu, X.; Banks, J.A.; Wisecaver, J.H.; Zhou, Y. Conservation and diversification of HAIRY MERISTEM gene family in land plants. Plant J. 2021, 106, 366–378. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, X.; Engstrom, E.M.; Nimchuk, Z.L.; Pruneda-Paz, J.L.; Tarr, P.T.; Yan, A.; Kay, S.A.; Meyerowitz, E.M. Control of plant stem cell function by conserved interacting transcriptional regulators. Nature 2015, 517, 377. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Yan, A.; Han, H.; Li, T.; Geng, Y.; Liu, X.; Meyerowitz, E.M. HAIRY MERISTEM with WUSCHEL confines CLAVATA3 expression to the outer apical meristem layers. Science 2018, 361, 502–506. [Google Scholar] [CrossRef] [Green Version]
- Han, H.; Geng, Y.; Guo, L.; Yan, A.; Meyerowitz, E.M.; Liu, X.; Zhou, Y. The overlapping and distinct roles of HAM family genes in Arabidopsis shoot meristems. Front. Plant Sci. 2020, 11, 1375. [Google Scholar] [CrossRef]
- Han, H.; Liu, X.; Zhou, Y. Transcriptional circuits in control of shoot stem cell homeostasis. Curr. Opin. Plant Biol. 2020, 53, 50–56. [Google Scholar] [CrossRef]
- Gruel, J.; Deichmann, J.; Landrein, B.; Hitchcock, T.; Jönsson, H. The interaction of transcription factors controls the spatial layout of plant aerial stem cell niches. NPJ Syst. Biol. Appl. 2018, 4, 36. [Google Scholar] [CrossRef] [Green Version]
- Biedermann, S.; Laux, T. Plant Development: Adding HAM to Stem Cell Control. Curr. Biol. 2018, 28, R1261–R1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devers, E.A.; Branscheid, A.; May, P.; Krajinski, F. Stars and symbiosis: MicroRNA-and microRNA*-mediated transcript cleavage involved in arbuscular mycorrhizal symbiosis. Plant Physiol. 2011, 156, 1990–2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauressergues, D.; Delaux, P.M.; Formey, D.; Lelandais-Brière, C.; Fort, S.; Cottaz, S.; Bécard, G.; Niebel, A.; Roux, C.; Combier, J.P. The microRNA miR171h modulates arbuscular mycorrhizal colonization of Medicago truncatula by targeting NSP2. Plant J. 2012, 72, 512–522. [Google Scholar] [CrossRef] [PubMed]
- Ariel, F.; Brault-Hernandez, M.; Laffont, C.; Huault, E.; Brault, M.; Plet, J.; Moison, M.; Blanchet, S.; Ichanté, J.L.; Chabaud, M. Two direct targets of cytokinin signaling regulate symbiotic nodulation in Medicago truncatula. Plant Cell 2012, 24, 3838–3852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maillet, F.; Poinsot, V.; André, O.; Puech-Pagès, V.; Haouy, A.; Gueunier, M.; Cromer, L.; Giraudet, D.; Formey, D.; Niebel, A. Fungal lipochitooligosaccharide symbiotic signals in arbuscular mycorrhiza. Nature 2011, 469, 58–63. [Google Scholar] [CrossRef]
- Hendelman, A.; Kravchik, M.; Stav, R.; Frank, W.; Arazi, T. Tomato HAIRY MERISTEM genes are involved in meristem maintenance and compound leaf morphogenesis. J. Exp. Bot. 2016, 67, 6187–6200. [Google Scholar] [CrossRef] [Green Version]
- De Luis, A.; Markmann, K.; Cognat, V.; Holt, D.B.; Charpentier, M.; Parniske, M.; Stougaard, J.; Voinnet, O. Two microRNAs linked to nodule infection and nitrogen-fixing ability in the legume Lotus japonicus. Plant Physiol. 2012, 160, 2137–2154. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.; Wu, Y.; Liu, C.-C.; Liu, L.-W.; Ma, F.-F.; Wu, X.-Y.; Wu, M.; Hang, Y.-Y.; Chen, J.-Q.; Shao, Z.-Q. Identification of arbuscular mycorrhiza (AM)-responsive microRNAs in tomato. Front. Plant Sci. 2016, 7, 429. [Google Scholar] [CrossRef] [Green Version]
- Manavella, P.A.; Koenig, D.; Rubio-Somoza, I.; Burbano, H.A.; Becker, C.; Weigel, D. Tissue-specific silencing of Arabidopsis SU (VAR) 3-9 HOMOLOG8 by miR171a. Plant Physiol. 2013, 161, 805–812. [Google Scholar] [CrossRef] [Green Version]
- Treiber, T.; Treiber, N.; Meister, G. Regulation of microRNA biogenesis and its crosstalk with other cellular pathways. Nat. Rev. Mol. Cell Biol. 2019, 20, 5–20. [Google Scholar] [CrossRef]
- Manavella, P.A.; Koenig, D.; Weigel, D. Plant secondary siRNA production determined by microRNA-duplex structure. Proc. Natl. Acad. Sci. USA 2012, 109, 2461–2466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medley, J.C.; Panzade, G.; Zinovyeva, A.Y. microRNA strand selection: Unwinding the rules. Wiley Interdiscip. Rev. RNA 2021, 12, e1627. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.-w.; Meng, J.; Cui, J.; Luan, Y.-s. Characterization and function of microRNA∗ s in plants. Front. Plant Sci. 2017, 8, 2200. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Wang, H. The miR156/SPL module, a regulatory hub and versatile toolbox, gears up crops for enhanced agronomic traits. Mol. Plant 2015, 8, 677–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jerome Jeyakumar, J.M.; Ali, A.; Wang, W.-M.; Thiruvengadam, M. Characterizing the role of the miR156-SPL Network in plant development and stress response. Plants 2020, 9, 1206. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Ye, M.; Sang, M.; Wu, R. A regulatory network for miR156-SPL module in Arabidopsis thaliana. Int. J. Mol. Sci. 2019, 20, 6166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takanashi, H.; Sumiyoshi, H.; Mogi, M.; Hayashi, Y.; Ohnishi, T.; Tsutsumi, N. miRNAs control HAM1 functions at the single-cell-layer level and are essential for normal embryogenesis in Arabidopsis. Plant Mol. Biol. 2018, 96, 627–640. [Google Scholar] [CrossRef]
- Huang, W.; Peng, S.; Xian, Z.; Lin, D.; Hu, G.; Yang, L.; Ren, M.; Li, Z. Overexpression of a tomato miR171 target gene Sl GRAS 24 impacts multiple agronomical traits via regulating gibberellin and auxin homeostasis. Plant Biotechnol. J. 2017, 15, 472–488. [Google Scholar] [CrossRef]
- Huang, W.; Xian, Z.; Kang, X.; Tang, N.; Li, Z. Genome-wide identification, phylogeny and expression analysis of GRAS gene family in tomato. BMC Plant Biol. 2015, 15, 209. [Google Scholar] [CrossRef] [Green Version]
- David-Schwartz, R.; Borovsky, Y.; Zemach, H.; Paran, I. CaHAM is autoregulated and regulates CaSTM expression and is required for shoot apical meristem organization in pepper. Plant Sci. 2013, 203, 8–16. [Google Scholar] [CrossRef]
- Fan, T.; Li, X.; Yang, W.; Xia, K.; Ouyang, J.; Zhang, M. Rice osa-miR171c mediates phase change from vegetative to reproductive development and shoot apical meristem maintenance by repressing four OsHAM transcription factors. PLoS ONE 2015, 10, e0125833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curaba, J.; Talbot, M.; Li, Z.; Helliwell, C. Over-expression of microRNA171 affects phase transitions and floral meristem determinancy in barley. BMC Plant Biol. 2013, 13, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couzigou, J.-M.; Lauressergues, D.; André, O.; Gutjahr, C.; Guillotin, B.; Bécard, G.; Combier, J.-P. Positive gene regulation by a natural protective miRNA enables arbuscular mycorrhizal symbiosis. Cell Host Microbe 2017, 21, 106–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofferek, V.; Mendrinna, A.; Gaude, N.; Krajinski, F.; Devers, E.A. MiR171h restricts root symbioses and shows like its target NSP2 a complex transcriptional regulation in Medicago truncatula. BMC Plant Biol. 2014, 14, 199. [Google Scholar] [CrossRef]
- Devers, E.A. MiR 171 h restricts root symbioses and shows, like its target NSP 2, a complex transcriptional regulation in Medicago truncatula. In The Model Legume Medicago Truncatula; John Wiley & Sons: Hoboken, NJ, USA, 2019; pp. 975–986. [Google Scholar]
- Cho, J.; Paszkowski, J. Regulation of rice root development by a retrotransposon acting as a microRNA sponge. eLife 2017, 6, e30038. [Google Scholar] [CrossRef] [Green Version]
- Franco-Zorrilla, J.M.; Valli, A.; Todesco, M.; Mateos, I.; Puga, M.I.; Rubio-Somoza, I.; Leyva, A.; Weigel, D.; García, J.A.; Paz-Ares, J. Target mimicry provides a new mechanism for regulation of microRNA activity. Nat. Genet. 2007, 39, 1033–1037. [Google Scholar] [CrossRef]
- Yan, J.; Gu, Y.; Jia, X.; Kang, W.; Pan, S.; Tang, X.; Chen, X.; Tang, G. Effective small RNA destruction by the expression of a short tandem target mimic in Arabidopsis. Plant Cell 2012, 24, 415–427. [Google Scholar] [CrossRef] [Green Version]
- Todesco, M.; Rubio-Somoza, I.; Paz-Ares, J.; Weigel, D. A collection of target mimics for comprehensive analysis of microRNA function in Arabidopsis thaliana. PLoS Genet. 2010, 6, e1001031. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhang, J.; Yan, J.; Gou, F.; Mao, Y.; Tang, G.; Botella, J.R.; Zhu, J.-K. Short tandem target mimic rice lines uncover functions of miRNAs in regulating important agronomic traits. Proc. Natl. Acad. Sci. USA 2017, 114, 5277–5282. [Google Scholar] [CrossRef] [Green Version]
- Kravchik, M.; Stav, R.; Belausov, E.; Arazi, T. Functional characterization of microRNA171 family in tomato. Plants 2019, 8, 10. [Google Scholar] [CrossRef] [Green Version]
- Han, H.; Yan, A.; Li, L.; Zhu, Y.; Feng, B.; Liu, X.; Zhou, Y. A signal cascade originated from epidermis defines apical-basal patterning of Arabidopsis shoot apical meristems. Nat. Commun. 2020, 11, 1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parizotto, E.A.; Dunoyer, P.; Rahm, N.; Himber, C.; Voinnet, O. In vivo investigation of the transcription, processing, endonucleolytic activity, and functional relevance of the spatial distribution of a plant miRNA. Genes Dev. 2004, 18, 2237–2242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, M.; Katsumata, H.; Komeda, Y.; Takahashi, T. Regulation of shoot epidermal cell differentiation by a pair of homeodomain proteins in Arabidopsis. Development 2003, 130, 635–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, M.; Katsumata, H.; Abe, M.; Yabe, N.; Komeda, Y.; Yamamoto, K.T.; Takahashi, T. Characterization of the class IV homeodomain-leucine zipper gene family in Arabidopsis. Plant Physiol. 2006, 141, 1363–1375. [Google Scholar] [CrossRef] [Green Version]
- Lu, P.; Porat, R.; Nadeau, J.A.; O’Neill, S.D. Identification of a meristem L1 layer-specific gene in Arabidopsis that is expressed during embryonic pattern formation and defines a new class of homeobox genes. Plant Cell 1996, 8, 2155–2168. [Google Scholar]
- Nagata, K.; Ishikawa, T.; Kawai-Yamada, M.; Takahashi, T.; Abe, M. Ceramides mediate positional signals in Arabidopsis thaliana protoderm differentiation. Development 2021, 148, dev194969. [Google Scholar] [CrossRef]
- Iida, H.; Takada, S. A Quarter Century History of ATML1. Plant Embryog. 2021, 10, 1. [Google Scholar]
- Siré, C.; Moreno, A.B.; Garcia-Chapa, M.; López-Moya, J.J.; San Segundo, B. Diurnal oscillation in the accumulation of Arabidopsis microRNAs, miR167, miR168, miR171 and miR398. FEBS Lett. 2009, 583, 1039–1044. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.-H.; Tian, X.; Li, Y.-J.; Wu, C.-A.; Zheng, C.-C. Microarray-based analysis of stress-regulated microRNAs in Arabidopsis thaliana. RNA 2008, 14, 836–843. [Google Scholar] [CrossRef] [Green Version]
- Hwang, E.-W.; Shin, S.-J.; Yu, B.-K.; Byun, M.-O.; Kwon, H.-B. miR171 family members are involved in drought response in Solanum tuberosum. J. Plant Biol. 2011, 54, 43–48. [Google Scholar] [CrossRef]
- Kantar, M.; Unver, T.; Budak, H. Regulation of barley miRNAs upon dehydration stress correlated with target gene expression. Funct. Integr. Genom. 2010, 10, 493–507. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Elling, A.A.; Chen, B.; Deng, X. Differential expression of microRNAs in maize inbred and hybrid lines during salt and drought stress. Am. J. Plant Sci. 2010, 1, 69. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.-C.; Zhou, H.; Li, Y.; Chen, J.-Y.; Yang, J.-H.; Chen, Y.-Q.; Qu, L.-H. Rice embryogenic calli express a unique set of microRNAs, suggesting regulatory roles of microRNAs in plant post-embryogenic development. Febs Lett. 2006, 580, 5111–5116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Zhou, J.; Han, S.; Yang, W.; Li, W.; Wei, H.; Li, X.; Qi, L. Four abiotic stress-induced miRNA families differentially regulated in the embryogenic and non-embryogenic callus tissues of Larix leptolepis. Biochem. Biophys. Res. Commun. 2010, 398, 355–360. [Google Scholar] [CrossRef]
- Wu, X.-M.; Liu, M.-Y.; Ge, X.-X.; Xu, Q.; Guo, W.-W. Stage and tissue-specific modulation of ten conserved miRNAs and their targets during somatic embryogenesis of Valencia sweet orange. Planta 2011, 233, 495–505. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, S.; Han, S.; Wu, T.; Li, X.; Li, W.; Qi, L. Genome-wide identification of microRNAs in larch and stage-specific modulation of 11 conserved microRNAs and their targets during somatic embryogenesis. Planta 2012, 236, 647–657. [Google Scholar] [CrossRef]
- Li, W.-F.; Zhang, S.-G.; Han, S.-Y.; Wu, T.; Zhang, J.-H.; Qi, L.-W. The post-transcriptional regulation of LaSCL6 by miR171 during maintenance of embryogenic potential in Larix kaempferi (Lamb.) Carr. Tree Genet. Genomes 2014, 10, 223–229. [Google Scholar] [CrossRef]
- Li, H.; Zhang, J.; Yang, Y.; Jia, N.; Wang, C.; Sun, H. miR171 and its target gene SCL6 contribute to embryogenic callus induction and torpedo-shaped embryo formation during somatic embryogenesis in two lily species. Plant Cell Tissue Organ. Cult. (PCTOC) 2017, 130, 591–600. [Google Scholar] [CrossRef]
- Li, H.; Wang, J.; Yan, R.; Wang, C.; Sun, H. Functional characterization of the MiR171a promoter and endogenous target mimics identification in Lilium pumilum DC. Fisch. during somatic embryogenesis. Plant Cell Tissue Organ. Cult. (PCTOC) 2021, 144, 345–357. [Google Scholar] [CrossRef]
- Lauressergues, D.; Couzigou, J.-M.; San Clemente, H.; Martinez, Y.; Dunand, C.; Bécard, G.; Combier, J.-P. Primary transcripts of microRNAs encode regulatory peptides. Nature 2015, 520, 90–93. [Google Scholar] [CrossRef]
- Chen, Q.-J.; Deng, B.-H.; Gao, J.; Zhao, Z.-Y.; Chen, Z.-L.; Song, S.-R.; Wang, L.; Zhao, L.-P.; Xu, W.-P.; Zhang, C.-X.; et al. A miRNA-encoded small peptide, vvi-miPEP171d1, regulates adventitious root formation. Plant Physiol. 2020, 183, 656–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuperus, J.T.; Fahlgren, N.; Carrington, J.C. Evolution and functional diversification of MIRNA genes. Plant Cell 2011, 23, 431–442. [Google Scholar] [CrossRef] [Green Version]
- Jones-Rhoades, M.W.; Bartel, D.P. Computational identification of plant microRNAs and their targets, including a stress-induced miRNA. Mol. Cell 2004, 14, 787–799. [Google Scholar] [CrossRef]
- Geng, Y.; Zhou, Y. N-terminal region is required for functions of the HAM family member. Plant Signal. Behav. 2021, 1940001. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.; Zhou, Y. HAM Gene Family and Shoot Meristem Development. Front. Plant Sci. 2021, 12, 800332. [Google Scholar] [CrossRef] [PubMed]
- Beheshti, H.; Strotbek, C.; Arif, M.A.; Klingl, A.; Top, O.; Frank, W. PpGRAS12 acts as a positive regulator of meristem formation in Physcomitrium patens. Plant Mol. Biol. 2021, 107, 293–305. [Google Scholar] [CrossRef]
- Lin, P.-C.; Lu, C.-W.; Shen, B.-N.; Lee, G.-Z.; Bowman, J.L.; Arteaga-Vazquez, M.A.; Liu, L.-Y.D.; Hong, S.-F.; Lo, C.-F.; Su, G.-M. Identification of miRNAs and their targets in the liverwort Marchantia polymorpha by integrating RNA-Seq and degradome analyses. Plant Cell Physiol. 2016, 57, 339–358. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.S.; Bowman, J.L. Micro RNA s in Marchantia polymorpha. New Phytol. 2018, 220, 409–416. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Leng, X.; Sun, X.; Mu, Q.; Wang, B.; Li, X.; Wang, C.; Fang, J. Discovery of conservation and diversification of miR171 genes by phylogenetic analysis based on global genomes. Plant Genome 2015, 8. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Pan, X.; Cannon, C.H.; Cobb, G.P.; Anderson, T.A. Conservation and divergence of plant microRNA genes. Plant J. 2006, 46, 243–259. [Google Scholar] [CrossRef]
- Nozawa, M.; Miura, S.; Nei, M. Origins and evolution of microRNA genes in plant species. Genome Biol. Evol. 2012, 4, 230–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veena, G.; Muralidhara, B.; Ahmad, I.; Rajan, S. Phylogenetic Analysis Reveals the Conservation of micro RNA 171 genes in diverse Fruit Species. Vegetos 2016, 29, 13. [Google Scholar] [CrossRef]
- Branscheid, A.; Devers, E.A.; May, P.; Krajinski, F. Distribution pattern of small RNA and degradome reads provides information on miRNA gene structure and regulation. Plant Signal. Behav. 2011, 6, 1609–1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plackett, A.R.; Di Stilio, V.S.; Langdale, J.A. Ferns: The missing link in shoot evolution and development. Front. Plant Sci. 2015, 6, 972. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Yan, A.; McAdam, S.A.M.; Banks, J.A.; Zhang, S.; Zhou, Y. Timing of meristem initiation and maintenance determines the morphology of fern gametophytes. J. Exp. Bot. 2021, 72, 6990–7001. [Google Scholar] [CrossRef]
Figure 1.
The sequence of miR171a is complementary to the transcripts of Arabidopsis HAM1 HAM2 and HAM3 genes, which belong to the Type II group in the HAM family.
Figure 1.
The sequence of miR171a is complementary to the transcripts of Arabidopsis HAM1 HAM2 and HAM3 genes, which belong to the Type II group in the HAM family.

Figure 2.
miR171 functions as a regulatory hub that participates in diverse developmental processes. Created with BioRender.com.
Figure 2.
miR171 functions as a regulatory hub that participates in diverse developmental processes. Created with BioRender.com.

Figure 3.
Diagrams illustrating the expression patterns of the L1-specific transcription factors (ATML1 and PDF2) and MIR171 transcriptional reporters (left), the miR171-insensitive HAM2 transcriptional reporter (middle), and the mature miR171 and the miR171-sensitive HAM2 translational reporter (right) in Arabidopsis SAMs. The Arabidopsis SAM consists of three clonally distinct cell layers, including the L1, L2, and L3/corpus, which are indicated here. L1 represents the epidermal layer, L2 represents the subepidermal layer, and L3/corpus includes all the cells beneath the subepidermal layer.
Figure 3.
Diagrams illustrating the expression patterns of the L1-specific transcription factors (ATML1 and PDF2) and MIR171 transcriptional reporters (left), the miR171-insensitive HAM2 transcriptional reporter (middle), and the mature miR171 and the miR171-sensitive HAM2 translational reporter (right) in Arabidopsis SAMs. The Arabidopsis SAM consists of three clonally distinct cell layers, including the L1, L2, and L3/corpus, which are indicated here. L1 represents the epidermal layer, L2 represents the subepidermal layer, and L3/corpus includes all the cells beneath the subepidermal layer.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Han, H.; Zhou, Y. Function and Regulation of microRNA171 in Plant Stem Cell Homeostasis and Developmental Programing. Int. J. Mol. Sci. 2022, 23, 2544. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052544
AMA Style
Han H, Zhou Y. Function and Regulation of microRNA171 in Plant Stem Cell Homeostasis and Developmental Programing. International Journal of Molecular Sciences. 2022; 23(5):2544. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052544
Chicago/Turabian StyleHan, Han, and Yun Zhou. 2022. "Function and Regulation of microRNA171 in Plant Stem Cell Homeostasis and Developmental Programing" International Journal of Molecular Sciences 23, no. 5: 2544. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052544
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.