
Recent Advances in the Production of Genome-Edited Rats


1
Department of Genome Medicine, National Center for Child Health and Development, Tokyo 157-8535, Japan
2
Division of Biomedical Engineering, National Defense Medical College Research Institute, Saitama 359-8513, Japan
3
Department of Pediatric Dentistry, Graduate School of Medical and Dental Sciences, Kagoshima University, Kagoshima 890-8544, Japan
4
Laboratory Animal Facilities & Services, Preeminent Medical Photonics Education & Research Center, Hamamatsu University School of Medicine, Hamamatsu, Shizuoka 431-3192, Japan
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2022, 23(5), 2548; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052548
Submission received: 31 January 2022
/
Revised: 17 February 2022
/
Accepted: 21 February 2022
/
Published: 25 February 2022
(This article belongs to the Special Issue Gene Editing and Delivery in Animal Genetic Engineering)
Abstract
:The rat is an important animal model for understanding gene function and developing human disease models. Knocking out a gene function in rats was difficult until recently, when a series of genome editing (GE) technologies, including zinc-finger nucleases (ZFNs), transcription activator-like effector nucleases (TALENs), and the type II bacterial clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated Cas9 (CRISPR/Cas9) systems were successfully applied for gene modification (as exemplified by gene-specific knockout and knock-in) in the endogenous target genes of various organisms including rats. Owing to its simple application for gene modification and its ease of use, the CRISPR/Cas9 system is now commonly used worldwide. The most important aspect of this process is the selection of the method used to deliver GE components to rat embryos. In earlier stages, the microinjection (MI) of GE components into the cytoplasm and/or nuclei of a zygote was frequently employed. However, this method is associated with the use of an expensive manipulator system, the skills required to operate it, and the egg transfer (ET) of MI-treated embryos to recipient females for further development. In vitro electroporation (EP) of zygotes is next recognized as a simple and rapid method to introduce GE components to produce GE animals. Furthermore, in vitro transduction of rat embryos with adeno-associated viruses is potentially effective for obtaining GE rats. However, these two approaches also require ET. The use of gene-engineered embryonic stem cells or spermatogonial stem cells appears to be of interest to obtain GE rats; however, the procedure itself is difficult and laborious. Genome-editing via oviductal nucleic acids delivery (GONAD) (or improved GONAD (i-GONAD)) is a novel method allowing for the in situ production of GE zygotes existing within the oviductal lumen. This can be performed by the simple intraoviductal injection of GE components and subsequent in vivo EP toward the injected oviducts and does not require ET. In this review, we describe the development of various approaches for producing GE rats together with an assessment of their technical advantages and limitations, and present new GE-related technologies and current achievements using those rats in relation to human diseases.
1. Introduction
Rats (Rattus norvegicus) and mice (Mus musculus) belong to the same rodent family and have been the most widely used models in biomedical research for many years. However, there are several differences between these two animals. For example, the rat is larger (roughly about eight- to ten-fold) in size than the mouse, which provides a number of practical advantages, as exemplified by easier and more rapid microsurgery, multiple sampling of larger blood and tissue volumes, and precise injection of substances into blood vessels or the brain (reviewed by Kjell and Olson [1]). Additionally, mice and rats differ in their physiology, behavior, and neurology. Therefore, rats are considered as useful animals suitable for toxicological, neurobehavioral, and cardiovascular studies compared with other currently available experimental animals (reviewed by Jacob [2]).
There are some examples for the production of genetically modified (GM) rats reported approximately 20 years ago. These GM rats were transgenic (Tg) rats as a “gain of function” model to examine the role of plasma membrane calmodulin-dependent calcium ATPase isoform 4 (PMCA4) cDNA under control of the cardiac-specific promoter [3] or sarcoplasmic reticulum Ca2+-ATPase (SERCA2a) under control of the ubiquitous promoter [4]. Unfortunately, the production of GM rats, as exemplified by knockout (KO) or knock-in (KI) rats, has not been possible until about 10 years ago. The main reason for this is the inability to establish and maintain rat embryonic stem (ES) cells, which is much more complicated when compared with establishing and maintaining mouse ES cells. For example, in mice, ES cells were already available in the late 1980s [5,6], whereas rat ES cell production began in the late 2000s [7,8]. Since the establishment of rat ES cells, ES cell-mediated gene targeting was performed in rats for the production of KO [9,10,11,12] and KI rats [13,14,15]. For cases in which ES cells are still absent, several alternative approaches, such as transposon-mediated mutagenesis and chemical mutagenesis using N-ethyl-N-nitrosourea, have been developed to assess gene function (reviewed by Zan et al. [16]).
During the period of 2009 to 2013, genome editing (GE) technologies, such as zinc-finger nucleases (ZFNs), transcription activator-like effector nucleases (TALENs), and clustered regularly interspaced short palindromic repeat-associated protein 9 (CRISPR/Cas9) nucleases systems, all of which can induce a double-stranded break (DSB) at a specific site in the genome, appeared; since then, the production of GM rats was accelerated. Using these GE systems, many GM rats have been produced using the direct microinjection (MI) of GE components (including engineered endonucleases) into the cytoplasm or nucleus of a zygote (fertilized one-cell embryo), in vitro electroporation (EP) of isolated zygotes in the presence of GE components, and in vivo EP after the instillation of GE components into the oviductal lumen of a pregnant female (corresponding to zygote to the 2-cell stage), which is termed “genome-editing via oviductal nucleic acids delivery (GONAD) (or improved GONAD (i-GONAD))”. Furthermore, it has become possible to engineer the genome of cultured spermatogonial stem cells (SSCs), precursor cells for spermatogenesis. The transplantation of these GM SSCs into the recipient male rat testes and subsequent mating of these transplanted males with wild-type (WT) females led to the production of GM rats.
In this review, we first describe the delivery method of GE components into rat zygotes for producing GM rats. Second, we describe the novel techniques that may be potentially useful for efficient production of GM rats with multiple functions. Third, we highlight that those GE technologies are useful for producing many types of rat models for human genetic diseases.
2. What Is GE Technology?
As mentioned above, site-specific engineered nucleases are used in GE techniques. ZFN, TALEN, and CRISPR/Cas9 can all bind DNA and induce DSBs, which are subsequently repaired by non-homologous end joining (NHEJ) mechanisms or by homology-directed repair (HDR). In the case when the template DNA is absent, the DSB is repaired via the NHEJ pathway where insertion or deletion of nucleotides (hereafter called “indels”) frequently occurs in the cleaved area (reviewed by Harrison et al. [17] and Hsu et al. [18]). The indels often generate frameshift mutations of the amino acid sequence (thus leading to the generation of abnormal proteins) or the formation of a premature stop codon (thus leading to stoppage of protein synthesis). In contrast, in the presence of the template DNA (also called donor DNA) that is homologous to the target site, the template DNA is inserted into the cleaved area through HDR. Notably, HDR-mediated editing preferentially occurs in dividing cells. In contrast, NHEJ-mediated editing occurs in cells independent of the cell cycle [19].
ZFN consists of two components: the DNA-binding domain of ZF protein (capable of binding to the target DNA) and the DNA cleavage domain of Fok I endonuclease (capable of cleaving the target DNA). ZF protein contains three to six Cys2-His2 fingers, which recognize a triplet nucleotide code. After recognizing the binding site, it becomes a dimetric form, activates Fok I, and then cleaves DNA [20,21].
Similar to ZFN, TALEN comprises two components: transcriptional activator-like effector (TALE) protein and Fok I nuclease (termed TALE nucleases) [22,23,24]. TALE contains a 33–35 amino acid sequence that flanks a central DNA-binding region (amino acids 12 and 13). Notably, the design and engineering of TALENs are simpler than that of ZFN and, thus, TALENs can be engineered faster.
CRISPR/Cas9 contains two components: single-guide RNA (sgRNA) and the Cas9 protein. sgRNA comprises two components, a target-specific CRISPR RNA (crRNA) and a trans-activating crRNA (tracrRNA). These two parts can be mixed as a crRNA/tracrRNA duplex and work to guide the Cas9 protein to bind to a specific chromosomal DNA site [25,26,27]. After binding to the specific site, Cas9 cleaves one of the three bases upstream of the protospacer adjacent motif (PAM) of the DNA strand to induce a DSB. This system is different from the other GE tools such as ZFN and TALEN, and the synthesis of sgRNA is a prerequisite for this system. This development dramatically reduces both the complexity and time required for the design and implementation of gene editing.
In Table 1, a summary of GE rats produced from 2009 to 2021 is provided.
3. Delivery Method
For the production of GE rats, the choice of the delivery method for GE components in rat zygotes is important. The methods for the production of GE rats achieved by delivering GE reagents can be largely divided into six groups: the first is MI of GE reagents (in the form of DNA, mRNA, or protein) into the cytoplasm or nucleus of a zygote (Figure 1A); the second is in vitro EP of zygotes in the presence of GE reagents (Figure 1B); the third is in vivo EP of zygotes after instillation of GE reagents into the oviductal lumen of a pregnant female (Figure 1C); the fourth is in vitro viral infection of zygotes with recombinant adeno-associated virus (rAAV) (Figure 1D); the fifth is SSC-mediated production of GE rats (Figure 1E); the sixth is ES cell-mediated production of GE rats (Figure 1F).
In the following sections, each of these methods is described in detail.
3.1. MI-Based Production of GE Rats
Since the first development of pronuclear MI in mice by Gordon et al. [154], numerous GM mice and other animals such as rats, pigs, cows, and nonhuman primates have been created (reviewed by Clark et al. [155]). These animals are mainly produced by the introduction of exogenous DNA called “transgene”, which comprises a tissue-specific or ubiquitous promoter, intron, cDNA as a gene of interest (GOI), and polyadenine tails (poly(A) site), into a male pronucleus of a zygote. The mode of transgene integration into the host chromosomes is generally random, leading to frequent transgene silencing. In contrast with this transgenic (Tg) system, GE-based modulation of endogenous target genes does not always require the chromosomal integration of the GE components.
In 2009, ZFN-based GE in rats was first reported by Guerts et al. [28], who performed a single pronuclear injection (PI) of plasmid DNA (ranging from 0.4 to 10 ng/µL) or mRNA (ranging from 0.4 to 10 ng/µL) or intracytoplasmic injection (IC) of mRNA (from 2 to 10 ng/µL) encoding ZFNs in rat zygotes (from Dahl salt-sensitive (SS), Sprague–Dawley (SD), or Fawn-hooded hypertensive (FHH) strains) to induce indels in different target genes (coding for integrated reporter green fluorescent protein (GFP), immunoglobulin M (IgM), or Ras-related protein Rab-38 (Rab38)). The PN of mRNA, PN of plasmid DNA, or IC of mRNA resulted in production of pups with the GE efficiencies of 5–29%, 6–11%, or 6–75%, respectively. These GE efficiencies shown in this first study accelerated the researchers to generate more numbers of GM rats with these technologies. For example, rat models for X-linked severe combined immunodeficiency (X-SCID) were successfully created by PI of mRNAs encoding custom-designed ZFNs (targeted to interleukin 2 receptor gamma (Il2rg) locus) with GE efficiencies of over 20% [29].
In 2011, TALEN-based GE in rats was first reported by Tesson et al. [32], who intended to disrupt the rat IgM locus to create heritable mutations that could eliminate IgM function. Seven out of seventy-four (10%) rat zygotes injected with plasmid DNA (0.4 to 10 ng/µL) and 51/88 (58%) of rat embryos injected with mRNA (4 and 10 ng/µL) were modified at IgM. Of the IgM-mutated mRNA-injected rats, the frequency of biallelically modified rats was dose-dependent: 8/15 (53%) of rats injected with 10 ng/µL mRNA, 5/27 (19%) at 4 ng/µL, and none at 0.8 ng/µL.
In 2011, the 2nd wave of production of ZFN-based GE was brought by Cui et al. [33], who developed a conditional KI system in rats. They used ZFN technology in embryos to introduce sequence-specific modifications (KI) through homologous recombination (HR) in SD and Long–Evans (LE) rats. Briefly, Cui et al. [33] first constructed donors with an eight-base pair (bp) Not I restriction site inserted between the ZFN-binding sites, flanked by ~800 bp of immediate homology on each side. Donor plasmid DNA (1 ng/µL) and respective ZFN mRNA (2.5 ng/µL) were co-injected into the pronucleus of rat zygotes. The loci targeted were rat multidrug resistance protein 1a (Mdr1a) and pregnane × receptor (Pxr) loci. When ZFN-treated fetuses were examined, 1 of 15 (7%) and 1 of 8 (13%) were GM fetuses with KI at the Mdr1a or Pxr loci, respectively. Notably, successful germ-line transmission of the indel mutations that occurred in the born founder rats was also confirmed. Next, they constructed GFP donors, replacing the Not I site with a 1.5-kilobase (kb) human phosphoglycerate kinase (PGK) promoter-driven GFP cassette. GFP is in the opposite orientation of transcription for the rat Mdr1a and Pxr loci. In the case of MI with GFP cassette into the Mdr1a locus of SD rats, of the 439 embryos transferred to recipients, 83 pups were born, and 2 (2%) were pups with successful KI and 21 (25%) were those having NHEJ-based mutations, suggesting that NHEJ occurs at a higher rate than targeted integration. In the eyes of founder Mdr1a pup No. 3, PGK-GFP was expressed and visually detectable.
In contrast, Brown et al. [36] first provided a system enabling the conditional ablation of a target gene by the ZFN/Cre system through MI of ZFNs into rat zygotes. Briefly, they first generated GM rats with loxP-flanked (floxed) alleles at the glutamate ionotropic receptor N-methyl-D-aspartate (NMDA) type subunit 1 (Grin1) or corticotropin releasing hormone receptor 1 (Crhr1) locus by co-injecting mRNA encoding two pairs of ZFNs for each gene and two donors (plasmids carrying the fragment in which loxP site is surrounded by left homology arm (800 bp) and right homology arm (800 bp) in both sides) for each target into the pronucleus of zygotes. Each ZFN pair cleaved in an intron flanking the exons of interest and delivered one loxP site. In case of Grin1 targeting, among 80 live births, 6 (8%) were identified as those containing loxP in both insertion sites. Additionally, 48 animals had single loxP insertions, and another 17 had only NHEJ events. Brown et al. [36] also generated a GM rat expressing Cre under the control of an endogenous tyrosine hydroxylase (Th) promoter by inserting an internal ribosomal entry site (IRES)-Cre cassette immediately after the translational stop site of the Th open reading frame by pronuclear MI of a pair of ZFNs along with a donor plasmid. Of the 41 live-born pups, 9 (22%) were founders. Through crossing between these two lines (the floxed line and the Cre-expressing line), Cre-dependent and tissue-specific gene disruption successfully occurred in vivo in the bigenic offspring.
In 2013, the production of GE rats using the CRISPR/Cas9 system was reported [37,38]. This CRISPR/Cas9 system enables multigene KO in one shot of gene delivery to mice [156,157]. This property is especially beneficial for the purpose of creating disease model animals, as certain types of diseases are known to be caused by multigene defects. Li et al. [37] first attempted to generate KO rats with multiple mutations (at least in two different genes) using a CRISPR/Cas9 system. They injected two sgRNAs (12.5 ng/µL for each) (targeting rat melanocortin 3 receptor (Mc3r) and melanocortin 4 receptor (Mc4r)) and Cas9 mRNA (25 ng/µL) into rat zygotes. The efficiency of indel formation caused by the Cas9 nuclease was considerably different because 13 of 15 (87%) F0 pups exhibited the KO phenotype for Mc4r, while 1 of 15 (7%) exhibited the KO phenotype for Mc3r. The authors concluded that a single injection can induce disruption of at least two different genes in the rat and also suggested that targeting efficiency depends on the choice of suitable gRNA. Furthermore, Li et al. [37] first demonstrated successful germ-line transmission of the KO phenotype obtained through the CRISPR/Cas9 system, although germ-line transmission has already been known using ZFNs and TALENs by others [28,32,33]. Similar to Li et al. [37], Li et al. [38] attempted to generate multiple gene mutations in rats in a germ-line competent manner using a CRISPR/Cas9 system. They designed six sgRNAs targeting six different genomic sites encoding rat Tet methylcytosine dioxygenase 1 (Tet1) (sgTet1-1 and sgTet1-2), Tet methylcytosine dioxygenase 2 (Tet2) (sgTet2-1 and sgTet2-2), and Tet methylcytosine dioxygenase 3 (Tet3) (sgTet3-1 and sgTet3-2) to increase the likelihood of successful targeting. Co-injection of sgRNAs for the three Tet genes (sgTet1-1, sgTet2-1, and sgTet3-1) together with Cas9 mRNA into rat zygotes resulted in the generation of a total of 22 newborn pups. Of these pups, 13 (59%) contained mutations of all three Tet genes. Notably, ~60% of them had a biallelic or monoallelic mutation for each targeted locus. Li et al. [38] also demonstrated that mutations in the founder rats were transmitted to the next generation.
Ménoret et al. [35] demonstrated, for the first time, that meganucleases—naturally occurring restriction enzymes derived from lower-order animals and plants—can be used as a fourth editing platform. They microinjected a plasmid encoding an engineered meganuclease for recombination, activating gene 1 (Rag1) into the pronuclei of rat zygotes. Of the microinjected zygotes, 0.6% were found to be mutated rats. However, this technique exhibits design complexity and associated costs, making it accessible to only a few laboratories.
Although MI of GE reagents into zygotes is one of the most useful tools for generating GM animals, it has advantages as well as disadvantages. The advantages of this technology include the delivery of known quantities of nucleic acids into a zygote irrespective of the type of zygote and the introduction of a large-size cargo (carrying a GOI), which is a significant limiting factor when using viral vectors for gene delivery, as suggested by Sato et al. [158]. The disadvantages of this technology include the requirement of a micromanipulator system and egg transfer (ET) to pseudo-pregnant females for allowing the further development of MI-treated embryos.
3.2. In Vitro EP-Based Production of GE Rats
EP is known to be a useful and powerful gene delivery tool which can enable the transfer of exogenous substances (i.e., DNA, mRNA, and protein) into a cell by forming transient pores into the cell membranes under electrical stimulation in vitro and in vivo (reviewed by Young and Dean [162]).
In 2014, Kaneko et al. [43] first demonstrated that rat zygotes can be genome-edited by in vitro EP, which is shown in Figure 1B, schematically. Since then, many researchers have successfully induced gene edits using this technology in mice [163,164,165,166,167,168], rats [95,111], bovines [169], and pigs [170,171,172]. The merit of this technology is that it is simple, rapid, and convenient for GE in zygotes, compared to the previous MI-based technique. Notably, approximately 30 to 50 zygotes can be edited with one shot of in vitro EP. Furthermore, EP only requires a square pulse generator, called an electroporator, and not a more expensive micromanipulator system. According to Kaneko et al. [43], intact F344/Stm rat zygotes were subjected to in vitro EP using an NEPA21 electroporator (NEPA GENE Co. Ltd. Chiba, Japan) in the presence of a phosphate-buffered saline (PBS)-based solution containing ZFN (40 μg/mL) mRNA (targeted to the rat Il2rg gene) under the EP condition of a poring pulse (Pp) (voltage: 225 V; pulse interval: 50 ms; pulse width: 1.5 and 2.5 ms; number of pulses: 4). The NEPA21 electroporator employs a 3-step pulse system. In the first step, the Pp makes micro-holes in the ZP and oolemma. In the second step, several first transfer pulses (Tp) transfer mRNA into the cytoplasm. In the third step, the polarity-changed second Tp increase the chance of transferring mRNA into embryos. When rat embryos were electroporated with a pulse width adjusted to 1.5 or 2.5 ms, 73 and 75% of the resulting offspring, respectively, had an edited Il2rg locus. Notably, they reported no appreciable reduction in the developmental ability of the EP-treated embryos. Furthermore, the germ-line transmission of the ZFN EP-induced editing was also confirmed in the next generation. They named this technology “Technique for Animal Knockout system by Electroporation (TAKE)”. Since then, many GE rats have been produced using EP-based GE technology [54,90,91,95,111,117,121,125,131,140,141,144]. Notably, in vitro EP can also be applied to frozen mouse and rat zygotes for the efficient introduction of CRISPR components, without affecting the embryo viability or development [173,174]. This will facilitate GE rat production in a more convenient manner.
At the initial stages of GE rat production, mRNAs encoding ZFNs, TALENs, or Cas9 have been frequently used [28,29,30,31,32,33,34,36,37,38,39,40,41,42]. However, mRNA transfection still relies on the translation of the mRNA into protein, resulting in variability of GE efficiency with timing. In the case of CRISPR/Cas9-based GE, instead of using Cas9 mRNA, the use of the Cas9 protein allows transient and faster editing [175]. For example, the recombinant Cas9 protein and the chemically synthesized guide RNA (gRNA) can be combined on the bench before being delivered into embryos; this Cas9-gRNA complex, which is called a ribonucleoprotein (RNP), is active immediately. If the RNP enters the nuclei of cells, GE should commence quickly, which will, in turn, lead to increased GE efficiency. Using this RNP-based GE system, a number of GE rats have been produced to date [57,58,68,90,91,95,111,113,115,117,121,126,131,142,150].
Although in vitro EP enables the effective delivery of a short (<1 kb) single-stranded DNA (ssDNA) or synthetic single-stranded oligodeoxynucleotide (ssODN) into mouse zygotes, it has been difficult to introduce longer (>1 kb) double-stranded DNA (dsDNA) such as plasmid DNA. Only a few laboratories have successfully achieved this, although extensive exploration of optimal EP conditions, which is very laborious and time-consuming, is strictly required [176]. Notably, Bagheri et al. [177] overcame this limitation by first injecting all GE components, including long plasmid-sized DNA templates, into the sub-zona pellucida (ZP) space, called the “peri-vitelline space”. Therefore, they were retained, supporting subsequent in vitro EP. Bagheri et al. [177] suggest that this simple and well-tolerated method achieves intracellular reagent concentrations sufficient to provide precise gene edits.
As for the HDR-based KI by in vitro EP, Miyasaka et al. [95] first designed long single-stranded DNA (lssDNA) (constructed using a simple method including custom-made plasmids using nicking endonucleases as previously reported by Yoshimi et al. [64]) as the HDR template. The lssDNA comprises a targeted exon (exon 2 of the rat vesicle-associated membrane protein-associated protein B/C (Vapb) gene with a P56S mutation, which is associated with amyotrophic lateral sclerosis (ALS) in humans) flanked by two loxP sites. In vitro EP of rat zygotes was performed in the presence of Cas9 mRNA (400 ng/μL), lssDNA (40 ng/μL), and two gRNAs (200 ng/μL) in a chamber with 40 μL of serum-free media (Opti-MEM) with a 5-mm gap electrode in a NEPA21 electroporator under the following conditions: Pp of voltage 225 V, pulse width—2.0 ms, pulse interval—50 ms, and number of pulses—4. The first and second Tp were of voltage 20 V, pulse width—50 ms, pulse interval 50—ms, and number of pulses—5. Consequently, of six pups born, three showed floxed alleles with the P56S mutation. They called this technology “CRISPR with lssDNA inducing conditional knockout alleles (CLICK)”. The CLICK technology will be useful as a tool for producing GM zygotes carrying transgenes driving tissue-specific Cre recombinase, which, in turn, will be applicable to the one-step generation of conditional KO animals.
When compared to the MI-based technique, in vitro EP allows the simultaneous processing of many zygotes in a short time (e.g., a batch of 30 to 50 zygotes in few seconds) without requiring expensive equipment and operators with extensive training and expertise. The disadvantages of this technology include the requirement of ET of the EP-treated embryos to pseudo-pregnant females to allow further development of the treated embryos.
3.3. GONAD-Based Production of GE Rats
In 2015, a novel method (termed “GONAD”) enabling in situ CRISPR/Cas9-based GE toward early embryos was first reported by Takahashi et al. [180] using mice. In this system, both Cas9 mRNA (up to 1.1 μg/μL) and sgRNA (0.6 μg/μL) were co-injected into the oviductal lumen of pregnant females at Day 1.4 of pregnancy (corresponding to the 2-cell stage). Technically, this was simply achieved by inserting a glass micropipette through the oviductal wall under observation using a dissecting microscope, as shown in Figure 1C schematically. Next, a small amount of a solution (1–1.5 µL) containing GE components was injected into the oviductal lumen with the aid of a mouthpiece-controlled micropipette. Subsequently, in vivo EP was performed using tweezer-type electrodes and a square-wave pulse generator (T820; BTX Genetronics Inc. San Diego, CA, USA) under the EP condition of eight square-wave pulses with a pulse duration of 5 ms and an electric field intensity of 50 V. GE occurrence in the 2-cell embryos can be later checked by molecular analysis of the mid-gestational fetuses dissected after the GONAD. The KO efficiency of these fetuses was approximately 29%. Later, this technology was elaborated by the same group, who injected a solution containing RNP (comprising Cas9 protein (1 µg/µL Integrated DNA Technologies, Inc. Coralville, IA, USA) and sgRNA (30 µM)) (for KO experiment), RNP + ssODN (1–2 µg/µL) (for KI experiment), or RNP + ssDNA (0.85–1.4 µg/µL) (for KI experiment) at Day 0.7 of pregnancy (corresponding to the late 1-cell stage) [181]. In vivo EP using the NEPA21 electroporator was performed under the following conditions: Pp: 50 V, 5-ms pulse, 50-ms pulse interval, three pulses, 10% decay (±pulse orientation); Tp: 10 V, 50 ms pulse, 50 ms pulse interval, three pulses, and 40% decay (±pulse orientation). This modification resulted in 97% of the embryos exhibiting indels in the target locus and approximately 50% of embryos containing KI alleles. Thus, this improved technology was re-named the “improved GONAD (i-GONAD)”. Since the successful development of GONAD/i-GONAD in mice, the technology has proven useful for other species, such as rats [90,91,121] and hamsters [182].
Successful i-GONAD in rats was first reported in 2018 [90,91]. For examples, Takabayashi et al. [90] demonstrated that the appearance of non-pigmented eyes in fetuses (showing the KO phenotype due to indels at the tyrosinase gene (Tyr), a gene coding for protein essential for eye pigmentation) was induced with an efficiency of 56%, when F1 embryos from pigmented Brown Norway (BN) × albino SD rat crosses were used for i-GONAD targeting of Tyr. In this case, the in vivo EP condition used was almost the same as that used for i-GONAD in mice. They also tested the possibility of KI (targeted Tyr locus) using albino Lewis (LEW) rats. The i-GONAD-mediated KI efficiency, which was evaluated by the presence of fetuses with pigmented eyes, was as low as ~5%. Kobayashi et al. [91] provided data similar to that of Takabayashi et al. [90] using the NEPA21 electroporator under the following EP conditions: Pp: 30, 40, or 50 V, 5-ms pulse, 50-ms pulse interval, number of pulses 3, and 10% decay (±pulse orientation); Tp: 10 V, 50-ms pulse, 50-ms pulse, number of pulses 6, and 40% decay (±pulse orientation). According to Kobayashi et al. [91], intraoviductal injection of RNP (1 μg/μL of Cas9 protein +30 μM of sgRNA) targeted to Tyr into oviducts of pregnant albino WKY females (which had been successfully mated with dark agouti (DA) males) or pigmented DA females (which had been successfully mated to Wistar–Kyoto (WKY) males) resulted in the production of GE pups with efficiencies of 59% or 42%, respectively, when 50 V of the voltage (Pp) was employed. These results suggest no significant difference in the gene-editing efficiency between these two strains. They also attempted to recover the coat-color mutation in WKY females using an ssODN-based KI approach. Consequently, the KI efficiency was 27% in the pups born. They named this rat-based i-GONAD as “rGONAD”.
For performing i-GONAD (or rGONAD) in rats, the exploration of optimal EP conditions is strictly required. For example, when a current of >500 mA was employed, albino SD and albino LEW rats were successfully genome-edited; however, the acquisition of offspring derived from pigmented BN rats (fetuses/newborns) failed. In contrast, i-GONAD under a current of 100–300 mA using the NEPA21 electroporator led to the production of GE BN rats with efficiencies of 75% to 100% [90]. Similarly, i-GONAD under a current of 150–200 mA using another electroporator CUY21EDIT II (BEX Co., Tokyo, Japan) led to the production of genome-edited BN rats with efficiencies of 24% to 55% [121].
The advantages of this technology are that GONAD/i-GONAD requires an electroporator but no other special equipment. Furthermore, no recipients are required unlike the case of MI- or in vitro EP-based GE rat production. Disadvantages of this technology are that technical skill may be required for intraoviductal injection of a solution into the lumen of oviducts under a dissecting microscope using a mouthpiece-controlled glass micropipette.
The detailed protocols for GONAD/i-GONAD have been reported by Gurumurthy et al. [183] and Ohtsuka and Sato [184] in mice. The GONAD/i-GONAD-based production of GE rats is also possible using the same approach shown in mice. Notably, Namba et al. [185] demonstrated the protocols for GONAD/i-GONAD in rats.
3.4. rAAV-Mediated Production of GE Rats
rAAVs are non-pathogenic and non-enveloped ssDNA viruses enabling highly efficient transduction of both dividing and non-dividing cells (reviewed by Daya and Berns [186]). Unlike other viral vectors such as retroviruses and adenoviruses, rAAVs have been used for manipulating the target genome sequences through simple co-incubation with ZP-enclosed preimplantation mammalian embryos for the production of GM animals such as mice [168,187,188,189,190,191]. For example, Mizuno et al. [187] observed that, among the rAAVs tested, the rAAV with serotype 6 (which is hereinafter referred to as rAAV-6) showed the highest transduction efficiency when ZP-intact murine embryos were co-cultured with rAAVs. More importantly, Mizuno et al. [187] demonstrated successful CRISPR-based KI in mice. As shown in Figure 1D, zygotes were first subjected to in vitro EP in the presence of RNP and then infected with rAAV-6 carrying a 1.8-kb GFP expression cassette flanked by two 100-bp Rosa26 homology arms. The KI efficiencies in the Rosa26 locus were 16% and 6%, when assessed at the blastocyst and newborn stages, respectively. Notably, Mizuno et al. [187] demonstrated that both ZP-intact rat and bovine embryos were effectively infected by rAAV-6, suggesting that rat genome can be effectively edited through the simple incubation of ZP-intact rat zygotes with a solution containing rAAVs.
This rAAV-based gene delivery system to early embryos was also observed in vivo. Yoon et al. [188] injected a solution containing two vectors, rAAV6-Cas9 (carrying spCas9 gene derived from Streptococcus pyogenes) and rAAV6-gTyr (carrying gRNA expression unit targeted Tyr), into the oviducts of pregnant female mice at Day 0.5 of pregnancy, similar to GONAD/i-GONAD. Out of the 29 pups that were obtained, three (10%) were found to have indels. All mutated founder mice generated albino offspring, indicating germ-line transmission. These results suggest that AAV is a powerful tool for inducing GE in the ZP-enclosed early embryos in vivo. According to Sato et al. [158], this in vivo approach is referred to as “AAV-based GONAD”.
The advantages of this technology are that no special equipment is required in the case of using the AAV-based GONAD. Notably, the use of highly concentrated rAAV would be one of the key factors for achieving a high rate of transduction of preimplantation embryos. According to Yoon et al. [188], zygotes treated with 6 × 108 genome copies (GCs) for one day in vitro resulted in live births with 100% indel frequency after ET. However, the editing frequency dropped to 25% for fetuses and 20% for newborns at 6 × 107 GCs. No edited animals were detected from the 6 × 106 GC treatment group. These results clearly suggest that gene delivery and GE efficiency are rAAV dose-dependent.
3.5. SSC-Mediated Production of GE Rats
Cultured SSCs can display long-term sperm-forming potential when they are transplanted into the seminiferous tubules (STs) of sterile rat testes (shown in Figure 1E). It is now possible to culture SSCs for a long time and perform gene modification toward SSCs derived from mice and rats (reviewed by Sato et al. [158]).
Chapman et al. [60] first demonstrated that CRISPR/Cas9-mediated GE can be successfully applied in SSCs to create KO rats. They used SSCs isolated from tgGCS-EGFP rats (with the genetic background of SD) showing germ cell-specific expression of enhanced green fluorescent protein (EGFP) [192]. The SSCs were then transfected with the pX330 vector (conferring expression of both Cas9 and gRNA targeted epithelial stromal interaction 1 (Epsti1) gene) together with a neomycin resistance gene expression plasmid. The surviving clones after G418 selection were then transplanted into the STs of recipient sterile rats to promote spermatogenesis. When the recipient rats transplanted with G418-selected cells were mated with WT females at ~65 days post-transplantation, a total of 87 pups were obtained. Of these, ~10% (9) were found to harbor Epsti1 mutants. A similar attempt was also made by Noto et al. [102], who successfully obtained immunodeficient rats by grafting recombination activating gene 2 (Rag2) KO SSCs into rat testes.
3.6. ES Cell-Mediated Production of GE Rats
As mentioned previously, acquisition of a rat ES cell line is now possible. Yamamoto et al. [14] first succeeded in producing KI rats using CRISPR/Cas9-engineered ES cells. Initially, they obtained GM ES cell clones using a conventional gene-targeting technology but failed because of no recombinant ES cells were obtained. Next, in vitro EP of a Cas9-expressing plasmid, gRNA-expressing plasmid, and KI vector (carrying two homology arms) in rat ES cells was performed. Consequently, the number of drug-resistant colonies increased and the HR efficiency was considerably enhanced from 0 to 36%. These recombinant clones resulted in the successful production of chimeras, which were later proven to have the ability to transmit the KI allele to their offspring after mating with WT rats (as shown in Figure 1F). According to Yamamoto et al. [14], the methodology presented can overcome the problem encountered in conventional gene targeting that prevented the production of humanized rats. Later, successful GM rat production through ES cells carrying CRISPR/Cas9-based KI of the Discosoma coral red fluorescent protein (dsRed) gene at the Sry-box transcription factor 10 (Sox10) locus was reported by Chen et al. [15].
4. Other Techniques and Factors to Modify the Rat Genome
As shown above, GE tools such as ZFNs, TALENs, and CRISPR/Cas9 are considered useful for the production of GE rats. However, several techniques and factors still influence their performance, which must be addressed. These include the allele-specific GE using ssODNs, large genomic fragment deletion, ssODN-mediated KI without the need to construct homology arms, the use of inhibitors for efficient KI, the importance of choosing the appropriate gRNA for efficient KI, and base editor systems enabling precise genome modifications.
In this section, these techniques or factors are described in detail.
4.1. Allele-Specific GE for the Correction of Mutated Phenotypes
Numerous reports have demonstrated that CRISPR/Cas9 editors combined with ~100 bp ssODNs, instead of using a relatively long template (~1 kb), allow for precise HDR-mediated GE in mice [156,157,193]. These results suggest that ssODNs co-injected as donor templates preferentially support the activation of HDR relative to the NHEJ pathway.
To explore the applications of ssODN-mediated GE in rats, Yoshimi et al. [46] successfully used ssODNs to manipulate the coat-color genetics of F344 rats. They used a WT ssODN to exchange a single base pair in Tyr, thus reverting the albino allele back to the WT sequence and restoring a non-albino pigmentation pattern. A similar strategy using ssODN for the correction of mutated phenotypes in rats was also employed by others [64,90,91,106,114,125,126,127,129,141,142].
4.2. Large Genomic Fragment Deletion
The GE system has enabled the creation of targeted deletion spanning the large genomic region over 10 kb in size via the direct MI of two sgRNAs (~10–30 ng/μL) and TALEN mRNA (45 ng/μL) or Cas9 mRNA (~67–100 ng/µL) into the pronuclei or cytoplasm of mouse zygotes [194,195]. Wang et al. [58] achieved 53-kb deletion of a fragment spanning long non-coding RNA (lncRNA) GM14005 genomic DNA by injecting the Cas9 protein (30 nM) into the cytoplasm of mouse zygotes together with two sgRNAs (12.5 ng/µL). Among five F0 pups, one founder was confirmed as a mouse with a large deletion. Wang et al. [58] also showed that the 53-kb deletion was efficiently transmitted to the F1 generation.
Yoshimi et al. [46] succeeded in the deletion of a 7-kb fragment from the rat genome. They used two flanking gRNAs to delete a 7098-bp endogenous retroviral element (ERV) within the first intron of the Kit gene, using ssODN containing homology arms on either side of the junction to bridge across the deletion (Figure 2). They injected 100 ng/μL Cas9 mRNA, 50 ng/μL gRNAs (targeted Kit), and 120 bp ssODNs (50 ng/μL) into the pronuclei of Long–Evans hooded (LEH) rat zygotes. Thus, GE rats exhibited restoration of the function of the Kit gene with a non-hooded pigmentation pattern. The frequency of the appropriately edited pups was 4%, showing the feasibility of performing template-directed correction of mutated portions using ssODNs.
If both DSBs occur at the same time, the result will be a large deletion of the region of interest. To the best of our knowledge, the biggest deletion achieved to date in rats is 24.4 mega base pairs (Mb) [196].
4.3. ssODN-Mediated KI with CRISPR/Cas9 for Large Genomic Regions in Rat Zygotes
Although the CRISPR/Cas9 system is recognized as a powerful tool for generating GM animals, the creation of targeted KI animals via HR is still challenging. Yoshimi et al. [64] developed a novel KI method without the need to construct homology arms, thus simplifying the process of genome engineering in living organisms. In other words, they tried to knock-in longer plasmids (including the 1.2-kb chicken β-actin-based promoter (called “CAG”) and a 0.8-kb GFP cassette sequence) at the targeted site (rat Rosa26 locus) (Figure 3). In more detail, they constructed two gRNAs: one targeting the rat Rosa26 locus to cleave genomic DNA, and the other targeting the 5′ end of the CAG promoter sequence for concurrent cleavage of the plasmid DNA (Figure 3). Two 80-bp ssODNs (ssODN-1 and -2) were designed to ligate the two cut ends, as shown in (Figure 3). A mixture of 100 ng/μL of the Cas9-poly(A) mRNA (which appended an 81-bp poly(A) to the transcription terminator to increase the efficacy of GE with the CRISPR/Cas9 system), 50 ng/μL of each of the two gRNAs (gRNA:rRosa26 and gRNA:CAGGS), 50 ng/μL of each of the two ssODNs (ssODN-1 and -2), and 5 ng/μL of the CAG-GFP plasmid was co-microinjected into the pronuclei of rat zygotes. Of the 17 pups obtained, four (#6, #7, #8, and #11) were selected as candidates with successful KI. Sequence analysis revealed that #11 had accurate conjunction at both of the two cut ends, #6 had a 6-bp deletion on one side, and #7 had several base-pair deletions and insertions on both sides. They confirmed germ-line transmission of the GFP allele. No off-target effect was observed at any candidate sites in these rats. Furthermore, uniform expression of the GFP gene in most cells and tissues examined was confirmed. Based on these observations, this strategy was called “two-hit by gRNA and two-oligo with plasmid (2H2OP)”.
4.4. Use of Inhibitor for Efficient KI
KI is an event of precise modifications at a target locus and is often mediated by HR in the presence of donor DNA, as exemplified by ssODN. Generally, HDR-mediated KI is more difficult than NHEJ-based indels. For example, in proliferating human cells, NHEJ has been reported to repair 75% DSBs, whereas HDR repaired the remaining 25% [197]. To enhance the HDR efficiency, several approaches are now being attempted. In the case of a CRISPR/Cas9-mediated KI event, Scr7, which was first identified as an anti-cancer compound and was considered a potential NHEJ inhibitor, was used to increase the HR-mediated precise genetic modification [198,199]. For example, co-injection of murine zygotes with a mixture containing Cas9 mRNA, sgRNA, template ssODNs, and Scr7 significantly improved the efficiency of HDR-mediated insertional mutagenesis [198]. Chu et al. [200] also demonstrated the usefulness of Scr7 for abolishing NHEJ activity and increasing HDR in both human and mouse cell lines. However, the function of Scr7 in promoting HDR remains controversial. Some researchers have demonstrated that Scr7 failed to increase HDR rates in rabbit embryos [201] and porcine fetal fibroblasts [202].
In rats, Ma et al. [68] first demonstrated that both Scr7 and Cas9 protein can increase the precise modification in rat embryos when the fatty acid binding protein 2 (Fabp2) and dysbindin (dystrobrevin binding protein 1) domain containing 1 (Dbndd1) loci were selected to perform KI of a fragment containing Cre or Cre-ERT2 (Cre recombinase-estrogen receptor T2) gene, respectively. For example, when a mixture containing Cas9 mRNA (25 ng/µL), sgRNA (10 ng/µL; targeting Fabp2), plasmid template (4 ng/µL; for Fabp2), and Scr7 (0, 0.5, 1 or 2 μM) was injected into both the cytoplasm and pronuclei of rat zygotes, two (18%) of 11 pups born exhibited KI in the absence of Scr7. In contrast, three (33%) of 9 and 11 (39%) of 28 pups were found to exhibit KI after MI with 2 or 1 μM Scr7, respectively.
However, the opposite results were recently achieved by Takabayashi et al. [90] and Aoshima et al. [141]. Takabayashi et al. [90] performed i-GONAD using a solution containing 1 μM Scr7, RNP (1 μg/μL of Cas9 protein +30 µM of sgRNA), and 2 µg/µL of ssODN resulted in the production of 27 fetuses, of which only two (7%) had pigmented eyes. Notably, in the absence of Scr7, i-GONAD-mediated KI efficiency was as low as ~5%, suggesting that addition of Scr7 appeared not to affect i-GONAD-mediated KI efficiency in rats. Aoshima et al. [141] examined whether commercially available KI-enhancing drugs (including Scr7, L755,507, RS-1, and HDR enhancer), all of which are known to enhance KI efficiency, can increase the KI efficiency using i-GONAD in rats. Aoshima et al. [141] demonstrated that some drugs (e.g., Scr7, L755,507, and HDR enhancer) were found to be slightly effective in vivo, but their effects were not statistically significant when compared to those in the control group (without any KI-enhancing drugs).
4.5. Choice of gRNA Is Very Important for Achieving High KI Efficiency in Rats
According to Raveux et al. [203], ssODN-mediated KI efficiency varied from 0% to 40%, depending on the type of Cas9, the injection site, the length of the homology arm, and the gRNA-binding site. The nucleotide composition of a target sequence is one of the most important factors determining KI and KO efficiency. For example, Graf et al. [204] screened the published gRNA activity datasets and demonstrated that TT- and GCC-motifs located in the four PAM-proximal bases of the targeting sequence (the efficiency-modulating sequence) were sufficient to block CRISPR/Cas9-mediated KI. Moreover, the presence of these motifs in gRNAs resulted in a 10-fold reduction in gene KO frequencies.
Aoshima et al. [141] constructed three gRNAs showing high efficiencies, ranging from 51% to 57%, based on the CHOPCHOP predictions [205]. These three gRNAs (termed crRNA1, crRNA2, and crRNA3) used here recognize the Tyr sequence containing a point mutation (G to A) and overlap with each other (Figure 4). When KI-based i-GONAD was performed using ssODN (2 μg/μL), Cas9 protein (1 μg/μL), and gRNA (crRNA1, 2 or 3) (30 μM), the KI efficiency in fetuses generated after i-GONAD with crRNA2 was significantly higher (24%) than that obtained after i-GONAD with crRNA1 (5%) or crRNA3 (0%). The discrepancy between predicted and actual efficiencies was unexpected. Notably, no inhibitory sequences exist in the gRNAs used, unlike the suggestion of Graf et al. [204]. Furthermore, in the case of in vitro GE, the GE efficiency is known to vary, with predicted efficiencies typically higher than observed levels [156,206,207]. The data of Aoshima et al. [141] suggest the importance of selecting the appropriate gRNA for effective KI in rats.
4.6. Base Editor Systems as An Efficient Tool Enabling Precise Genome Modifications
Single-base editing systems without the induction of DSB or template DNA, mediated by cytidine deaminase combined with Cas9 nickase (nCas9) or dead Cas9 (dCas9), were recently developed to modify mammalian genomic DNA. This system is called the “cytosine base editor (CBE)” which achieves C·G to T·A conversion with a window of approximately five nucleotides [208,209]. Furthermore, Gaudelli et al. [210] developed a new base-editing (BE)-based system, called the “adenine base editor (ABE)”, which efficiently enables the induction of A·T to G·C conversion through fusion of an engineered tRNA adenosine deaminase (ecTadA) with nCas9. Thus, together with CBE, ABE enables the introduction of all four nucleotide transitions (C to T, A to G, T to C, and G to A) in the target genomic sequence [210]. Indeed, using these systems, the correction of genetic disease has been successfully performed in mice [211,212].
With respect to the production of GM rats using the BE systems, Yang et al. [97] reported the successful production of KO rats through MI of ABE mRNA together with sgRNA (targeted glycogen storage disease type 2 (Gsd2) gene) to generate an animal model for glycogen storage disease type II. Consequently, 85% (28/33) of rats had single or multiple A > G substitutions between position A3 and A7 in the target protein (GSD2), leading to I646V or D645G mutations. Similarly, Ma et al. [104] injected ABE mRNA together with the sgRNA-targeted hemogen (Hemgn), N-deacetylase (Ndst1), or N-sulfotransferase 4 (Ndst4) gene into rat zygotes. For Hemgn targeting, the zygotes were microinjected with ABE mRNA and sgRNA (targeted Hemgn); a total of 99 injected zygotes were transferred to three pseudo-pregnant female rats, and 15 pups were born. Consequently, fourteen rats (14/15) contained an A to G conversion at the 14th base distal from the PAM, indicating the high base-editing efficiency. According to Ma et al. [104], these BE tools can provide a much safer approach compared with the WT CRISPR/Cas9 system for the gene correction of human disease.
Recently, Qi et al. [153] demonstrated that a bacterial toxin deaminase (DddA) from Burkholderia cenocepacia, which can convert cytosine to uracil specifically within dsDNA, can be used for mediating mitochondrial DNA (mtDNA) editing in rats. It has been demonstrated that the toxin domain of DddA (DddAtox, 1264–1427 amino acids) is engineered and incorporated with the mitoTALE system to efficiently achieve C ∙ G-to-T ∙ A conversion in mtDNA of human cell lines [213]. This system is therefore termed “DddA-derived cytosine base editor (DdCBE)” [213]. Qi et al. [153] injected mRNAs of a DdCBE pair with the best performance in rat zygotes, but the DdCBE-mediated BE failed. They reasoned that this failure was due to the inactive mtDNA replication in zygotes because this system works during the period when mtDNA replication is active. To overcome this problem, the DdCBE pair was cloned into the piggyBack (PB) transposon vector, and then, the resulting PB vector was co-injected into rat zygotes with PB transposase mRNA. Thus, one out of four pups obtained was successfully edited with 36% efficiency for one target locus. According to Qi et al. [153], almost all DdCBE pairs failed to integrate into the rat genome by the transposon system; however, the expression of the DdCBE pair from the PB vector might have lasted longer than that from mRNA injection, facilitating C ∙ G-to-T ∙ A conversion in mtDNA. Using the DdCBE system, animals with precise mtDNA mutations can be produced on demand.
5. Disease Models in Rats
For a decade, the advent of genetic engineering tools such as ZFNs, TALENs, and CRISPR/Cas9 has led to a revolution in obtaining specific and targeted genetic mutations in rats for the study of human genetic diseases. Several important genetic diseases have been modeled in rats. A brief description of the most useful models is provided below.
5.1. Models for Cardiovascular Diseases
Monoallelic mutations in the gene encoding bone morphogenetic protein receptor 2 (Bmpr2) are the main genetic risk factors for heritable pulmonary arterial hypertension (PAH) with incomplete penetrance. Several Bmpr2 Tg mice have been reported to develop mild spontaneous PAH. Ranchoux et al. [53] generated Bmpr2 KO rats using the ZFN technology. The resulting KO rats with a heterozygous 140 bp deletion in the first exon of Bmpr2 displayed intense pulmonary vascular remodeling. The same group [116] also generated rat lines with mutations (deletion of 71 bp in exon 1 of Bmpr2) and showed that the heterozygous rats developed age-dependent spontaneous PAH with a low penetrance (16–27%), similar to that in humans. They concluded that this new genetic rat model represents a promising tool to study PAH pathogenesis. Concerning the pathogenesis of PAH, mutations in the potassium channel subfamily K member 3 (KCNK3) gene, which encodes an outward rectifier K+ channel, have been identified in PAH patients. Lambert et al. [109] generated Kcnk3 KO rats through the CRISPR/Cas9 technology. The resulting rats developed age-dependent PAH associated with low serum-albumin concentrations. Lambert et al. [109] concluded that KCNK3 loss of function is a key event in PAH pathogenesis.
5.2. Models for Neurological Diseases
Animal models are required to understand the pathogenesis of autism spectrum disorder (ASD). Despite the apparent advantages of mice for neural studies, rats have not been widely used for ASD studies, probably owing to the lack of convenient genome manipulation tools. Hamilton et al. [42] generated two rat models for ASD, one syndromic and one non-syndromic model, through the ZFN system, by destroying a gene (Fmr1) coding the Fragile X mental retardation protein (FMRP), the protein responsible for the pathogenesis of Fragile X syndrome (FXS), or a gene (Ngln3) coding for Neuroligin3 (NLGN3), a member of the neuroligin synaptic cell-adhesion protein family, respectively. Both FMRP and NLGN3 have been implicated in the pathogenesis of human ASD. Both KO rat lines exhibited abnormalities in ASD-relevant phenotypes including juvenile play, perseverative behaviors, and sensorimotor gating, suggesting the utility of these rats as genetic models for investigating ASD-relevant genes. Later, Tian et al. [77] produced Fmr1 KO rats through the CRISPR/Cas9 technology targeted to exon 4 of Fmr1. Consistent with the previous reports, deletion of the Fmr1 gene in rats specifically impairs long-term synaptic plasticity and hippocampus-dependent learning in a manner resembling the key symptoms of FXS.
DEP-domain containing 5 gene (Depdc5), encoding a repressor of the mechanistic target of rapamycin complex 1 (mTORC1) signaling pathway, has recently emerged as a major gene mutated in familial focal epilepsies and focal cortical dysplasia. Marsan et al. [67] produced a Depdc5 KO rat using the TALEN technology. Homozygous KO embryos died from Day 14.5 of pregnancy. Heterozygous KO rats developed normally and exhibited no spontaneous electroclinical seizures; however, they showed altered cortical neuron excitability and firing patterns. This rat model is considered a relevant model to study pathogenic mechanisms underlying those disorders.
One subtype of ASD is associated with mutations in the methyl-CpG-binding protein 2 (Mecp2) gene, causing an X-linked neurodevelopmental disorder called Rett syndrome (RS). Patients with RS have cognitive defects and circadian clock dysfunction, as exemplified by abnormal sleep patterns. According to Zhai et al. [61], there is an urgent need for new animal models for RS because the existing Mecp2 KO mouse models fail to fully mimic the pathogenesis and symptoms of patients with RS. Mecp2 KO rats were successfully produced by the two groups using ZFN [62] and CRISPR/Cas9 [61] technologies. The resulting KO rats exhibited significant abnormalities in growth (body weight loss) as well as behavioral function (anxiety tendency and cognitive deficits) [61,62]. Because these phenotypes well recapitulate the major symptoms of RS patients, these Mecp2 KO rats will provide an alternative tool for future studies of MeCP2 functions.
Mutations in fused in sarcoma (Fus), a nuclear DNA/RNA-binding protein, cause familial ALS and occasionally frontotemporal dementia. Zhang et al. [87] produced KI rats expressing a Fus point mutation (R521C) as a model for ALS using the CRISPR/Cas9 technology. The mutant animals developed adult-onset learning and memory behavioral deficits, with reduced spine density in the hippocampal neurons. Remarkably, the sleep–wake cycle and circadian abnormalities preceded the onset of cognitive deficits. These results suggest a new role of Fus in sleep and circadian regulation and demonstrate that functional changes in FUS could cause sleep–wake and circadian disturbances as early symptoms.
Emmert et al. [113] produced a novel rat model of X-linked hydrocephalus (XLH) by CRISPR-mediated mutation in the L1 cell-adhesion molecule (L1cam) gene on the X chromosome. Hemizygous male mutants developed hydrocephalus and delayed development. The mutant rats did not show reactive gliosis, but exhibited hypomyelination and increased extracellular fluid in the corpus callosum.
GABAergic dysfunctions have been implicated in the pathogenesis of schizophrenia, especially the associated cognitive impairments. The level of the GABA synthetic enzyme glutamate decarboxylase 67 kDa isoform (GAD67) encoded by the GAD1 gene is downregulated in the brains of schizophrenia patients. Furthermore, a schizophrenia patient harboring a homozygous mutation of GAD1 has recently been discovered. Gad1 KO mice exhibited perinatal lethality [214], which precluded characterization at adult stages. Fujihara et al. [125] generated Gad1 KO rats using the CRISPR/Cas9 technology. Surprisingly, 33% Gad1 KO rats survived to adulthood, which made further characterization possible. The Gad1 KO rats exhibited impairments in both spatial reference and working memory without affecting adult neurogenesis in the hippocampus. In addition, Gad1 KO rats showed a wide range of behavioral alterations, such as enhanced sensitivity to an NMDA receptor antagonist, hypoactivity in a novel environment, and decreased preference for social novelty. These results suggest that Gad1 KO rats could be a novel model for studying cognitive deficits. Furthermore, Fujihara et al. [125] claimed the necessity to check species differences in the mode of phenotype manifestation when animal models of human diseases are considered.
Angelman syndrome (AS) is a rare genetic disorder characterized by severe intellectual disability, seizures, lack of speech, and ataxia. The gene responsible for AS is the ubiquitin protein ligase E3A (Ube3a) gene, which encodes for ubiquitin ligase E6-associated protein (E6AP). Dodge et al. [122] generated Ube3a KO rats using the CRISPR/Cas9 system. The resulting KO rats phenotypically mirrored human AS with deficits in motor coordination as well as learning and memory. This model can, thus, offer a new avenue for the study of AS.
Koster et al. [142] produced a pigmented KO rat model for lecithin retinol acyltransferase (LRAT) using the CRISPR/Cas9 system. The introduced mutation (c.12delA) is based on a patient group harboring a homozygous frameshift mutation in the Lrat gene (c.12delC), causing a dysfunctional visual (retinoid) cycle. The resulting KO rats exhibited progressively reduced electroretinography potentials from two weeks of age onwards and overall retinal thinning. Vision-based behavioral assays confirmed the reduced vision. These KO rats are a novel animal model for retinal dystrophy, especially for early-onset retinal dystrophies.
5.3. Models for Muscular Diseases
Duchenne muscular dystrophy (DMD) is an X-linked lethal muscle disorder caused by mutations in the Duchenne muscular dystrophy (Dmd) gene encoding dystrophin. DMD model animals, such as Mdx (X-linked muscular dystrophy) mice and canine X-linked muscular dystrophy dogs, have been widely utilized in the development of a treatment for DMD. However, according to Larcher et al. [51], large animal models such as dogs are expensive and difficult to handle. In contrast, Mdx mice only partially mimic the human disease, with limited chronic muscular lesions and muscle weakness. Their small size also imposes limitations on analyses. In this context, a rat model could represent a useful alternative because rats are bigger than mice and could better reflect the lesions and functional abnormalities observed in DMD patients.
Dmd KO rats were successfully produced by two groups using CRISPR/Cas9 technology targeting two exons of the rat Dmd [50] or using TALENs targeting exon 23 [51]. These resulting KO rats exhibited a decline in muscle strength and the emergence of degenerative/regenerative phenotypes in the skeletal muscle, heart, and diaphragm. Furthermore, these phenotypes were transmitted to the next generation [50]. Notably, Dmd KO rats, but not mice, present cardiovascular alterations close to those observed in humans, which are the main cause of death of patients [51].
Desminopathy is a clinically heterogeneous muscle disease caused by over 60 different mutations in the desmin (DES) gene. The most common mutation with a clinical phenotype in humans is an exchange of arginine to proline at position 350 of desmin leading to p.R350P. Langer et al. [126] first produced a KI rat model for a muscle disease through the CRISPR/Cas9 technology using ssODN carrying the missense mutation Des c.1045-1046 (AGG > CCG) in exon 6 of Des. While muscle weights did not differ between the mutant rats and WT rats, the levels of many muscle-related proteins such as dystrophin, syntrophin, dysferlin, and annexin A2 increased in the mutant rats, showing the phenotype of desminopathy. This rat model will be a useful tool for furthering our understanding of the disease and testing therapeutic approaches to delay disease progression.
5.4. Models for Pulmonary Diseases
Cystic fibrosis (CF) is characterized by airway and digestive pathology with a reduced life expectancy and is one of the most common genetic diseases in western populations. The most common mutation is the missense mutation p.Phe508del (or F508del) in the cystic fibrosis transmembrane conductance regulator (CFTR) gene, which leads to abnormal CFTR function and mucus accumulation. Tuggle et al. [52] first generated Cftr KO rats using the ZFN technology. The resulting KO rats lacked CFTR activity and exhibited abnormalities in the ileum and increased intracellular mucus in the proximal nasal septa. Airway surface liquid and periciliary liquid depths were reduced, and the submucosal gland size was abnormal. Similar results have also been observed in another Cftr KO rat developed by Dreano et al. [106] using the CRISPR/Cas9 technology. Furthermore, Dreano et al. [106] produced another KI rats (carrying F508del mutation). These rats showed residual CFTR activity and milder phenotype than the Cftr KO rats. These findings suggest that these rat models can be a CF animal model that recapitulates various aspects of the human disease. Notably, a humanized CF rat strain expressing the G551D variant was produced through KI of the human CFTR cDNA carrying the G551D mutation downstream of the endogenous Cftr promoter using the ZFN technology [130].
5.5. Models for Metabolic Diseases
The low-density lipoprotein receptor (LDLR) and apolipoprotein E (APOE) genes control normal levels of cholesterol and other forms of fat in the blood. A deficiency in ApoE is involved in several age-related fatty acid diseases. Wei et al. [56] established ApoE KO rats through TALEN-mediated gene targeting. After being fed with a high-cholesterol diet (HCD) for 12 weeks, the ApoE KO rats displayed typical dyslipidemia, although there was no obvious atherosclerotic lesion in the en face aortas. Notably, partial ligation caused the formation of plaques consisting of lipids and macrophages in carotid arteries from ApoE KO rats. Wei et al. [56] concluded that the ApoE KO rats can be a novel model for dyslipidemia. Lee et al. [110] produced ApoE KO rats using Cas12a (previously named Cpf1), an RNA-guided endonuclease, as a part of the CRISPR system. The resulting KO rats displayed hyperlipidemia and aortic lesions.
A deficiency in LDLR is a cause of familial hypercholesterolemia (FH). Zhao et al. [92] and Lee et al. [110] created Ldlr KO rats. These ApoE and Ldlr KO rats mimic pathological changes observed in hyperlipidemia and atherosclerosis in humans with genetic deficiencies and in normal individuals, suggesting usefulness in the research of atherosclerosis.
Melanocortin-3 and -4 receptors (MC3R and MC4R) regulate energy homeostasis. You et al. [70] generated Mc3r and Mc4r single- and double-KO (DKO) rats using the CRISPR/Cas9 system. Mc3r KO rats displayed hypophagia and decreased body weight, whereas Mc4r KO and DKO rats exhibited hyperphagia and increased body weight. All three mutants showed increased white adipose tissue mass and adipocyte size. These mutant rats will be important in defining the complicated signaling pathways of MC3R and MC4R. According to You et al. [70], both Mc4r KO and DKO rats are good models for obesity and diabetes research.
Hereditary tyrosinemia type I (HT1) is caused by a deficiency in fumarylacetoacetate hydrolase (FAH) enzyme. Fah-deficient mice and pigs are phenotypically analogous to human HT1, but do not recapitulate all chronic features of the human disorder, especially liver fibrosis and cirrhosis. Zhang et al. [63] produced Fah KO rats through MI of CRISPR/Cas9 components to obtain HT1 models. The Fah KO rats faithfully represented hypertyrosinemia, liver failure, and renal tubular damage. More importantly, they developed remarkable liver fibrosis and cirrhosis, which have not been observed in Fah mutant mice or pigs. These data suggest that Fah KO rats may be used as an animal model of HT1 with liver cirrhosis.
Pseudoxanthoma elasticum (PXE)—a heritable ectopic mineralization disorder—is caused by mutations in the ATP-binding cassette subtype C number 6 (ABCC6) gene primarily expressed in the liver and kidneys. These mutations result in generalized arterial calcification throughout the body in infancy. Li et al. [75] generated Abcc6 KO rats as models of PXE using the ZFN technology. The plasma inorganic pyrophosphate (PPi) level was reduced (<30%), leading to a lowered PPi/inorganic phosphate plasma ratio. When in situ liver and kidney perfusions were performed, the PPi levels in the perfusates were significantly reduced, but those in the liver of WT rats remained high. Li et al. [75] speculate that hepatic ABCC6 may play a critical role in contributing to plasma PPi levels, identifying the liver as a target of molecular correction to counteract ectopic mineralization in PXE.
Wolfram syndrome (WS) is a rare autosomal-recessive disorder caused by mutations in the Wolfram syndrome 1 (WFS1) gene and characterized by juvenile-onset diabetes, optic atrophy, hearing loss, and a number of other complications. According to Plaas et al. [76], no mutant Wfs1 mice displayed fasting hyperglycemia. In other words, previous mouse models of WS involved only partial diabetes and other disease symptoms. Plaas et al. [76] generated Wfs1 KO rats, in which exon 5 of the Wfs1 gene was deleted, resulting in a loss of 27 amino acids from the WFS1 protein. The resulting KO rats showed progressive glucose intolerance, glycosuria, hyperglycemia, and severe body weight loss by 12 months of age. They also exhibited neuronal abnormality such as axonal degeneration and disorganization of the myelin. The phenotype of these KO rats indicates that they have the core symptoms of WS.
Leptin is a cytokine-like hormone principally produced by white adipose tissues. Defects in leptin production cause severe obesity. Leptin receptor (Lepr) encoded by the diabetes (db) gene is highly expressed in the choroid plexus. Bao et al. [59] produced Lepr KO rats through the CRISPR/Cas9 technology. The resulting KO rats exhibited obesity, hyperphagia, hyperglycemia, glucose intolerance, hyperinsulinemia, and dyslipidemia. In contrast, Chen et al. [79] generated Lepr KO rats through the TALEN technology. These rat models could complement the existing models (db/db mice and Zucker rats) [215,216] and be useful for biomedical and pharmacological research in obesity and diabetes.
Angiopoietin-like protein 8 (ANGPTL8) is a liver- and adipocyte-derived protein that controls plasma triglyceride levels. Izumi et al. [89] generated Angptl8 KO rats through the CRISPR/Cas9 technology to clarify the roles of Angptl8 in glucose and lipid metabolism. The resulting KO rats exhibited decreased body weight and fat content, associated with impaired lipogenesis in adipocytes. Izumi et al. [89] suggest that ANGPTL8 might be an important therapeutic target for obesity and dyslipidemia.
Hereditary aceruloplasminemia (HA) is a genetic disease characterized by iron accumulation in the liver and brain. The mutation of the ceruloplasmin (Cp gene is thought to be related to HA pathogenesis. Kenawi et al. [115] generated Cp KO rats through the CRISPR/Cas9 system. The Cp KO rats exhibited decreased iron concentration, transferrin saturation, plasma ceruloplasmin, and ferroxidase activity, which is considered essential for macrophage iron release. Thus, Cp KO rats can mimic the iron hepatosplenic phenotype in HA, which will form the basis to understand and treat the disease.
5.6. Models for Kidney Diseases
The renin (REN)-angiotensin system plays an important role in the control of blood pressure and renal function. Moreno et al. [31] produced Ren KO rats through ZFN-mediated GE system targeted to exon 5 of Ren. The resulting rats exhibited reduced body weight, lower blood pressure, and abnormal renal morphology (as exemplified by cortical interstitial fibrosis and abnormally shaped glomeruli). These results suggest the role of REN in the regulation of blood pressure and kidney function.
Primary hyperoxaluria type 1 (PH1) is an inherited disease caused by mutations in the mitochondrial localized alanine-glyoxylate aminotransferase (Agxt) gene, leading to abnormal metabolism of glyoxylic acid in the liver, subsequent endogenous oxalate overproduction, and deposition of oxalate in multiple organs, mainly the kidney. Patients with PH1 often suffer from recurrent urinary tract stones and finally renal failure. There is no effective treatment other than combined liver–kidney transplantation. Zheng et al. [94] produced Agxt KO rats as PH1 models through the CRISPR/Cas9 system. The resulting Agxt KO rats excreted more oxalate in the urine than WT animals and exhibited crystalluria with mild fibrosis in the kidney. These data suggest that Agxt-deficiency in mitochondria impairs glyoxylic acid metabolism and leads to PH1 in rats. This rat strain would be a valuable tool for developing innovative drugs and therapeutics.
Urate oxidase (uricase) encoded by UOX gene is a key enzyme whose disfunction causes hyperuricemia. Because of the low survival rate of Uox-deficient mice [217], Yu et al. [128] generated Uox KO rats through the CRISPR/Cas9 system targeting the exons 2 to 4 of Uox. The resulting Uox KO rats, called “Kunming-DY rats”, were apparently healthy, with more than a 95% survival up to one year. The male rats’ serum uric acid increased at levels significantly higher than those of WT rats. Kunming-DY rats exhibited histological renal changes including mild glomerular/tubular lesions, suggesting an alternative model animal to study hyperuricemia and associated diseases mimicking human conditions.
5.7. Models for Ophthalmology Diseases
The aryl hydrocarbon receptor (AHR) is a ligand-activated transcription factor which plays a role in the development of multiple tissues and is activated by a large number of ligands, including 2,3,7,8-tetrachlorodibenzo-p-dioxin. To examine the roles of the AHR in both normal biological development and response to environmental chemicals, Harrill et al. [40] produced Ahr KO rats through the ZFN technology targeting exon 2 of the Ahr and compared with an existing Ahr KO mouse model. Ahr KO rats, but not Ahr KO mice, displayed pathological alterations to the urinary tract, as exemplified by bilateral renal dilation (hydronephrosis). In contrast, abnormalities in vascular development were observed in Ahr KO mice, but not in rats. These findings suggest the differences in the role of AHR in tissue development, homeostasis, and toxicity between rats and mice.
5.8. Models for Hematological Systems
Hemophilia A is a genetic bleeding disorder resulting from factor VIII (FVIII or F8) deficiency. In preclinical hemophilia research, an animal model that reflects both the phenotype and pathology of the disease is required. Nielsen et al. [41] produced KO rats lacking detectable F8 activity through the ZFN technology targeting exon 16 of the F8 gene. Episodes of spontaneous bleeding requiring treatments were observed in 70% of F8 KO rats. Shi et al. [120] produced F8 KO rats in which nearly the entire rat F8 gene was inverted, causing translational stop six amino acids after the signal sequence, through the CRISPR/Cas9 technology to examine whether platelet F8 expression can prevent severe spontaneous bleeding in F8 KO rats. They showed that the severe spontaneous bleeding phenotype in F8 KO rats was successfully rescued by platelet-specific F8 expression through bone marrow cell transplantation.
Autoimmune regulator (AIRE) deficiency in humans induces a life-threatening generalized autoimmune disease called autoimmune polyendocrinopathy–candidiasis–ectodermal dystrophy (APECED), and no curative treatments are available. Several models of AIRE-deficient mice have been generated; although they have been useful in understanding the role of AIRE in central tolerance, they do not reproduce the APECED symptoms accurately and, thus, there is still a need for an animal model displaying APECED-like disease. Ossart et al. [86] produced an Aire KO rat model using ZFN technology. The resulting KO rats exhibited several of the key symptoms of APECED disease, including alopecia, skin depigmentation, and nail dystrophy, which are much more pronounced than those in Aire KO mice and closer to manifestations in humans.
5.9. Others
Estrogens play pivotal roles in the development and function of many organ systems, including the reproductive system. Rumi et al. [48] generated estrogen receptor 1 (Esr1) KO rats using the ZFN system targeting exon 3 of Esr1. Both male and female Esr1 KO rats were infertile. Esr1 KO males had small testes with distended and dysplastic STs, whereas Esr1 KO females possessed large polycystic ovaries, thread-like uteri, and poorly developed mammary glands. In addition, the uteri of Esr1 KO rats failed to 17β-estradiol treatment. This rat model provides a new experimental tool for investigating the pathophysiology of estrogen action.
The forkhead box N1 (FOXN1) gene is known as a critical factor for the differentiation of thymic and skin epithelial cells. Goto et al. [69] generated Foxn1 KO rats through the CRISPR/Cas9 technology. The resulting Foxn1 KO rats exhibited thymus deficiency and incomplete hairless, which was characterized by splicing variants.
Multidrug resistance 1 (MDR1; also known as P-glycoprotein) is a key efflux transporter that plays an important role not only in the transport of endogenous and exogenous substances, but also in tumor MDR, one of the most important impediments to the effective chemotherapy of cancer. In rodents, two isoforms, Mdr1a and Mdr1b, encode MDR1. Liang et al. [112] produced DKO rats (in which both Mdr1a and Mdr1b had been disrupted) through the CRISPR/Cas9 system. Pharmacokinetic studies of digoxin, a typical substrate of MDR1, confirmed the deficiency of MDR1 in resulting KO rats. This rat model is a useful tool for studying the function of MDR1 in drug absorption, tumor MDR, and drug-target validation.
6. Perspective
Genome-engineering technology, especially the CRISPR/Cas9 system, has opened ways to create KO at the specific genes and targeted KI, perform gene replacement, and conditionally ablate gene expression with more ease. Furthermore, recently, CRISPR-based new tools such as use of other types of Cas9 endonucleases (i.e., Staphylococcus aureus Cas9 (SaCas9), Francisella novicida Cas9 (FnCas9), Neisseria meningitidis Cas9 (NmCas9), Streptococcus thermophilus Cas9 (St1Cas9), and Brevibacillus laterosporus Cas9 (BlatCas9)) and Cas12a (Cpf1) endonucleases (i.e., Acidaminococcus sp. Cpf1 (AsCpf1) and Lachnospiraceae bacterium Cpf1 (LbCpf1)) (reviewed by Nakade et al. [218]), prime editing (PE), and BE have emerged. For example, BE systems, as exemplified by ABE, have already been applied in rats [97,104]. The PE system is the latest addition to the CRISPR genome-engineering toolkit and represents a novel approach to expand the scope of donor-free precise DNA editing [219]. PE-based manipulation of mouse genome has already been successfully performed [220]. This approach will soon be applied to modify the rat genome. Manipulation of mtDNA by DdCBE coupled with the PB transposon system has become possible in rats [153]. These new technologies, together with the newly developed gene delivery tools such as in vitro or in vivo EP-based gene delivery (TAKE and i-GONAD/rGONAD), will accelerate the creation of more GM rats, which will shed light on the new role of the previously known genes, whose function had been investigated using mouse models but was judged as being far from the function inferred from human diseases.
7. Conclusions
To date, tools for modifying the mouse genome as exemplified by traditional gene-targeting-based KO and KI technologies have been extensively employed in mice and have proven useful for understanding the molecular mechanism underlying human diseases. However, despite these efforts, the consequences obtained from the KO or KI mice were often different from those observed in human diseases. According to Barbaric et al. [221], discrepancies between the symptoms of human diseases and mouse phenotypes may be ascribed to redundant gene networks or alternative pathways or modifiers. This point was further strengthened when a number of GM rats were successfully produced through newly developed GE technologies (see Table 1). For example, Dmd KO rats (but not mice) presented cardiovascular alterations close to those observed in humans, which are the main cause of death in patients. Furthermore, Fah KO rats developed remarkable liver fibrosis and cirrhosis, which have not been observed in Fah mutant mice or pigs. These findings encourage the speculation that rats may better mimic the human situation than mice.
Author Contributions
M.S. and S.T. drafted the manuscript; S.N. and E.I. reviewed the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was partly supported by JSPS KAKENHI (No. 24580411 for M.S.; No. 16H05049 for S.N.; No. 21K10165 for E.I.; No. 16K07087 for S.T.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Conflicts of Interest
The founding sponsors had no role in the design of the study, the collection, analyses, or interpretation of data, the writing of the manuscript, or the decision to publish the results.
Abbreviations
| Abcc6 | ATP binding cassette subtype C number 6 |
| ABE | Adenine base editor |
| Abhd6 | Alpha/beta-hydrolase domain 6 |
| Adora2a | Adenosine 2a receptor |
| α-Gal A | α-galactosidase A |
| Agxt | Alanine-glyoxylate aminotransferase |
| Ahr | Aryl hydrocarbon receptor |
| Aire | Autoimmune regulator |
| ALS | Amyotrophic lateral sclerosis |
| Angptl8 | Angiopoietin-like protein 8 |
| Anks3 | Ankyrin repeat and sterile alpha motif domain containing 3 |
| APECED | Autoimmune polyendocrinopathy-candidiasis-ectodermal dystrophy |
| ApoE | Apolipoprotein E |
| AS | Angelman syndrome |
| AsCpf1 | Acidaminococcus sp. Cpf1 |
| ASD | Autism spectrum disorder |
| Asip | Agouti signaling protein |
| Atg16l1 | Autophagy-related 16-like 1 |
| B2m | Beta-2-microglobulin |
| BE | Base-editing |
| BlatCas9 | Brevibacillus laterosporus Cas9 |
| Bmpr2 | Bone morphogenetic protein receptor 2 |
| BN | Brown Norway |
| C3 | Complement C3 |
| C9orf72 | Chromosome 9 open reading frame 72 |
| Cas12a | CRISPR-associated protein 12a |
| Calb2 | Calbindin 2 |
| Carns1 | Carnosine synthase 1 |
| Cas9n | Cas9 nickase |
| CBE | Cytosine base editor |
| Ccdc85c | Coiled-coil domain containing 85C |
| Cck | Cholecystokinin |
| CD | Crohn’s disease |
| CES2 | Carboxylesterase 2 |
| Ces2a | Carboxylesterase 2A |
| CF | Cystic fibrosis |
| Cftr | Cystic fibrosis transmembrane conductance regulator |
| CIPN | Chemotherapy-induced peripheral neuropathy |
| Cirp | Cold-inducible RNA-binding protein |
| CLICK | CRISPR with lssDNA inducing conditional knockout alleles |
| Cp | Ceruloplasmin |
| Cpf1 | CRISPR from Prevotella and Francisella1 |
| Cplx1 | Complexin I |
| Cre-ERT2 | Cre recombinase-estrogen receptor T2 |
| Crhr1 | Corticotropin releasing hormone receptor 1 |
| CRISPR/Cas9 | Clustered regularly interspaced palindrome repeats (CRISPR)/Caspase 9 (Cas9) |
| Cas12a | Type V CRISPR-effector protein (formerly Cpf1) |
| crRNA | CRISPR RNA |
| CS | Cockayne syndrome |
| Csb | Cockayne syndrome B |
| Cyp2c11 | Cytochrome P450, subfamily 2, polypeptide 11 |
| Cyp2d | Cytochrome P450 family 2 subfamily D member 6 |
| Cyp2e1 | Cytochrome P450 2E1 |
| Cyp2J2 | Cytochrome P450 family 2 subfamily J member 2 |
| Cyp2J3 | Cytochrome P450, family 2, subfamily j, polypeptide 3 |
| Cyp2J10 | Cytochrome P450, family 2, subfamily j, polypeptide 10 |
| Cyp3a1 | Cytochrome P450 3A1 |
| Cyp3a2 | Cytochrome P450 3A2 |
| Cyp24a1 | Cytochrome P450 family 24 subfamily A member 1 |
| Cyp27b1 | Cytochrome P450 family 27 subfamily B member 1 |
| DA | Dark agouti |
| Dat | Dopamine transporter |
| db | Diabetes |
| Dbndd1 | Dysbindin (dystrobrevin binding protein 1) domain containing 1 |
| Defb23 | Defensin beta 23 |
| Defb26 | Defensin beta 26 |
| Defb42 | Defensin beta 42 |
| dCas9 | Dead Cas9 |
| DdCBE | DddA-derived cytosine base editor |
| DddA | Bacterial toxin deaminase |
| Depdc5 | DEP domain-containing 5 |
| Des | Desmin |
| DdCBE | DddA-derived cytosine base editor |
| DMD | Duchenne muscular dystrophy |
| Dmd | Dystrophin-coding gene (dystrophin) |
| DKO | Double KO |
| DNA-PKcs | DNA-dependent protein kinase catalytic subunits |
| Dnah17 | Dynein axonemal heavy chain 17 |
| Drd1a | Dopamine receptor D1A |
| DSB | Double-stranded break |
| dsDNA | Double-stranded DNA |
| dsRed | Discosoma coral red fluorescent protein |
| EGFP | Enhanced green fluorescent protein |
| EndoMT | Endothelial-to-mesenchymal transition |
| EP | Electroporation |
| Epsti1 | Epithelial stromal interaction 1 |
| Erbb3 | Erb-B2 receptor tyrosine kinase 3 |
| ERV | Endogenous retroviral element |
| ES | Embryonic stem |
| Esr1 | Estrogen receptor 1 |
| ET | Egg transfer |
| F344 | Fisher 344 |
| F8 | Factor 8 |
| Fabp2 | Fatty acid binding protein 2 |
| Fah | Fumarylacetoacetate hydrolase |
| FD | Fabry disease |
| Fh | Fumarate hydratase |
| FH | Familial hypercholesterolemia |
| FHH | Fawn-hooded hypertensive |
| Fmr1 | Fragile X mental retardation 1 |
| FMRP | Fragile X mental retardation protein |
| FnCas9 | Francisella novicida Cas9 |
| Foxp3 | Forkhead box P3 |
| Foxn1 | Forkhead box N1 |
| FSG | F344-scid gamma |
| Fus | Fused in sarcoma |
| FXS | Fragile X syndrome |
| Gaa | Acid alpha-glucosidase |
| Gad1 | Glutamate decarboxylase 1 |
| GAD67 | Glutamate decarboxylase 67-kDa isoform |
| GE | Genome editing or genome-edited |
| GFP | Green fluorescent protein |
| G6pd | Glucose-6-phosphate dehydrogenase |
| GM | Genetically modified |
| GOI | Gene of interest |
| GONAD | Genome-editing via oviductal nucleic acids delivery |
| GR | Glucocorticoid receptors |
| GRdim | Glucocorticoid receptor mutant |
| Grin1 | Glutamate ionotropic receptor N-methyl-D-aspartate (NMDA) type subunit 1 |
| gRNA | Guide RNA |
| Gsd2 | Glycogen storage disease type 2 |
| GSDII | Glycogen storage disease type II (GSDII) |
| 2H2OP | Two-hit by gRNA and two-oligo with plasmid |
| HA | Hereditary aceruloplasminemia |
| HCD | High-cholesterol diet |
| HDR | Homology directed repair |
| Hemgn | Hemogen |
| hhy | Hemorrhagic hydrocephalus |
| Hprt1 | Hypoxanthine phosphoribosyltransferase 1 |
| HR | Homologous recombination |
| HT1 | Hereditary tyrosinemia type I |
| Htr7 | 5-hydroxytryptamine receptor 7 |
| IC | Intracytoplasmic injection |
| Ighm | Immunoglobulin heavy constant mu |
| IgM | Immunoglobulin M |
| i-GONAD | Improved GONAD |
| Il2rg | Interleukin 2 receptor gamma |
| indels | Insertion or deletion of nucleotides |
| iNOS | Inducible nitric oxide synthase |
| IRES | Internal ribosomal entry site |
| IVF | In vitro fertilized |
| Kat II | Kynurenine aminotransferase II |
| Kcnj16 | Potassium inwardly-rectifying channel, subfamily J, member 16 |
| Kcnk3 | Potassium channel subfamily K member 3 |
| KI | Knock-in |
| Kit | Kit proto-oncogene |
| KO | Knockout |
| L1cam | L1 cell adhesion molecule |
| LbCpf1 | Lachnospiraceae bacterium Cpf1 |
| Ldlr | Low-density lipoprotein receptor |
| Lrat | Lecithin retinol acyltransferase |
| Lepr | Leptin receptor |
| LE | Long–Evans |
| LEH | Long–Evans hooded |
| Lepr | Leptin receptor |
| LEW | Lewis |
| Lgr5 | Leucine-rich repeat-containing G-protein coupled receptor 5 |
| lncRNA | Long non-coding RNA |
| Lrat | Lecithin retinol acyltransferase |
| lssDNA | Long single-stranded DNA |
| Mc3r | Melanocortin 3 receptor |
| Mc4r | Melanocortin 4 receptor |
| Mdr1a | Multidrug resistance protein 1a |
| Mdr1b | Multidrug resistance protein 1b |
| Mdx | X-linked muscular dystrophy |
| Mecp2 | Methyl-CpG-binding protein 2 |
| MI | Microinjection |
| mitoTALE | TALE nickase enabling mitochondrial gene editing |
| Mkx | Mohawk homeobox |
| Mnx1 | Gene encoding homeobox protein HB9 |
| mtDNA | Mitochondrial DNA |
| mTORC1 | Rapamycin complex 1 signaling pathway |
| Myl4 | Myosin light-chain 4 |
| NAFLD | Nonalcoholic fatty liver disease |
| nCas9 | Cas9 nickase |
| Ndst1 | N-deacetylase |
| Ndst4 | N-sulfotransferase 4 |
| Nes | Nestin |
| Nfatc1 | Nuclear factor of activated T-cells |
| Ngln3 | Neuroligin 3 |
| NHEJ | Non-homologous end joining |
| Nkx3.1 | NK3 homeobox 1 |
| NmCas9 | Neisseria meningitidis Cas9 |
| Nr3c1 | Nuclear receptor subfamily 3, group C, member 1 |
| Nrf2 | Nuclear factor (erythroid-derived 2)-like-2 |
| Oatp1b2 | Organic anion transport polypeptide 1b2 |
| Ogdh | Oxoglutarate dehydrogenase |
| PAH | Pulmonary arterial hypertension |
| PAM | Protospacer adjacent motif |
| PB | PiggyBack transposon |
| PBS | Phosphate-buffered saline |
| PE | Prime editing |
| PGK | Phosphoglycerate kinase |
| PH | Pulmonary hypertension |
| PH1 | Primary hyperoxaluria type 1 |
| PI | Pronuclear injection |
| PMCA4 | Plasma membrane calmodulin–dependent calcium ATPase isoform 4 |
| PMS | Phelan–McDermid syndrome |
| Pnpla5 | Patatin-like phospholipase domain containing 5 |
| poly(A) | Polyadenine tails |
| Pomc | Proopiomelanocortin |
| Pp | Poring pulse |
| PPi | Pyrophosphate |
| Prf1 | Perforin 1 |
| Prkdc | DNA-dependent protein kinase catalytic subunit |
| Psen1 | Presenilin 1 |
| p52SHC/p46SHC | Adaptor protein 1 (shc1), encodes 3 main protein isoforms |
| Pvalb | Parvalbumin |
| PVG | Piebald Virol Glaxo |
| PXE | Pseudoxanthoma elasticum |
| Pxr | Pregnane X receptor |
| rAAV | Recombinant adeno-associated virus |
| Rab38 | Ras-related protein Rab-38 |
| Rag1 | Recombination activating gene 1 |
| Rag2 | Recombination activating gene 2 |
| RD | Retinal dystrophy |
| Ren | Renin |
| Rffl-lnc1 | A gene located within the 5′UTR intronic region of the rififylin (Rffl) gene |
| rGONAD | Rat-based i-GONAD |
| RNaseT2 | Ribonuclease T2 |
| RNP | Ribonucleoprotein |
| Rosa26 | Gt(ROSA)26Sor |
| RP | Retinitis pigmentosa |
| RS | Rett syndrome |
| SaCas9 | Staphylococcus aureus Cas9 |
| SCID | Severe combined immunodeficiency |
| SD | Sprague–Dawley |
| SERCA2a | Sarcoplasmic reticulum Ca2+-ATPase |
| sgRNA | Single-guide RNA |
| Shank3 | SH3 and multiple ankyrin repeat domains 3 |
| Sirpa | Signal regulatory protein alpha |
| Slc6 | Sodium- and chloride-dependent members of the solute carrier family 6 |
| Slc16a2 | Solute carrier family 16 member 2 coding for monocarboxylate transporter 8 (MCT8) |
| Slco1b2 | Solute carrier organic anion transporter family member 1B2 |
| SNP | Single-nucleotide polymorphism |
| Sox10 | Sry-box transcription factor 10 |
| Sox16 | SRY (sex determining region Y)-box 16 |
| SS | Dahl salt-sensitive |
| SSCs | Spermatogonial stem cells |
| ssDNA | Single-stranded DNA |
| ssODN | Single-stranded oligodeoxynucleotide |
| St1Cas9 | Streptococcus thermophilus Cas9 |
| STs | Seminiferous tubules |
| TAKE | Technique for Animal Knockout system by Electroporation |
| TALE | Transcriptional activator-like effector |
| TALENs | Transcription activator-like effector nucleases |
| Tet1 | Tet methylcytosine dioxygenase 1 |
| Tet2 | Tet methylcytosine dioxygenase 2 |
| Tet3 | Tet methylcytosine dioxygenase 3 |
| Tg | Transgenic |
| Th | Tyrosine hydroxylase |
| Thy1 | Thy-1 cell surface antigen |
| Tie2 | Tunica interna endothelial cell kinase 2 |
| Tlr4 | Toll-like receptor 4 |
| TM/Kyo | TRM or tremor rat |
| Tp | Transfer pulse |
| Tp53 | Tumor protein p53 |
| tracrRNA | Trans-activating CRISPR RNA |
| Trdmt1 | TRNA aspartic acid methyltransferase 1 |
| Trpa1 | Transient receptor potential family member ankyrin 1 |
| Trpv4 | Transient receptor potential cation channel subfamily V member 4 |
| Tyr | Tyrosinase |
| Ube3a | Ubiquitin protein ligase E3A |
| Uox | Urate oxidase (uricase) |
| Vapb | Vesicle-associated membrane protein-associated protein B/C |
| Vdr | Vitamin D receptor |
| Wfs1 | Wolfram syndrome 1 |
| WI | Wistar |
| WI-IM | Wistar–Imamichi |
| WKY | Wistar–Kyoto |
| Wnt1 | Wnt family member 1 |
| WS | Wolfram syndrome |
| WT | Wild-type |
| XLH | X-linked hydrocephalus |
| X-SCID | X-linked severe combined immunodeficiency |
| ZF | Zinc-finger |
| ZFNs | Zinc-finger nucleases |
| ZP | Zona pellucida |
References
- Kjell, J.; Olson, L. Rat models of spinal cord injury: From pathology to potential therapies. Dis. Models Mech. 2016, 9, 1125–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacob, H.J. The Rat: A model used in biomedical research. In Rat Genomics: Methods and Protocols Methods in Molecular Biology; Anegon, I., Ed.; Humana Press: Totowa, NJ, USA, 2010; pp. 1–11. [Google Scholar] [CrossRef]
- Hammes, A.; Oberdorf-Maass, S.; Rother, T.; Nething, K.; Gollnick, F.; Linz, K.W.; Meyer, R.; Hu, K.; Han, H.; Gaudron, P.; et al. Overexpression of the sarcolemmal calcium pump in the myocardium of transgenic rats. Circ. Res. 1998, 83, 877–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vetter, R.; Rehfeld, U.; Reissfelder, C.; Weifl, W.; Wagner, K.D.; Günther, J.; Hammes, A.; Tschöpe, C.; Dillmann, W.; Paul, M. Transgenic overexpression of the sarcoplasmic reticulum Ca2+ ATPase improves reticular Ca2+ handling in normal and diabetic rat hearts. FASEB J. 2002, 16, 1657–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, K.R.; Capecchi, M.R. Site-directed mutagenesis by gene targeting in mouse embryo-derived stem cells. Cell 1987, 51, 503–512. [Google Scholar] [CrossRef]
- Doetschman, T.; Gregg, R.G.; Maeda, N.; Hooper, M.L.; Melton, D.W.; Thompson, S.; Smithies, O. Targetted correction of a mutant HPRT gene in mouse embryonic stem cells. Nature 1987, 330, 576–578. [Google Scholar] [CrossRef]
- Buehr, M.; Meek, S.; Blair, K.; Yang, J.; Ure, J.; Silva, J.; McLay, R.; Hall, J.; Ying, Q.L.; Smith, A. Capture of authentic embryonic stem cells from rat blastocysts. Cell 2008, 135, 1287–1298. [Google Scholar] [CrossRef]
- Li, P.; Tong, C.; Mehrian-Shai, R.; Jia, L.; Wu, N.; Yan, Y.; Maxson, R.E.; Schulze, E.N.; Song, H.; Hsieh, C.L.; et al. Germline competent embryonic stem cells derived from rat blastocysts. Cell 2008, 135, 1299–1310. [Google Scholar] [CrossRef] [Green Version]
- Kawamata, M.; Ochiya, T. Generation of genetically modified rats from embryonic stem cells. Proc. Natl. Acad. Sci. USA 2010, 107, 14223–14228. [Google Scholar] [CrossRef] [Green Version]
- Meek, S.; Buehr, M.; Sutherland, L.; Thomson, A.; Mullins, J.J.; Smith, A.J.; Burdon, T. Efficient gene targeting by homologous recombination in rat embryonic stem cells. PLoS ONE 2010, 5, e14225. [Google Scholar] [CrossRef]
- Tong, C.; Li, P.; Wu, N.L.; Yan, Y.; Ying, Q.L. Production of p53 gene knockout rats by homologous recombination in embryonic stem cells. Nature 2010, 467, 211–213. [Google Scholar] [CrossRef] [Green Version]
- Lan, H.; Li, S.; Guo, Z.; Men, H.; Wu, Y.; Li, N.; Bryda, E.C.; Capecchi, M.R.; Wu, S. Efficient generation of selection-gene-free rat knockout models by homologous recombination in ES cells. FEBS Lett. 2016, 590, 3416–3424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, T.; Yamaguchi, T.; Hamanaka, S.; Kato-Itoh, M.; Yamazaki, Y.; Ibata, M.; Sato, H.; Lee, Y.S.; Usui, J.; Knisely, A.S.; et al. Generation of rat pancreas in mouse by interspecific blastocyst injection of pluripotent stem cells. Cell 2010, 142, 787–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, S.; Ooshima, Y.; Nakata, M.; Yano, T.; Nishimura, N.; Nishigaki, R.; Satomi, Y.; Matsumoto, H.; Matsumoto, Y.; Takeyama, M. Efficient gene-targeting in rat embryonic stem cells by CRISPR/Cas and generation of human kynurenine aminotransferase II (KAT II) knock-in rat. Transgenic Res. 2015, 24, 991–1001. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Spitzer, S.; Agathou, S.; Karadottir, R.T.; Smith, A. Gene editing in rat embryonic stem cells to produce in vitro models and in vivo reporters. Stem Cell Rep. 2017, 9, 1262–1274. [Google Scholar] [CrossRef] [Green Version]
- Zan, Y.; Haag, J.D.; Chen, K.S.; Shepel, L.A.; Wigington, D.; Wang, Y.R.; Hu, R.; Lopez-Guajardo, C.C.; Brose, H.L.; Porter, K.I.; et al. Production of knockout rats using ENU mutagenesis and a yeast-based screening assay. Nat. Biotechnol. 2003, 21, 645–651. [Google Scholar] [CrossRef]
- Harrison, M.M.; Jenkins, B.V.; O’Connor-Giles, K.M.; Wildonger, J.A. CRISPR view of development. Genes Dev. 2014, 28, 1859–1872. [Google Scholar] [CrossRef] [Green Version]
- Hsu, P.D.; Lander, E.S.; Zhang, F. Development and applications of CRISPR-Cas9 for genome engineering. Cell 2014, 157, 1262–1278. [Google Scholar] [CrossRef] [Green Version]
- Sonoda, E.; Hochegger, H.; Saberi, A.; Taniguchi, Y.; Takeda, S. Differential usage of non-homologous end-joining and homologous recombination in double strand break repair. DNA Repair 2006, 5, 1021–1029. [Google Scholar] [CrossRef]
- Kim, Y.G.; Cha, J.; Chandrasegaran, S. Hybrid restriction enzymes: Zinc finger fusions to Fok I cleavage domain. Proc. Natl. Acad. Sci. USA 1996, 93, 1156–1160. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, S.A.; Nekludova, L.; Pabo, C.O. DNA recognition by Cys2His2 zinc finger proteins. Annu. Rev. Biophys. Biomol. Struct. 2000, 29, 183–212. [Google Scholar] [CrossRef]
- Moscou, M.J.; Bogdanove, A.J. A simple cipher governs DNA recognition by TAL effectors. Science 2009, 326, 1501. [Google Scholar] [CrossRef] [PubMed]
- Christian, M.; Cermak, T.; Doyle, E.L.; Schmidt, C.; Zhang, F.; Hummel, A.; Bogdanove, A.J.; Voytas, D.F. Targeting DNA double-strand breaks with TAL effector nucleases. Genetics 2010, 186, 757–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Huang, S.; Jiang, W.Z.; Wright, D.; Spalding, M.H.; Weeks, D.P.; Yang, B. TAL nucleases (TALNs): Hybrid proteins composed of TAL effectors and FokI DNA-cleavage domain. Nucleic Acids Res. 2011, 39, 359–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Gaj, T.; Gersbach, C.A.; Barbas, C.F., III. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2013, 31, 397–405. [Google Scholar] [CrossRef] [Green Version]
- Doudna, J.A.; Charpentier, E. Genome editing. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef]
- Geurts, A.M.; Cost, G.J.; Freyvert, Y.; Zeitler, B.; Miller, J.C.; Choi, V.M.; Jenkins, S.S.; Wood, A.; Cui, X.; Meng, X.; et al. Knockout rats via embryo microinjection of zinc-finger nucleases. Science 2009, 325, 433. [Google Scholar] [CrossRef] [Green Version]
- Mashimo, T.; Takizawa, A.; Voigt, B.; Yoshimi, K.; Hiai, H.; Kuramoto, T.; Serikawa, T. Generation of knockout rats with X-linked severe combined immunodeficiency (X-SCID) using zinc-finger nucleases. PLoS ONE 2010, 5, e8870. [Google Scholar] [CrossRef]
- Ménoret, S.; Iscache, A.L.; Tesson, L.; Rémy, S.; Usal, C.; Osborn, M.J.; Cost, G.J.; Brüggemann, M.; Buelow, R.; Anegon, I. Characterization of immunoglobulin heavy chain knockout rats. Eur. J. Immunol. 2010, 40, 2932–2941. [Google Scholar] [CrossRef]
- Moreno, C.; Hoffman, M.; Stodola, T.J.; Didier, D.N.; Lazar, J.; Geurts, A.M.; North, P.E.; Jacob, H.J.; Greene, A.S. Creation and characterization of a renin knockout rat. Hypertension 2011, 57, 614–619. [Google Scholar] [CrossRef] [Green Version]
- Tesson, L.; Usal, C.; Ménoret, S.; Leung, E.; Niles, B.J.; Remy, S.; Santiago, Y.; Vincent, A.I.; Meng, X.; Zhang, L.; et al. Knockout rats generated by embryo microinjection of TALENs. Nat. Biotechnol. 2011, 29, 695–696. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Ji, D.; Fisher, D.A.; Wu, Y.; Briner, D.M.; Weinstein, E.J. Targeted integration in rat and mouse embryos with zinc-finger nucleases. Nat. Biotechnol. 2011, 29, 64–67. [Google Scholar] [CrossRef]
- Mashimo, T.; Takizawa, A.; Kobayashi, J.; Kunihiro, Y.; Yoshimi, K.; Ishida, S.; Tanabe, K.; Yanagi, A.; Tachibana, A.; Hirose, J.; et al. Generation and characterization of severe combined immunodeficiency rats. Cell Rep. 2012, 2, 685–694. [Google Scholar] [CrossRef] [Green Version]
- Ménoret, S.; Fontanière, S.; Jantz, D.; Tesson, L.; Thinard, R.; Rémy, S.; Usal, C.; Ouisse, L.H.; Fraichard, A.; Anegon, I. Generation of Rag1-knockout immunodeficient rats and mice using engineered meganucleases. FASEB J. 2013, 27, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.J.; Fisher, D.A.; Kouranova, E.; McCoy, A.; Forbes, K.; Wu, Y.; Henry, R.; Ji, D.; Chambers, A.; Warren, J.; et al. Whole-rat conditional gene knockout via genome editing. Nat. Methods 2013, 10, 638–640. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Qiu, Z.; Shao, Y.; Chen, Y.; Guan, Y.; Liu, M.; Li, Y.; Gao, N.; Wang, L.; Lu, X.; et al. Heritable gene targeting in the mouse and rat using a CRISPR-Cas system. Nat. Biotechnol. 2013, 31, 681–683. [Google Scholar] [CrossRef]
- Li, W.; Teng, F.; Li, T.; Zhou, Q. Simultaneous generation and germline transmission of multiple gene mutations in rat using CRISPR-Cas systems. Nat. Biotechnol. 2013, 31, 684–686. [Google Scholar] [CrossRef]
- Ferguson, C.; McKay, M.; Harris, R.A.; Homanics, G.E. Toll-like receptor 4 (Tlr4) knockout rats produced by transcriptional activator-like effector nuclease (TALEN)-mediated gene inactivation. Alcohol 2013, 47, 595–599. [Google Scholar] [CrossRef] [Green Version]
- Harrill, J.A.; Hukkanen, R.R.; Lawson, M.; Martin, G.; Gilger, B.; Soldatow, V.; Lecluyse, E.L.; Budinsky, R.A.; Rowlands, J.C.; Thomas, R.S. Knockout of the aryl hydrocarbon receptor results in distinct hepatic and renal phenotypes in rats and mice. Toxicol. Appl. Pharmacol. 2013, 272, 503–518. [Google Scholar] [CrossRef]
- Nielsen, L.N.; Wiinberg, B.; Häger, M.; Holmberg, H.L.; Hansen, J.J.; Roepstorff, K.; Tranholm, M. A novel F8−/− rat as a translational model of human hemophilia A. J. Thromb. Haemost. 2014, 12, 1274–1282. [Google Scholar] [CrossRef]
- Hamilton, S.M.; Green, J.R.; Veeraragavan, S.; Yuva, L.; McCoy, A.; Wu, Y.; Warren, J.; Little, L.; Ji, D.; Cui, X.; et al. Fmr1 and Nlgn3 knockout rats: Novel tools for investigating autism spectrum disorders. Behav. Neurosci. 2014, 128, 103–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneko, T.; Sakuma, T.; Yamamoto, T.; Mashimo, T. Simple knockout by electroporation of engineered endonucleases into intact rat embryos. Sci. Rep. 2014, 4, 6382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remy, S.; Tesson, L.; Menoret, S.; Usal, C.; De Cian, A.; Thepenier, V.; Thinard, R.; Baron, D.; Charpentier, M.; Renaud, J.B.; et al. Efficient gene targeting by homology-directed repair in rat zygotes using TALE nucleases. Genome Res. 2014, 24, 1371–1383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Ma, J.; Zhang, X.; Chen, W.; Yu, L.; Lu, Y.; Bai, L.; Shen, B.; Huang, X.; Zhang, L. Generation of eGFP and Cre knockin rats by CRISPR/Cas9. FEBS J. 2014, 281, 3779–3790. [Google Scholar] [CrossRef]
- Yoshimi, K.; Kaneko, T.; Voigt, B.; Mashimo, T. Allele-specific genome editing and correction of disease-associated phenotypes in rats using the CRISPR–Cas platform. Nat. Commun. 2014, 5, 4240. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Shen, B.; Zhang, X.; Lu, Y.; Chen, W.; Ma, J.; Huang, X.; Zhang, L. Heritable multiplex genetic engineering in rats using CRISPR/Cas9. PLoS ONE 2014, 9, e89413. [Google Scholar] [CrossRef]
- Rumi, M.A.K.; Dhakal, P.; Kubota, K.; Chakraborty, D.; Lei, T.; Larson, M.A.; Wolfe, M.W.; Roby, K.F.; Vivian, J.L.; Soares, M.J. Generation of Esr1-knockout rats using zinc finger nuclease-mediated genome editing. Endocrinology 2014, 155, 1991–1999. [Google Scholar] [CrossRef] [Green Version]
- De León, V.P.; Anne-Marie, M.; Laurent, T.; Ignacio, A.; Edith, H. Generation of TALEN-mediated GRdim knock-in rats by homologous recombination. PLoS ONE 2014, 9, e88146. [Google Scholar] [CrossRef]
- Nakamura, K.; Fujii, W.; Tsuboi, M.; Tanihata, J.; Teramoto, N.; Takeuchi, S.; Naito, K.; Yamanouchi, K.; Nishihara, M. Generation of muscular dystrophy model rats with a CRISPR/Cas system. Sci. Rep. 2014, 4, 5635. [Google Scholar] [CrossRef]
- Larcher, T.; Lafoux, A.; Tesson, L.; Remy, S.; Thepenier, V.; François, V.; Le Guiner, C.; Goubin, H.; Dutilleul, M.; Guigand, L.; et al. Characterization of dystrophin deficient rats: A new model for Duchenne muscular dystrophy. PLoS ONE 2014, 9, e110371. [Google Scholar] [CrossRef] [Green Version]
- Tuggle, K.L.; Birket, S.E.; Cui, X.; Hong, J.; Warren, J.; Reid, L.; Chambers, A.; Ji, D.; Gamber, K.; Chu, K.K.; et al. Characterization of defects in ion transport and tissue development in cystic fibrosis transmembrane conductance regulator (CFTR)-knockout rats. PLoS ONE 2014, 9, e91253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranchoux, B.; Antigny, F.; Rucker-Martin, C.; Hautefort, A.; Péchoux, C.; Bogaard, H.J.; Dorfmüller, P.; Remy, S.; Lecerf, F.; Planté, S.; et al. Endothelial-to-mesenchymal transition in pulmonary hypertension. Circulation 2015, 131, 1006–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneko, T.; Mashimo, T. Simple genome editing of rodent intact embryos by electroporation. PLoS ONE 2015, 10, e0142755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Xie, D.; Huang, J.; Lv, F.; Shi, D.; Liu, Y.; Lin, L.; Geng, L.; Wu, Y.; Liang, D.; et al. Cold-inducible RNA-binding protein regulates cardiac repolarization by targeting transient outward potassium channels. Circ. Res. 2015, 116, 1655–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, S.; Zhang, Y.; Su, L.; He, K.; Wang, Q.; Zhang, Y.; Yang, D.; Yang, Y.; Ma, S. Apolipoprotein E-deficient rats develop atherosclerotic plaques in partially ligated carotid arteries. Atherosclerosis 2015, 243, 589–592. [Google Scholar] [CrossRef] [PubMed]
- Ménoret, S.; De Cian, A.; Tesson, L.; Remy, S.; Usal, C.; Boulé, J.B.; Boix, C.; Fontanière, S.; Crénéguy, A.; Nguyen, T.H.; et al. Homology-directed repair in rodent zygotes using Cas9 and TALEN engineered proteins. Sci. Rep. 2015, 5, 14410. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Shao, Y.; Guan, Y.; Li, L.; Wu, L.; Chen, F.; Liu, M.; Chen, H.; Ma, Y.; Ma, X.; et al. Large genomic fragment deletion and functional gene cassette knock-in via Cas9 protein mediated genome editing in one-cell rodent embryos. Sci. Rep. 2015, 5, 17517. [Google Scholar] [CrossRef] [Green Version]
- Bao, D.; Ma, Y.; Zhang, X.; Guan, F.; Chen, W.; Gao, K.; Qin, C.; Zhang, L. Preliminary characterization of a leptin receptor knockout rat created by CRISPR/Cas9 system. Sci. Rep. 2015, 5, 15942. [Google Scholar] [CrossRef] [Green Version]
- Chapman, K.M.; Medrano, G.A.; Jaichander, P.; Chaudhary, J.; Waits, A.E.; Nobrega, M.A.; Hotaling, J.M.; Ober, C.; Hamra, F.K. Targeted germline modifications in rats using CRISPR/Cas9 and spermatogonial stem cells. Cell Rep. 2015, 10, 1828–1835. [Google Scholar] [CrossRef] [Green Version]
- Zhai, W.; Hu, H.; Le, L.; Zhuang, F.; Wang, K.; Zhao, Y.; Wang, K.Z.; Liu, X.M.; Sun, D.; Wang, X.Y.; et al. Generation and analysis of the Rett syndrome-associated MeCP2-null rat model. Yi Chuan 2016, 38, 1004–1011. [Google Scholar] [CrossRef]
- Patterson, K.C.; Hawkins, V.E.; Arps, K.M.; Mulkey, D.K.; Olsen, M.L. MeCP2 deficiency results in robust Rett-like behavioural and motor deficits in male and female rats. Hum. Mol. Genet. 2016, 25, 3303–3320. [Google Scholar] [CrossRef]
- Zhang, L.; Shao, Y.; Li, L.; Tian, F.; Cen, J.; Chen, X.; Hu, D.; Zhou, Y.; Xie, W.; Zheng, Y.; et al. Efficient liver repopulation of transplanted hepatocyte prevents cirrhosis in a rat model of hereditary tyrosinemia type I. Sci. Rep. 2016, 6, 31460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimi, K.; Kunihiro, Y.; Kaneko, T.; Nagahora, H.; Voigt, B.; Mashimo, T. ssODN-mediated knock-in with CRISPR-Cas for large genomic regions in zygotes. Nat. Commun. 2016, 7, 10431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renaud, J.B.; Boix, C.; Charpentier, M.; De Cian, A.; Cochennec, J.; Duvernois-Berthet, E.; Perrouault, L.; Tesson, L.; Edouard, J.; Thinard, R.; et al. Improved genome editing efficiency and flexibility using modified oligonucleotides with TALEN and CRISPR-Cas9 nucleases. Cell Rep. 2016, 14, 2263–2272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, D.; Zhong, Y.; Li, X.; Li, Y.; Li, X.; Cao, J.; Fan, Z.; Fan, H.; Yuan, L.; Xu, B.; et al. Generation of TALEN-mediated FH knockout rat model. Oncotarget 2016, 7, 61656–61669. [Google Scholar] [CrossRef] [Green Version]
- Marsan, E.; Ishida, S.; Schramm, A.; Weckhuysen, S.; Muraca, G.; Lecas, S.; Liang, N.; Treins, C.; Pende, M.; Roussel, D.; et al. Depdc5 knockout rat: A novel model of mTORopathy. Neurobiol. Dis. 2016, 89, 180–189. [Google Scholar] [CrossRef]
- Ma, Y.; Chen, W.; Zhang, X.; Yu, L.; Dong, W.; Pan, S.; Gao, S.; Huang, X.; Zhang, L. Increasing the efficiency of CRISPR/Cas9-mediated precise genome editing in rats by inhibiting NHEJ and using Cas9 protein. RNA Biol. 2016, 13, 605–612. [Google Scholar] [CrossRef] [Green Version]
- Goto, T.; Hara, H.; Nakauchi, H.; Hochi, S.; Hirabayashi, M. Hypomorphic phenotype of Foxn1 gene-modified rats by CRISPR/Cas9 system. Transgenic Res. 2016, 25, 533–544. [Google Scholar] [CrossRef]
- You, P.; Hu, H.; Chen, Y.; Zhao, Y.; Yang, Y.; Wang, T.; Xing, R.; Shao, Y.; Zhang, W.; Li, D.; et al. Effects of melanocortin 3 and 4 receptor deficiency on energy homeostasis in rats. Sci. Rep. 2016, 6, 34938. [Google Scholar] [CrossRef] [Green Version]
- Priestley, J.R.C.; Kautenburg, K.E.; Casati, M.C.; Endres, B.T.; Geurts, A.M.; Lombard, J.H. The NRF2 knockout rat: A new animal model to study endothelial dysfunction, oxidant stress, and microvascular rarefaction. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H478–H487. [Google Scholar] [CrossRef] [Green Version]
- Taguchi, K.; Takaku, M.; Egner, P.A.; Morita, M.; Kaneko, T.; Mashimo, T.; Kensler, T.W.; Yamamoto, M. Generation of a new model rat: Nrf2 knockout rats are sensitive to aflatoxin B1 toxicity. Toxicol. Sci. 2016, 152, 40–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Tang, Y.; Lu, J.; Shao, Y.; Qin, X.; Li, Y.; Wang, L.; Li, D.; Liu, M. Characterization of novel cytochrome P450 2E1 knockout rat model generated by CRISPR/Cas9. Biochem. Pharmacol. 2016, 105, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Ito, Y.; Shinohara, M.; Yamashita, S.; Ichinose, S.; Kishida, A.; Oyaizu, T.; Kayama, T.; Nakamichi, R.; Koda, N.; et al. Gene targeting of the transcription factor Mohawk in rats causes heterotopic ossification of Achilles tendon via failed tenogenesi. Proc. Natl. Acad. Sci. USA 2016, 113, 7840–7845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Kingman, J.; van de Wetering, K.; Tannouri, S.; Sundberg, J.P.; Uitto, J. Abcc6 knockout rat model highlights the role of liver in PPi homeostasis in Pseudoxanthoma elasticum. J. Investig. Dermatol. 2017, 137, 1025–1032. [Google Scholar] [CrossRef] [PubMed]
- Plaas, M.; Seppa, K.; Reimets, R.; Jagomäe, T.; Toots, M.; Koppel, T.; Vallisoo, T.; Nigul, M.; Heinla, I.; Meier, R.; et al. Wfs1-deficient rats develop primary symptoms of Wolfram syndrome: Insulin-dependent diabetes, optic nerve atrophy and medullary degeneration. Sci. Rep. 2017, 7, 10220. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Yang, C.; Shang, S.; Cai, Y.; Deng, X.; Zhang, J.; Shao, F.; Zhu, D.; Liu, Y.; Chen, G.; et al. Loss of FMRP impaired hippocampal long-term plasticity and spatial learning in rats. Front. Mol. Neurosci. 2017, 10, 269. [Google Scholar] [CrossRef] [Green Version]
- Peng, W.; Li, M.; Li, H.; Tang, K.; Zhuang, J.; Zhang, J.; Xiao, J.; Jiang, H.; Li, D.; Yu, Y.; et al. Dysfunction of myosin light-chain 4 (MYL4) leads to heritable atrial cardiomyopathy with electrical, contractile, and structural components: Evidence from genetically-engineered rats. J. Am. Heart Assoc. 2017, 6, e007030. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Lu, W.; Gao, N.; Long, Y.; Shao, Y.; Liu, M.; Chen, H.; Ye, S.; Ma, X.; Liu, M.; et al. Generation of obese rat model by transcription activator-like effector nucleases targeting the leptin receptor gene. Sci. China Life Sci. 2017, 60, 152–157. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Gao, Q.; Zhang, X.; Huang, L.; Xu, K.; Hu, Y.; Liu, L.; Mu, Y.; Li, K. PNPLA5-knockout rats induced by CRISPR/Cas9 exhibit abnormal bleeding and lipid level. J. Integr. Agric. 2017, 16, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Harony-Nicolas, H.; Kay, M.; du Hoffmann, J.; Klein, M.E.; Bozdagi-Gunal, O.; Riad, M.; Daskalakis, N.P.; Sonar, S.; Castillo, P.E.; Hof, P.R.; et al. Oxytocin improves behavioral and electrophysiological deficits in a novel Shank3-deficient rat. eLife 2017, 6, e18904. [Google Scholar] [CrossRef]
- Cheng, X.; Waghulde, H.; Mell, B.; Morgan, E.E.; Pruett-Miller, S.M.; Joe, B. Positional cloning of quantitative trait nucleotides for blood pressure and cardiac QT-interval by targeted CRISPR/Cas9 editing of a novel long non-coding RNA. PLoS Genet. 2017, 13, e1006961. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Lai, D.H.; Wilson, R.A.; Chen, Y.F.; Wang, L.F.; Yu, Z.L.; Li, M.Y.; He, P.; Hide, G.; Sun, X.; et al. Nitric oxide blocks the development of the human parasite Schistosoma japonicum. Proc. Natl. Acad. Sci. USA 2017, 114, 10214–10219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palygin, O.; Levchenko, V.; Ilatovskaya, D.V.; Pavlov, T.S.; Pochynyuk, O.M.; Jacob, H.J.; Geurts, A.M.; Hodges, M.R.; Staruschenko, A. Essential role of Kir5.1 channels in renal salt handling and blood pressure control. JCI Insight 2017, 2, e92331. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Shao, Y.; Qin, X.; Liu, D.; Chen, A.; Li, D.; Liu, M.; Wang, X. CRISPR knockout rat cytochrome P450 3A1/2 model for advancing drug metabolism and pharmacokinetics research. Sci. Rep. 2017, 7, 42922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ossart, J.; Moreau, A.; Autrusseau, E.; Ménoret, S.; Martin, J.C.; Besnard, M.; Ouisse, L.H.; Tesson, L.; Flippe, L.; Kisand, K.; et al. Breakdown of immune tolerance in AIRE-deficient rats induces a severe autoimmune polyendocrinopathy–candiasis–ectodermal dystrophy–like autoimmune disease. J. Immunol. 2018, 201, 874–887. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Jiang, X.; Xu, M.; Wang, H.; Sang, X.; Qin, M.; Bao, P.; Wang, R.; Zhang, C.; Lu, H.; et al. Sleep and circadian abnormalities precede cognitive deficits in R521C FUS knockin rats. Neurobiol. Aging 2018, 72, 159–170. [Google Scholar] [CrossRef]
- Ménoret, S.; Ouisse, L.-H.; Tesson, L.; Delbos, F.; Garnier, D.; Remy, S.; Usal, C.; Concordet, J.P.; Giovannangeli, C.; Chenouard, V.; et al. Generation of immunodeficient rats with Rag1 and Il2rg gene deletions and human tissue grafting models. Transplantation 2018, 102, 1271–1278. [Google Scholar] [CrossRef]
- Izumi, R.; Kusakabe, T.; Noguchi, M.; Iwakura, H.; Tanaka, T.; Miyazawa, T.; Aotani, D.; Hosoda, K.; Kangawa, K.; Nakao, K. CRISPR/Cas9-mediated Angptl8 knockout suppresses plasma triglyceride concentrations and adiposity in rats. J. Lipid Res. 2018, 59, 1575–1585. [Google Scholar] [CrossRef] [Green Version]
- Takabayashi, S.; Aoshima, T.; Kabashima, K.; Aoto, K.; Ohtsuka, M.; Sato, M. i-GONAD (improved genome-editing via oviductal nucleic acids delivery), a convenient in vivo tool to produce genome-edited rats. Sci. Rep. 2018, 8, 12059. [Google Scholar] [CrossRef]
- Kobayashi, T.; Namba, M.; Koyano, T.; Fukushima, M.; Sato, M.; Ohtsuka, M.; Matsuyama, M. Successful production of genome-edited rats by the rGONAD method. BMC Biotechnol. 2018, 18, 19. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Yang, Y.; Xing, R.; Cui, X.; Xiao, Y.; Xie, L.; You, P.; Wang, T.; Zeng, L.; Peng, W.; et al. Hyperlipidemia induces typical atherosclerosis development in Ldlr and Apoe deficient rats. Atherosclerosis 2018, 271, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Wang, L.; Guo, N.; Wang, S.; Yang, L.; Li, Y.; Wang, M.; Yin, S.; Han, H.; Zeng, L.; et al. Cas9-nickase-mediated genome editing corrects hereditary tyrosinemia in rats. J. Biol. Chem. 2018, 293, 6883–6892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, R.; Fang, X.; He, L.; Shao, Y.; Guo, N.; Wang, L.; Liu, M.; Li, D.; Geng, H. Generation of a primary hyperoxaluria type 1 disease model via CRISPR/Cas9 system in rats. Curr. Mol. Med. 2018, 18, 436–447. [Google Scholar] [CrossRef]
- Miyasaka, Y.; Uno, Y.; Yoshimi, K.; Kunihiro, Y.; Yoshimura, T.; Tanaka, T.; Ishikubo, H.; Hiraoka, Y.; Takemoto, N.; Tanaka, T.; et al. CLICK: One-step generation of conditional knockout mice. BMC Genom. 2018, 19, 318. [Google Scholar] [CrossRef] [Green Version]
- Sinkevicius, K.W.; Morrison, T.R.; Kulkarni, P.; Cagliostro, M.K.C.; Iriah, S.; Malmberg, S.; Sabrick, J.; Honeycutt, J.A.; Askew, K.L.; Trivedi, M.; et al. RNaseT2 knockout rats exhibit hippocampal neuropathology and deficits in memory. Dis. Models Mech. 2018, 11, dmm032631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Zhang, X.; Wang, L.; Yin, S.; Zhu, B.; Xie, L.; Duan, Q.; Hu, H.; Zheng, R.; Wei, Y.; et al. Increasing targeting scope of adenosine base editors in mouse and rat embryos through fusion of TadA deaminase with Cas9 variants. Protein Cell 2018, 9, 814–819. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.J.; Aoki, K.; Moehring, F.; Murphy, C.A.; O’Hara, C.L.; Tiemeyer, M.; Stucky, C.L.; Dahms, N.M. Neuropathic pain in a Fabry disease rat model. JCI Insight 2018, 3, e99171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Z.; Li, L.; Li, X.; Zhang, M.; Zhong, Y.; Li, Y.; Yu, D.; Cao, J.; Zhao, J.; Xiaoming, D.; et al. Generation of an oxoglutarate dehydrogenase knockout rat model and the effect of a high-fat diet. RSC Adv. 2018, 8, 16636–16644. [Google Scholar] [CrossRef] [Green Version]
- Leo, D.; Sukhanov, I.; Zoratto, F.; Illiano, P.; Caffino, L.; Sanna, F.; Messa, G.; Emanuele, M.; Esposito, A.; Dorofeikova, M.; et al. Pronounced hyperactivity, cognitive dysfunctions, and BDNF dysregulation in dopamine transporter knock-out rats. J. Neurosci. 2018, 38, 1959–1972. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Zhang, L.; Xie, M.; Li, Y.; Huang, P.; Saunders, T.L.; Fox, D.A.; Rosenquist, R.; Lin, F. Role of complement in a rat model of paclitaxel-induced peripheral neuropathy. J. Immunol. 2018, 200, 4094–4101. [Google Scholar] [CrossRef]
- Noto, F.K.; Adjan-Steffey, V.; Tong, M.; Ravichandran, K.; Zhang, W.; Arey, A.; McClain, C.B.; Ostertag, E.; Mazhar, S.; Sangodkar, J.; et al. Sprague dawley Rag2-null rats created from engineered spermatogonial stem cells are immunodeficient and permissive to human xenografts. Mol. Cancer Ther. 2018, 17, 2481–2489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Zhou, Y.; Xie, S.; Yin, Q.; Tang, C.; Ni, Z.; Fei, J.; Zhang, Y. CRISPR/Cas9-mediated genome editing reveals the synergistic effects of β-defensin family members on sperm maturation in rat epididymis. FASEB J. 2018, 32, 1354–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Yu, L.; Zhang, X.; Xin, C.; Huang, S.; Bai, L.; Chen, W.; Gao, R.; Li, J.; Pan, S.; et al. Highly efficient and precise base editing by engineered dCas9-guide tRNA adenosine deaminase in rats. Cell Discov. 2018, 4, 39. [Google Scholar] [CrossRef]
- Wei, Y.; Yang, L.; Zhang, X.; Sui, D.; Wang, C.; Wang, K.; Shan, M.; Guo, D.; Wang, H. Generation and characterization of a CYP2C11-null rat model by using the CRISPR/Cas9 method. Drug Metab. Dispos. 2018, 46, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Dreano, E.; Bacchetta, M.; Simonin, J.; Galmiche, L.; Usal, C.; Slimani, L.; Sadoine, J.; Tesson, L.; Anegon, I.; Concordet, J.P.; et al. Characterization of two rat models of cystic fibrosis—KO and F508del CFTR—Generated by Crispr-Cas9. Anim. Models Exp. Med. 2019, 2, 297–311. [Google Scholar] [CrossRef]
- Miller, J.J.; Aoki, K.; Mascari, C.A.; Beltrame, A.K.; Sokumbi, O.; North, P.E.; Tiemeyer, M.; Kriegel, A.J.; Dahms, N.M. α-Galactosidase A-deficient rats accumulate glycosphingolipids and develop cardiorenal phenotypes of Fabry disease. FASEB J. 2019, 33, 418–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, T.J.; Lan, X.Y.; Wei, M.P.; Zhai, F.J.; Boeckers, T.M.; Wang, J.N.; Yuan, S.; Jin, M.Y.; Xie, Y.F.; Dang, W.W.; et al. Altered behaviors and impaired synaptic function in a novel rat model with a complete Shank3 deletion. Front. Cell. Neurosci. 2019, 13, 111. [Google Scholar] [CrossRef]
- Lambert, M.; Capuano, V.; Boet, A.; Tesson, L.; Bertero, T.; Nakhleh, M.K.; Remy, S.; Anegon, I.; Pechoux, C.; Hautefort, A.; et al. Characterization of Kcnk3-mutated rat, a novel model of pulmonary hypertension. Circ. Res. 2019, 125, 678–695. [Google Scholar] [CrossRef]
- Lee, J.G.; Ha, C.H.; Yoon, B.; Cheong, S.A.; Kim, G.; Lee, D.J.; Woo, D.C.; Kim, Y.H.; Nam, S.Y.; Lee, S.W.; et al. Knockout rat models mimicking human atherosclerosis created by Cpf1-mediated gene targeting. Sci. Rep. 2019, 9, 2628. [Google Scholar] [CrossRef] [Green Version]
- Honda, A.; Tachibana, R.; Hamada, K.; Morita, K.; Mizuno, N.; Morita, K.; Asano, M. Efficient derivation of knock-out and knock-in rats using embryos obtained by in vitro fertilization. Sci. Rep. 2019, 9, 11571. [Google Scholar] [CrossRef]
- Liang, C.; Zhao, J.; Lu, J.; Zhang, Y.; Ma, X.; Shang, X.; Li, Y.; Ma, X.; Liu, M.; Wang, X. Development and characterization of MDR1 (Mdr1a/b) CRISPR/Cas9 knockout rat model. Drug Metab. Dispos. 2019, 47, 71–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emmert, A.S.; Vuong, S.M.; Shula, C.; Lindquist, D.; Yuan, W.; Hu, Y.C.; Mangano, F.T.; Goto, J. Characterization of a novel rat model of X-linked hydrocephalus by CRISPR-mediated mutation in L1cam. J. Neurosurg. 2019, 132, 945–958. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wu, Z.; Liu, L.; Liu, J.; Wang, Y. Rat model of Cockayne syndrome neurological disease. Cell Rep. 2019, 29, 800–809.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenawi, M.; Rouger, E.; Island, M.L.; Leroyer, P.; Robin, F.; Rémy, S.; Tesson, L.; Anegon, I.; Nay, K.; Derbré, F.; et al. Ceruloplasmin deficiency does not induce macrophagic iron overload: Lessons from a new rat model of hereditary aceruloplasminemia. FASEB J. 2019, 33, 13492–13502. [Google Scholar] [CrossRef] [Green Version]
- Hautefort, A.; Mendes-Ferreira, P.; Sabourin, J.; Manaud, G.; Bertero, T.; Rucker-Martin, C.; Riou, M.; Adão, R.; Manoury, B.; Lambert, M.; et al. Bmpr2 mutant rats develop pulmonary and cardiac characteristics of pulmonary arterial hypertension. Circulation 2019, 139, 932–948. [Google Scholar] [CrossRef]
- Demireva, E.Y.; Xie, H.; Flood, E.D.; Thompson, J.M.; Seitz, B.M.; Watts, S.W. Creation of the 5-hydroxytryptamine receptor 7 knockout rat as a tool for cardiovascular research. Physiol. Genom. 2019, 51, 290–301. [Google Scholar] [CrossRef]
- Scheimann, J.R.; Moloney, R.D.; Mahbod, P.; Morano, R.L.; Fitzgerald, M.; Hoskins, O.; Packard, B.A.; Cotella, E.M.; Hu, Y.C.; Herman, J.P. Conditional deletion of glucocorticoid receptors in rat brain results in sex-specific deficits in fear and coping behaviors. eLife 2019, 8, e44672. [Google Scholar] [CrossRef]
- Pettibone, J.R.; Yu, J.Y.; Derman, R.C.; Faust, T.W.; Hughes, E.D.; Filipiak, W.E.; Saunders, T.L.; Ferrario, C.R.; Berke, J.D. Knock-in rat lines with Cre recombinase at the dopamine D1 and adenosine 2a receptor loci. eNeuro 2019, 6, ENEURO.0163-19.2019. [Google Scholar] [CrossRef] [Green Version]
- Shi, Q.; Mattson, J.G.; Fahs, S.A.; Geurts, A.M.; Weiler, H.; Montgomery, R.R. The severe spontaneous bleeding phenotype in a novel hemophilia A rat model is rescued by platelet FVIII expression. Blood Adv. 2020, 4, 55–65. [Google Scholar] [CrossRef]
- Takabayashi, S.; Aoshima, T.; Kobayashi, Y.; Takagi, H.; Akasaka, E.; Sato, M. Successful i-GONAD in Brown Norway rats by modification of in vivo electroporation conditions. OBM Genet. 2020, 4, 9. [Google Scholar] [CrossRef]
- Dodge, A.; Peters, M.M.; Greene, H.E.; Dietrick, C.; Botelho, R.; Chung, D.; Willman, J.; Nenninger, A.W.; Ciarlone, S.; Kamath, S.G.; et al. Generation of a novel rat model of Angelman syndrome with a complete Ube3a gene deletion. Autism Res. 2020, 13, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhao, X.M.; Liu, J.; Wang, Y.Y.; Xiong, L.L.; He, X.Y.; Wang, T.H. Complexin I knockout rats exhibit a complex neurobehavioral phenotype including profound ataxia and marked deficits in lifespan. Pflugers Arch. 2020, 472, 117–133. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Schiel, J.A.; Maksimova, E.; Strezoska, Z.; Zhao, G.; Anderson, E.M.; Wu, Y.; Warren, J.; Bartels, A.; van Brabant Smith, A.; et al. ErCas12a CRISPR-MAD7 for model generation in human cells, mice, and rats. CRISPR J. 2020, 3, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Fujihara, K.; Yamada, K.; Ichitani, Y.; Kakizaki, T.; Jiang, W.; Miyata, S.; Suto, T.; Kato, D.; Saito, S.; Watanabe, M.; et al. CRISPR/Cas9-engineered Gad1 elimination in rats leads to complex behavioral changes: Implications for schizophrenia. Transl. Psychiatry 2020, 10, 426. [Google Scholar] [CrossRef]
- Langer, H.T.; Mossakowski, A.A.; Willis, B.J.; Grimsrud, K.N.; Wood, J.A.; Lloyd, K.; Zbinden-Foncea, H.; Baar, K. Generation of desminopathy in rats using CRISPR-Cas9. J. Cachexia Sarcopenia Muscle 2020, 11, 1364–1376. [Google Scholar] [CrossRef]
- Zheng, R.; Li, Y.; Wang, L.; Fang, X.; Zhang, J.; He, L.; Yang, L.; Li, D.; Geng, H. CRISPR/Cas9–mediated metabolic pathway reprogramming in a novel humanized rat model ameliorates primary hyperoxaluria type 1. Kidney Int. 2020, 98, 947–957. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, N.; Dong, X.; Fan, N.; Wang, L.; Xu, Y.; Chen, H.; Duan, W. Uricase-deficient rat is generated with CRISPR/Cas9 technique. PeerJ 2020, 8, e8971. [Google Scholar] [CrossRef]
- Nishikawa, M.; Yasuda, K.; Takamatsu, M.; Abe, K.; Okamoto, K.; Horibe, K.; Mano, H.; Nakagawa, K.; Tsugawa, N.; Hirota, Y.; et al. Generation of novel genetically modified rats to reveal the molecular mechanisms of vitamin D actions. Sci. Rep. 2020, 10, 5677. [Google Scholar] [CrossRef]
- Birket, S.E.; Davis, J.M.; Fernandez-Petty, C.M.; Henderson, A.G.; Oden, A.M.; Tang, L.; Wen, H.; Hong, J.; Fu, L.; Chambers, A.; et al. Ivacaftor reverses airway mucus abnormalities in a rat model harboring a humanized G551D-CFTR. Am. J. Respir. Crit. Care Med. 2020, 202, 1271–1282. [Google Scholar] [CrossRef]
- Bae, H.S.; Jin, Y.K.; Ham, S.; Kim, H.K.; Shin, H.; Cho, G.B.; Lee, K.J.; Lee, H.; Kim, K.M.; Koo, O.J.; et al. CRISRP/Cas9-mediated knockout of Mct8 reveals a functional involvement of Mct8 in testis and sperm development in a rat. Sci. Rep. 2020, 10, 11148. [Google Scholar] [CrossRef]
- Sanna, F.; Bratzu, J.; Serra, M.P.; Leo, D.; Quartu, M.; Boi, M.; Espinoza, S.; Gainetdinov, R.R.; Melis, M.R.; Argiolas, A. Altered sexual behavior in dopamine transporter (DAT) knockout male rats: A behavioral, neurochemical and intracerebral microdialysis study. Front. Behav. Neurosci. 2020, 14, 58. [Google Scholar] [CrossRef] [PubMed]
- Reese, R.M.; Dourado, M.; Anderson, K.; Warming, S.; Stark, K.L.; Balestrini, A.; Suto, E.; Lee, W.; Riol-Blanco, L.; Shields, S.D.; et al. Behavioral characterization of a CRISPR-generated TRPA1 knockout rat in models of pain, itch, and asthma. Sci. Rep. 2020, 10, 979. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Chen, A.; Ma, X.; Shang, X.; Zhang, Y.; Guo, Y.; Liu, M.; Wang, X. Generation and characterization of cytochrome P450 2J3/10 CRISPR/Cas9 knockout rat model. Drug Metab. Dispos. 2020, 48, 1129–1136. [Google Scholar] [CrossRef] [PubMed]
- Konishi, S.; Tanaka, N.; Mashimo, T.; Yamamoto, T.; Sakuma, T.; Kaneko, T.; Tanaka, M.; Izawa, T.; Yamate, J.; Kuwamura, M. Pathological characteristics of Ccdc85c knockout rats: A rat model of genetic hydrocephalus. Exp. Anim. 2020, 69, 26–33. [Google Scholar] [CrossRef]
- Tambini, M.D.; D’Adamio, L. Knock-in rats with homozygous PSEN1L435F Alzheimer mutation are viable and show selective γ-secretase activity loss causing low Aβ40/42 and high Aβ43. J. Biol. Chem. 2020, 295, 7442–7451. [Google Scholar] [CrossRef] [Green Version]
- Dong, W.; Zhang, L.; Sun, C.; Gao, X.; Guan, F.; Li, J.; Chen, W.; Ma, Y.; Zhang, L. Knock in of a hexanucleotide repeat expansion in the C9orf72 gene induces ALS in rats. Anim. Models Exp. Med. 2020, 3, 237–244. [Google Scholar] [CrossRef]
- Kitagawa, A.; Kizub, I.; Jacob, C.; Michael, K.; D’Alessandro, A.; Reisz, J.A.; Grzybowski, M.; Geurts, A.M.; Rocic, P.; Gupte, R.; et al. CRISPR-mediated single nucleotide polymorphism modeling in rats reveals insight into reduced cardiovascular risk associated with Mediterranean G6PD variant. Hypertension 2020, 76, 523–532. [Google Scholar] [CrossRef]
- Ma, X.; Shang, X.; Qin, X.; Lu, J.; Liu, M.; Wang, X. Characterization of organic anion transporting polypeptide 1b2 knockout rats generated by CRISPR/Cas9: A novel model for drug transport and hyperbilirubinemia disease. Acta Pharm. Sin. B 2020, 10, 850–860. [Google Scholar] [CrossRef]
- Yoshimi, K.; Oka, Y.; Miyasaka, Y.; Kotani, Y.; Yasumura, M.; Uno, Y.; Hattori, K.; Tanigawa, A.; Sato, M.; Oya, M.; et al. Combi-CRISPR: Combination of NHEJ and HDR provides efficient and precise plasmid-based knock-ins in mice and rats. Hum. Genet. 2021, 140, 277–287. [Google Scholar] [CrossRef]
- Aoshima, T.; Kobayashi, Y.; Takagi, H.; Iijima, K.; Sato, M.; Takabayashi, S. Modification of improved-genome editing via oviductal nucleic acids delivery (i-GONAD)-mediated knock-in in rats. BMC Biotechnol. 2021, 21, 63. [Google Scholar] [CrossRef]
- Koster, C.; van den Hurk, K.T.; Lewallen, C.F.; Talib, M.; Ten Brink, J.B.; Boon, C.; Bergen, A.A. The Lrat−/− rat: CRISPR/Cas9 construction and phenotyping of a new animal model for retinitis pigmentosa. Int. J. Mol. Sci. 2021, 22, 7234. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, K.; Nishikawa, M.; Okamoto, K.; Horibe, K.; Mano, H.; Yamaguchi, M.; Okon, R.; Nakagawa, K.; Tsugawa, N.; Okano, T.; et al. Elucidation of metabolic pathways of 25-hydroxyvitamin D3 mediated by CYP24A1 and CYP3A using Cyp24a1 knockout rats generated by CRISPR/Cas9 system. J. Biol. Chem. 2021, 296, 100668. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Kim, U.; Yang, H.; Ryu, B.; Kim, J.; Sakuma, T.; Yamamoto, T.; Park, J.H. TALEN-mediated generation of Nkx3.1 knockout rat model. Prostate 2021, 81, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, K.; Kadekawa, K.; Nishijima, S.; Sakanashi, M.; Okitsu-Sakurayama, S.; Higa-Nakamine, S.; Yamamoto, H.; Sugaya, K. Phenotypic characterization of the endocannabinoid-degrading enzyme alpha/beta-hydrolase domain 6 knockout rat. Cannabis Cannabinoid Res. 2021. [Google Scholar] [CrossRef]
- Miller, B.; Kostrominova, T.Y.; Geurts, A.M.; Sorokin, A. Double p52Shc/p46Shc rat knockout demonstrates severe gait abnormalities accompanied by dilated cardiomyopathy. Int. J. Mol. Sci. 2021, 22, 5237. [Google Scholar] [CrossRef]
- De Souza Gonçalves, L.; Sales, L.P.; Saito, T.R.; Campos, J.C.; Fernandes, A.L.; Natali, J.; Jensen, L.; Arnold, A.; Ramalho, L.; Bechara, L.; et al. Histidine dipeptides are key regulators of excitation-contraction coupling in cardiac muscle: Evidence from a novel CARNS1 knockout rat model. Redox Biol. 2021, 44, 102016. [Google Scholar] [CrossRef]
- Soga, M.; Izumi, T.; Nanchi, I.; Horita, N.; Yamamoto, M.; Kawasaki, S.; Ogawa, K.; Fujita, M.; Morioka, Y. Suppression of joint pain in transient receptor potential vanilloid 4 knockout rats with monoiodoacetate-induced osteoarthritis. Pain Rep. 2021, 6, e951. [Google Scholar] [CrossRef]
- Liu, J.; Shang, X.; Huang, S.; Xu, Y.; Lu, J.; Zhang, Y.; Liu, Z.; Wang, X. Construction and characterization of CRISPR/Cas9 knockout rat model of carboxylesterase 2a gene. Mol. Pharmacol. 2021, 100, 480–490. [Google Scholar] [CrossRef]
- Zhang, H.; Zheng, Q.; Chen-Tsai, R.Y. Establishment of a Cre-rat resource for creating conditional and physiological relevant models of human diseases. Transgenic Res. 2021, 30, 91–104. [Google Scholar] [CrossRef]
- Chesney, K.L.; Men, H.; Hankins, M.A.; Bryda, E.C. The Atg16l1 gene: Characterization of wild type, knock-in, and knock-out phenotypes in rats. Physiol. Genom. 2021, 53, 269–281. [Google Scholar] [CrossRef]
- Chen, L.; Ouyang, J.; Li, X.; Xiao, X.; Sun, W.; Li, S.; Zhou, L.; Liao, Y.; Zhang, Q. DNAH17 is essential for rat spermatogenesis and fertility. J. Genet. 2021, 100, 14. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Chen, X.; Guo, J.; Zhang, X.; Sun, H.; Wang, J.; Qian, X.; Li, B.; Tan, L.; Yu, L.; et al. Precision modeling of mitochondrial disease in rats via DdCBE-mediated mtDNA editing. Cell Discov. 2021, 7, 95. [Google Scholar] [CrossRef] [PubMed]
- Gordon, J.W.; Scangos, G.A.; Plotkin, D.J.; Barbosa, J.A.; Ruddle, F.H. Genetic transformation of mouse embryos by microinjection of purified DNA. Proc. Natl. Acad. Sci. USA 1980, 77, 7380–7384. [Google Scholar] [CrossRef] [Green Version]
- Clark, J.F.; Dinsmore, C.J.; Soriano, P. A most formidable arsenal: Genetic technologies for building a better mouse. Genes Dev. 2020, 34, 1256–1286. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yang, H.; Shivalila, C.S.; Dawlaty, M.M.; Cheng, A.W.; Zhang, F.; Jaenisch, R. One-step generation of mice carrying mutations in multiple genes by CRISPR/Cas-mediated genome engineering. Cell 2013, 153, 910–918. [Google Scholar] [CrossRef] [Green Version]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex genome engineering using CRISPR/Cas systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [Green Version]
- Sato, M.; Takabayashi, S.; Akasaka, E.; Nakamura, S. Recent advances and future perspectives of in vivo targeted delivery of genome-editing reagents to germ cells, embryos, and fetuses in mice. Cells 2020, 9, 799. [Google Scholar] [CrossRef] [Green Version]
- Shao, Y.; Guan, Y.; Wang, L.; Qiu, Z.; Liu, M.; Chen, Y.; Wu, L.; Li, Y.; Ma, X.; Liu, M.; et al. CRISPR/Cas-mediated genome editing in the rat via direct injection of one-cell embryos. Nat. Protoc. 2014, 9, 2493–2512. [Google Scholar] [CrossRef]
- Ménoret, S.; Tesson, L.; Rémy, S.; Usal, C.; Thépenier, V.; Thinard, R.; Ouisse, L.H.; De Cian, A.; Giovannangeli, C.; Concordet, J.P.; et al. Gene targeting in rats using transcription activator-like effector nucleases. Methods 2014, 69, 102–107. [Google Scholar] [CrossRef]
- Tesson, L.; Remy, S.; Ménoret, S.; Usal, C.; Thinard, R.; Savignard, C.; De Cian, A.; Giovannangeli, C.; Concordet, J.P.; Anegon, I. Genome editing in rats using TALE nucleases. Methods Mol. Biol. 2016, 1338, 245–259. [Google Scholar] [CrossRef]
- Young, J.L.; Dean, D.A. Electroporation-mediated gene delivery. Adv. Genet. 2015, 89, 49–88. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Takemoto, T. Electroporation enables the efficient mRNA delivery into the mouse zygotes and facilitates CRISPR/Cas9-based genome editing. Sci. Rep. 2015, 5, 11315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, W.; Dion, S.L.; Kutny, P.M.; Zhang, Y.; Cheng, A.W.; Jillette, N.L.; Malhotra, A.; Geurts, A.M.; Chen, Y.G.; Wang, H. Efficient CRISPR/Cas9-mediated genome editing in mice by zygote electroporation of nuclease. Genetics 2015, 200, 423–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Lee, B.; Lee, A.Y.; Modzelewski, A.J.; He, L. Highly efficient mouse genome editing by CRISPR ribonucleoprotein electroporation of zygotes. J. Biol. Chem. 2016, 291, 14457–14467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tröder, S.E.; Ebert, L.K.; Butt, L.; Assenmacher, S.; Schermer, B.; Zevnik, B. An optimized electroporation approach for efficient CRISPR/Cas9 genome editing in murine zygotes. PLoS ONE 2018, 13, e0196891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, M.; Py, B.F.; Bosc, C.; Laubreton, D.; Moutin, M.J.; Marvel, J.; Flamant, F.; Markossian, S. Electroporation of mice zygotes with dual guide RNA/Cas9 complexes for simple and efficient cloning-free genome editing. Sci. Rep. 2018, 8, 474. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Sun, S.; Moonen, D.; Lee, C.; Lee, A.Y.; Schaffer, D.V.; He, L. CRISPR-READI: Efficient generation of knockin mice by CRISPR RNP electroporation and AAV donor infection. Cell Rep. 2019, 27, 3780–3789.e4. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.; Gaynor, P.; Cole, S.; Brophy, B.; Oback, B.; Laible, G. Developing the laboratory conditions for bovine zygote-mediated genome editing by electroporation. In Proceedings of the World Congress on Genetics Applied to Livestock Production; 2018. Available online: http://www.wcgalp.org/system/files/proceedings/2018/developing-condition-bovine-zygote-mediated-genome-editing-electroporation.pdf (accessed on 30 January 2022).
- Tanihara, F.; Takemoto, T.; Kitagawa, E.; Rao, S.; Do, L.T.; Onishi, A.; Yamashita, Y.; Kosugi, C.; Suzuki, H.; Sembon, S.; et al. Somatic cell reprogramming-free generation of genetically modified pigs. Sci. Adv. 2016, 2, e1600803. [Google Scholar] [CrossRef] [Green Version]
- Tanihara, F.; Hirata, M.; Nguyen, N.T.; Le, Q.A.; Wittayarat, M.; Fahrudin, M.; Hirano, T.; Otoi, T. Generation of CD163-edited pig via electroporation of the CRISPR/Cas9 system into porcine in vitro-fertilized zygotes. Anim. Biotechnol. 2021, 32, 147–154. [Google Scholar] [CrossRef]
- Sato, M.; Jin, H.; Akasaka, E.; Miyoshi, M. In vitro electroporation in the presence of CRISPR/Cas9 reagents as a safe and useful method for producing biallelic knock out porcine embryos. OBM Genet. 2021, 5, 15. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Sakuma, T.; Takeo, T.; Nakagata, N.; Yamamoto, T. Electroporation-mediated genome editing in vitrified/warmed mouse zygotes created by IVF via ultra-superovulation. Exp. Anim. 2018, 67, 535–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneko, T.; Nakagawa, Y. Genome editing of rodents by electroporation of CRISPR/Cas9 into frozen-warmed pronuclear-stage embryos. Cryobiology 2020, 92, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, D.; Cho, S.W.; Kim, J.; Kim, J.S. Highly efficient RNA-guided genome editing in human cells via delivery of purified Cas9 ribonucleoproteins. Genome Res. 2014, 24, 1012–1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakim, B.A.; Tyagi, V.; Agnihotri, S.K.; Amar Nath, A.; Agrawal, A.K.; Jain, A.; Singh, D.; Konwar, R.; Sachdev, M. Electroporation of mouse follicles, oocytes and embryos without manipulating zona pellucida. J. Dev. Biol. 2021, 9, 13. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, H.; Friedman, H.; Shao, H.; Chong, Y.; Lo, C.A.; Emran, F.; Kays, I.; Yang, X.J.; Cooper, E.; Chen, B.E.; et al. TIE: A method to electroporate long DNA templates into preimplantation embryos for CRISPR-Cas9 gene editing. CRISPR J. 2018, 1, 223–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneko, T. Genome editing of rat. Methods Mol. Biol. 2017, 1630, 101–108. [Google Scholar] [CrossRef]
- Remy, S.; Chenouard, V.; Tesson, L.; Usal, C.; Ménoret, S.; Brusselle, L.; Heslan, J.M.; Nguyen, T.H.; Bellien, J.; Merot, J.; et al. Generation of gene-edited rats by delivery of CRISPR/Cas9 protein and donor DNA into intact zygotes using electroporation. Sci. Rep. 2017, 7, 16554. [Google Scholar] [CrossRef]
- Takahashi, G.; Gurumurthy, C.B.; Wada, K.; Miura, H.; Sato, M.; Ohtsuka, M. GONAD: Genome-editing via Oviductal Nucleic Acids Delivery system: A novel microinjection independent genome engineering method in mice. Sci. Rep. 2015, 5, 11406. [Google Scholar] [CrossRef] [Green Version]
- Ohtsuka, M.; Sato, M.; Miura, H.; Takabayashi, S.; Matsuyama, M.; Koyano, T.; Arifin, N.; Nakamura, S.; Wada, K.; Gurumurthy, C.B. i-GONAD: A robust method for in situ germline genome engineering using CRISPR nucleases. Genome Biol. 2018, 19, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirose, M.; Honda, A.; Fulka, H.; Tamura-Nakano, M.; Matoba, S.; Tomishima, T.; Mochida, K.; Hasegawa, A.; Nagashima, K.; Inoue, K.; et al. Acrosin is essential for sperm penetration through the zona pellucida in hamsters. Proc. Natl. Acad. Sci. USA 2020, 117, 2513–2518. [Google Scholar] [CrossRef]
- Gurumurthy, C.B.; Sato, M.; Nakamura, A.; Inui, M.; Kawano, N.; Islam, M.A.; Ogiwara, S.; Takabayashi, S.; Matsuyama, M.; Nakagawa, S.; et al. Creation of CRISPR-based germline-genome-engineered mice without ex vivo handling of zygotes by i-GONAD. Nat. Protoc. 2019, 14, 2452–2482. [Google Scholar] [CrossRef] [PubMed]
- Ohtsuka, M.; Sato, M. i-GONAD: A method for generating genome-edited animals without ex vivo handling of embryos. Dev. Growth Differ. 2019, 61, 306–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Namba, M.; Kobayashi, T.; Koyano, T.; Kohno, M.; Ohtsuka, M.; Matsuyama, M. GONAD: A new method for germline genome-editing in mice and rats. Dev. Growth Differ. 2021, 63, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Daya, S.; Berns, K.I. Gene therapy using adeno-associated virus vectors. Clin. Microbiol. Rev. 2008, 21, 583–593. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, N.; Mizutani, E.; Sato, H.; Kasai, M.; Ogawa, A.; Suchy, F.; Yamaguchi, T.; Nakauchi, H. Intra-embryo gene cassette knockin by CRISPR/Cas9-mediated genome editing with adeno-associated viral vector. iScience 2018, 9, 286–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, Y.; Wang, D.; Tai, P.W.L.; Riley, J.; Gao, G.; Rivera-Pérez, J.A. Streamlined ex vivo and in vivo genome editing in mouse embryos using recombinant adeno-associated viruses. Nat. Commun. 2018, 9, 412. [Google Scholar] [CrossRef]
- Edraki, A.; Mir, A.; Ibraheim, R.; Gainetdinov, I.; Yoon, Y.; Song, C.Q.; Cao, Y.; Gallant, J.; Xue, W.; Rivera-Pérez, J.A.; et al. A compact, high-accuracy Cas9 with a dinucleotide PAM for in vivo genome editing. Mol. Cell 2019, 73, 714–726.e4. [Google Scholar] [CrossRef] [Green Version]
- Romeo, C.; Chen, S.H.; Goulding, E.; Van Gorder, L.; Schwartz, M.; Walker, M.; Scott, G.; Scappini, E.; Ray, M.; Martin, N.P. AAV diffuses across zona pellucida for effortless gene delivery to fertilized eggs. Biochem. Biophys. Res. Commun. 2020, 526, 85–90. [Google Scholar] [CrossRef]
- Wang, D.; Niu, Y.; Ren, L.; Kang, Y.; Tai, P.W.L.; Si, C.; Mendonca, C.A.; Ma, H.; Gao, G.; Ji, W. Gene delivery to nonhuman primate preimplantation embryos using recombinant adeno-associated virus. Adv. Sci. 2019, 6, 1900440. [Google Scholar] [CrossRef]
- Cronkhite, J.T.; Norlander, C.; Furth, J.K.; Levan, G.; Garbers, D.L.; Hammer, R.E. Male and female germline specific expression of anEGFP reporter gene in a unique strain of transgenic rats. Dev. Biol. 2005, 284, 171–183. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Liang, D.; Wang, Y.; Bai, M.; Tang, W.; Bao, S.; Yan, Z.; Li, D.; Li, J. Correction of a genetic disease in mouse via use of CRISPR-Cas9. Cell Stem Cell 2013, 13, 659–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, W.; Kawasaki, K.; Sugiura, K.; Naito, K. Efficient generation of large-scale genome-modified mice using gRNA and CAS9 endonuclease. Nucleic Acids Res. 2013, 41, e187. [Google Scholar] [CrossRef] [PubMed]
- Brandl, C.; Ortiz, O.; Röttig, B.; Wefers, B.; Wurst, W.; Kühn, R. Creation of targeted genomic deletions using TALEN or CRISPR/Cas nuclease pairs in one-cell mouse embryos. FEBS Open Bio 2015, 5, 26–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birling, M.C.; Schaeffer, L.; André, P.; Lindner, L.; Maréchal, D.; Ayadi, A.; Sorg, T.; Pavlovic, G.; Hérault, Y. Efficient and rapid generation of large genomic variants in rats and mice using CRISMERE. Sci. Rep. 2017, 7, 43331. [Google Scholar] [CrossRef] [Green Version]
- Mao, Z.; Bozzella, M.; Seluanov, A.; Gorbunova, V. Comparison of nonhomologous end joining and homologous recombination in human cells. DNA Repair 2008, 7, 1765–1771. [Google Scholar] [CrossRef] [Green Version]
- Maruyama, T.; Dougan, S.K.; Truttmann, M.C.; Bilate, A.M.; Ingram, J.R.; Ploegh, H.L. Increasing the efficiency of precise genome editing with CRISPR-Cas9 by inhibition of nonhomologous end joining. Nat. Biotechnol. 2015, 33, 538–542. [Google Scholar] [CrossRef]
- Singh, P.; Schimenti, J.C.; Bolcun-Filas, E. A mouse geneticist’s practical guide to CRISPR applications. Genetics 2015, 199, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Chu, V.T.; Weber, T.; Wefers, B.; Wurst, W.; Sander, S.; Rajewsky, K.; Kühn, R. Increasing the efficiency of homology-directed repair for CRISPR-Cas9-induced precise gene editing in mammalian cells. Nat. Biotechnol. 2015, 33, 543–548. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Yang, D.; Xu, J.; Zhu, T.; Chen, Y.E.; Zhang, J. RS-1 enhances CRISPR/Cas9- and TALEN-mediated knock-in efficiency. Nat. Commun. 2016, 7, 10548. [Google Scholar] [CrossRef] [Green Version]
- Gerlach, M.; Kraft, T.; Brenner, B.; Petersen, B.; Niemann, H.; Montag, J. Efficient knock-in of a point mutation in porcine fibroblasts using the CRISPR/Cas9-GMNN fusion gene. Genes 2018, 9, 296. [Google Scholar] [CrossRef] [Green Version]
- Raveux, A.; Vandormael-Pournin, S.; Cohen-Tannoudji, M. Optimization of the production of knock-in alleles by CRISPR/Cas9 microinjection into the mouse zygote. Sci. Rep. 2017, 7, 42661. [Google Scholar] [CrossRef] [PubMed]
- Graf, R.; Li, X.; Chu, V.T.; Rajewsky, K. sgRNA sequence motifs blocking efficient CRISPR/Cas9-mediated gene editing. Cell Rep. 2019, 26, 1098–1103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labun, K.; Montague, T.G.; Gagnon, J.A.; Thyme, S.B.; Valen, E. CHOPCHOP v2: A web tool for the next generation of CRISPR genome engineering. Nucleic Acids Res. 2016, 44, W272–W276. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.W.; Kim, S.; Kim, J.M.; Kim, J.S. Targeted genome engineering in human cells with the Cas9 RNA-guided endonuclease. Nat. Biotechnol. 2013, 31, 230–232. [Google Scholar] [CrossRef] [PubMed]
- Mali, P.; Yang, L.; Esvelt, K.M.; Aach, J.; Guell, M.; DiCarlo, J.E.; Norville, J.E.; Church, G.M. RNA-guided human genome engineering via Cas9. Science 2013, 339, 823–826. [Google Scholar] [CrossRef] [Green Version]
- Komor, A.C.; Kim, Y.B.; Packer, M.S.; Zuris, J.A.; Liu, D.R. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature 2016, 533, 420–424. [Google Scholar] [CrossRef] [Green Version]
- Nishida, K.; Arazoe, T.; Yachie, N.; Banno, S.; Kakimoto, M.; Tabata, M.; Mochizuki, M.; Miyabe, A.; Araki, M.; Hara, K.Y.; et al. Targeted nucleotide editing using hybrid prokaryotic and vertebrate adaptive immune systems. Science 2016, 353, aaf8729. [Google Scholar] [CrossRef]
- Gaudelli, N.M.; Komor, A.C.; Rees, H.A.; Packer, M.S.; Badran, A.H.; Bryson, D.I.; Liu, D.R. Programmable base editing of A•T to G•C in genomic DNA without DNA cleavage. Nature 2017, 551, 464–471. [Google Scholar] [CrossRef]
- Ryu, S.M.; Koo, T.; Kim, K.; Lim, K.; Baek, G.; Kim, S.T.; Kim, H.S.; Kim, D.E.; Lee, H.; Chung, E.; et al. Adenine base editing in mouse embryos and an adult mouse model of Duchenne muscular dystrophy. Nat. Biotechnol. 2018, 36, 536–539. [Google Scholar] [CrossRef]
- Liu, Z.; Lu, Z.; Yang, G.; Huang, S.; Li, G.; Feng, S.; Liu, Y.; Li, J.; Yu, W.; Zhang, Y.; et al. Efficient generation of mouse models of human diseases via ABE- and BE-mediated base editing. Nat. Commun. 2018, 9, 2338. [Google Scholar] [CrossRef] [Green Version]
- Mok, B.Y.; de Moraes, M.H.; Zeng, J.; Bosch, D.E.; Kotrys, V.; Raguram, A.; Hsu, F.; Radey, M.C.; Peterson, S.B.; Mootha, V.K.; et al. A bacterial cytidine deaminase toxin enables CRISPR-free mitochondrial base editing. Nature 2020, 583, 631–637. [Google Scholar] [CrossRef]
- Asada, H.; Kawamura, Y.; Maruyama, K.; Kume, H.; Ding, R.G.; Kanbara, N.; Kuzume, H.; Sanbo, M.; Yagi, T.; Obata, K. Cleft palate and decreased brain gamma-aminobutyric acid in mice lacking the 67-kDa isoform of glutamic acid decarboxylase. Proc. Natl. Acad. Sci. USA 1997, 94, 6496–6499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chua, S.C., Jr.; Chung, W.K.; Wu-Peng, X.S.; Zhang, Y.; Liu, S.M.; Tartaglia, L.; Leibel, R.L. Phenotypes of mouse diabetes and rat fatty due to mutations in the OB (leptin) receptor. Science 1996, 271, 994–996. [Google Scholar] [CrossRef]
- Argilés, J.M. The obese Zucker rat: A choice for fat metabolism 1968-1988: Twenty years of research on the insights of the Zucker mutation. Prog. Lipid Res. 1989, 28, 53–66. [Google Scholar] [CrossRef]
- Wu, X.; Wakamiya, M.; Vaishnav, S.; Geske, R.; Montgomery, C., Jr.; Jones, P.; Bradley, A.; Caskey, C.T. Hyperuricemia and urate nephropathy in urate oxidase-deficient mice. Proc. Natl. Acad. Sci. USA 1994, 91, 742–746. [Google Scholar] [CrossRef] [Green Version]
- Nakade, S.; Yamamoto, T.; Sakuma, T. Cas9, Cpf1 and C2c1/2/3-What’s next? Bioengineered 2017, 8, 265–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anzalone, A.V.; Randolph, P.B.; Davis, J.R.; Sousa, A.A.; Koblan, L.W.; Levy, J.M.; Chen, P.J.; Wilson, C.; Newby, G.A.; Raguram, A.; et al. Search-and-replace genome editing without double-strand breaks or donor DNA. Nature 2019, 576, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, X.; He, S.; Huang, S.; Li, C.; Chen, Y.; Liu, Z.; Huang, X.; Wang, X. Efficient generation of mouse models with the prime editing system. Cell Discov. 2020, 6, 27. [Google Scholar] [CrossRef] [PubMed]
- Barbaric, I.; Gaynor Miller, G.; Dear, T.N. Appearances can be deceiving: Phenotypes of knockout mice. Brief. Funct. Genom. 2007, 6, 91–103. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Schematic illustration of genome-edited (GE) rat production through microinjection (MI) (A), in vitro electroporation (EP) (B), genome-editing via oviductal nucleic acids delivery (GONAD) (or improved genome-editing via oviductal nucleic acids delivery (i-GONAD)) (C), adeno-associated virus (AAV)-based GONAD, a technique enabling in vitro viral infection of zygotes with recombinant AAV (rAAV) (D), spermatogonial stem cell (SCC)-mediated transgenesis (E), and embryonic stem (ES) cell-mediated transgenesis (F). This figure was drawn in-house, based on the data shown in the paper of Sato et al. [158].
Figure 1.
Schematic illustration of genome-edited (GE) rat production through microinjection (MI) (A), in vitro electroporation (EP) (B), genome-editing via oviductal nucleic acids delivery (GONAD) (or improved genome-editing via oviductal nucleic acids delivery (i-GONAD)) (C), adeno-associated virus (AAV)-based GONAD, a technique enabling in vitro viral infection of zygotes with recombinant AAV (rAAV) (D), spermatogonial stem cell (SCC)-mediated transgenesis (E), and embryonic stem (ES) cell-mediated transgenesis (F). This figure was drawn in-house, based on the data shown in the paper of Sato et al. [158].

Figure 2.
Schematic illustration of single-stranded oligodeoxynucleotide (ssODN)-based deletion of a large fragment spanning a 7,098-bp endogenous retrovirus (ERV) element within the first intron of the Kit gene. This figure was drawn in-house, based on the data shown in the paper of Yoshimi et al. [46].
Figure 2.
Schematic illustration of single-stranded oligodeoxynucleotide (ssODN)-based deletion of a large fragment spanning a 7,098-bp endogenous retrovirus (ERV) element within the first intron of the Kit gene. This figure was drawn in-house, based on the data shown in the paper of Yoshimi et al. [46].

Figure 3.
Schematic representation of the two-hit by gRNA and two-oligo with plasmid (2H2OP) method for the production of CAG-GFP knock-in (KI) rats generated using the CRISPR/Cas9 system. In the first step, Cas9, together with two gRNAs targeting the rat Rosa26 locus and the CAG promoter in the GFP plasmids, cuts the target sites. In the second step, two ssODNs ligate each cut end to join the genomic DNA and the plasmid DNA via HDR. This figure was drawn in-house, based on the data shown in the paper of Yoshimi et al. [64]. Abbreviations: CAG, chicken β-actin-based promoter; GFP, green fluorescent protein; KI, knock-in; ssODNs, single-stranded oligodeoxynucleotides; pA, poly(A) sites.
Figure 3.
Schematic representation of the two-hit by gRNA and two-oligo with plasmid (2H2OP) method for the production of CAG-GFP knock-in (KI) rats generated using the CRISPR/Cas9 system. In the first step, Cas9, together with two gRNAs targeting the rat Rosa26 locus and the CAG promoter in the GFP plasmids, cuts the target sites. In the second step, two ssODNs ligate each cut end to join the genomic DNA and the plasmid DNA via HDR. This figure was drawn in-house, based on the data shown in the paper of Yoshimi et al. [64]. Abbreviations: CAG, chicken β-actin-based promoter; GFP, green fluorescent protein; KI, knock-in; ssODNs, single-stranded oligodeoxynucleotides; pA, poly(A) sites.

Figure 4.
Schematic representation of knock-in (KI) experiment in rats toward the mutated Tyr locus performed by Aoshima et al. [141]. The target sequence (exon 2 of Tyr) recognized by crRNA1, 2, and 3 is shown in green. The PAM sequences are underlined. Single-stranded oligodeoxynucleotide (ssODN) (containing wild-type nucleotide “G” that corresponds to mutated nucleotide “A”) is shown in orange below the target sequence. The nucleotide “A/T” marked in red is the mutation causative of the albino phenotype. This figure was drawn in-house, based on the data shown in the paper of Aoshima et al. [141].
Figure 4.
Schematic representation of knock-in (KI) experiment in rats toward the mutated Tyr locus performed by Aoshima et al. [141]. The target sequence (exon 2 of Tyr) recognized by crRNA1, 2, and 3 is shown in green. The PAM sequences are underlined. Single-stranded oligodeoxynucleotide (ssODN) (containing wild-type nucleotide “G” that corresponds to mutated nucleotide “A”) is shown in orange below the target sequence. The nucleotide “A/T” marked in red is the mutation causative of the albino phenotype. This figure was drawn in-house, based on the data shown in the paper of Aoshima et al. [141].


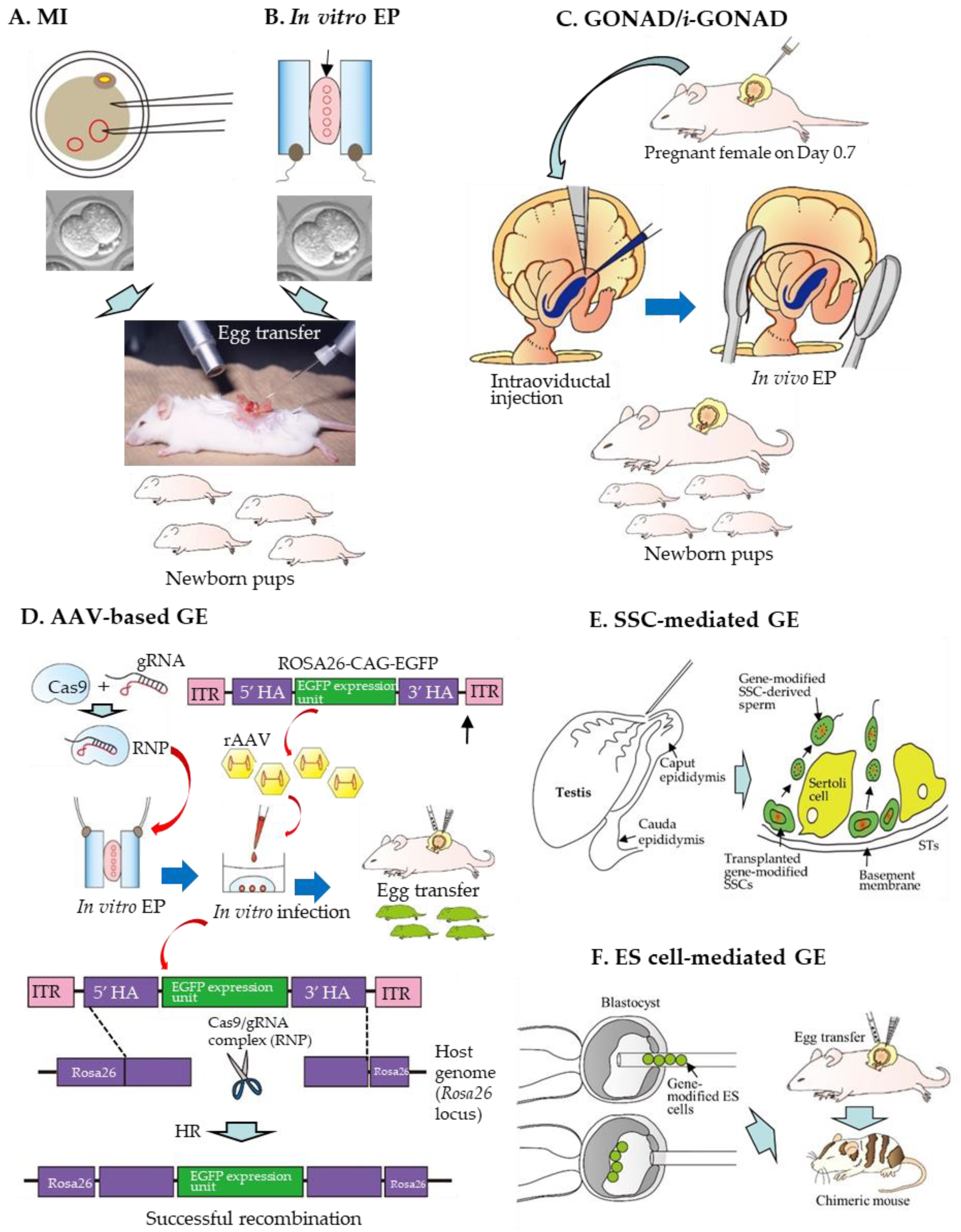{kind=link}
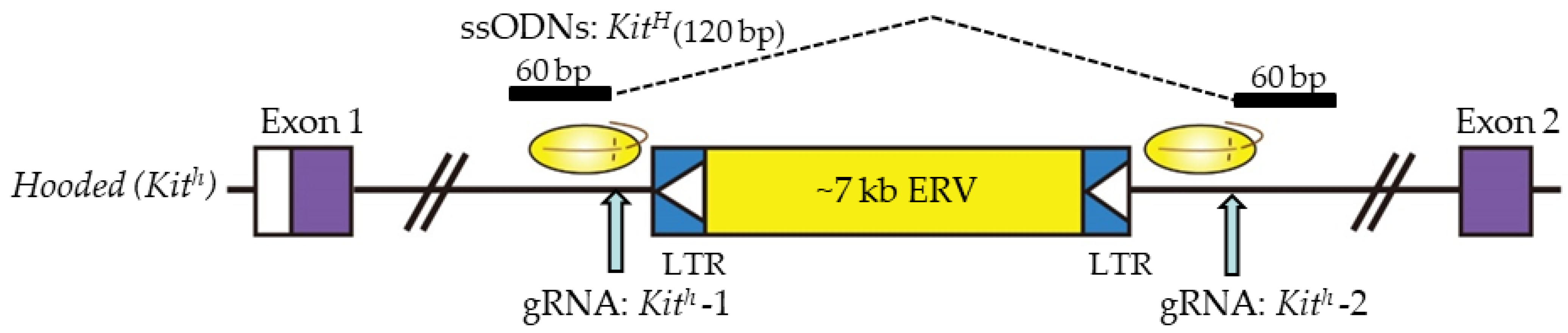{kind=link}
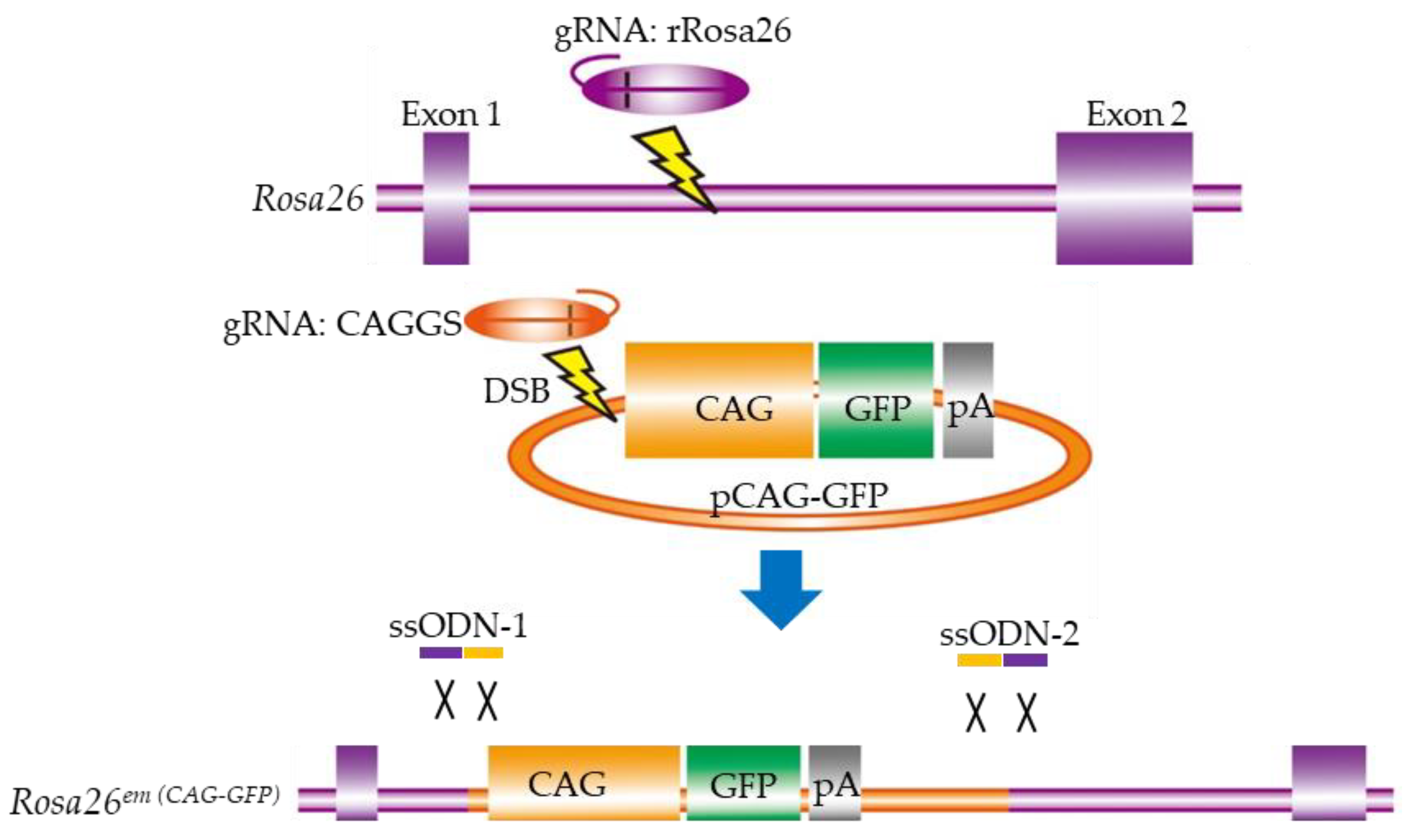{kind=link}
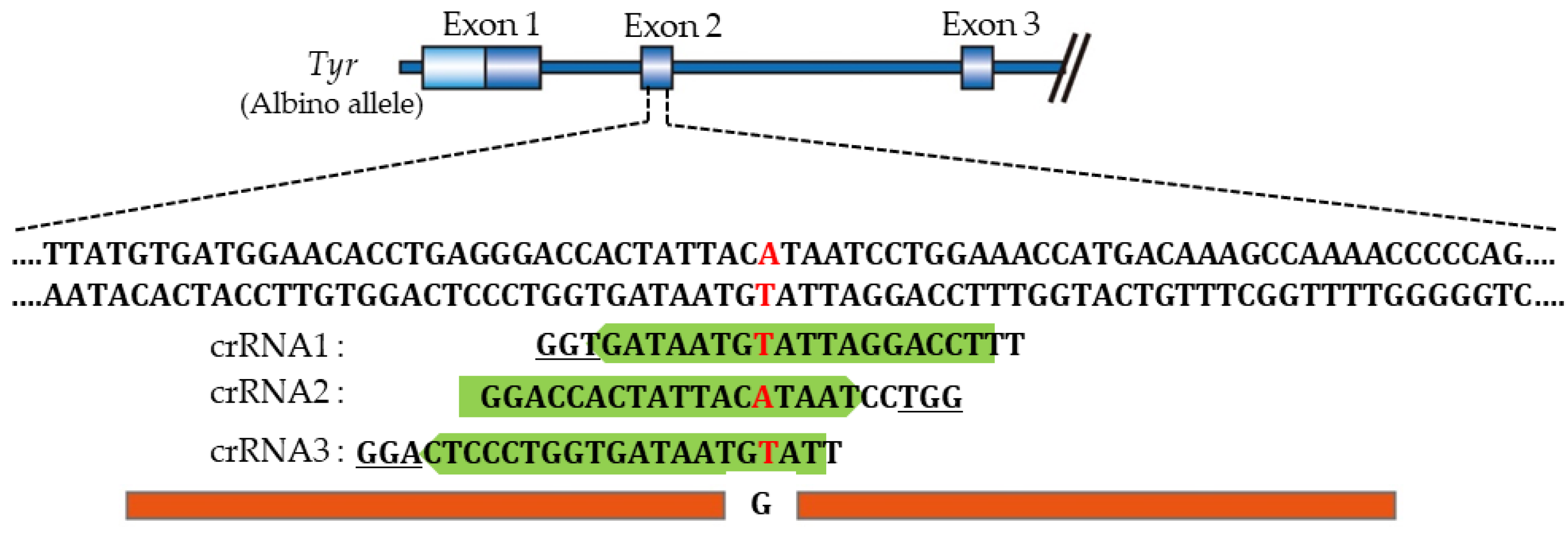{kind=link}
Table 1.
Summary of the production of genome-edited (GE) rats from 2009 to 2021.
| Method for Gene Modification | GE Tool (Mode for Gene Modification) | Rat Strain | Outcome | Target Gene | References |
|---|---|---|---|---|---|
| MI DNA or mRNA | ZFNs (indels) | SS FHH SD | Knockout (KO) rats first produced using ZFN technology with high frequency (25 to 100% disruption); showing successful germ-line transmission. | IgM Rab38 | Geurts et al. (2009) [28] |
| MI mRNA | ZFNs (indels) | F344/Stm TM/Kyo | KO rats generated as a model for X-linked severe combined immunodeficiency (X-SCID) with rates greater than 20%; showing successful germ-line transmission. | Il2rg | Mashimo et al. (2010) [29] |
| MI mRNA | ZFNs (indels) | SD | KO rats generated to show the absence of antibody-mediated hyperacute allograft rejection; can be used for biomedical research. | IgM | Ménoret et al. (2010) [30] |
| MI mRNA | ZFNs (indels) | SS | Renin-angiotensin system-related KO rats first produced. | Ren | Moreno et al. (2011) [31] |
| MI mRNA or plasmid DNA | TALENs (indels) | Unknown | KO rats first produced using TALEN technology. | IgM | Tesson et al. (2011) [32] |
| MI mRNA/plasmid DNA | ZFN (KI) | SD LEH | Knock-in (KI) rats first produced using ZFN technology; showing successful germ-line transmission of KI phenotype. | Mdr1a Pxr | Cui et al. (2011) [33] |
| MI mRNA | ZFNs (indels) | F344/Stm WI | KO rats lacking either DNA-dependent protein kinase catalytic subunit (Prkdc) or Prkdc and interleukin 2 receptor gamma (Il2rg) generated; double knockout (DKO) F344-scid gamma (FSG) rats showed more immunocompromised phenotypes, such as the abolishment of natural killer cells. | Prkdc Il2rg | Mashimo et al. (2012) [34] |
| MI plasmid DNA | Engineered meganuclease | SD | KO immunodeficient rats first produced using meganucleases; showing decreased numbers of immature and mature T and B lymphocytes and normal natural killer cells. | Rag1 | Ménoret et al. (2013) [35] |
| MI mRNA/plasmid DNA | ZFNs (KI) | SD | LoxP-flanked (floxed) rats first produced using ZFN technology; can be useful for Cre-dependent gene disruption in vivo. | Grin1 Crhr1 Tp53 | Brown et al. (2013) [36] |
| MI mRNA/sgRNAs | CRISPR/Cas9 (indels) | SD | KO rats first produced using CRISPR/Cas9 technology with multiple gene mutations (melanocortin 3 receptor (Mc3r) and melanocortin 4 receptor (Mc4r)) in a germ-line competent manner. | Mc3r Mc4r | Li D et al. (2013) [37] |
| MI mRNA/sgRNAs | CRISPR/Cas9 (indels) | SD | KO rats first produced using CRISPR/Cas9 technology with multiple gene mutations (Tet methylcytosine dioxygenase 1 (Tet1), Tet methylcytosine dioxygenase 2 (Tet2) and Tet methylcytosine dioxygenase 3 (Tet3)) in a germ-line competent manner. | Tet1 Tet2 Tet3 | Li W et al. (2013) [38] |
| MI mRNAs | TALENs (indels) | WI | KO rats generated with a markedly attenuated response to a lipopolysaccharide challenge; can be used as a model for studying ethanol action and general inflammatory conditions including septic shock. | Tlr4 | Ferguson et al. (2013) [39] |
| MI mRNA | ZFNs (indels) | SD | KO rats generated to examine the roles of aryl hydrocarbon receptor (Ahr); showing alterations to the urinary tract, including hydronephrosis and hydroureter; suggesting a role of Ahr in tissue development, homeostasis, and toxicity. | Ahr | Harrill et al. (2013) [40] |
| MI mRNA | ZFNs (indels) | SD | KO rats generated as a model for hemophilia A; showing a prolonged activated partial thromboplastin time and clot formation time; can be useful for the study to treat hemophilia A. | F8 | Nielsen et al. (2013) [41] |
| MI mRNA | ZFNs (indels) | Unknown | KO rat lines generated as a model for autism spectrum disorder (ASD); showing abnormalities in ASD-relevant phenotypes including juvenile play, perseverative behaviors, and sensorimotor gating. | Fmrp Nlgn3 | Hamilton et al. (2013) [42] |
| In vitro EP mRNA/gRNA | ZFN (indels) TALEN (indels) CRISPR/Cas9 (indels) | F344/Stm | KO rats first produced using in vitro electroporation (EP) (called “Technique for Animal Knockout system by Electroporation (TAKE)”); in vitro EP resulted in an embryonic survival rate (91%) and a GE rate (73%). | Il2rg | Kaneko et al. (2014) [43] |
| MI mRNA/DNA donor | TALENs (KI) | SD | Homology-directed repair (HDR)-modified KI rats generated with multiple gene mutations with high efficiency (0.62%–5.13%). | Hprt1 Rosa26 Ighm | Remy et al. (2014) [44] |
| MI mRNA/sgRNAs | CRISPR/Cas9 (KI) | SD | KI rats first generated using CRISPR/Cas9-based two-cut strategy with an efficiency up to 54%. | Trdmt1 Nes Cck | Ma et al. (2014) [45] |
| MI mRNA/sgRNAs | CRISPR/Cas9 (indels, KI) | F344/Stm DA PVG | Using single-stranded oligodeoxynucleotide (ssODN) donor as templates, three recessive phenotypes (including the albino phenotype by single-nucleotide polymorphism (SNP) exchange, the non-agouti phenotype by integration of a 19-bp DNA fragment, and the hooded phenotype by removal of a retrovirus-derived insertional DNA fragment) were successfully corrected. | Tyr Asip Kit | Yoshimi et al. (2014) [46] |
| MI mRNA/sgRNAs | CRISPR/Cas9 (indels) | SD | KO rats generated with multiple gene mutations (four genes) in a germ-line competent manner. | ApoE B2m Prf1 Prkdc | Ma et al. (2014) [47] |
| MI mRNAs | ZFNs (indels) | SD | KO rats generated to examine the function of estrogen receptor 1 (Esr1); showing infertility, due to small testes in males, and large polycystic ovaries, thread-like uteri, and poorly developed mammary glands in females; can be a new tool for investigating the pathophysiology of estrogen action. | Esr1 | Rumi et al. (2014) [48] |
| MI mRNAs/DNA donor | TALENs (KI) | SD | KI rats (carrying a pA476T mutation) first produced using TALEN technology with an efficiency of 17%. | Nr3c1 | Ponce de Leon et al. (2014) [49] |
| MI mRNA/gRNAs | CRISPR/Cas9 (indels) | WI-IM | KO rats generated as a model for Duchenne muscular dystrophy (DMD); showing a decline in muscle strength, and the emergence of degenerative/regenerative phenotypes in the skeletal muscle, heart, and diaphragm; can be useful for developing therapeutic methods to treat DMD. | Dmd | Nakamura et al. (2014) [50] |
| MI mRNA | TALENs (indels) | SD | KO rats generated as a model for DMD; showing reduction in muscle strength and a decrease in spontaneous motor activity with dilated cardiomyopathy; can be useful as an animal model of DMD. | Dmd | Larcher et al. (2014) [51] |
| MI mRNA | ZFNs (indels) | SD | KO rats generated as a model for cystic fibrosis (CF); showing abnormalities in the ileum and increased intracellular mucus in the proximal nasal septa as well as reduced airway surface liquid and periciliary liquid depth; recapitulating many aspects of CF disease. | Cftr | Tuggle et al. (2014) [52] |
| MI mRNA | ZFNs (indels) | SD | KO rats generated to know that the bone morphogenetic protein receptor 2 (Bmpr2) mutations are linked to pulmonary arterial hypertension (PAH); displaying an intense pulmonary vascular remodeling at 3 months; suggesting that endothelial-to-mesenchymal transition (EndoMT) is linked to alterations in BPMR2 signaling. | Bmpr2 | Ranchoux et al. (2015) [53] |
| In vitro EP mRNA/gRNA/ssODN | CRISPR/Cas9 (indels, KI) | WI | KI and KO rats produced through in vitro EP in the presence of CRISPR/Cas9 components with efficiencies of 33% and 88%, respectively. | Il2rg | Kaneko and Mashimo (2015) [54] |
| MI mRNAs | TALENs (indels) | Unknown | KO rats generated to examine the function of cold-inducible RNA-binding protein (Cirp) in heart; CIRP modulates cardiac repolarization by negatively adjusting the expression and function of Ito channels. | Cirp | Li et al. (2015) [55] |
| MI mRNAs | TALENs (indels) | SD | KO rats displayed typical dyslipidemia, although no obvious atherosclerotic lesion was noted in the enface aortas and aortic root; can be a novel model for dyslipidemia and is used in the research of atherosclerosis. | ApoE | Wei et al. (2015) [56] |
| MI protein/gRNA | CRISPR/Cas9 (KI) TALENs (KI) | SD | Cas9 protein was more efficient at HDR than Cas9 mRNA, while TALEN protein was less efficient than TALEN mRNA for inducing HDR. | Rosa26 Foxp3 Anks3 | Ménoret et al. (2015) [57] |
| MI protein/sgRNA/DNA donor | CRISPR/Cas9 (KI) | SD | A DNA cassette (composed of a green fluorescent protein (GFP) reporter sequence flanked by the two pairs of lox sites) was successfully inserted in the reverse orientation into the target (leucine-rich repeat-containing G-protein coupled receptor 5 (Lgr5)) locus, which was later inverted by Cre-mediated recombination. | Lgr5 | Wang et al. (2015) [58] |
| ES cell-mediated GE plasmid DNA | CRISPR/Cas9 (KI) | ES cells | The first production of KI rats using CRISPR/Cas9-engineered embryonic stem (ES) cells; in vitro EP of Cas9 plasmid, sgRNA plasmid, and a KI vector led to enhanced homologous recombination (HR) efficiency up to 36%. | Kat II | Yamamoto et al. (2015) [14] |
| MI mRNA/gRNA | CRISPR/Cas9 (indels) | SD | KO rats generated as a model for obesity; showing obesity, hyperphagia, hyperglycemia, glucose intolerance, hyperinsulinemia, and dyslipidemia, as well as a decrease in bone volume and bone mineral density of the femur; can be useful for the research on obesity and diabetes. | Lepr | Bao et al. (2015) [59] |
| SSC-mediated GE plasmid DNA | CRISPR/Cas9 (indels) | SD-derived SSCs | The first creation of spermatogonial stem cell (SSC)-derived GE KO rats; when recipient male rats were transplanted with engineered SSCs into seminiferous tubules (STs), and then mated with wild-type (WT) females, the resulting progeny harbored indels with ~10% efficiency. | Epsti1 Erbb3 | Chapman et al. (2015) [60] |
| MI mRNA/sgRNA | CRISPR/Cas9 (indels) | SD | KO rats generated as a model for Rett syndrome (RS); showing body weight loss, anxiety tendency, and cognitive deficits; recapitulating the major symptoms of RS patients. | Mecp2 | Zhai et al. (2016) [61] |
| MI mRNA | ZFNs (indels) | SD | KO rats generated as a model for RS; showing behavioral and motor deficits in male and female rats; can be used in RS research. | Mecp2 | Patterson et al. (2016) [62] |
| MI mRNA/sgRNA | CRISPR/Cas9 (indels) | SD | KO rats generated as a model for hereditary tyrosinemia type I (HT1); showing major phenotypic manifestations of human HT1, including hypertyrosinemia, liver failure, and renal tubular damage; also showing remarkable liver fibrosis and cirrhosis; can be used as a model of HT1 with liver cirrhosis. | Fah | Zhang et al. (2016) [63] |
| MI mRNA/gRNA/ lssODN | CRISPR/Cas9 (KI) | F344/Stm | The ssODN-mediated KI rats were successfully generated through two-hit by gRNA and two-oligo with plasmid (2H2OP) system; can be applied to any target site with any donor vector without the need to construct homology arms, thus simplifying genome engineering in living organisms. | Tyr Thy1 Rosa26 Cyp2d Sirpa | Yoshimi et al. (2016) [64] |
| MI mRNA/gRNA/ ssODN | CRISPR/Cas9 (KI) | SD | Usage of chemically modified (phosphorothioate-modified) ssODN donors is shown to be effective in improving KI efficiency. | Cftr | Renaud et al. (2016) [65] |
| MI mRNA | TALENs (indels) | SD | Homozygous KO rats are embryonically lethal; both male and female fumarate hydratase (Fh)+/- KO rats exhibited reduced litter size; also showing hematopoietic and kidney dysfunction; Fh+/- KO rats can be useful for further functional fumarate hydratase studies. | Fh | Yu et al. (2016) [66] |
| MI mRNA | TALENs (indels) | F344 | Homozygous KO rats are embryonically lethal; DEP domain-containing 5 (Depdc5)(+/-) rats developed normally and exhibited no spontaneous electroclinical seizures, but had altered cortical neuron excitability and firing patterns; can be a model for mTORopathy. | Depdc5 | Marsan et al. (2016) [67] |
| MI protein (or mRNA)/sgRNA/donor plasmid | CRISPR/Cas9 (KI) | SD | Both Scr7 (non-homologous end joining (NHEJ) inhibitor) and Cas9 protein were used to increase KI efficiency; consequently, this combination successfully increased the HR-mediated precise modification. | Fabp2 Dbndd1 | Ma et al. (2016) [68] |
| MI mRNA/sgRNAs | CRISPR/Cas9 (indels) | WI | KO rats generated to examine the function of forkhead box N1 (Foxn1) that is known to a critical factor for differentiation of thymic and skin epithelial cells; showing thymus deficiency and incomplete hairless. | Foxn1 | Goto et al. (2016) [69] |
| MI mRNA/sgRNAs | CRISPR/Cas9 (indels) | SD | Mc3r and Mc4r single-KO or DKO rats generated to examine Mc3r and Mc4r more deeply; Mc3r KO rats displayed hypophagia and decreased body weight, while Mc4r KO and DKO exhibited hyperphagia and increased body weight; both Mc4r KO and DKO are good models for obesity and diabetes research. | Mc3r Mc4r | You et al. (2016) [70] |
| MI mRNA | TALENs (indels) | SD | KO rats generated to evaluate the role of reduced nuclear factor (erythroid-derived 2)-like-2 (NRF2)-regulated antioxidant defenses; suppression of NRF2 antioxidant defenses plays a role in the development of salt-induced oxidant stress, endothelial dysfunction, and microvessel rarefaction; can be used as therapeutics to ameliorate vascular oxidant stress in humans. | Nrf2 | Priestley et al. (2016) [71] |
| MI mRNA | ZFNs (indels) | F344/Stm | KO rats generated to evaluate the role of reduced NRF2-regulated antioxidant defenses were sensitive to aflatoxin B1 toxicity; can be a new model animal in toxicology. | Nrf2 | Taguchi et al. (2016) [72] |
| MI mRNA/gRNA | CRISPR/Cas9 (indels) | Unknown | KO rats generated to investigate the functions of cytochrome P450 2E1 (CYP2E1) were viable and fertile and did not display any obvious physiological abnormities; can be a powerful tool for the study of CYP2E1 in the chemical metabolism, toxicity, carcinogenicity. | Cyp2e1 | Wang et al. (2016) [73] |
| MI mRNA/gRNA | CRISPR/Cas9 (indels) | WI | KO rats generated to examine the function of Mohawk homeobox (Mkx) in tenogenesis; showing systemic hypoplasia of tendons and earlier heterotopic ossification of the Achilles tendon; can be used for studying tendon physiology and tissue engineering. | Mkx | Suzuki et al. (2016) [74] |
| MI mRNA | ZFNs (indels) | SD | KO rats generated as a model for pseudoxanthoma elasticum, a heritable ectopic mineralization disorder; showing reduced levels of plasma inorganic pyrophosphate (PPi), suggesting a critical role of hepatic ATP binding cassette subtype C number 6 (ABCC6) in contributing to plasma PPi levels. | Abcc6 | Li et al. (2017) [75] |
| MI mRNA | ZFNs (indels) | SD | KO rats generated as model of Wolfram syndrome (WS), a rare autosomal-recessive disorder characterized by juvenile-onset diabetes, optic atrophy, and hearing loss; exhibiting the core symptoms of WS. | Wfs1 | Plaas et al. (2017) [76] |
| MI mRNA/sgRNA | CRISPR/Cas9 (indels) | SD | KO rats specifically impairs long-term synaptic plasticity and hippocampus-dependent learning in a manner resembling the key symptoms of Fragile X syndrome (FXS). | Fmr1 | Tian et al. (2017) [77] |
| MI mRNA/gRNA | CRISPR/Cas9 (indels) | Unknown | KO rats generated to investigate the role of myosin light-chain 4 (MYL4) in atrial cardiomyopathy; showing progressive atrial cardiomyopathy; suggesting that Myl4 is as a key gene required for atrial contractile, electrical, and structural integrity. | Myl4 | Peng et al. (2017) [78] |
| MI mRNA | TALENs (indels) | SD | KO rats generated as obese rat model; showing early onset of obesity and infertility. | Lepr | Chen et al. (2017) [79] |
| MI mRNA/gRNA | CRISPR/Cas9 (indels) | SD | KO rats generated to determine the function of patatin-like phospholipase domain containing 5 (Pnpla5); were viable, but showed a variety of abnormalities related to lipid metabolism; can be a possibly model for the treatment of cardiovascular diseases. | Pnpla5 | Liu et al. (2017) [80] |
| MI mRNA | ZFNs (indels) | SD | KO rats generated as a model for Phelan–McDermid syndrome (PMS); exhibit impaired long-term social recognition memory and attention, and reduced synaptic plasticity in the hippocampal-medial prefrontal cortex pathway. | Shank3 | Harony-Nicolas et al. (2017) [81] |
| MI mRNA/gRNA (or mRNA/gRNA/ssODN) | CRISPR/Cas9 (KI, indels) | SS | GM rats generated to assess the candidacy of a novel predicted long non-coding RNA (lncRNA) showing strong association of cardiac QT-interval; KO rats showed aberrant, short QT-intervals, and elevated blood pressure; KI rats showed the rescued phenotype. | Rffl-lnc1 | Cheng et al. (2017) [82] |
| MI mRNA | TALENs (indels) | SD | KO rats generated to examine why rats are naturally resistant to Schistosoma japonicum infection; high expression levels of inducible nitric oxide synthase (iNOS) in rats play an important role in blocking the egg-induced granuloma formation of S. japonicum; providing insights for understanding the pathogenesis of human schistosomiasis. | iNOS | Shen et al. (2017) [83] |
| MI mRNA | ZFNs (indels) | SS | KO rats generated to define the importance of Kir5.1 (encoded by potassium inwardly-rectifying channel, subfamily J, member 16 (Kcnj16)) in blood pressure control under salt-induced hypertension; exhibiting hypokalemia and reduced blood pressure, and 100% mortality within a few days triggered by salt wasting; demonstrating that Kir5.1 channels are key regulators of renal salt handling. | Kcnj16 | Palygin et al. (2017) [84] |
| ES cell-mediated GE plasmid | CRISPR/Cas9 (KI) | ES cells | KI rats derived from the CRISPR/Cas9-engineered ES cells; two clones showing non-disruptive KI of Discosoma coral red fluorescent protein (dsRed) at the Sry-box transcription factor 10 (Sox10) locus produced germ-line chimeras; showing expression of dsRed in the Sox10-expressing cells (oligodendrocyte lineage cells) in vivo. | Sox10 | Chen et al. (2017) [15] |
| MI mRNA/gRNA | CRISPR/Cas9 (indels) | SD | DKO rats generated to investigate cytochrome P450 3A (CYP3A) functions; were viable and fertile, and had no obvious physiological abnormities; can be a powerful tool for the study of the physiological and pharmacological roles of CYP3A, especially in drug and chemical metabolism in vivo. | Cyp3a1 Cyp3a2 | Lu et al. (2017) [85] |
| MI mRNA | ZFNs (indels) | BN | KO rats generated as a model for autoimmune polyendocrinopathy-candidiasis-ectodermal dystrophy (APECED); displaying several of the key symptoms of APECED, including alopecia, skin depigmentation, and nail dystrophy | Aire | Ossart et al. (2018) [86] |
| MI mRNA/gRNA/ssODN | CRISPR/Cas9 (KI) | SD | KI rats (carrying a Fused in sarcoma (Fus)-point mutation (R521C)) generated as a model for amyotrophic lateral sclerosis (ALS); developing adult-onset learning and memory behavioral deficits cognitive impairment; suggesting a new role of FUS in sleep and circadian regulation. | Fus | Zhang et al. (2018) [87] |
| MI mRNA | TALENs (indels) | SD | Immunodeficient rats with DKO rats generated for obtaining more profound immunosuppressed phenotype; can be useful as recipients for long-term studies concerning tissue humanization of different tissues. | Il2rg Rag1 | Ménoret et al. (2018) [88] |
| MI mRNA/gRNA | CRISPR/Cas9 (indels) | F344/Stm | KO rats generated to clarify the roles of angiopoietin-like protein 8 (ANGPTL8) in glucose and lipid metabolism; showing lower body weight and fat content, associated with impaired lipogenesis in adipocytes; can be an important therapeutic target for obesity and dyslipidemia. | Angptl8 | Izumi et al. (2018) [89] |
| i-GONAD protein/gRNA (or protein/gRNA/ssODN) | CRISPR/Cas9 (indels, KI) | SD LEW BN | Improved GONAD (i-GONAD) was found useful for producing KO and KI rats with efficiencies of 62 to 97% and 9%, respectively. | Tyr Sox16 | Takabayashi et al. (2018) [90] |
| i-GONAD (rGONAD) protein/gRNA (or protein/gRNA/ssODN) | CRISPR/Cas9 (indels, KI) | WKY DA | Most suitable condition for in vivo gene delivery towards rat preimplantation embryos was assessed; i-GONAD was found useful for producing KO and KI rats with relative high efficiency. | Tyr | Kobayashi et al. (2018) [91] |
| MI mRNA/gRNA | CRISPR/Cas9 (indels) | Unknown | DKO rats (lacking both apolipoprotein E (ApoE) and low-density lipoprotein receptor (Ldlr)) generated as a model for familial hypercholesterolemia; showing hypercholesterolemia, hepatosteatosis, and atherosclerosis. | ApoE Ldlr | Zhao et al. (2018) [92] |
| MI mRNA/gRNA | CRISPR/Cas9 (indels) | SD | KO rats generated to investigate whether Cas9 nickase (Cas9n)-mediated GE can efficiently correct the HT1; tail-vein injection of adenoviral vectors carrying Cas9n, repair donor template, and sgRNA successfully corrected the phenotype of fumarylacetoacetate hydrolase (Fah) KO rats. | Fah | Shao et al. (2018) [93] |
| MI mRNA/gRNA | CRISPR/Cas9 (indels) | Unknown | KO rats generated as a model for primary hyperoxaluria type 1 (PH1); showing crystalluria with abnormal phenotypes, such as a slight dilatation of renal tubules with mild fibrosis in the kidney; suggesting impairment of glyoxylic acid metabolism; can be a useful tool for the development and evaluation of drugs and therapeutics. | Agxt | Zheng et al. (2018) [94] |
| In vitro EP (protein/gRNAs/lssDNA) | CRISPR/Cas9 (KI) | F344 | Efficient approaches enabling the quick generation of floxed alleles in mice and rats were taken using in vitro EP in the presence of Cas9 protein, gRNAs, and long single-stranded DNA (lssDNA); homozygous KI oocytes carrying tissue-specific Cre gene were successfully obtained with high efficiency. | Vapb | Miyasaka et al. (2018) [95] |
| MI mRNA/sgRNA | CRISPR/Cas9 (indels) | SD | KO rats generated as a model for RNase T2 deficiency; showing no evidence of cystic lesions, but hippocampal neuroinflammation, altered lysosomal function, and cognitive defects; can be a useful model to study the RNase T2 function. | RNase T2 | Sinkevicius et al. (2018) [96] |
| MI mRNA/sgRNA | ABEs (indels) | Unknown | KO rats generated as a model for Glycogen storage disease type II (GSDII) with an efficiency of 85%; the adenine base editor (ABE) system is a powerful and convenient tool to introduce precise base conversions in rodents. | Gaa | Yang et al. (2018) [97] |
| MI plasmid DNA | CRISPR/Cas9 (indels) | DA | KO rats generated as a model for Fabry disease (FD); showing substantial serum and tissue accumulation of α-galactosyl glycosphingolipids and had pronounced mechanical pain behavior. | α-Gal A | Miller et al. (2018) [98] |
| MI mRNA | TALENs (indels) | SD | Oxoglutarate dehydrogenase (Ogdh)−/− rats are lethal; Ogdh+/− rats had higher body weight; treatment of Ogdh+/− rats with a high-fat diet resulted in liver dysfunction; Ogdh+/− rats can be used as model for investigations of metabolic syndrome and obesity-related hepatic carcinogenesis. | Ogdh | Fan et al. (2018) [99] |
| MI mRNA | ZFNs (indels) | WI | KO rats developed normally, but exhibited spontaneous locomotor hyperactivity and cognitive dysfunctions; can be a model for human diseases involving aberrant dopamine functions. | Dat | Leo et al. (2018) [100] |
| MI plasmid | CRISPR/Cas9 (indels) | F344 SD | KO rats generated to know the role of complement in chemotherapy-induced peripheral neuropathy (CIPN); suggesting that complement may be a new target for the development of novel therapeutics to manage CIPN. | C3 | Xu et al. (2018) [101] |
| SSC-mediated GE plasmid | TALENs (indels) | SD-derived SSCs | SSCs-derived GE KO rats showed immunodeficiency with lacked mature B and T cells; allowing growth of human tumors; can be used for xenograft studies. | Rag2 | Noto et al. (2018) [102] |
| MI mRNA/sgRNAs | CRISPR/Cas9 (indels) | Unknown | Multiple-gene KO rats generated to examine the function of defensin beta (Defb) expressed abundantly in the epididymis; showing subfertility and decreased motility of sperm; suggesting that Defb family members affect sperm maturation by a synergistic pattern in the epididymis. | Defb23 Defb26 Defb42 | Zhang et al. (2018) [103] |
| MI mRNA/sgRNA | ABE (indels) | SD | For hemogen (Hemgn) targeting, out of 15 rats obtained, 14 contained an A to G conversion at the 14th base distal from the protospacer adjacent motif (PAM), indicating the high base-editing efficiency. | Hemgn Ndst1 Ndst4 | Ma et al. (2018) [104] |
| MI mRNA/sgRNA | CRISPR/Cas9 (indels) | SD | KO rats generated to assess the roles of cytochrome P450, subfamily 2, polypeptide 11 (CYP2C11); were viable and had no obvious abnormalities, with the exception of reduced fertility; can be a valuable tool to study the in vivo function of CYP2C11. | Cyp2c11 | Wei et al. (2018) [105] |
| MI mRNA/gRNA (or mRNA/gRNA ssODN) | CRISPR/Cas9 (indels, KI) | Unknown | KI (carrying F508del mutation) and KO rats generated as a model for CF; both rats exhibit CF phenotypic anomalies such as vas deferens agenesis and tooth enamel defects; can be used as a novel resource to advance the development of CF therapeutics. | Cftr | Dreano et al. (2019) [106] |
| MI plasmid DNA | CRISPR/Cas9 (indels) | DA | KO rats generated as a model for FD; showing pronounced renal tubule dysfunction and mitral valve thickening; can be further used to study disease mechanisms and test therapies. | α-Gal A | Miller et al. (2019) [107] |
| MI mRNA/sgRNA | CRISPR/Cas9 (indels) | SD | KO rats generated a model for ASDs; showing impaired social memory (but not impaired social interaction behaviors), impaired learning and memory, increased anxiety-like behavior, increased mechanical pain threshold, and decreased thermal sensation. | Shank3 | Song et al. (2019) [108] |
| MI mRNA/sgRNA | CRISPR/Cas9 (indels) | SD | KO rats generated as a model for pulmonary hypertension (PH); showing PH pathogenesis associated with low serum-albumin concentration; can be used for understanding the mechanisms of PH and testing therapeutic molecules. | Kcnk3 | Lambert et al. (2019) [109] |
| MI mRNA/sgRNA | CRISPR-Cpf1 (indels) | SD | ApoE KO rats displayed hyperlipidemia and aortic lesions; showing that the Cpf1 system can target single or multiple genes efficiently; these rats can be helpful for understanding initial-stage atherosclerosis. | ApoE Ldlr | Lee et al. (2019) [110] |
| In vitro EP (protein/sgRNA/plasmid DNA) rAAV | CRISPR/Cas9 (KI, indels) | BN | Superovulation is successfully conducted to Brown Norway (BN), which is a very difficult rat strain to superovulate; in vitro EP or infection with recombinant adeno-associated virus (rAAV) enables KO or KI in the in vitro fertilized (IVF) rat embryos efficiently. | Tyr | Honda et al. (2019) [111] |
| MI mRNA/sgRNA | CRISPR/Cas9 (indels) | SD | KO rats exhibited complete loss of multidrug resistance protein 1 (MDR1) in the liver, small intestine, brain, and kidney; can be useful for studying the function of MDR1 in drug absorption, tumor multidrug, and drug target validation. | Mdr1a/b | Liang et al. (2019) [112] |
| MI protein/sgRNA | CRISPR/Cas9 (indels) | SD | KO rats generated as a model for X-linked hydrocephalus (XLH); hemizygous male mutants developed hydrocephalus and delayed development. | L1cam | Emmert et al. (2019) [113] |
| MI mRNA/sgRNA/ssODN | CRISPR/Cas9 (KI) | SD | KI rats (carrying a R571X mutation) generated to mimic Cockayne syndrome (CS); showing atrophy and dysmyelination in the cerebellar cortex; RNA-seq suggests that transcription dysregulation could contribute to the CS pathogenesis. | Csb | Xu et al. (2019) [114] |
| MI protein/sgRNA | CRISPR/Cas9 (indels) | SD | KO rats generated to understand the mechanisms leading to iron excess in HA; showing decreased iron concentration and transferrin saturation; suggesting that the reported role of ceruloplasmin (Cp) cannot fully explain the iron hepatosplenic phenotype in hereditary aceruloplasminemia (HA). | Cp | Kenawi et al. (2019) [115] |
| MI mRNA | ZFNs | SD | KO rats generated as a model for PAH; showing age-dependent spontaneous PAH with a low penetrance (16%-27%), similar to that in humans; can be a promising tool to study the pathogenesis of PAH. | Bmpr2 | Hautefort et al. (2019) [116] |
| In vitro EP protein/gRNA | CRISPR/Cas9 (indels) | SD | KO rats generated to examine the role of 5-hydroxytryptamine receptor 7 (Htr7) in blood pressure regulation; neither the male nor female KO rats were different in body size, fat weights, or mass of organs (kidney, heart, and brain) important to blood pressure; can be used to investigate the importance of Htr7 in blood pressure regulation. | Htr7 | Demireva et al. (2019) [117] |
| MI mRNA/two sgRNAs/donor plasmid containing two flanking loxP sites | CRISPR/Cas9 (KI) | SD | KI rats generated to create a conditional glucocorticoid receptors (GR) knockdown using a dual sgRNA strategy; affording high-precision knockdown of GR across a variety of contexts, ranging from neuronal depletion to circuit-wide manipulations. | Nr3c1 | Scheimann et al. (2019) [118] |
| MI mRNA/ sgRNA/donor plasmid | CRISPR/Cas9 (KI) | LE | Two KI rat lines (D1-Cre and A2a-Cre) generated for Cre-mediated gene manipulation; will be used to study both normal brain functions and neurological and psychiatric pathophysiology. | Drd1a Adora2a | Pettibone et al. (2019) [119] |
| MI mRNA/gRNA | CRISPR/Cas9 (indels) | SS | KO rats generated as a model for hemophilia; showing spontaneous bleeding in the soft tissue, muscles, or joints occurred in 100% of factor 8 (F8)-/- rats; bone marrow reconstitution rescued the spontaneous bleeding in rat HA hemophilia. | F8 | Shi et al. (2020) [120] |
| i-GONAD protein/gRNAs | CRISPR/Cas9 (indels) | BN | i-GONAD under EP conditions with current values ranging from 150 to 200 mA enabled production of KO rats with efficiencies ranging from 75% to 24%. | Tyr | Takabayashi et al. (2020) [121] |
| MI mRNA/gRNA | CRISPR/Cas9 (indels) | SD | KO rats generated as a model for Angelman syndrome (AS); showing deficits in motor coordination as well as learning and memory, phenotypically mirroring human AS; can be a model for the study of AS. | Ube3a | Dodge et al. (2020) [122] |
| MI mRNA/gRNA | CRISPR/Cas9 (indels) | SD | KO rats generated to explore the function of complexin I (Cplx1); showing a complex neurobehavioral phenotype including profound ataxia, dystonia, movement and exploratory deficits, and increased anxiety and sensory deficits, but had normal cognitive function. | Cplx1 | Xu et al. (2020) [123] |
| MI mRNA (or protein)/gRNAs/plasmid donor | Cas12a/Cpf1 (MAD7) (indels, KI) | SD | MAD7 is capable of generating indels, small DNA insertion, and fluorescent gene tagging of endogenous genes, such as Cas9; causing indels with 20% efficiency in rats; MAD7 can expand the CRISPR toolbox for genome engineering. | Calb2 | Liu et al. (2020) [124] |
| In vitro EP mRNA/gRNA (or mRNA/gRNA/ lssDNA) | CRISPR/Cas9 (indels, KI) | LE | KO rats generated to know the role of glutamate decarboxylase 1 (GAD1) deficiency in pathogenesis of schizophrenia; showing complex behavioral changes, such as hypoactivity in a novel environment and decreased preference for social novelty; can be a model for studying the mechanism of schizophrenia. | Gad1 | Fujihara et al. (2020) [125] |
| MI protein/sgRNA/ssODN | CRISPR/Cas9 (KI) | SD | KI rats (carrying a R350P mutation) generated as a model for desminopathy; showing the phenotype of desminopathy; can be a useful tool for understanding of the disease and testing therapeutic approaches to delay disease progression. | Des | Langer et al. (2020) [126] |
| MI mRNA/sgRNA/ssODN | CRISPR/Cas9 (KI) | SD | KI rats (carrying a D205N mutation) generated as a model for primary hyperoxaluria type 1 (PH1); showing hyperoxaluria at 1 month of age and exhibited severe renal calcium oxalate deposition after ethylene glycol challenge; can be a useful model for understanding the disease and developing therapeutics. | Agxt | Zheng et al. (2020) [127] |
| MI mRNA/gRNA | CRISPR/Cas9 (indels) | SD | KO rats generated a model for hyperuricemia; were apparently healthy with more than a 95% survival up to one year and showed renal abnormality such as mild glomerular/tubular lesions; can be useful for studying hyperuricemia. | Uox | Yu et al. (2020) [128] |
| MI mRNA/sgRNA (or mRNA/sgRNA/ssODN) | CRISPR/Cas9 (KI, indels) | WI | Cytochrome P450 family 27 subfamily B member 1 (Cyp27b1) KO, vitamin D receptor (Vdr) KO, and Vdr KI (carrying a R270L mutation) rats generated as a model for human type II rickets; showing growth retardation and abnormal bone formation; can be useful for elucidating the molecular mechanism of vitamin D action. | Cyp27b1 Vdr | Nishikawa et al. (2020) [129] |
| MI mRNA/donor DNA | ZFNs (KI) | SD | KI rats (carrying a G551D variant) generated as a model for CF; showing that the epithelia recapitulates the expected absence of cystic fibrosis transmembrane conductance regulator (CFTR) activity, which was restored with ivacaftor, a medication used to treat CF. | Cftr | Birket et al. (2020) [130] |
| In vitro EP protein/gRNA | CRISPR/Cas9 (indels) | SD | KO rats generated to investigate the role of monocarboxylate transporter 8 (MCT8) (encoded by solute carrier family 16 member 2 coding for monocarboxylate transporter 8 (Slc16a2)) during spermatogenesis; showing growth delay during postnatal development with reduced sperm motility and viability; suggesting the role of MCT8 in spermatogenesis. | Slc16a2 | Bae et al. (2020) [131] |
| MI mRNA | ZFNs (indels) | WI | KO rats generated to examine whether dopamine plays a key role in sexual behavior; showing more rapid acquisition of stable sexual activity levels and to higher levels of sexual motivation and activity; can be confirmed that dopamine has a key role in sexual behavior. | Dat | Sanna et al. (2020) [132] |
| MI mRNA/sgRNA | CRISPR/Cas9 (indels) | SD | KO rats generated to enable more sophisticated modeling of pain, itch, and asthma; showing apparently normal behavioral responses to pain and itch, although transient receptor potential family member ankyrin 1 (TRPA1)-dependent immune cell infiltration into the lung was seen under the asthma-inducing condition; useful for studying that TRPA1 can be as a drug target. | Trpa1 | Reese et al. (2020) [133] |
| MI mRNA/sgRNA | CRISPR/Cas9 (indels) | SD | KO rats generated to study the function of cytochrome P450 family 2 subfamily J (Cyp2J) isoform genes in vivo; were viable and fertile; can be a useful tool to study the function of Cyp2J in drug metabolism and cardiovascular disease. | Cyp2J2 Cyp2J3 Cyp2J10 | Lu et al. (2020) [134] |
| MI mRNA | TALENs (indels) | F344 | KO rats generated to study the role of coiled-coil domain containing 85C (Ccdc85c) in the pathogenesis of spontaneous hydrocephalus shown in hemorrhagic hydrocephalus (hhy) mice; showing non-obstructive hydrocephalus, subcortical heterotopia, and intracranial hemorrhage; suggesting a role of Ccdc85c in cerebral development. | Ccdc85c | Konishi et al. (2020) [135] |
| MI mRNA/gRNA/ssODN | CRISPR/Cas9 (KI) | LE | KI rats (carrying a Psen1LF mutation) generated to determine potential early pathogenic changes caused by this mutation; survived into adulthood; showing increased levels of Aβ43, a longer and potentially more amyloidogenic Aβ form. | Psen1 | Tambini and D’Adamio (2020) [136] |
| MI protein/sgRNAs/donor DNA | CRISPR/Cas9 (KI) | SD | KI rats (carrying GGGGCC (G4C2) repeats) generated as a model for ALS; showing motor deficits from 4 months of age; can be used for investigating the neurotoxicity in chromosome 9 open reading frame 72 (C9orf72)-related ALS. | C9orf72 | Dong et al. (2020) [137] |
| MI protein/sgRNAs/donor DNA | CRISPR/Cas9 (KI) | SD | KI rats generated as a model for the Mediterranean SNP G6PDS188F; showing glucose-6-phosphate dehydrogenase (G6PD) activity, but not expression, was reduced to 20% of WT littermates; replacement of a single amino acid (S188F) in G6PD, a rate-limiting enzyme in the pentose phosphate pathway, may be related to pathogenesis of vascular diseases. | G6pd | Kitagawa et al. (2020) [138] |
| MI mRNA/sgRNA | CRISPR/Cas9 (indels) | SD | KO rats generated to examine the role of solute carrier organic anion transporter family member 1B2 (Slco1b2); in those rats, the serum levels of bilirubin and bile acids, the substrates of organic anion transporting polypeptide organic anion transport polypeptide 1b2 (OATP1B2), increased; can be a disease model to study hyperbilirubinemia-related diseases. | Slco1b2 | Ma et al. (2020) [139] |
| MI, in vitro EP mRNA/sgRNAs/dsDNA donor vectors (or mRNA/sgRNAs/lssDNA) | CRISPR/Cas9 (KI) | F344 | A new powerful method, called Combi-CRISPR, was developed for achieving plasmid-based KI in rat embryos using the CRISPR/Cas9 system; in other words, this method is the combination of highly efficient editing via NHEJ and the low-efficiency but precise editing via HDR. | Pvalb Th | Yoshimi et al. (2021) [140] |
| i-GONAD protein/gRNA/ssODN | CRISPR/Cas9 (KI) | SD | Effects of 3 gRNAs (which recognize different portions of the target locus, but also overlap each other in the target locus) and of commercially available KI-enhancing drugs on the KI efficiency; KI efficiency largely depends on the type of gRNA used; none of drugs are significantly effective for KI efficiency. | Tyr | Aoshima et al. (2021) [141] |
| MI protein/gRNA (or protein/gRNA/ssODN) | CRISPR/Cas9 (indels, KI) | BN | KO rats generated as a model for retinal dystrophy (RD); showing reduced vision and structural abnormalities, such as overall retinal thinning; can be new animal model for retinal dystrophy. | Lrat | Koster et al. (2021) [142] |
| MI mRNA/sgRNA | CRISPR/Cas9 (indels) | WI | KO rats generated to investigate cytochrome P450 family 24 subfamily A member 1 (CYP24A1)-dependent or -independent metabolism of 25(OH)D3, the prohormone of calcitriol; found to be valuable for metabolic studies of vitamin D and its analogs. | Cyp24a1 | Yasuda et al. (2021) [143] |
| In vitro EP mRNA | TALENs (indels) | SD | KO rats generated to elucidate the functions of NK3 homeobox 1 (Nkx3.1); showing reduced fertility, decreased prostate weights, and increased epithelial cell layers; can be a model for studying the role of Nkx3.1 in decreased prostate weights, fertility, and breast cancer, as well as in prostate cancer. | Nkx3.1 | Lee et al. (2021) [144] |
| MI mRNA/sgRNA | CRISPR/Cas9 (indels) | Unknown | KO rats generated to elucidate the roles of alpha/beta-hydrolase domain 6 (ABHD6) in vivo; showing normal appearance, but caused more frequent urination in the stimulated bladder. | Abhd6 | Noguchi et al. (2021) [145] |
| MI plasmid | CRISPR/Cas9 (indels) | SS | KO rats lacking adaptor protein 1 (shc1), encodes 3 main protein isoforms (p52Shc and p46Shc isoforms) generated to further investigate the function of Shc1; showing severe gait abnormalities accompanied by dilated cardiomyopathy. | p52Shc p46Shc | Miller et al. (2021) [146] |
| MI mRNA/sgRNAs | CRISPR/Cas9 (indels) | WI | KO rats generated to investigate the in vivo roles of carnosine synthase 1 (Carns1); showing a significant impairment of contractile function in the cardiac muscle; suggesting a role of high-cholesterol diet (HCD) in the regulation of Ca2+ handling and excitation–contraction coupling of cardiac muscle. | Carns1 | de Souza Gonçalves et al. (2021) [147] |
| MI mRNA/sgRNAs | CRISPR/Cas9 (indels) | SD | KO rats generated to examine the function of transient receptor potential cation channel subfamily V member 4 (Trpv4) in regulating osteoarthritic pain; showing suppression of joint pain under the monoiodoacetate-induced osteoarthritic pain model; suggesting that inhibition of TRPV4 might be a novel potent analgesic strategy for treating osteoarthritic pain. | Trpv4 | Soga et al. (2021) [148] |
| MI mRNA/sgRNAs | CRISPR/Cas9 (indels) | SD | KO rats generated to examine the role of carboxylesterase 2A (Ces2a) in drug metabolism mediated by carboxylesterase 2 (CES2); showing obesity, insulin resistance, and liver fat accumulation, which are consistent with the symptoms of nonalcoholic fatty liver disease (NAFLD); can be used as a model for NAFLD. | Ces2a | Liu et al. (2021) [149] |
| MI protein/two plasmids (one containing gRNA and the other containing Cre-ERT2-mCherry cassette as donor plasmid) | CRISPR/Cas9 (KI) | SD | Generation of six Cre driver rats allowing for the controlled gene expression or conditional KO in a temporal and spatial manner through the Cre-ERT2/loxP system. | Wnt1 Pomc Mnx1 Drd1a Gad67 Tie2 | Zhang et al. (2021) [150] |
| MI mRNA/gRNA/ssODN | CRISPR/Cas9 (KI) | SD F344 | KI rats (carrying a T300A mutation) generated as a model for Crohn’s disease (CD); showing morphological abnormalities in both Paneth and goblet cells, but do not develop spontaneous intestinal permeability or inflammatory bowel disease; can be used for the study of both autophagy and CD susceptibility. | Atg16l1 | Chesney et al. (2021) [151] |
| MI mRNA/gRNA | CRISPR/Cas9 (indels) | SD | KO rats generated to examine the function of dynein axonemal heavy chain 17 (Dnah17) in spermatogenesis; showing infertility with significantly decreased number of sperm; DNAH17 is critical for spermatogenesis. | Dnah17 | Chen et al. (2021) [152] |
| MI PB vector carrying DdCBE pair/PB transposase mRNA | DdCBE (gene correction) | SD | DddA-derived cytosine base editor (DdCBE) was applied to explore the possible production of GE rats as a mitochondrial disease model with pathogenic mitochondrial DNA (mtDNA) mutations; MI of piggyBack transposon (PB) vector carrying DdCBE pair with PB transposase mRNA resulted in production of pups with mtDNA mutations with 36% efficiency. | G7755 | Qi et al. (2021) [153] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sato, M.; Nakamura, S.; Inada, E.; Takabayashi, S. Recent Advances in the Production of Genome-Edited Rats. Int. J. Mol. Sci. 2022, 23, 2548. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052548
AMA Style
Sato M, Nakamura S, Inada E, Takabayashi S. Recent Advances in the Production of Genome-Edited Rats. International Journal of Molecular Sciences. 2022; 23(5):2548. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052548
Chicago/Turabian StyleSato, Masahiro, Shingo Nakamura, Emi Inada, and Shuji Takabayashi. 2022. "Recent Advances in the Production of Genome-Edited Rats" International Journal of Molecular Sciences 23, no. 5: 2548. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052548
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.