
An Efficient Way to Screen Inhibitors of Energy-Coupling Factor (ECF) Transporters in a Bacterial Uptake Assay

 and
and
Abstract
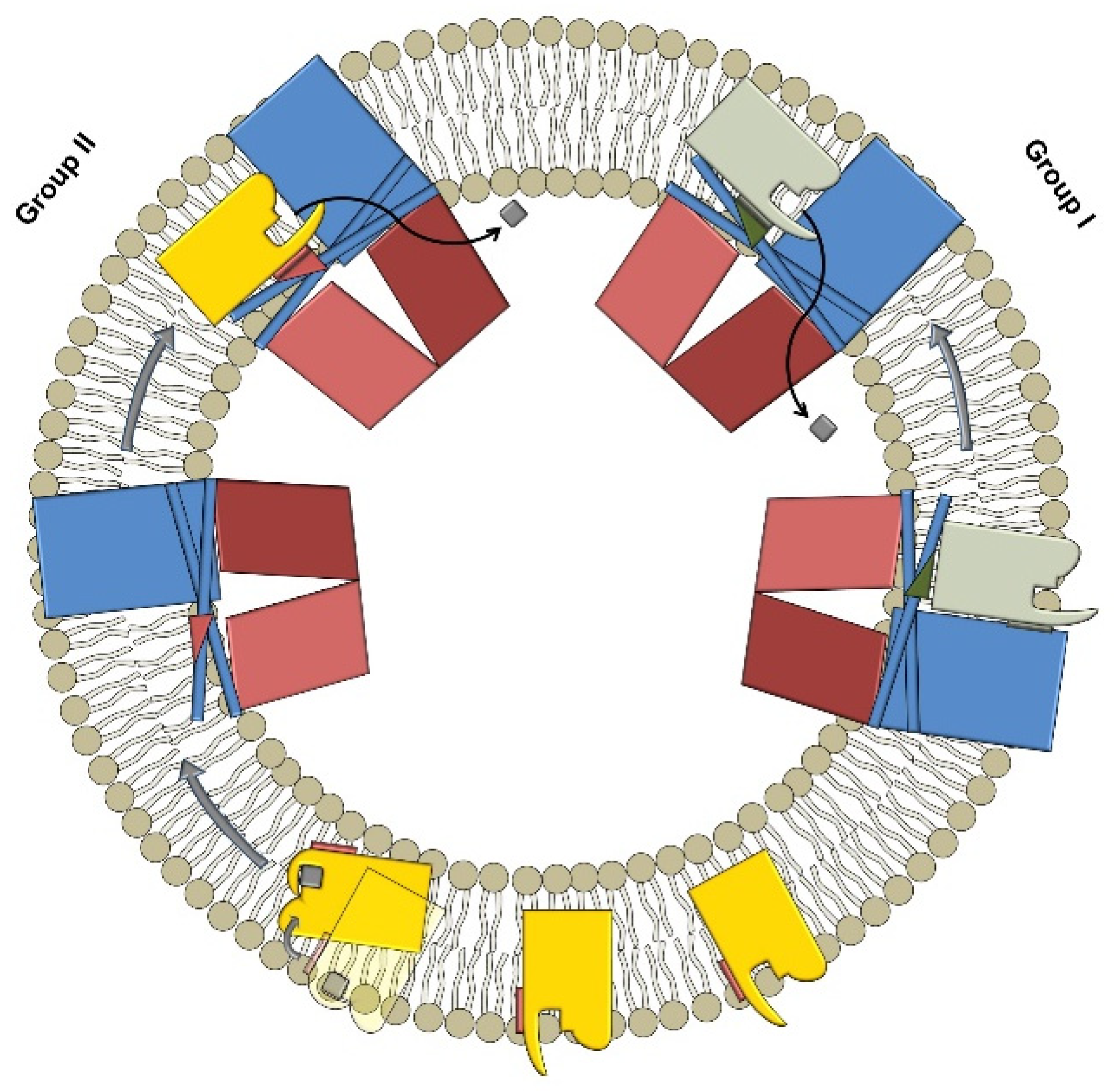
:1. Introduction
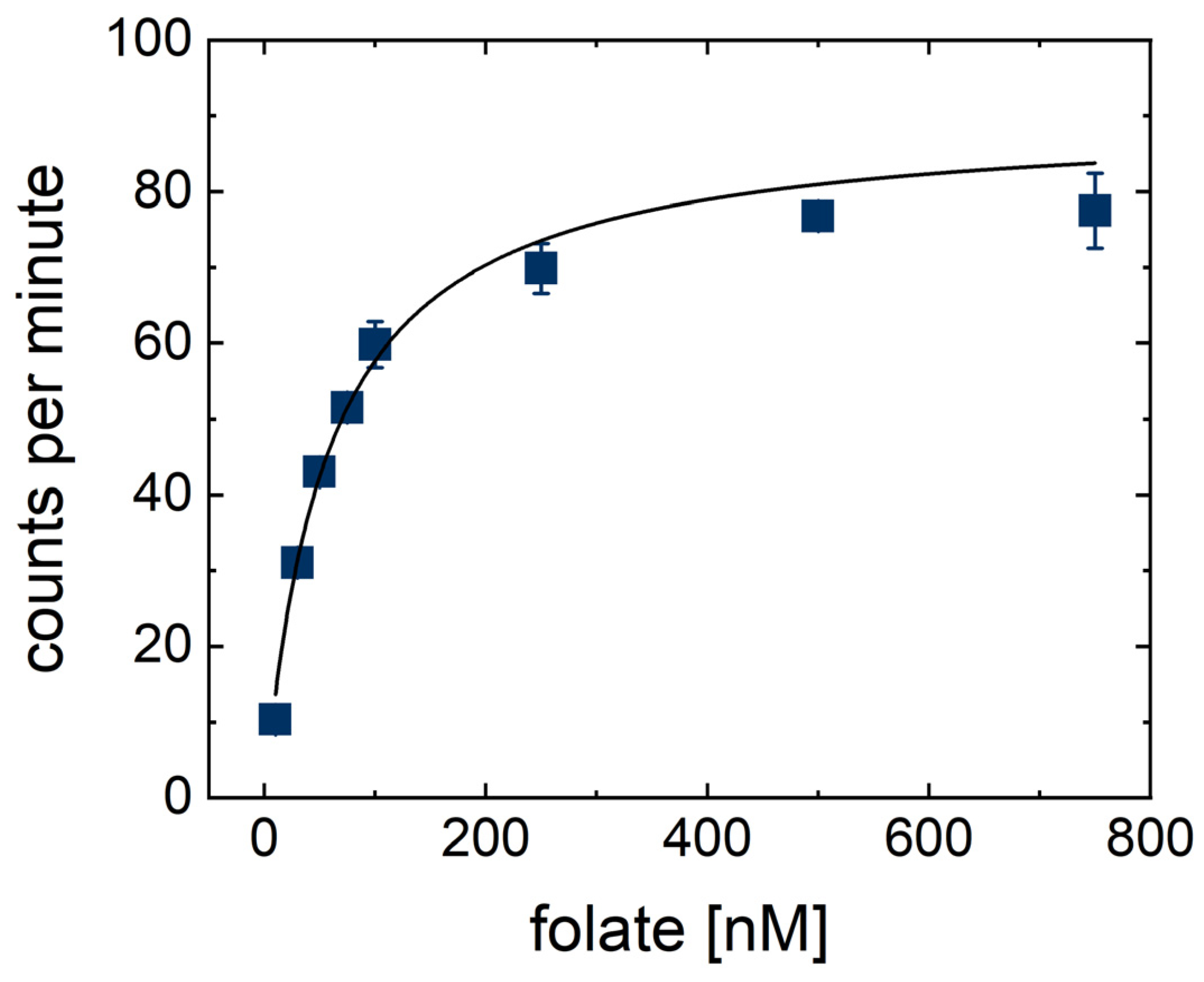
2. Results
3. Discussion
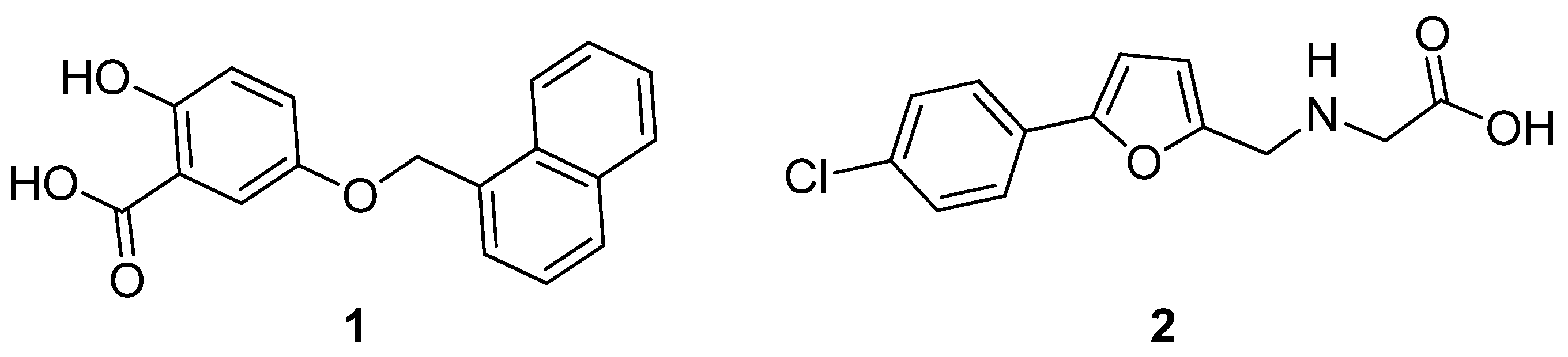
Structure–Activity Relationship (SAR) Study
4. Materials and Methods
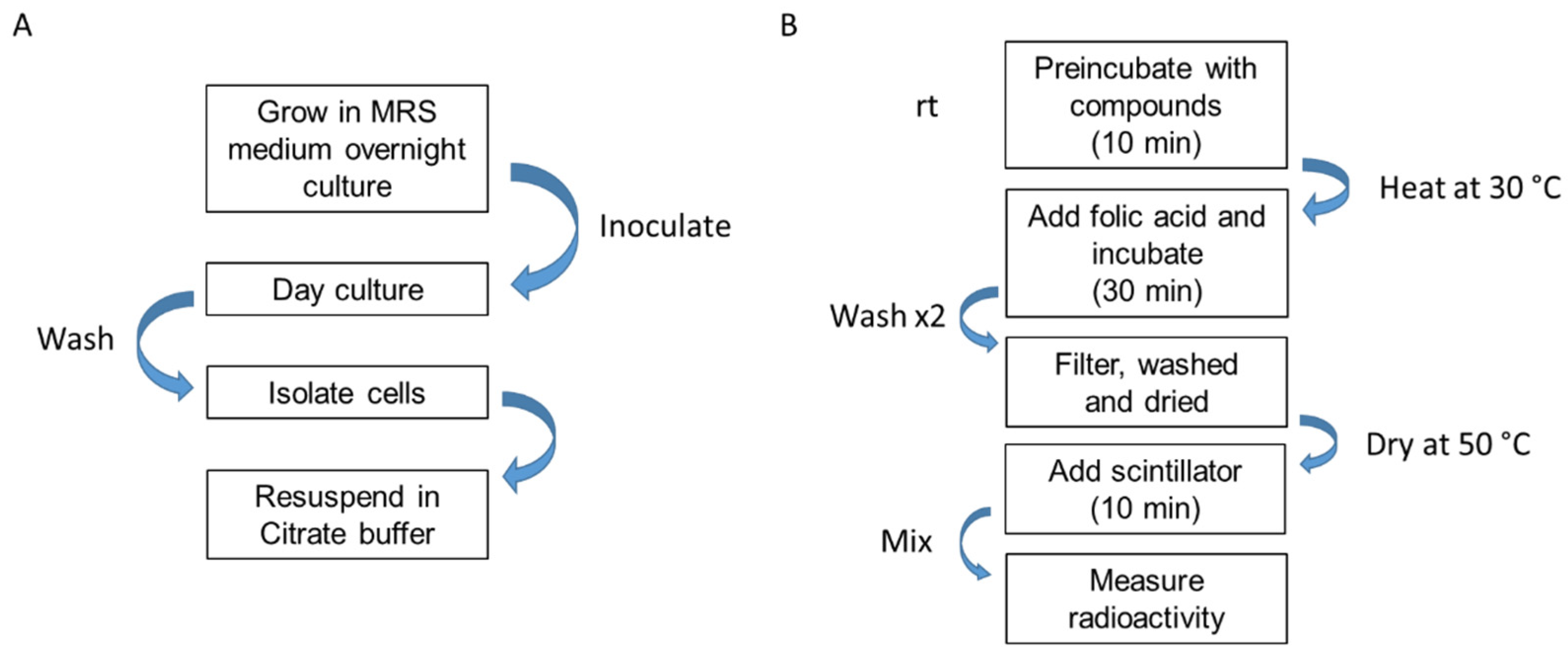
ECF-T Inhibition Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Slotboom, D.J. Structural and mechanistic insights into prokaryotic energy-coupling factor transporters. Nat. Rev. Microbiol. 2014, 12, 79–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodionov, D.A.; Hebbeln, P.; Eudes, A.; ter Beek, J.; Rodionova, I.A.; Erkens, G.B.; Slotboom, D.J.; Gelfand, M.S.; Osterman, A.L.; Hanson, A.D.; et al. Eitinger A novel class of modular transporters for vitamins in prokaryotes. J. Bacteriol. 2009, 191, 42–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodionov, D.A.; Hebbeln, P.; Gelfand, M.S.; Eitinger, T. Comparative and functional genomic analysis of prokaryotic nickel and cobalt uptake transporters: Evidence for a novel group of ATP-binding cassette transporters. J. Bacteriol. 2006, 188, 317–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eudes, A.; Erkens, G.B.; Slotboom, D.J.; Rodionov, D.A.; Naponelli, V.; Hanson, A.D. Identification of genes encoding the folate- and thiamine-binding membrane proteins in Firmicutes. J. Bacteriol. 2008, 190, 7591–7594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bousis, S.; Setyawati, I.; Diamanti, E.; Slotboom, D.J.; Hirsch, A.K.H. Energy-Coupling Factor Transporters as Novel Antimicrobial Targets. Adv. Therap. 2018, 233, 1800066. [Google Scholar] [CrossRef] [Green Version]
- Rice, L.B. Federal funding for the study of antimicrobial resistance in nosocomial pathogens: No ESKAPE. J. Infect. Dis. 2008, 197, 1079–1081. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Priority Pathogens List for R&D of New Antibiotics. Available online: http://www.who.int/en/news-room/detail/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 27 February 2017).
- Henderson, G.B.; Zevely, E.M.; Huennekens, F.M. Mechanism of folate transport in Lactobacillus casei: Evidence for a component shared with the thiamine and biotin transport systems. J. Bacteriol. 1979, 137, 1308–1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diamanti, E.; Setyawati, I.; Bousis, S.; Souza, P.C.T.; Gómez, L.M.; Swier, L.J.Y.M.; Haupenthal, J.; Gibson, P.S.; Volz, C.; Stanek, W.K.; et al. Targeting the Energy-Coupling Factor (ECF) Transporters: Identification of New Tool Compounds. Available online: https://0-doi-org.brum.beds.ac.uk/10.26434/chemrxiv-2021-xq08b-v2 (accessed on 1 January 2022). [CrossRef]
- Kiefer, A.; Bousis, S.; Hamed, M.M.; Diamanti, E.; Haupenthal, J.; Hirsch, A.K.H. Structure-Guided Optimization of Small-Molecule Folate Uptake Inhibitors Targeting the Energy-Coupling Factor Transporters. Available online: https://0-doi-org.brum.beds.ac.uk/10.26434/chemrxiv-2021-gstjz (accessed on 1 January 2022). [CrossRef]
- Henderson, G.B.; Zevely, E.M.; Huennekens, F.M. Coupling of energy to folate transport in Lactobacillus casei. J. Bacteriol. 1979, 139, 552–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drost, M.; Diamanti, E.; Fuhrmann, K.; Goes, A.; Shams, A.; Haupenthal, J.; Koch, M.; Hirsch, A.K.H.; Fuhrmann, G. Bacteriomimetic Liposomes Improve Antibiotic Activity of a Novel Energy-Coupling Factor Transporter Inhibitor. Pharmaceutics 2022, 14, 4. [Google Scholar] [CrossRef] [PubMed]
- The UniProt Consortium. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; UGENE Team. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Uptake Assay | Uptake into Proteoliposomes |
|---|---|
| Medium to high throughput | Low throughput |
| No protein purification | Requires protein purification |
| Multiparameter optimization | Single-parameter optimization |
| Monitor the uptake of several vitamins | Monitor the uptake of specific vitamin |
| Name | IC50 | Assay Type of ECF-FolT |
|---|---|---|
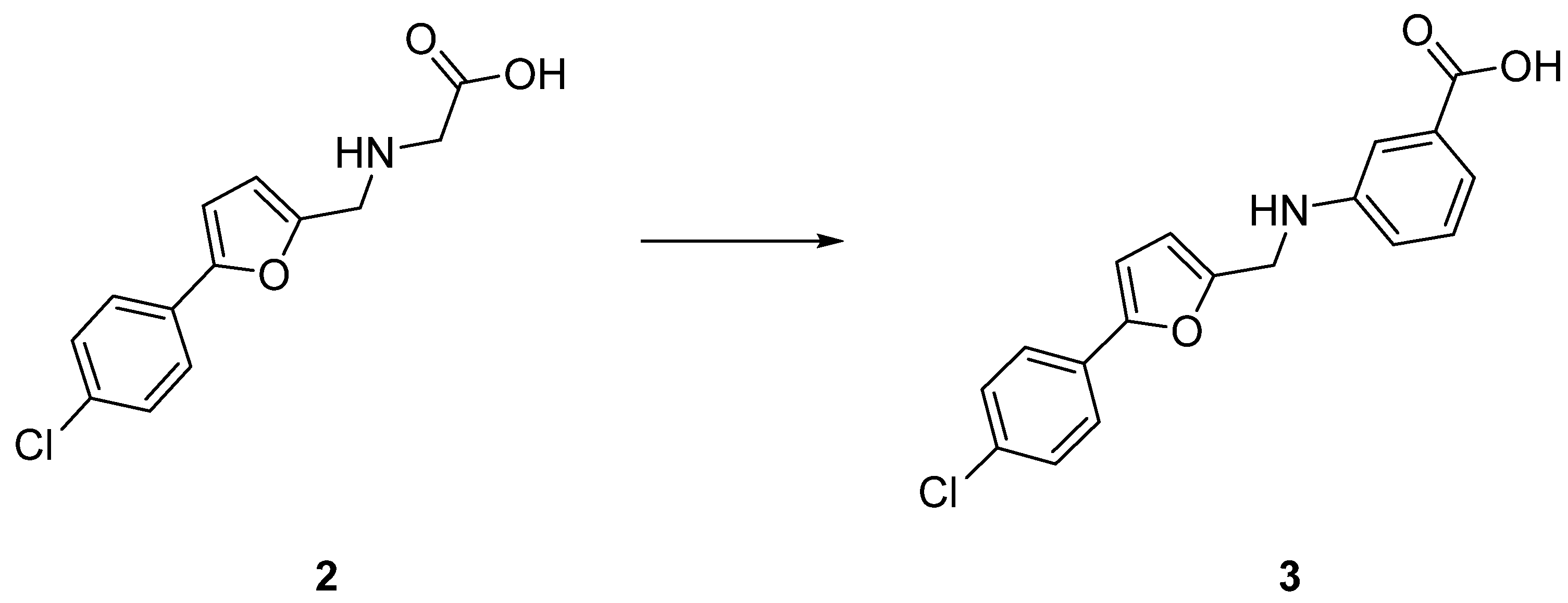
| 2 | >1.2 mM | Proteoliposome uptake |
| 3 | 204 ± 22 µM | Bacterial uptake |

| Name | R1 | R2 | R3 | R4 | R5 | R6 | Inhibition at 200 μM |
|---|---|---|---|---|---|---|---|
| 3 | Cl | H | H | H | COOH | H | 43% (IC50 = 204 ± 22 μM) |
| 231 | Cl | H | H | COOH | H | H | 11% at 50 µM |
| 24 | Cl | H | H | H | H | COOH | n.i at 500 μM |
| 25 | H | Cl | H | H | COOH | H | 41% |
| 26 | H | H | Cl | H | COOH | H | 36% |

| Name | Ar | Inhibition at 200 μM |
|---|---|---|
| 3 |  | 43% |
| 27 |  | 8% |
| Indicator Strain | Auxotrophic | MIC (μg/mL) |
|---|---|---|
| Staphylococcus aureus str. Newman | no | 23 |
| Streptococcus pneumoniae DSM-20566 | yes | 21 |
| Streptococcus pneumoniae DSM-11865 1 | yes | 10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bousis, S.; Winkler, S.; Haupenthal, J.; Fulco, F.; Diamanti, E.; Hirsch, A.K.H. An Efficient Way to Screen Inhibitors of Energy-Coupling Factor (ECF) Transporters in a Bacterial Uptake Assay. Int. J. Mol. Sci. 2022, 23, 2637. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052637
Bousis S, Winkler S, Haupenthal J, Fulco F, Diamanti E, Hirsch AKH. An Efficient Way to Screen Inhibitors of Energy-Coupling Factor (ECF) Transporters in a Bacterial Uptake Assay. International Journal of Molecular Sciences. 2022; 23(5):2637. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052637
Chicago/Turabian StyleBousis, Spyridon, Steffen Winkler, Jörg Haupenthal, Francesco Fulco, Eleonora Diamanti, and Anna K. H. Hirsch. 2022. "An Efficient Way to Screen Inhibitors of Energy-Coupling Factor (ECF) Transporters in a Bacterial Uptake Assay" International Journal of Molecular Sciences 23, no. 5: 2637. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052637