
Generation of a Dystrophin Mutant in Dog by Nuclear Transfer Using CRISPR/Cas9-Mediated Somatic Cells: A Preliminary Study

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
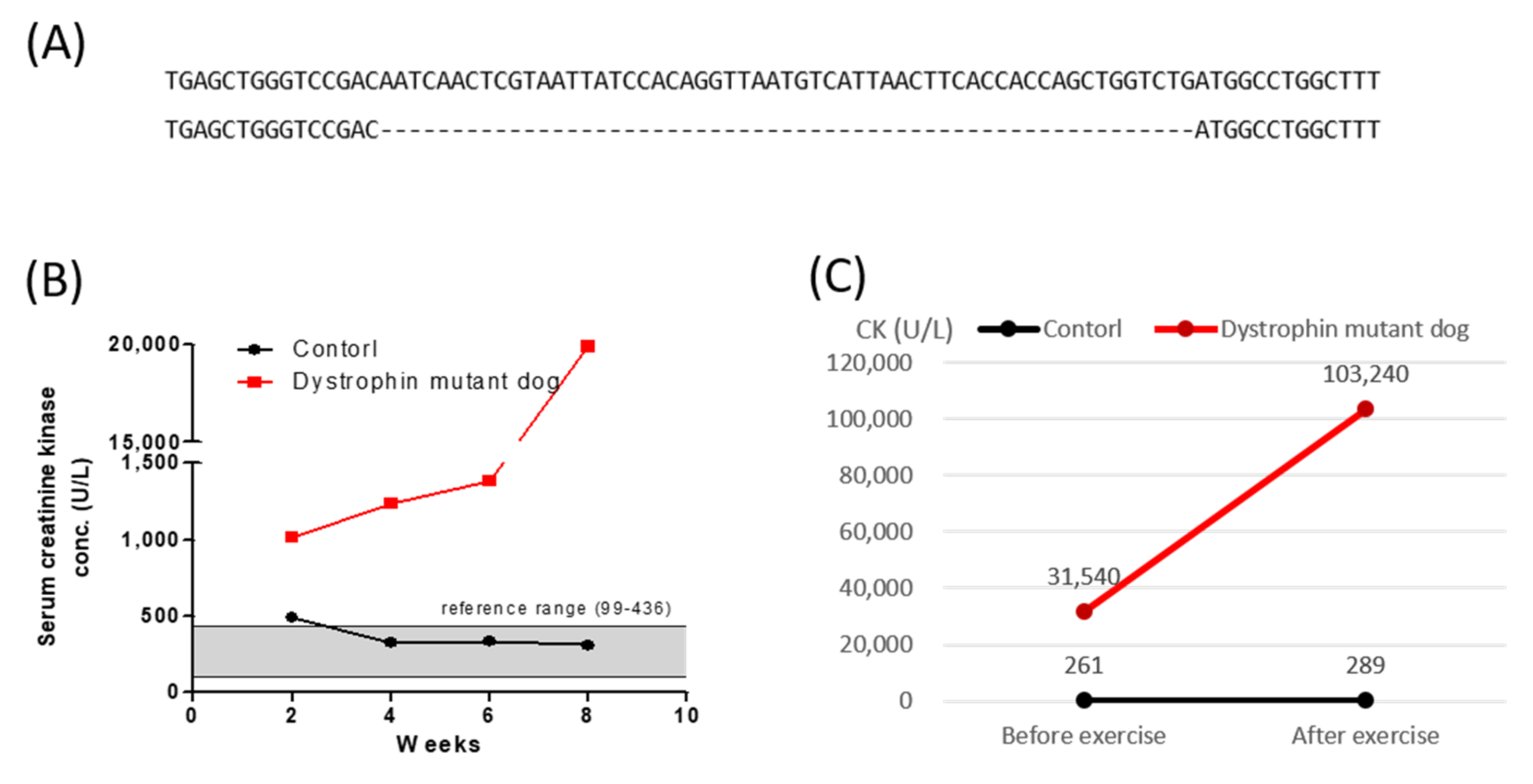
2.1. Generation of Dystrophin Mutant Cloned Dog by SCNT
2.2. Creatine Kinase and Electrocardiographic Analysis
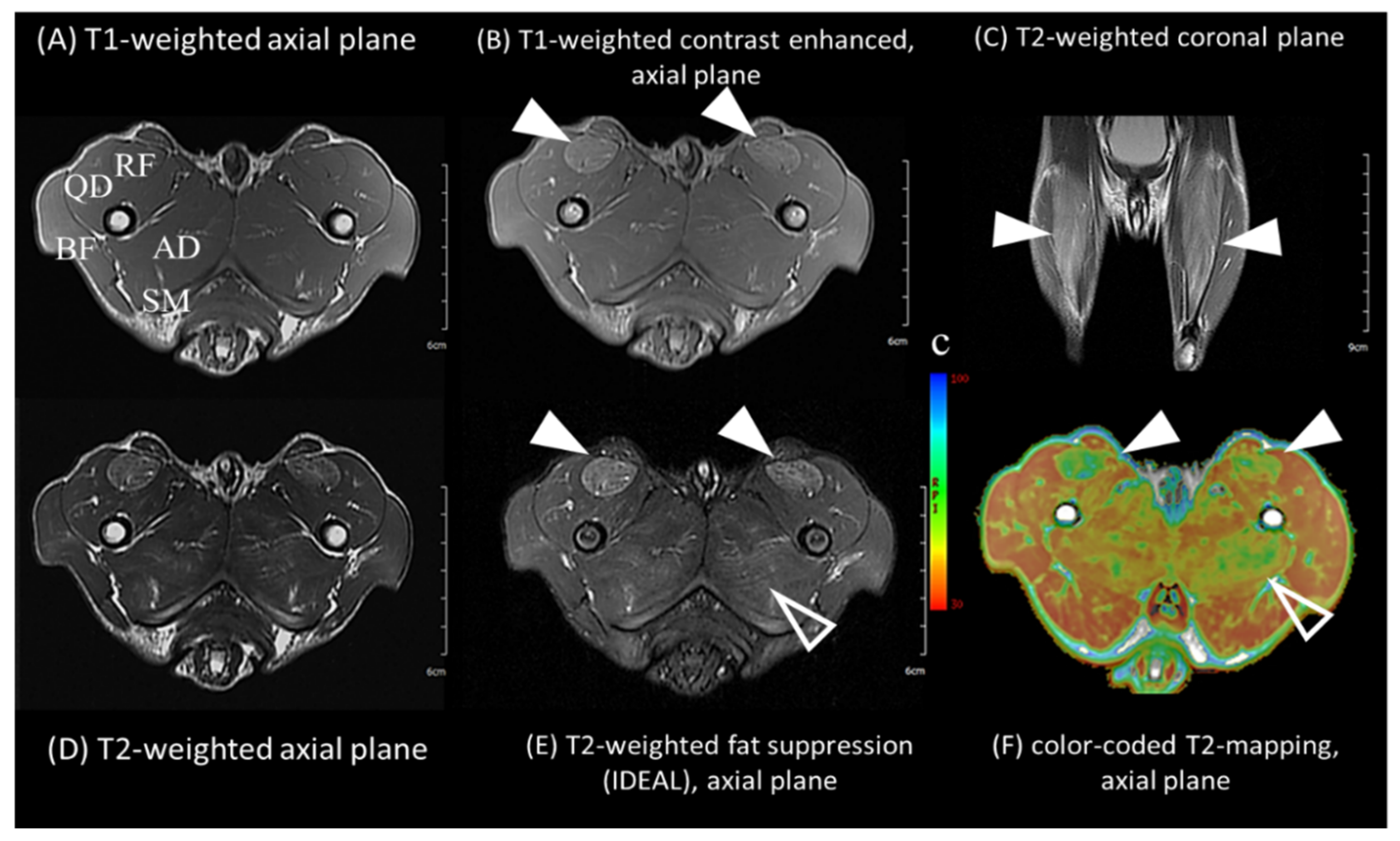
2.3. Muscle Magnetic Resonance Imaging
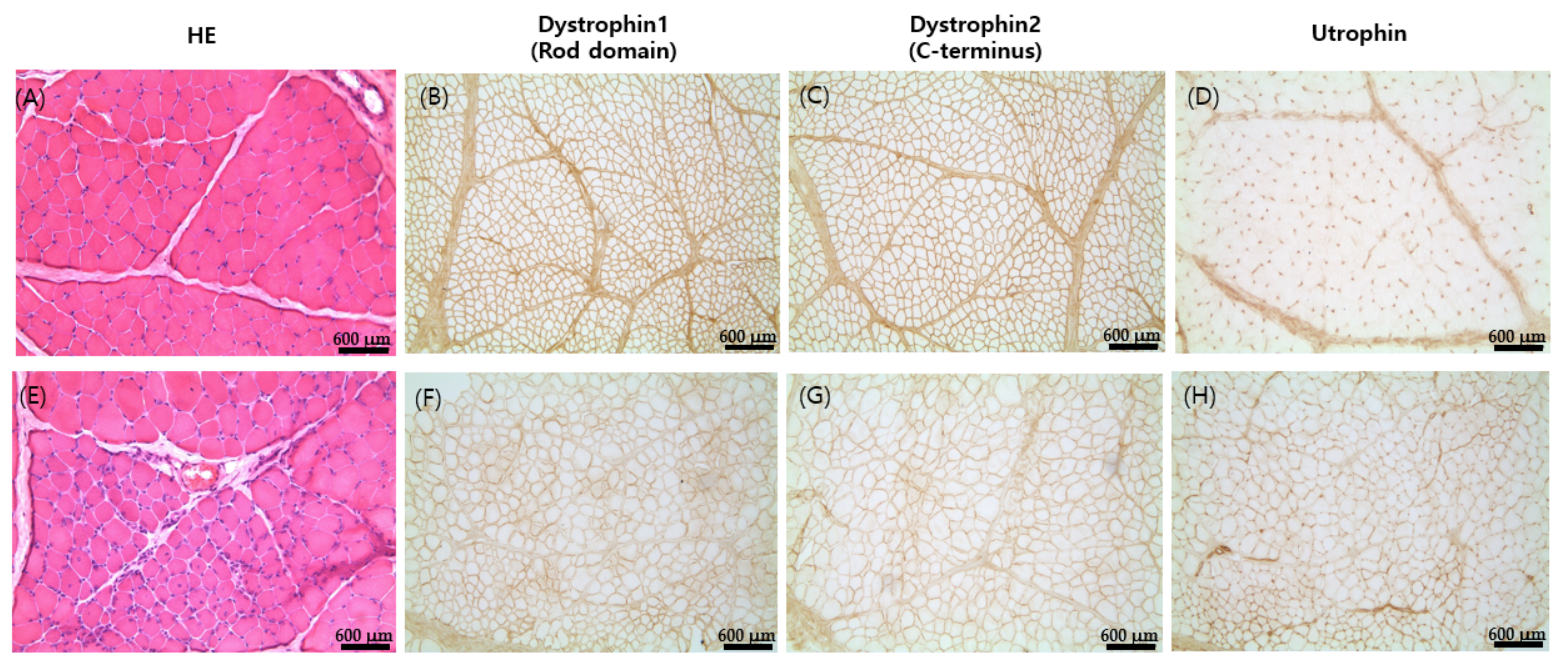
2.4. Histopathological Analysis
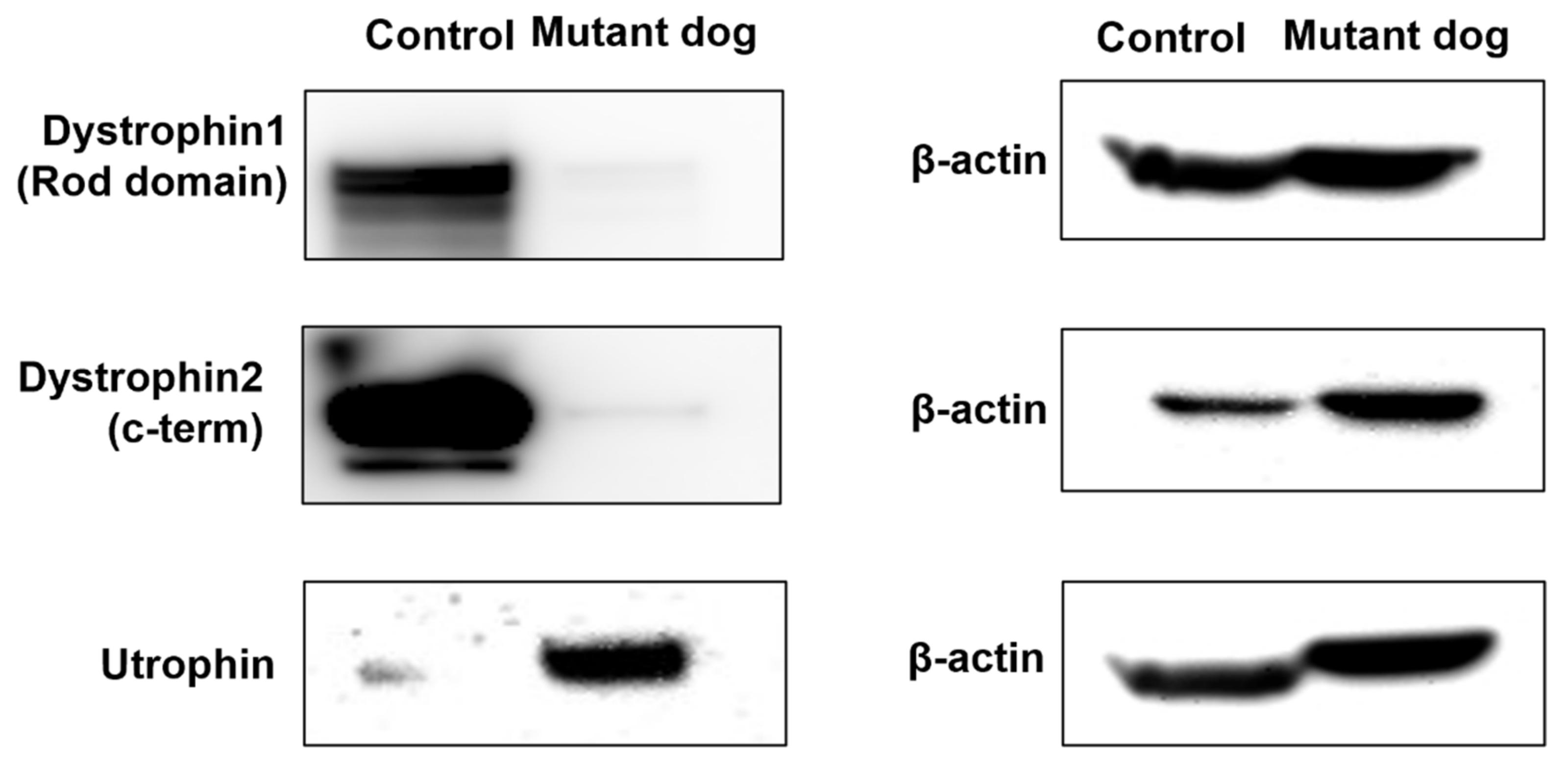
2.5. Western Blotting
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Generation of CRISPR/Cas9
4.3. Cell Culture and Transfections
4.4. In Vivo Matured Oocyte Collection
4.5. Somatic Cell Nuclear Transfer and Embryo Transfer
4.6. T7 Endonuclease I Assay and Sequencing
4.7. Deep Sequencing Analysis
4.8. Creatine Kinase
4.9. Anesthesia for Magnetic Resonance Imaging
4.10. Non-Quantitative (Conventional) Magnetic Resonance Imaging
4.11. Quantitative (T2 Mapping) Magnetic Resonance Imaging
4.12. Biopsy and Histopathological Analysis
4.13. Western Blotting
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Wayne, R.K.; Ostrander, E.A. Lessons learned from the dog genome. Trends Genet. 2007, 23, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Glickman, L.T.; Raghavan, M.; Knapp, D.W.; Bonney, P.L.; Dawson, M.H. Herbicide exposure and the risk of transitional cell carcinoma of the urinary bladder in Scottish Terriers. J. Am. Vet. Med. Assoc. 2004, 224, 1290–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, E.P.; Brown, R.H., Jr.; Kunkel, L.M. Dystrophin: The protein product of the Duchenne muscular dystrophy locus. Cell 1987, 51, 919–928. [Google Scholar] [CrossRef]
- Guiraud, S.; Aartsma-Rus, A.; Vieira, N.M.; Davies, K.E.; van Ommen, G.J.; Kunkel, L.M. The Pathogenesis and Therapy of Muscular Dystrophies. Annu. Rev. Genom. Hum. Genet. 2015, 16, 281–308. [Google Scholar] [CrossRef] [Green Version]
- Shelton, G.D.; Engvall, E. Canine and feline models of human inherited muscle diseases. Neuromuscul. Disord. 2005, 15, 127–138. [Google Scholar] [CrossRef]
- Chamberlain, J.S.; Metzger, J.; Reyes, M.; Townsend, D.; Faulkner, J.A. Dystrophin-deficient mdx mice display a reduced life span and are susceptible to spontaneous rhabdomyosarcoma. FASEB J. 2007, 21, 2195–2204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGreevy, J.W.; Hakim, C.H.; McIntosh, M.A.; Duan, D. Animal models of Duchenne muscular dystrophy: From basic mechanisms to gene therapy. Dis. Model. Mech. 2015, 8, 195–213. [Google Scholar] [CrossRef] [Green Version]
- Duan, D. Duchenne muscular dystrophy gene therapy in the canine model. Hum. Gene Clin. Dev. 2015, 26, 57–69. [Google Scholar] [CrossRef]
- Duan, D. Duchenne muscular dystrophy gene therapy: Lost in translation? Res. Rep. Biol. 2011, 2011, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Valentine, B.A.; Cooper, B.J.; de Lahunta, A.; O’Quinn, R.; Blue, J.T. Canine X-linked muscular dystrophy. An animal model of Duchenne muscular dystrophy: Clinical studies. J. Neurol. Sci. 1988, 88, 69–81. [Google Scholar] [CrossRef]
- Kornegay, J.N. The golden retriever model of Duchenne muscular dystrophy. Skelet. Muscle 2017, 7, 9. [Google Scholar] [CrossRef] [PubMed]
- Sharp, N.J.; Kornegay, J.N.; Van Camp, S.D.; Herbstreith, M.H.; Secore, S.L.; Kettle, S.; Hung, W.Y.; Constantinou, C.D.; Dykstra, M.J.; Roses, A.D.; et al. An error in dystrophin mRNA processing in golden retriever muscular dystrophy, an animal homologue of Duchenne muscular dystrophy. Genomics 1992, 13, 115–121. [Google Scholar] [CrossRef]
- Schatzberg, S.J.; Olby, N.J.; Breen, M.; Anderson, L.V.B.; Langford, C.F.; Dickens, H.F.; Wilton, S.D.; Zeiss, C.J.; Binns, M.M.; Kornegay, J.N.; et al. Molecular analysis of a spontaneous dystrophin ‘knockout’ dog. Neuromuscul. Disord. 1999, 9, 289–295. [Google Scholar] [CrossRef]
- Walmsley, G.L.; Arechavala-Gomeza, V.; Fernandez-Fuente, M.; Burke, M.M.; Nagel, N.; Holder, A.; Stanley, R.; Chandler, K.; Marks, S.L.; Muntoni, F.; et al. A Duchenne Muscular Dystrophy Gene Hot Spot Mutation in Dystrophin-Deficient Cavalier King Charles Spaniels Is Amenable to Exon 51 Skipping. PLoS ONE 2010, 5, e8647. [Google Scholar] [CrossRef]
- Shimatsu, Y.; Katagiri, K.; Furuta, T.; Nakura, M.; Tanioka, Y.; Yuasa, K.; Tomohiro, M.; Kornegay, J.N.; Nonaka, I.; Takeda, S. Canine X-linked muscular dystrophy in Japan (CXMDj). Exp. Anim. 2003, 52, 93–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.L.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.B.; Jiang, W.Y.; Marraffini, L.A.; et al. Multiplex Genome Engineering Using CRISPR/Cas Systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mali, P.; Yang, L.H.; Esvelt, K.M.; Aach, J.; Guell, M.; DiCarlo, J.E.; Norville, J.E.; Church, G.M. RNA-Guided Human Genome Engineering via Cas9. Science 2013, 339, 823–826. [Google Scholar] [CrossRef] [Green Version]
- Ni, W.; Qiao, J.; Hu, S.W.; Zhao, X.X.; Regouski, M.; Yang, M.; Polejaeva, I.A.; Chen, C.F. Efficient Gene Knockout in Goats Using CRISPR/Cas9 System. PLoS ONE 2014, 9, e106718. [Google Scholar] [CrossRef] [Green Version]
- Han, H.; Wang, T.; Lian, L.; Tian, X.; Hu, R.; Deng, S.; Li, K.; Wang, F.; Li, N.; Liu, G.; et al. One-step generation of myostatin gene knockout sheep via the CRISPR/Cas9 system. Front. Agric. Sci. Eng. 2014, 1, 2–5. [Google Scholar]
- Kim, G.A.; Oh, H.J.; Kim, M.J.; Jo, Y.K.; Choi, J.; Park, J.E.; Park, E.J.; Lim, S.H.; Yoon, B.I.; Kang, S.K.; et al. Survival of Skin Graft between Transgenic Cloned Dogs and Non-Transgenic Cloned Dogs. PLoS ONE 2014, 9, e108330. [Google Scholar] [CrossRef]
- Skrzyszowska, M.; Samiec, M. Generating Cloned Goats by Somatic Cell Nuclear Transfer-Molecular Determinants and Application to Transgenics and Biomedicine. Int. J. Mol. Sci. 2021, 22, 7490. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Zheng, Y.H.; Kang, Y.; Yang, W.L.; Niu, Y.Y.; Guo, X.Y.; Tu, Z.C.; Si, C.Y.; Wang, H.; Xing, R.X.; et al. Functional disruption of the dystrophin gene in rhesus monkey using CRISPR/Cas9. Hum. Mol. Genet. 2015, 24, 3764–3774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.Q.; Xin, J.G.; Fan, N.N.; Zou, Q.J.; Huang, J.; Ouyang, Z.; Zhao, Y.; Zhao, B.T.; Liu, Z.M.; Lai, S.S.; et al. Generation of CRISPR/Cas9-mediated gene-targeted pigs via somatic cell nuclear transfer. Cell Mol. Life Sci. 2015, 72, 1175–1184. [Google Scholar] [CrossRef] [PubMed]
- Eun, K.; Hong, N.; Jeong, Y.W.; Park, M.G.; Hwang, S.U.; Jeong, Y.I.K.; Choi, E.J.; Olsson, P.O.; Hwang, W.S.; Hyun, S.H.; et al. Transcriptional activities of human elongation factor-1alpha and cytomegalovirus promoter in transgenic dogs generated by somatic cell nuclear transfer. PLoS ONE 2020, 15, e0233784. [Google Scholar] [CrossRef]
- Li, P.; Estrada, J.L.; Burlak, C.; Montgomery, J.; Butler, J.R.; Santos, R.M.; Wang, Z.Y.; Paris, L.L.; Blankenship, R.L.; Downey, S.M.; et al. Efficient generation of genetically distinct pigs in a single pregnancy using multiplexed single-guide RNA and carbohydrate selection. Xenotransplantation 2015, 22, 20–31. [Google Scholar] [CrossRef]
- Chen, F.J.; Wang, Y.; Yuan, Y.L.; Zhang, W.; Ren, Z.J.; Jin, Y.; Liu, X.R.; Xiong, Q.; Chen, Q.; Zhang, M.L.; et al. Generation of B Cell-Deficient Pigs by Highly Efficient CRISPR/Cas9-Mediated Gene Targeting. J. Genet. Genom. 2015, 42, 437–444. [Google Scholar] [CrossRef]
- Samiec, M.; Skrzyszowska, M. The possibilities of practical application of transgenic mammalian species generated by somatic cell cloning in pharmacology, veterinary medicine and xenotransplantology. Pol. J. Vet. Sci. 2011, 14, 329–340. [Google Scholar] [CrossRef] [Green Version]
- Feng, C.; Wang, X.M.; Shi, H.; Yan, Q.M.; Zheng, M.; Li, J.; Zhang, Q.J.; Qin, Y.M.; Zhong, Y.G.; Mi, J.D.; et al. Generation of ApoE deficient dogs via combination of embryo injection of CRISPR/Cas9 with somatic cell nuclear transfer. J. Genet. Genom. 2018, 45, 47–50. [Google Scholar] [CrossRef]
- Zou, Q.J.; Wang, X.M.; Liu, Y.Z.; Ouyang, Z.; Long, H.B.; Wei, S.; Xin, J.G.; Zhao, B.T.; Lai, S.S.; Shen, J.; et al. Generation of gene-target dogs using CRISPR/Cas9 system. J. Mol. Cell Biol. 2015, 7, 580–583. [Google Scholar] [CrossRef] [Green Version]
- Bae, S.; Park, J.; Kim, J.S. Cas-OFFinder: A fast and versatile algorithm that searches for potential off-target sites of Cas9 RNA-guided endonucleases. Bioinformatics 2014, 30, 1473–1475. [Google Scholar] [CrossRef] [Green Version]
- Aartsma-Rus, A.; Van Deutekom, J.C.T.; Fokkema, I.F.; Van Ommen, G.J.B.; Den Dunnen, J.T. Entries in the Leiden Duchenne muscular dystrophy mutation database: An overview of mutation types and paradoxical cases that confirm the reading-frame rule. Muscle Nerve 2006, 34, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Kornegay, J.N.; Cundiff, D.D.; Bogan, D.J.; Bogan, J.R.; Okamura, C.S. The cranial sartorius muscle undergoes true hypertrophy in dogs with golden retriever muscular dystrophy. Neuromuscul. Disord. 2003, 13, 493–500. [Google Scholar] [CrossRef]
- Rowland, L.P. Biochemistry of muscle membranes in Duchenne muscular dystrophy. Muscle Nerve 1980, 3, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Manera, J.; Llauger, J.; Gallardo, E.; Illa, I. Muscle MRI in muscular dystrophies. Acta Myol. 2015, 34, 95–108. [Google Scholar]
- Finanger, E.L.; Russman, B.; Forbes, S.C.; Rooney, W.D.; Walter, G.A.; Vandenborne, K. Use of Skeletal Muscle MRI in Diagnosis and Monitoring Disease Progression in Duchenne Muscular Dystrophy. Phys. Med. Rehabil. Clin. N. Am. 2012, 23, 1. [Google Scholar] [CrossRef] [Green Version]
- Maron, B.J.; Towbin, J.A.; Thiene, G.; Antzelevitch, C.; Corrado, D.; Arnett, D.; Moss, A.J.; Seidman, C.E.; Young, J.B. Contemporary definitions and classification of the cardiomyopathies—An American Heart Association Scientific Statement from the Council on Clinical Cardiology, Heart Failure and Transplantation Committee; Quality of Care and Outcomes Research and Functional Genomics and Translational Biology Interdisciplinary Working Groups; And Council on Epidemiology and Prevention. Circulation 2006, 113, 1807–1816. [Google Scholar]
- McNally, E.M. Duchenne muscular dystrophy: How bad is the heart? Heart 2008, 94, 976–977. [Google Scholar] [CrossRef]
- Yugeta, N.; Urasawa, N.; Fujii, Y.; Yoshimura, M.; Yuasa, K.; Wada, M.R.; Nakura, M.; Shimatsu, Y.; Tomohiro, M.; Takahashi, A.; et al. Cardiac involvement in Beagle-based canine X-linked muscular dystrophy in Japan (CXMDJ): Electrocardiographic, echocardiographic, and morphologic studies. BMC Cardiovasc. Disord. 2006, 6, 47. [Google Scholar] [CrossRef] [Green Version]
- Arahata, K.; Ishiura, S.; Ishiguro, T.; Tsukahara, T.; Suhara, Y.; Eguchi, C.; Ishihara, T.; Nonaka, I.; Ozawa, E.; Sugita, H. Immunostaining of skeletal and cardiac muscle surface membrane with antibody against Duchenne muscular dystrophy peptide. Nature 1988, 333, 861–863. [Google Scholar] [CrossRef]
- Monaco, A.P.; Bertelson, C.J.; Liechti-Gallati, S.; Moser, H.; Kunkel, L.M. An explanation for the phenotypic differences between patients bearing partial deletions of the DMD locus. Genomics 1988, 2, 90–95. [Google Scholar] [CrossRef]
- Amoasii, L.; Hildyard, J.C.W.; Li, H.; Sanchez-Ortiz, E.; Mireault, A.; Caballero, D.; Harron, R.; Stathopoulou, T.R.; Massey, C.; Shelton, J.M.; et al. Gene editing restores dystrophin expression in a canine model of Duchenne muscular dystrophy. Science 2018, 362, 86–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Recipient | No. In Vivo Matured Oocytes | No. Reconstructed Oocytes | No. Transferred Embryos | Pregnancy | No. Births |
|---|---|---|---|---|---|
| A | 16 | 6 | 6 | + | 1 |
| B | 13 | 7 | 7 | - | 0 |
| C | 20 | 13 | 13 | - | 0 |
| Total | 49 | 26 | 26 | 1 (33.3%) ∫ | 1 (3.84%) ∫∫ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oh, H.J.; Chung, E.; Kim, J.; Kim, M.J.; Kim, G.A.; Lee, S.H.; Ra, K.; Eom, K.; Park, S.; Chae, J.-H.; et al. Generation of a Dystrophin Mutant in Dog by Nuclear Transfer Using CRISPR/Cas9-Mediated Somatic Cells: A Preliminary Study. Int. J. Mol. Sci. 2022, 23, 2898. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052898
Oh HJ, Chung E, Kim J, Kim MJ, Kim GA, Lee SH, Ra K, Eom K, Park S, Chae J-H, et al. Generation of a Dystrophin Mutant in Dog by Nuclear Transfer Using CRISPR/Cas9-Mediated Somatic Cells: A Preliminary Study. International Journal of Molecular Sciences. 2022; 23(5):2898. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052898
Chicago/Turabian StyleOh, Hyun Ju, Eugene Chung, Jaehwan Kim, Min Jung Kim, Geon A. Kim, Seok Hee Lee, Kihae Ra, Kidong Eom, Soojin Park, Jong-Hee Chae, and et al. 2022. "Generation of a Dystrophin Mutant in Dog by Nuclear Transfer Using CRISPR/Cas9-Mediated Somatic Cells: A Preliminary Study" International Journal of Molecular Sciences 23, no. 5: 2898. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052898