
The Sleep-Promoting Ventrolateral Preoptic Nucleus: What Have We Learned over the Past 25 Years?

1
Department of Neurology, Beth Israel Deaconess Medical Center, Harvard Medical School, Boston, MA 02215, USA
2
Department of Neurological Surgery, University of California, Davis School of Medicine, Davis, CA 95618, USA
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2022, 23(6), 2905; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23062905
Submission received: 31 January 2022
/
Revised: 28 February 2022
/
Accepted: 2 March 2022
/
Published: 8 March 2022
(This article belongs to the Special Issue Molecular Circuits Regulating Sleep and Wakeful Consciousness)
Abstract
:For over a century, the role of the preoptic hypothalamus and adjacent basal forebrain in sleep–wake regulation has been recognized. However, for years, the identity and location of sleep- and wake-promoting neurons in this region remained largely unresolved. Twenty-five years ago, Saper and colleagues uncovered a small collection of sleep-active neurons in the ventrolateral preoptic nucleus (VLPO) of the preoptic hypothalamus, and since this seminal discovery the VLPO has been intensively investigated by labs around the world, including our own. Herein, we first review the history of the preoptic area, with an emphasis on the VLPO in sleep–wake control. We then attempt to synthesize our current understanding of the circuit, cellular and synaptic bases by which the VLPO both regulates and is itself regulated, in order to exert a powerful control over behavioral state, as well as examining data suggesting an involvement of the VLPO in other physiological processes.
1. The Hypothalamus and Sleep: A Brief History
During the epidemics of encephalitis lethargica, which both preceded and followed the 1918 influenza pandemic, the Austrian neurologist Baron Constantin von Economo undertook a series of clinicopathologic studies on individuals who were rendered insomniac by this previously unrecognized type of encephalitis [1,2]. What he discovered during his post mortem analysis of the brains of these insomniac individuals was the presence of inflammatory lesions involving the anterior hypothalamus and, to varying degrees, the adjacent basal ganglia, the latter of which would explain the chorea that was often associated with insomnia. Based upon these findings, von Economo hypothesized that the anterior hypothalamic region comprised a “schlafsteuerungszentrum” or “sleep center”. He further hypothesized that this sleep center actively inhibited the cerebral cortex and thalamus to produce, in his words, “brain sleep”. Less than a decade later, Hess reported that he could produce sleep in cats by stimulating various regions of the hypothalamus and midbrain [3], although his findings stood at variance with those of Ranson and colleagues [4,5,6], who were never able to produce sleep in cats or monkeys following scattered lesions of the hypothalamus. In 1946, Nauta undertook an extensive and detailed study of the role of the hypothalamus in the regulation of sleep, in part an attempt to reconcile the findings of von Economo, Ranson and Hess [7]. None of Nauta’s control lesions, which included incisions dorsal to the ventral thalamus and unilateral cuts to the hypothalamus, resulted in insomnia, nor did they result in marked changes in the sleep–wake cycle. Nauta then employed a parapharyngeal approach, along with a “tour de maître”, a small hook-shaped knife, to mechanically and systematically inflict “transverse” lesions in the rat hypothalamus. Nauta found that his “lesions”, when situated in the caudal hypothalamus, tended to produce mild-to-extreme sleepiness, whereas lesions encompassing the rostral lateral hypothalamus produced a state of insomnia. From this, Nauta concluded that the rostral half of the hypothalamus “roughly conforming to the suprachiasmatic and preoptic areas, is the site of a nervous structure which is of specific importance for the capacity of sleep”; he henceforth referred to this structure as a “sleep center”. Nauta also concluded via concurrent lesions to the rostral and caudal hypothalamus that active inhibition of the waking center(s) by the sleep center is how sleep is caused—his interpretation of the data was a prescient one and something we return to later in this review (see Section 5). A little over two decades later, in 1968, McGinty and Sterman reported that lesions of the basal forebrain (which included the tissue of the medially adjacent preoptic region) produced sleeplessness in cats, largely recapitulating Nauta’s findings [8].
2. Discovery of the Sleep-Active VLPO
A true watershed moment in the field of sleep science occurred when Saper and colleagues uncovered a bona fide ‘sleep center’ in the rostral hypothalamus, a previously unidentified “sleep-active” cell group in the ventrolateral preoptic area (VLPO) [9] of rats (See Table 1 for key references). The clearest density of these sleep-active VLPO neurons was located just lateral to the optic chiasm at the level of the crossing of the anterior commissure. Following the identification and anatomic characterization of the cellular VLPO, recordings made from other groups in the preoptic area, including presumptive VLPO neurons, revealed that many of these neurons fired more rapidly during sleep than during wakefulness, and more rapidly still when animals were recovering from sleep deprivation [10,11]. Some of the recorded VLPO neurons also showed a wake-active profile, although their role in sleep–wake regulation remains uncertain. These electrophysiological results correlated well with the finding that VLPO neurons accumulate cFos protein during sleep, but not during the accumulation of “sleep need” during prolonged wakefulness [9,12]. Subsequent single-unit recording studies found that VLPO neurons also start firing during sleep deprivation, a finding that was at some variance with the cFos studies [13]. To demonstrate that VLPO neurons were necessary to promote sleep, Lu and colleagues placed cell-specific lesions in the VLPO area in rats. Rats with verified bilateral VLPO lesions had a reduction of ~40–50% in total sleep time, along with sleep fragmentation; this was sustained for the post surgical survival period, which in some cases, was months [14]. These lesion findings were repeatedly replicated and have firmly established VLPO neurons as necessary for the maintenance of sleep [15,16,17,18]. However, animals with a complete, bilateral loss of VLPO still exhibit sleep. This is consistent with more recent studies that have identified and characterized other sleep-promoting systems in the brain, which can likely compensate, although clearly incompletely, following ablation of the VLPO. These sleep-promoting nodes include the GABAergic parafacial zone of the rostral medulla [19,20,21] and, most likely, the median preoptic nucleus (MnPO) [22] and are the focus of other reviews [23,24,25].
3. VLPO—Inputs and Outputs
In their initial paper, Saper and colleagues reported that sleep-active VLPO neurons provide heavy innervation of the tuberomammillary nucleus (TMN), an arousal-associated cell group in the caudal hypothalamus [9] (See Table 1 for key references). These authors subsequently showed that TMN-projecting VLPO neurons contain both GABA and the inhibitory peptide galanin, suggesting that the VLPO could exert potent inhibitory control over the TMN [26]. Gaus and colleagues later reported that most sleep-active VLPO neurons express mRNA for galanin in both nocturnal (mice and rats) and diurnal (degus) rodents as well as in cats [27]. On the basis of these findings, the authors hypothesized that GABA- and galanin-containing VLPO neurons might contribute to the sleep process through synaptic silencing of the brain’s arousal centers, which in many respects aligns with Von Economo’s original prediction [26]. Consistent with this theoretical model, it was later shown that VLPO galanin neurons provide innervation of most of the canonical components of the brain’s ascending arousal system, including the lateral hypothalamic orexin neurons, the serotonergic raphe neurons, the noradrenergic locus coeruleus and ventrolateral medullary neurons, and the glutamatergic parabrachial nucleus [28]. It had been previously reported that virtually all of these ‘arousal’ nodes provide reciprocal, and generally inhibitory, inputs to the VLPO [29]. The functional implications of these anatomic projections and connections are discussed below.
4. Cellular Anatomy of the VLPO
The VLPO was initially described as a cluster of sleep-active neurons that co-express GABA and the inhibitory neuropeptide, galanin, and project to cell bodies and proximal dendrites of the TMN [9,27] (See Table 2 for Key References). Subsequent investigative work revealed the VLPO to be a brain region of even greater cellular heterogeneity, containing multiple neurochemically and functionally distinct cell populations. For instance, in addition to galanin, several other peptides (including peptides expected to excite their post synaptic targets) are produced and differentially expressed by VLPO neurons, including cholecystokinin (CCK), corticotropin-releasing hormone (CRH), dynorphin A and substance P [30,31], although the roles of most of these peptides in VLPO function remain unclear. While some degree of cellular overlap in VLPO peptide expressions has also been reported, e.g., some CCK- and substance P-expressing neurons also express CRH [30], the co-localization of peptide within VLPO cells does not appear to be a common feature of this nucleus. For a visual depiction of the cellular anatomy of the VLPO, discussed in more detail below, see Figure 1.
4.1. The GABAergic VLPO
The VLPO appears to contain a large number of GABA-releasing neurons, as indicated by the dense expression of the vesicular GABA transporter (Vgat). VLPO GABAergic neurons, however, comprise at least two distinct subgroups. The first expresses galanin (Gal; VLPOGal), is sleep-active and is inhibited by wake-promoting inputs [13,27,32,33,34,35], while the second lacks galanin (VLPOGABA), is excited by arousal signals and might inhibit VLPOGal neurons to produce arousal [36,37]. The latter and more recently ‘discovered’ second subgroup of VLPOGABA neurons has received considerable attention as of late and is discussed in detail below.
4.2. The Galaninergic VLPO
Galanin is an inhibitory peptide, and upon its co-release with GABA, it is likely to contribute to the inhibition of wake-promoting neurons [38,39]. Supporting this concept are results from in vitro electrophysiology studies showing that the locus coeruleus (LC), TMN, dorsal raphe nucleus (DRN) and orexin neurons are all inhibited by galanin [39,40,41,42] and that all these regions are innervated by VLPO [26,28,43]. Nevertheless, the relative contribution of GABA versus galanin signaling to promoting NREM sleep remains unclear. Answering this question will likely require an intersectional genetic strategy to delete either the Vgat or the galanin genes selectively in galanin-expressing VLPO neurons, but to our knowledge, these tools are not currently available. Regarding whether or not the synaptic release of galanin by VLPO neurons is required to produce sleep, there remains a general consensus that galanin is the best available cellular ‘marker’ within the VLPO for identifying sleep-promoting neurons, and this is true across multiple species [27]. Further indications of strong phylogenetic conservation in the role of this cell group in sleep–wake regulation are found in post mortem studies in humans, which established a close correlation between the number of galanin-immunoreactive neurons in the intermediate nucleus of the hypothalamus (or human homolog of the VLPO) and the amount/percentage of consolidated sleep in aged individuals [44,45].
It is also worth noting that there appear to be at least two functionally distinct galanin-expressing populations in the VLPO region. The first of these subgroups is found within the anatomic boundaries of the VLPO “core” or “cluster” (cVLPOGal) whereas the second subgroup is located in a dorsal and medial position to the core in the so-called “extended” VLPO (eVLPOGal) [14,27,46]. From a functional standpoint, cVLPOGal neurons are thought to promote NREM sleep, whereas eVLPOGal neurons are more strongly linked to the regulation of REM sleep [14,46]. With respect to the latter, the idea of the role of eVLPOGal neurons in REM sleep regulation increasingly gained support. For example, single-unit recording studies demonstrated that the eVLPO region contains REM-active neurons [13,47,48,49], and tracing studies have shown that the eVLPOGal neurons innervate key pontine nuclei implicated in REM sleep regulation [46,50,51]. However, the lack of a selective marker for distinguishing between cVLPOGal and eVLPOGal neurons and the fact the cVLPO and the eVLPO are very small nuclei that are located in close proximity to one another, have proven a challenge for researchers seeking to “parse” the respective roles of these anatomically distinct nuclei in behavioral state control.
4.3. The Glutamatergic VLPO
In addition to GABA-releasing neurons, the VLPO also contains glutamate-releasing neurons, as indicated by the expression of the vesicular glutamate transporter 2 (Vglut2) (Allen Brain Atlas, n.d.). Interestingly, the activation of glutamatergic VLPO (VLPOVglut2) neurons suppresses sleep, delays REM sleep and consolidates wakefulness for about an hour [52]. The optogenetic stimulation of glutamatergic neurons in the preoptic area, including the VLPO, also produces rapid transitions from NREM to wakefulness [30]. Taken together, these findings suggest that VLPOVglut2 neurons, upon their activation, function to promote waking rather than sleep. The circuit bases by which glutamatergic VLPO neurons contribute to arousal remains unresolved, although there are data to suggest that VLPO glutamatergic neurons might promote wakefulness by activating other arousal centers, such as the TMN [30]. Regardless, the arousal response elicited by the acute activation of VLPO glutamatergic neurons is relatively short in duration when compared to the responses evoked by the comparable activation of other wake-promoting neurons in the brain, which in some cases, consistently produce sustained wakefulness for three to six hours, even when this activation occurs during a period of high sleep drive [53,54,55,56].
5. VLPO Regulation of Behavioral State: Circuit Basis of Sleep and Arousal
How the brain transitions between the behavioral states of wake and sleep (and REM sleep) remains incompletely understood. One highly influential circuit model for state transitions, termed the “flip-flop” switch model, posits reciprocal inhibitory interactions between sleep- and wake-promoting cell groups, similar to the function of an electronic “flip-flop” switch/circuit [25,57,58]. Within the framework of the flip-flop model, the VLPO represents the “sleep side”, whereas the ascending arousal nodes represent the “arousal side”. That these cell groups exhibit, respectively, sleep- and wake-active firing profiles has provided general support for the model. More specifically, in this flip-flop design, the VLPO inhibits the arousal nodes (e.g., TMN, LC and DR neurons) through the synaptic release of GABA and galanin, whereas the TMN, LC and DR inhibit the VLPO through the synaptic release of histamine, noradrenaline and serotonin, respectively [59,60]. By virtue of the self-reinforcing nature of these switches—that is, when each side is firing they reduce their own inhibitory feedback—the flip-flop switch is inherently stable in either end state, but avoids intermediate states. Hence, the flip-flop design ensures the stability of behavioral states, but also facilitates relatively rapid switching between behavioral states. One design flaw of flip-flop switches is that the frequency of unwanted state transitions can increase if one side of the switch is “weakened”, as the weakened side becomes less able to inhibit the other side, thereby biasing the switch toward a midpoint where smaller perturbations may trigger a state transition. As an example, cell loss in the VLPO, which occurs during aging, may weaken the “sleep side” of the switch, which would predictably lead to sleep fragmentation and daytime napping, both of which are frequent complaints in the elderly [44]. While many features of the theoretical framework of the flip-flop model were discovered experimentally, other features have proven more difficult to convincingly demonstrate. For example, the LC and DRN are directly inhibited by the VLPO through the release of GABA, and the LC and DRN directly inhibit VLPO sleep-active neurons through the release of noradrenaline and serotonin, whereas reciprocal inhibition between the VLPO and the TMN necessarily requires additional synaptic relays; this is discussed in more detail below. Specifically, while the VLPO directly inhibits TMN neurons through the release of GABA, histamine released from the TMN does not inhibit the VLPO directly, but rather activates GABAergic interneurons that, in turn, inhibit the VLPO, i.e., feedforward inhibition [36,61]. Hence, while reciprocal inhibition per the flip-flop model is conserved, it does require further circuit elements, which in this example takes the form of GABAergic interneurons. Additionally, and as previously indicated, behavioral state transitions are not always as rapid as would be predicted by the flip-flop model, instead they often occur gradually, i.e., over minutes rather than seconds.
Rather strikingly, given the influential nature of this model and with the exception of a recent paper showing that the noradrenergic LC promotes arousal by inhibiting the activity of the VLPO [37], there exists little in vivo evidence that monoaminergic or cholinergic inhibition of the VLPO promotes EEG or behavioral arousal. Conversely, the flip-flop model does not fully consider the possibility that non-aminergic and non-cholinergic cell groups that innervate the VLPO may also contribute to sleep–wake regulation. Inspired by this knowledge gap, our labs recently explored this question and found that wake-promoting (and wake-active) GABAergic neurons of the lateral hypothalamus (LHGABA) [54], when acutely activated, can rapidly produce arousal from deep sleep by inhibiting sleep-promoting galanin neurons of the VLPO [62]. These results were the first to identify a long-range, non-aminergic and non-cholinergic inhibitory input that can effectively “turn off” the sleep-promoting neurons of the VLPO to promote arousal in vivo. We also found that using conditional retrograde tracing, which the LHGABA neurons themselves receive as a prominent input from the VLPO, supported the hypothesis that sleep-promoting VLPO and wake-promoting LHGABA neurons could exist in a state of mutual inhibition. Therefore, LHGABA neurons may comprise an additional, non-aminergic and non-cholinergic circuit component of the flip-flop switch.
As indicated, our understanding of how VLPOGal neurons are regulated, both by long-range monoaminergic and cholinergic inputs as well as by local (inter)neurons, is relatively immature. We know for example that VLPOGal neurons are inhibited by monoamines [33,34,59,63] and that monoaminergic inputs to the VLPO originate from the LC and the C1/A1 adrenergic cell group of the ventrolateral medulla (VLM) [29]. Accordingly, electrical and optogenetic stimulation of the LC or of the VLM inhibits the firing of sleep-active neurons in VLPO via α2-mediated effects [37,64], and promotes cortical activation, presumably as a direct function of inhibiting sleep-active VLPO neurons. VLPOGal neurons are similarly strongly inhibited by cholinergic agonists [59,65], although the source of cholinergic input(s) to the VLPO remains unclear, in particular as retrograde tracing studies determined that cholinergic inputs to the VLPO are rare, if nonexistent [29]. The apparent mismatch between cholinergic innervation and cholinergic receptors in the VLPO area remains a mystery.
The VLPO also receives inputs from other canonical arousal-related nodes, including lateral hypothalamic orexin neurons and, as indicated, the histaminergic TMN [29]. Targeted infusion of orexin or histamine into the VLPO produces rapid arousal responses [36,66,67]. Interestingly, during wakefulness, orexin and histamine inhibit VLPOGal neurons, although they do not do so directly. Their arousal responses appear to be produced through an indirect feedforward GABAergic mechanism [36,61,68]. The source of increased GABAergic tone providing inhibitory control over VLPOGal neurons during arousal, which can potentially promote arousal, remains unresolved. Orexin and histamine could, for instance, activate terminals of GABAergic afferences to the VLPO, or they could activate collaterals from local (i.e., intra-VLPO) GABAergic neurons. The available data support the latter model [36,68].
The results from recent studies have further suggested that, similar to orexin and histamine, other arousal signals may also inhibit VLPOGal neurons through an internal GABAergic circuit. For instance, noradrenaline and carbachol inhibit VLPOGal neurons directly [33,34,59] and indirectly through increasing GABA release [37]. Noradrenaline and carbachol directly inhibit the VLPOGal neurons via α2 adrenergic and muscarinic receptors [37,48,65,69], respectively, and noradrenaline can further increase GABAergic-afferent input via α1 adrenergic receptors, indicating an additional and synergistic mechanism of inhibition [37].
In summary, the VLPO is regulated by afferent inputs from many sources [29]. While some of these inputs provide a direct synaptic regulation of the VLPOGal sleep-promoting population, some afferences also appear to operate through a local (i.e., intra-VLPO) GABAergic circuit. The available data support the hypothesis that local GABAergic neurons within the VLPO (VLPOGABA), which provide collaterals to VLPOGal neurons, function as an interface between afferent inputs to the VLPO and VLPOGal neurons, effectively serving as a common ‘point of entry’ through which wake and sleep signals control VLPOGal neurons to regulate arousal, e.g., wake-promoting signals (via feedforward inhibition) inhibit VLPOGal neurons to produce arousal- and sleep-promoting signals (via disinhibition), activating VLPOGal neurons to produce NREM sleep. This local GABAergic mechanism for promoting state transitions could be shared by most, if not all, wake- and sleep-promoting inputs that modulate VLPO activity. According to this model, VLPOGABA neurons are wake-active and possibly wake-promoting. Recordings in the VLPO have uncovered neurons that are active in wakefulness [13]. Additionally, unlike the activation of VLPOGal neurons, the optogenetic activation of GAD2-expressing VLPO neurons (which likely activates both VLPOGal and VLPOGABA neurons) produces arousal, suggesting that the VLPO contains wake-promoting GABAergic neurons that can override the co-activation of sleep-promoting VLPOGal neurons [30].
Sleep Promoting Signals in VLPO—The Role of Adenosine and Prostaglandin D2
Adenosine (AD) and prostaglandin D2 (PGD2) are arguably the most widely recognized and intensively investigated endogenous sleep-promoting signals [70,71,72] (See Table 1 for key references). They also appear to be inextricably linked to the mechanism by which they promote sleep. Both AD and PGD2 progressively accumulate in the extracellular space during wakefulness and their levels continue to rise during sleep deprivation, [73,74] eventually driving sleep by inhibiting wake-promoting neurons throughout the brain and/or disinhibiting VLPO neurons. In addition, PGD2 levels are elevated during inflammation and are thought to mediate, at least in part, the sleepiness associated with infections and inflammation. For example, PGD2 levels are elevated in the CSF of patients with African sleeping sickness, as well as in the serum of narcoleptic patients, the levels in the latter correlating with excessive daytime sleepiness but not cataplexy [75,76].
AD itself is a purine metabolite that has long been linked with the sleep process [77]. For example, the subarachnoid administration of AD agonists promotes sleep and induces the expression of the cFos protein in VLPO neurons [78]. AD appears to exert these effects, at least in part, via adenosine A1 receptors to disinhibit VLPOGal neurons, i.e., by reducing GABAergic synaptic inputs on sleep-promoting VLPO neurons [79,80]. Pharmacological studies also suggested that AD may directly activate A2a receptors [33], although to the authors’ knowledge, it is not yet been determined if the A2a receptor is expressed in VLPO neurons, VLPOGal or otherwise.
Prostaglandins (PG), on the other hand, are a group of lipid compounds also known as eicosanoids. The specific role of PGD2—which is the most abundant PG in the brains of rats [81] and other mammals, including humans [82]—in sleep regulation has been identified. Initial and seminal research on PGD2 in sleep regulation was performed by Uneuo and colleagues [83,84,85,86], who reported that infusion of PGD2 into the lateral ventricles or into the subarachnoid space at the base of the forebrain, but not other regions of brain parenchyma, could induce sleep. Similar hypnogenic effects of ICV infusion of PGD2 were subsequently demonstrated in primates [87]. Importantly, animals under PGD2-induced sleep remain arousable, and the EEG power spectrum of NREM sleep is the same as that of natural sleep [71]. The concentration of PGD2 in the CSF also shows a strong diurnal rhythm that oscillates in parallel with the sleep–wake cycle [88], and notably, the amplitude of this oscillation increases during sleep deprivation, i.e., with an increase in sleep propensity [74]. Within the brain, PGD2 is produced in the leptomeninges, choroid plexus, and oligodendrocytes [89]. The main cognate receptor for PGD2 is the DP1 receptor, which is primarily expressed in the leptomeninges of the ventral surface of the rostral basal forebrain of mice. These DP1-enriched leptomeninges were immunohistochemically defined as bilateral wings under the rostral basal forebrain lateral to the optic chiasm, and thus were in close proximity to the anterior preoptic hypothalamus region containing the sleep-promoting VLPO neurons [86]. In fact, administration of PGD2 into the subarachnoid space at the base of the rat forebrain produces sustained NREM sleep (with minimal effects on REM sleep), and this sleep-inducing effect is absent in DP1-KO mice [86], indicating that NREM sleep induction by PGD2 is mediated by the DP1 receptors. However, in DP1-KO mice, it was further observed that their baseline sleep–wake patterns were essentially identical to WT mice, suggesting that DP1 receptors may not be crucial for basal sleep–wake regulation, or that the lack of receptors in these mice was compensated, developmentally or otherwise [86]. Yet, DP1-KO mice also exhibited a strongly attenuated rebound of NREM sleep following sleep deprivation, suggesting that endogenous PGD2 might be involved in the homeostatic regulation of NREM sleep [71,90]. Additionally, while the cellular bases of these effects are incompletely understood, it was shown that the administration of PGD2 into the subarachnoid space markedly increases the expression of cFos in VLPO neurons and the basal forebrain leptomeninges [91]. Given that the DP1 receptors are not expressed in the brain parenchyma, but rather only in the leptomeninges [86,92], these results suggest that PGD2 may act via the leptomeninges to activate VLPO neurons, and thereby promote sleep. Of additional interest, PGD2 infusion into the subarachnoid space of mice increases extracellular adenosine concentrations in a dose-dependent manner [86], and this effect is absent in DP1-KO mice [86].
Therefore, in summary, one current model that remains to be critically tested is the idea that PGD2 acts via DP1 receptors in the leptomeninges to induce the production of AD. This then activates sleep-promoting VLPOGal neurons, and AD most likely carries out this process by acting in the A1 receptors (i.e., indirectly through disinhibition) but could possibly act via the direct activation of A2a receptors [78,85].
6. VLPO and Other Physiological Processes
6.1. VLPO and Thermoregulation
Recent work has suggested that the VLPO, or more likely a subset of VLPO neurons, may contribute to thermoregulatory processes (See Table 1 for key references). For example, in addition to triggering sleep, the selective chemoactivation of VLPOGal neurons produces a marked hypothermia [28]. This finding is perhaps unsurprising given that a rapid decrease in core temperature is known to occur upon the onset of natural sleep [93,94,95], and that the preoptic region, likely including the VLPO, contains heat-sensitive neurons (i.e., neurons that increase their firing rates in response to local temperature increases) [49,96,97]. It has also been hypothesized that the activation of heat-sensitive preoptic neurons by increased core temperature may also subserve the connection between fever and sleepiness [98,99]. Regardless, the reduction in core temperature that occurs upon sleep onset is largely due to skin vasodilation, i.e., heat loss [100], and this process is linked with the inhibition of thermoregulatory neurons in the dorsomedial hypothalamic nucleus (DMH) and the raphe pallidus (RPa) [101,102]. However, it is the authors’ surmise that the promotion of NREM sleep and hypothermia are controlled by two distinct, but partially intermixed, VLPOGal cell populations, with those generating NREM sleep mainly located in the cVLPO, and those inducing heat loss mainly located in the dorsal and medial eVLPO [28,103]. In general support of this concept, we note from the authors’ heatmaps in the chemogenetic study of VLPOGal neurons ([28], Figure 4) that the simultaneous or otherwise joint activation of cVLPOGal and eVLPOGal neurons was near-certain, and thus would trigger both NREM sleep and hypothermia. Furthermore, and in the same study, optogenetic activation, which permitted a more selective targeting of cVLPOGal neurons (i.e., with fiber optics placed just above the cVLPO) promoted NREM sleep, but not hypothermia [28], whereas the optogenetic stimulation of GABA neurons located more dorsally, in an area roughly corresponding to the eVLPO, produces hypothermia [103]. Finally, tracing and physiological studies have shown that the region dorsal and medial to the cVLPO provides an inhibitory innervation of the DMH and the RPa and that the activation of this circuit can trigger heat loss through arterial vasodilation in the tail [104,105,106]. Of interest, more recent data suggested that neurons within the cVLPO may also contribute to this pathway, but that the functional cell group involved is most likely the (putative wake-promoting) VLPOVglut2 and not the sleep-promoting cVLPOGal neurons [107].
6.2. VLPO as Metabolic Sensor
An association between sleep and satiety is long-established, albeit largely anecdotally. Known as post prandial somnolence, aka “food comas”, this phenomenon has historically been linked with changes in post meal insulin (or other metabolic factors) or the activation of ascending gastrointestinal (vagal) inputs to the brainstem, although neither of these theories have been rigorously tested. However, recent studies suggested that sleep-active VLPO neurons might be capable of directly sensing metabolic changes, and thus might contribute to the sleep or sleepiness that accompanies a large meal. The infusion of glucose into the VLPO also promotes NREM sleep and increases cFos expression [108]. Interestingly, glucose-driven cFos activation in the VLPO appears to be a direct response to glucose. Glucose within the physiological range selectively excites putative VLPO sleep-promoting neurons [108] via the same cellular mechanism that occurs in glucose-sensing pancreatic beta cells and POMC neurons in the arcuate nucleus [109,110]. Specifically, glucose is initially transported intracellularly and then catabolized via glycolysis, which leads to an increase in ATP levels and induces the closure of KATP channels and membrane depolarization [111]. Satiety signals may therefore promote sleep by activating sleep-active VLPO neurons, which is an interesting parallel phenomenon to that of the hunger- and hypoglycemia-driven activation of centers, which drive arousal to facilitate foraging [112]. Therefore, while wakefulness/alertness is enhanced with fasting by the activation of wake-promoting neurons, resting (i.e., sleeping/napping) is favored after food intake and direct activation of VLPO sleep-promoting neurons may contribute to this change in behavioral state [113,114,115,116].
6.3. VLPO and Anesthesia
Over the past two decades, there has been an increased interest in the relationship between sleep and general anesthesia [117,118]. One emerging hypothesis is that loss of consciousness in anesthesia might rely on the same circuitry that produces natural sleep, in particular for anesthetics that may interact with and activate sleep-promoting neurons in the preoptic area, including the VLPO. This hypothesis derives support from the finding that putative sleep-promoting neurons in the VLPO are directly activated by volatile anesthetics, such as isoflurane and halothane, and that general anesthesia induces cFos expression in VLPO neurons [63,69,119,120,121]. However, both VLPO lesion studies and the selective activation of GABAergic or glutamatergic neurons in VLPO have found an absence of effect of these manipulations on anesthetic induction or recovery time [16,52]. Interestingly, sleep deprivation increases sensitivity to anesthesia and prolongs time to emergence [122]. Therefore, sleep loss could explain why, paradoxically, the extent of cell loss following lesions of the VLPO correlates with deeper, rather than lighter, anesthesia and delayed, rather than quicker, emergence [16]. Hence, while sleep-promoting neurons in the VLPO are activated during anesthesia, their activation is unlikely to be necessary for the loss of consciousness induced by general anesthesia.
7. On the Horizon for VLPO Research
While a subjective assessment, the authors of this review have identified several current and important knowledge gaps regarding the neurobiology of the VLPO. The first of these gaps is the incompletely understood role that putative wake-promoting glutamatergic VLPO neurons play in the regulation of behavioral state (Figure 1). More specifically, we think it will be important to identify and characterize their afferent inputs and efferent outputs and to determine whether, in addition to glutamate, they express and co-release any of the neuropeptides found in the VLPO region. Another important knowledge gap is how transcriptional profiles may vary across VLPO cell subtypes. For example, do VLPOGABA neurons, which appear to function as a critical interface between VLPOGal neurons and modulatory afferent inputs, express unique GPCRs? If so, these could form the basis for newer and more targeted pharmacologic approaches to treating the crippling inability to maintain consolidated wakefulness in a host of neurodegenerative, neurodevelopmental, and neuropsychiatric diseases, in which disordered arousal, including even hyperarousal (e.g., insomnia), is a prominent clinical feature. The same question can be asked about the cVLPOGal and eVLPOGal cell populations, which themselves may be transcriptionally heterogeneous, and thus further subdivided into functionally distinct cell clusters.
8. Summary
Without doubt, challenges lie ahead as we seek a more refined understanding of the neurobiology of the VLPO, but the ever-expanding toolkit of newer methods and technologies will unquestionably facilitate deeper and more detailed interrogations of the cellular and molecular VLPO. It is our expectation that, within the next decade or two, experimental work will reveal druggable targets in key VLPO subpopulations that will in turn permit the development of newer drugs that can be clinically deployed for treating a wide range of sleep- and arousal-based disorders without side effects. Our confidence in this outcome is based on the fact that in the span of just 25 years, the field has gone from being unaware that the VLPO was a bona fide schlafsteuerungszentrum, to our current and impressively detailed understanding of the VLPO’s heterogeneous cellular composition, how these cells are modulated by pre-synaptic inputs, and a far more granular understanding of their intra-nucleus circuitry.

{kind=link}
Table 1.
Selected key references.
| The discovery of VLPO as sleep-promoting nucleus | |
| Afferent and efferent inputs and cellular composition | |
| Electrophysiology and pharmacological studies | In vitro electrophysiology studies |
| Regulation of behavioral states | |
| Other physiological processes | Thermoregulation:
|
Table 2.
Cellular anatomy of the VLPO: selected key references.
| The VLPO nucleus | |
| Galaninergic neurons |
|
| GABAergic neurons | |
| Glutamatergic neurons |
Funding
This work was supported by National Institutes of Health Grants R01NS091126 to E.A. and R01NS107315 to PMF.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Von Economo, C. Die Encephalitis Lethargica; Franz Deuticke: Vienna, Austria, 1918. [Google Scholar]
- Von Economo, C. Sleep as a Problem of Localization. J. Nerv. Ment. Dis. 1930, 71, 249–259. [Google Scholar] [CrossRef]
- Hess, W.R. Le sommeil. C. R. Soc. Biol. Paris 1931, 107, 1333–1360. [Google Scholar]
- Ranson, S.W.; Ingram, W.R. Catalepsy Caused by Lesions between the Mamillary Bodies and Third Nerve in the Cat. Am. J. Physiol. 1932, 101, 690–696. [Google Scholar] [CrossRef] [Green Version]
- Ingram, W.R.; Barris, R.W.; Ranson, S.W. Catalepsy: An Experimental Study. Arch. Neurol. Psychiatry 1936, 35, 1175–1197. [Google Scholar] [CrossRef]
- Ranson, S.W. Somnolence Caused by Hypothalamic Lesions in Monkeys. Arch. Neurol. Psychiatr. 1939, 41, 1–23. [Google Scholar] [CrossRef]
- Nauta, W.J. Hypothalamic Regulation of Sleep in Rats; an Experimental Study. J. Neurophysiol. 1946, 9, 285–316. [Google Scholar] [CrossRef]
- McGinty, D.J.; Sterman, M.B. Sleep Suppression after Basal Forebrain Lesions in the Cat. Science 1968, 160, 1253–1255. [Google Scholar] [CrossRef]
- Sherin, J.E.; Shiromani, P.J.; McCarley, R.W.; Saper, C.B. Activation of Ventrolateral Preoptic Neurons during Sleep. Science 1996, 271, 216–219. [Google Scholar] [CrossRef] [Green Version]
- Szymusiak, R.; McGinty, D. Sleep-Waking Discharge of Basal Forebrain Projection Neurons in Cats. Brain Res. Bull. 1989, 22, 423–430. [Google Scholar] [CrossRef]
- Takahashi, K.; Lin, J.S.; Sakai, K. Characterization and Mapping of Sleep-Waking Specific Neurons in the Basal Forebrain and Preoptic Hypothalamus in Mice. Neuroscience 2009, 161, 269–292. [Google Scholar] [CrossRef] [Green Version]
- Gvilia, I.; Xu, F.; McGinty, D.; Szymusiak, R. Homeostatic Regulation of Sleep: A Role for Preoptic Area Neurons. J. Neurosci. 2006, 26, 9426–9433. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.A.; Kumar, S.; McGinty, D.; Alam, M.N.; Szymusiak, R. Neuronal Activity in the Preoptic Hypothalamus during Sleep Deprivation and Recovery Sleep. J. Neurophysiol. 2014, 111, 287–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Greco, M.A.; Shiromani, P.; Saper, C.B. Effect of Lesions of the Ventrolateral Preoptic Nucleus on NREM and REM Sleep. J. Neurosci. 2000, 20, 3830–3842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arrigoni, E.; Lu, J.; Vetrivelan, R.; Saper, C.B. Long-Term Synaptic Plasticity Is Impaired in Rats with Lesions of the Ventrolateral Preoptic Nucleus. Eur. J. Neurosci. 2009, 30, 2112–2120. [Google Scholar] [CrossRef] [Green Version]
- Eikermann, M.; Vetrivelan, R.; Grosse-Sundrup, M.; Henry, M.E.; Hoffmann, U.; Yokota, S.; Saper, C.B.; Chamberlin, N.L. The Ventrolateral Preoptic Nucleus Is Not Required for Isoflurane General Anesthesia. Brain Res. 2011, 1426, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Vetrivelan, R.; Fuller, P.M.; Yokota, S.; Lu, J.; Saper, C.B. Metabolic Effects of Chronic Sleep Restriction in Rats. Sleep 2012, 35, 1511–1520. [Google Scholar] [CrossRef]
- Vetrivelan, R.; Kong, D.; Ferrari, L.L.; Arrigoni, E.; Madara, J.C.; Bandaru, S.S.; Lowell, B.B.; Lu, J.; Saper, C.B. Melanin-Concentrating Hormone Neurons Specifically Promote Rapid Eye Movement Sleep in Mice. Neuroscience 2016, 336, 102–113. [Google Scholar] [CrossRef] [Green Version]
- Anaclet, C.; Lin, J.S.; Vetrivelan, R.; Krenzer, M.; Vong, L.; Fuller, P.M.; Lu, J. Identification and Characterization of a Sleep-Active Cell Group in the Rostral Medullary Brainstem. J. Neurosci. 2012, 32, 17970–17976. [Google Scholar] [CrossRef]
- Anaclet, C.; Ferrari, L.; Arrigoni, E.; Bass, C.E.; Saper, C.B.; Lu, J.; Fuller, P.M. The GABAergic Parafacial Zone Is a Medullary Slow Wave Sleep-Promoting Center. Nat. Neurosci. 2014, 17, 1217–1224. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.A.; Kostin, A.; Siegel, J.; McGinty, D.; Szymusiak, R.; Alam, M.N. Characteristics of Sleep-Active Neurons in the Medullary Parafacial Zone in Rats. Sleep 2018, 41, zsy130. [Google Scholar] [CrossRef]
- Gong, H.; McGinty, D.; Guzman-Marin, R.; Chew, K.T.; Stewart, D.; Szymusiak, R. Activation of C-Fos in GABAergic Neurones in the Preoptic Area during Sleep and in Response to Sleep Deprivation. J. Physiol. 2004, 556, 935–946. [Google Scholar] [CrossRef] [PubMed]
- McGinty, D.; Szymusiak, R. Hypothalamic Regulation of Sleep and Arousal. Front. Biosci. 2003, 8, s1074–s1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anaclet, C.; Fuller, P.M. Brainstem Regulation of Slow-Wave-Sleep. Curr. Opin. Neurobiol. 2017, 44, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Saper, C.B.; Fuller, P.M. Wake-Sleep Circuitry: An Overview. Curr. Opin. Neurobiol. 2017, 44, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Sherin, J.E.; Elmquist, J.K.; Torrealba, F.; Saper, C.B. Innervation of Histaminergic Tuberomammillary Neurons by GABAergic and Galaninergic Neurons in the Ventrolateral Preoptic Nucleus of the Rat. J. Neurosci. 1998, 18, 4705–4721. [Google Scholar] [CrossRef] [Green Version]
- Gaus, S.E.; Strecker, R.E.; Tate, B.A.; Parker, R.A.; Saper, C.B. Ventrolateral Preoptic Nucleus Contains Sleep-Active, Galaninergic Neurons in Multiple Mammalian Species. Neuroscience 2002, 115, 285–294. [Google Scholar] [CrossRef]
- Kroeger, D.; Absi, G.; Gagliardi, C.; Bandaru, S.S.; Madara, J.C.; Ferrari, L.L.; Arrigoni, E.; Munzberg, H.; Scammell, T.E.; Saper, C.B.; et al. Galanin Neurons in the Ventrolateral Preoptic Area Promote Sleep and Heat Loss in Mice. Nat. Commun. 2018, 9, 4129. [Google Scholar] [CrossRef] [Green Version]
- Chou, T.C.; Bjorkum, A.A.; Gaus, S.E.; Lu, J.; Scammell, T.E.; Saper, C.B. Afferents to the Ventrolateral Preoptic Nucleus. J. Neurosci. 2002, 22, 977–990. [Google Scholar] [CrossRef] [Green Version]
- Chung, S.; Weber, F.; Zhong, P.; Tan, C.L.; Nguyen, T.N.; Beier, K.T.; Hormann, N.; Chang, W.C.; Zhang, Z.; Do, J.P.; et al. Identification of Preoptic Sleep Neurons Using Retrograde Labelling and Gene Profiling. Nature 2017, 545, 477–481. [Google Scholar] [CrossRef] [Green Version]
- Moffitt, J.R.; Bambah-Mukku, D.; Eichhorn, S.W.; Vaughn, E.; Shekhar, K.; Perez, J.D.; Rubinstein, N.D.; Hao, J.; Regev, A.; Dulac, C.; et al. Molecular, Spatial, and Functional Single-Cell Profiling of the Hypothalamic Preoptic Region. Science 2018, 362, eaau5324. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, S.; Jang, I.S.; Nabekura, J.; Akaike, N. Alpha 2-Adrenoceptor-Mediated Presynaptic Modulation of GABAergic Transmission in Mechanically Dissociated Rat Ventrolateral Preoptic Neurons. J. Neurophysiol. 2003, 89, 1640–1648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallopin, T.; Luppi, P.H.; Cauli, B.; Urade, Y.; Rossier, J.; Hayaishi, O.; Lambolez, B.; Fort, P. The Endogenous Somnogen Adenosine Excites a Subset of Sleep-Promoting Neurons via A2A Receptors in the Ventrolateral Preoptic Nucleus. Neuroscience 2005, 134, 1377–1390. [Google Scholar] [CrossRef] [PubMed]
- Sangare, A.; Dubourget, R.; Geoffroy, H.; Gallopin, T.; Rancillac, A. Serotonin Differentially Modulates Excitatory and Inhibitory Synaptic Inputs to Putative Sleep-Promoting Neurons of the Ventrolateral Preoptic Nucleus. Neuropharmacology 2016, 109, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Chauveau, F.; Claverie, D.; Lardant, E.; Varin, C.; Hardy, E.; Walter, A.; Canini, F.; Rouach, N.; Rancillac, A. Neuropeptide S Promotes Wakefulness through the Inhibition of Sleep-Promoting Ventrolateral Preoptic Nucleus Neurons. Sleep 2020, 43, zsz189. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.W.; Li, J.; Ye, J.H. Histamine Regulates Activities of Neurons in the Ventrolateral Preoptic Nucleus. J. Physiol. 2010, 588, 4103–4116. [Google Scholar] [CrossRef]
- Liang, Y.; Shi, W.; Xiang, A.; Hu, D.; Wang, L.; Zhang, L. The NAergic Locus Coeruleus-Ventrolateral Preoptic Area Neural Circuit Mediates Rapid Arousal from Sleep. Curr. Biol. 2021, 31, 3729–3742.e5. [Google Scholar] [CrossRef]
- Kask, K.; Langel, U.; Bartfai, T. Galanin—A Neuropeptide with Inhibitory Actions. Cell Mol. Neurobiol. 1995, 15, 653–673. [Google Scholar] [CrossRef]
- Pieribone, V.A.; Xu, Z.Q.; Zhang, X.; Grillner, S.; Bartfai, T.; Hokfelt, T. Galanin Induces a Hyperpolarization of Norepinephrine-Containing Locus Coeruleus Neurons in the Brainstem Slice. Neuroscience 1995, 64, 861–874. [Google Scholar] [CrossRef]
- Schonrock, B.; Busselberg, D.; Haas, H.L. Properties of Tuberomammillary Histamine Neurones and Their Response to Galanin. Agents Actions 1991, 33, 135–137. [Google Scholar] [CrossRef]
- Xu, Z.Q.; Zhang, X.; Pieribone, V.A.; Grillner, S.; Hökfelt, T. Galanin-5-Hydroxytryptamine Interactions: Electrophysiological, Immunohistochemical and in Situ Hybridization Studies on Rat Dorsal Raphe Neurons with a Note on Galanin R1 and R2 Receptors. Neuroscience 1998, 87, 79–94. [Google Scholar] [CrossRef]
- Goforth, P.B.; Leinninger, G.M.; Patterson, C.M.; Satin, L.S.; Myers, M.G. Leptin Acts via Lateral Hypothalamic Area Neurotensin Neurons to Inhibit Orexin Neurons by Multiple GABA-Independent Mechanisms. J. Neurosci. 2014, 34, 11405–11415. [Google Scholar] [CrossRef] [PubMed]
- Luppi, P.H.; Aston-Jones, G.; Akaoka, H.; Chouvet, G.; Jouvet, M. Afferent Projections to the Rat Locus Coeruleus Demonstrated by Retrograde and Anterograde Tracing with Cholera-Toxin B Subunit and Phaseolus Vulgaris Leucoagglutinin. Neuroscience 1995, 65, 119–160. [Google Scholar] [CrossRef]
- Lim, A.S.; Ellison, B.A.; Wang, J.L.; Yu, L.; Schneider, J.A.; Buchman, A.S.; Bennett, D.A.; Saper, C.B. Sleep Is Related to Neuron Numbers in the Ventrolateral Preoptic/Intermediate Nucleus in Older Adults with and without Alzheimer’s Disease. Brain 2014, 137, 2847–2861. [Google Scholar] [CrossRef] [PubMed]
- Saper, C.B. The Intermediate Nucleus in Humans: Cytoarchitecture, Chemoarchitecture, and Relation to Sleep, Sex, and Alzheimer Disease. Handb. Clin. Neurol. 2021, 179, 461–469. [Google Scholar] [CrossRef]
- Lu, J.; Bjorkum, A.A.; Xu, M.; Gaus, S.E.; Shiromani, P.J.; Saper, C.B. Selective Activation of the Extended Ventrolateral Preoptic Nucleus during Rapid Eye Movement Sleep. J. Neurosci. 2002, 22, 4568–4576. [Google Scholar] [CrossRef]
- Koyama, Y.; Hayaishi, O. Firing of Neurons in the Preoptic/Anterior Hypothalamic Areas in Rat: Its Possible Involvement in Slow Wave Sleep and Paradoxical Sleep. Neurosci. Res. 1994, 19, 31–38. [Google Scholar] [CrossRef]
- Osaka, T.; Matsumura, H. Noradrenaline Inhibits Preoptic Sleep-Active Neurons through Alpha 2-Receptors in the Rat. Neurosci. Res. 1995, 21, 323–330. [Google Scholar] [CrossRef]
- Szymusiak, R.; Alam, N.; Steininger, T.L.; McGinty, D. Sleep-Waking Discharge Patterns of Ventrolateral Preoptic/Anterior Hypothalamic Neurons in Rats. Brain Res. 1998, 803, 178–188. [Google Scholar] [CrossRef]
- Lu, J.; Sherman, D.; Devor, M.; Saper, C.B. A Putative Flip-Flop Switch for Control of REM Sleep. Nature 2006, 441, 589–594. [Google Scholar] [CrossRef]
- Hsieh, K.-C.; Gvilia, I.; Kumar, S.; Uschakov, A.; McGinty, D.; Alam, M.N.; Szymusiak, R. C-Fos Expression in Neurons Projecting from the Preoptic and Lateral Hypothalamic Areas to the Ventrolateral Periaqueductal Gray in Relation to Sleep States. Neuroscience 2011, 188, 55–67. [Google Scholar] [CrossRef] [Green Version]
- Vanini, G.; Bassana, M.; Mast, M.; Mondino, A.; Cerda, I.; Phyle, M.; Chen, V.; Colmenero, A.V.; Hambrecht-Wiedbusch, V.S.; Mashour, G.A. Activation of Preoptic GABAergic or Glutamatergic Neurons Modulates Sleep-Wake Architecture, but Not Anesthetic State Transitions. Curr. Biol. 2020, 30, 779–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anaclet, C.; Pedersen, N.P.; Ferrari, L.L.; Venner, A.; Bass, C.E.; Arrigoni, E.; Fuller, P.M. Basal Forebrain Control of Wakefulness and Cortical Rhythms. Nat. Commun. 2015, 6, 8744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venner, A.; Anaclet, C.; Broadhurst, R.Y.; Saper, C.B.; Fuller, P.M. A Novel Population of Wake-Promoting GABAergic Neurons in the Ventral Lateral Hypothalamus. Curr. Biol. 2016, 26, 2137–2143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, N.P.; Ferrari, L.; Venner, A.; Wang, J.L.; Abbott, S.B.G.; Vujovic, N.; Arrigoni, E.; Saper, C.B.; Fuller, P.M. Supramammillary Glutamate Neurons Are a Key Node of the Arousal System. Nat. Commun. 2017, 8, 1405. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Wang, D.-R.; Dong, H.; Chen, L.; Lu, J.; Lazarus, M.; Cherasse, Y.; Chen, G.-H.; Qu, W.-M.; Huang, Z.-L. Medial Parabrachial Nucleus Is Essential in Controlling Wakefulness in Rats. Front. Neurosci. 2021, 15, 645877. [Google Scholar] [CrossRef] [PubMed]
- Saper, C.B.; Fuller, P.M.; Pedersen, N.P.; Lu, J.; Scammell, T.E. Sleep State Switching. Neuron 2010, 68, 1023–1042. [Google Scholar] [CrossRef] [Green Version]
- Scammell, T.E.; Arrigoni, E.; Lipton, J.O. Neural Circuitry of Wakefulness and Sleep. Neuron 2017, 93, 747–765. [Google Scholar] [CrossRef] [Green Version]
- Gallopin, T.; Fort, P.; Eggermann, E.; Cauli, B.; Luppi, P.H.; Rossier, J.; Audinat, E.; Muhlethaler, M.; Serafin, M. Identification of Sleep-Promoting Neurons in Vitro. Nature 2000, 404, 992–995. [Google Scholar] [CrossRef]
- Gallopin, T.; Luppi, P.H.; Rambert, F.A.; Frydman, A.; Fort, P. Effect of the Wake-Promoting Agent Modafinil on Sleep-Promoting Neurons from the Ventrolateral Preoptic Nucleus: An in Vitro Pharmacologic Study. Sleep 2004, 27, 19–25. [Google Scholar]
- Williams, R.H.; Chee, M.J.; Kroeger, D.; Ferrari, L.L.; Maratos-Flier, E.; Scammell, T.E.; Arrigoni, E. Optogenetic-Mediated Release of Histamine Reveals Distal and Autoregulatory Mechanisms for Controlling Arousal. J. Neurosci. 2014, 34, 6023–6029. [Google Scholar] [CrossRef]
- Venner, A.; De Luca, R.; Sohn, L.T.; Bandaru, S.S.; Verstegen, A.M.J.; Arrigoni, E.; Fuller, P.M. An Inhibitory Lateral Hypothalamic-Preoptic Circuit Mediates Rapid Arousals from Sleep. Curr. Biol. 2019, 29, 4155–4168. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.T.; Chen, J.; Han, B.; Meng, Q.C.; Veasey, S.C.; Beck, S.G.; Kelz, M.B. Direct Activation of Sleep-Promoting VLPO Neurons by Volatile Anesthetics Contributes to Anesthetic Hypnosis. Curr. Biol. 2012, 22, 2008–2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osaka, T.; Matsumura, H. Noradrenergic Inputs to Sleep-Related Neurons in the Preoptic Area from the Locus Coeruleus and the Ventrolateral Medulla in the Rat. Neurosci. Res. 1994, 19, 39–50. [Google Scholar] [CrossRef]
- Saint-Mleux, B.; Eggermann, E.; Bisetti, A.; Bayer, L.; Machard, D.; Jones, B.E.; Muhlethaler, M.; Serafin, M. Nicotinic Enhancement of the Noradrenergic Inhibition of Sleep-Promoting Neurons in the Ventrolateral Preoptic Area. J. Neurosci. 2004, 24, 63–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Methippara, M.M.; Alam, M.N.; Szymusiak, R.; McGinty, D. Effects of Lateral Preoptic Area Application of Orexin-A on Sleep-Wakefulness. Neuroreport 2000, 11, 3423–3426. [Google Scholar] [CrossRef]
- Mavanji, V.; Perez-Leighton, C.E.; Kotz, C.M.; Billington, C.J.; Parthasarathy, S.; Sinton, C.M.; Teske, J.A. Promotion of Wakefulness and Energy Expenditure by Orexin-A in the Ventrolateral Preoptic Area. Sleep 2015, 38, 1361–1370. [Google Scholar] [CrossRef] [PubMed]
- De Luca, R.; Park, D.; Bandaru, S.S. Orexin Mediates Feed-Forward Inhibition of VLPO Sleep-Active Neurons—A Mechanism for Controlling Arousal. Sleep 2017, 40, A50. [Google Scholar] [CrossRef] [Green Version]
- McCarren, H.S.; Chalifoux, M.R.; Han, B.; Moore, J.T.; Meng, Q.C.; Baron-Hionis, N.; Sedigh-Sarvestani, M.; Contreras, D.; Beck, S.G.; Kelz, M.B. A2-Adrenergic Stimulation of the Ventrolateral Preoptic Nucleus Destabilizes the Anesthetic State. J. Neurosci. 2014, 34, 16385–16396. [Google Scholar] [CrossRef]
- Huang, Z.L.; Urade, Y.; Hayaishi, O. Prostaglandins and Adenosine in the Regulation of Sleep and Wakefulness. Curr. Opin. Pharmacol. 2007, 7, 33–38. [Google Scholar] [CrossRef]
- Urade, Y.; Hayaishi, O. Prostaglandin D2 and Sleep/Wake Regulation. Sleep Med. Rev. 2011, 15, 411–418. [Google Scholar] [CrossRef]
- Lazarus, M.; Chen, J.-F.; Huang, Z.-L.; Urade, Y.; Fredholm, B.B. Adenosine and Sleep. Handb. Exp. Pharmacol. 2019, 253, 359–381. [Google Scholar] [CrossRef] [PubMed]
- Porkka-Heiskanen, T.; Strecker, R.E.; Thakkar, M.; Bjorkum, A.A.; Greene, R.W.; McCarley, R.W. Adenosine: A Mediator of the Sleep-Inducing Effects of Prolonged Wakefulness. Science 1997, 276, 1265–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ram, A.; Pandey, H.P.; Matsumura, H.; Kasahara-Orita, K.; Nakajima, T.; Takahata, R.; Satoh, S.; Terao, A.; Hayaishi, O. CSF Levels of Prostaglandins, Especially the Level of Prostaglandin D2, Are Correlated with Increasing Propensity towards Sleep in Rats. Brain Res. 1997, 751, 81–89. [Google Scholar] [CrossRef]
- Pentreath, V.W.; Rees, K.; Owolabi, O.A.; Philip, K.A.; Doua, F. The Somnogenic T Lymphocyte Suppressor Prostaglandin D2 Is Selectively Elevated in Cerebrospinal Fluid of Advanced Sleeping Sickness Patients. Trans. R. Soc. Trop. Med. Hyg. 1990, 84, 795–799. [Google Scholar] [CrossRef]
- Jordan, W.; Tumani, H.; Cohrs, S.; Eggert, S.; Rodenbeck, A.; Brunner, E.; Rüther, E.; Hajak, G. Prostaglandin D Synthase (Beta-Trace) in Healthy Human Sleep. Sleep 2004, 27, 867–874. [Google Scholar] [CrossRef]
- Porkka-Heiskanen, T.; Kalinchuk, A.V. Adenosine, Energy Metabolism and Sleep Homeostasis. Sleep Med. Rev. 2011, 15, 123–135. [Google Scholar] [CrossRef]
- Scammell, T.E.; Gerashchenko, D.Y.; Mochizuki, T.; McCarthy, M.T.; Estabrooke, I.V.; Sears, C.A.; Saper, C.B.; Urade, Y.; Hayaishi, O. An Adenosine A2a Agonist Increases Sleep and Induces Fos in Ventrolateral Preoptic Neurons. Neuroscience 2001, 107, 653–663. [Google Scholar] [CrossRef]
- Chamberlin, N.L.; Arrigoni, E.; Chou, T.C.; Scammell, T.E.; Greene, R.W.; Saper, C.B. Effects of Adenosine on Gabaergic Synaptic Inputs to Identified Ventrolateral Preoptic Neurons. Neuroscience 2003, 119, 913–918. [Google Scholar] [CrossRef]
- Morairty, S.; Rainnie, D.; McCarley, R.; Greene, R. Disinhibition of Ventrolateral Preoptic Area Sleep-Active Neurons by Adenosine: A New Mechanism for Sleep Promotion. Neuroscience 2004, 123, 451–457. [Google Scholar] [CrossRef]
- Narumiya, S.; Ogorochi, T.; Nakao, K.; Hayaishi, O. Prostaglandin D2 in Rat Brain, Spinal Cord and Pituitary: Basal Level and Regional Distribution. Life Sci. 1982, 31, 2093–2103. [Google Scholar] [CrossRef]
- Ogorochi, T.; Narumiya, S.; Mizuno, N.; Yamashita, K.; Miyazaki, H.; Hayaishi, O. Regional Distribution of Prostaglandins D2, E2, and F2 Alpha and Related Enzymes in Postmortem Human Brain. J. Neurochem. 1984, 43, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Ueno, R.; Ishikawa, Y.; Nakayama, T.; Hayaishi, O. Prostaglandin D2 Induces Sleep When Microinjected into the Preoptic Area of Conscious Rats. Biochem. Biophys. Res. Commun. 1982, 109, 576–582. [Google Scholar] [CrossRef]
- Matsumura, H.; Nakajima, T.; Osaka, T.; Satoh, S.; Kawase, K.; Kubo, E.; Kantha, S.S.; Kasahara, K.; Hayaishi, O. Prostaglandin D2-Sensitive, Sleep-Promoting Zone Defined in the Ventral Surface of the Rostral Basal Forebrain. Proc. Natl. Acad. Sci. USA 1994, 91, 11998–12002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh, S.; Matsumura, H.; Suzuki, F.; Hayaishi, O. Promotion of Sleep Mediated by the A2a-Adenosine Receptor and Possible Involvement of This Receptor in the Sleep Induced by Prostaglandin D2 in Rats. Proc. Natl. Acad. Sci. USA 1996, 93, 5980–5984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizoguchi, A.; Eguchi, N.; Kimura, K.; Kiyohara, Y.; Qu, W.M.; Huang, Z.L.; Mochizuki, T.; Lazarus, M.; Kobayashi, T.; Kaneko, T.; et al. Dominant Localization of Prostaglandin D Receptors on Arachnoid Trabecular Cells in Mouse Basal Forebrain and Their Involvement in the Regulation of Non-Rapid Eye Movement Sleep. Proc. Natl. Acad. Sci. USA 2001, 98, 11674–11679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onoe, H.; Ueno, R.; Fujita, I.; Nishino, H.; Oomura, Y.; Hayaishi, O. Prostaglandin D2, a Cerebral Sleep-Inducing Substance in Monkeys. Proc. Natl. Acad. Sci. USA 1988, 85, 4082–4086. [Google Scholar] [CrossRef] [Green Version]
- Pandey, H.P.; Ram, A.; Matsumura, H.; Hayaishi, O. Concentration of Prostaglandin D2 in Cerebrospinal Fluid Exhibits a Circadian Alteration in Conscious Rats. Biochem. Mol. Biol. Int. 1995, 37, 431–437. [Google Scholar]
- Beuckmann, C.T.; Lazarus, M.; Gerashchenko, D.; Mizoguchi, A.; Nomura, S.; Mohri, I.; Uesugi, A.; Kaneko, T.; Mizuno, N.; Hayaishi, O.; et al. Cellular Localization of Lipocalin-Type Prostaglandin D Synthase (Beta-Trace) in the Central Nervous System of the Adult Rat. J. Comp. Neurol. 2000, 428, 62–78. [Google Scholar] [CrossRef]
- Hayaishi, O.; Urade, Y.; Eguchi, N.; Huang, Z.L. Genes for Prostaglandin d Synthase and Receptor as Well as Adenosine A2A Receptor Are Involved in the Homeostatic Regulation of Nrem Sleep. Arch. Ital. Biol. 2004, 142, 533–539. [Google Scholar]
- Scammell, T.; Gerashchenko, D.; Urade, Y.; Onoe, H.; Saper, C.; Hayaishi, O. Activation of Ventrolateral Preoptic Neurons by the Somnogen Prostaglandin D2. Proc. Natl. Acad. Sci. USA 1998, 95, 7754–7759. [Google Scholar] [CrossRef] [Green Version]
- Gerashchenko, D.; Beuckmann, C.T.; Kanaoka, Y.; Eguchi, N.; Gordon, W.C.; Urade, Y.; Bazan, N.G.; Hayaishi, O. Dominant Expression of Rat Prostanoid DP Receptor MRNA in Leptomeninges, Inner Segments of Photoreceptor Cells, Iris Epithelium, and Ciliary Processes. J. Neurochem. 1998, 71, 937–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alföldi, P.; Rubicsek, G.; Cserni, G.; Obál, F. Brain and Core Temperatures and Peripheral Vasomotion during Sleep and Wakefulness at Various Ambient Temperatures in the Rat. Pflugers Arch. 1990, 417, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Landolt, H.P.; Moser, S.; Wieser, H.G.; Borbély, A.A.; Dijk, D.J. Intracranial Temperature across 24-Hour Sleep-Wake Cycles in Humans. Neuroreport 1995, 6, 913–917. [Google Scholar] [CrossRef] [PubMed]
- Mochizuki, T.; Klerman, E.B.; Sakurai, T.; Scammell, T.E. Elevated Body Temperature during Sleep in Orexin Knockout Mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, R533–R540. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.N.; McGinty, D.; Szymusiak, R. Neuronal Discharge of Preoptic/Anterior Hypothalamic Thermosensitive Neurons: Relation to NREM Sleep. Am. J. Physiol. 1995, 269, R1240–R1249. [Google Scholar] [CrossRef]
- Gong, H.; Szymusiak, R.; King, J.; Steininger, T.; McGinty, D. Sleep-Related c-Fos Protein Expression in the Preoptic Hypothalamus: Effects of Ambient Warming. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, R2079–R2088. [Google Scholar] [CrossRef]
- Nakamura, K. Central Circuitries for Body Temperature Regulation and Fever. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R1207–R1228. [Google Scholar] [CrossRef] [Green Version]
- Saper, C.B.; Romanovsky, A.A.; Scammell, T.E. Neural Circuitry Engaged by Prostaglandins during the Sickness Syndrome. Nat. Neurosci. 2012, 15, 1088–1095. [Google Scholar] [CrossRef]
- Harding, E.C.; Franks, N.P.; Wisden, W. The Temperature Dependence of Sleep. Front. Neurosci. 2019, 13, 336. [Google Scholar] [CrossRef]
- Morrison, S.F.; Nakamura, K. Central Neural Pathways for Thermoregulation. Front. Biosci. 2011, 16, 74–104. [Google Scholar] [CrossRef] [Green Version]
- Morrison, S.F. Central Neural Control of Thermoregulation and Brown Adipose Tissue. Auton. Neurosci. 2016, 196, 14–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.-D.; Yang, W.Z.; Gao, C.; Fu, X.; Zhang, W.; Zhou, Q.; Chen, W.; Ni, X.; Lin, J.-K.; Yang, J.; et al. A Hypothalamic Circuit That Controls Body Temperature. Proc. Natl. Acad. Sci. USA 2017, 114, 2042–2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, M.; McKinley, M.J.; McAllen, R.M. Roles of Two Preoptic Cell Groups in Tonic and Febrile Control of Rat Tail Sympathetic Fibers. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R1248–R1257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, M.; McKinley, M.J.; McAllen, R.M. Preoptic-Raphé Connections for Thermoregulatory Vasomotor Control. J. Neurosci. 2011, 31, 5078–5088. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, K.; Li, X.; Cano, G.; Lazarus, M.; Saper, C.B. Parallel Preoptic Pathways for Thermoregulation. J. Neurosci. 2009, 29, 11954–11964. [Google Scholar] [CrossRef]
- Machado, N.L.S.; Bandaru, S.S.; Abbott, S.B.G.; Saper, C.B. EP3R-Expressing Glutamatergic Preoptic Neurons Mediate Inflammatory Fever. J. Neurosci. 2020, 40, 2573–2588. [Google Scholar] [CrossRef]
- Varin, C.; Rancillac, A.; Geoffroy, H.; Arthaud, S.; Fort, P.; Gallopin, T. Glucose Induces Slow-Wave Sleep by Exciting the Sleep-Promoting Neurons in the Ventrolateral Preoptic Nucleus: A New Link between Sleep and Metabolism. J. Neurosci. 2015, 35, 9900–9911. [Google Scholar] [CrossRef] [Green Version]
- Koster, J.C.; Marshall, B.A.; Ensor, N.; Corbett, J.A.; Nichols, C.G. Targeted Overactivity of Beta Cell K(ATP) Channels Induces Profound Neonatal Diabetes. Cell 2000, 100, 645–654. [Google Scholar] [CrossRef] [Green Version]
- Parton, L.E.; Ye, C.P.; Coppari, R.; Enriori, P.J.; Choi, B.; Zhang, C.-Y.; Xu, C.; Vianna, C.R.; Balthasar, N.; Lee, C.E.; et al. Glucose Sensing by POMC Neurons Regulates Glucose Homeostasis and Is Impaired in Obesity. Nature 2007, 449, 228–232. [Google Scholar] [CrossRef]
- Yoon, N.A.; Diano, S. Hypothalamic Glucose-Sensing Mechanisms. Diabetologia 2021, 64, 985–993. [Google Scholar] [CrossRef]
- Burdakov, D.; Jensen, L.T.; Alexopoulos, H.; Williams, R.H.; Fearon, I.M.; O’Kelly, I.; Gerasimenko, O.; Fugger, L.; Verkhratsky, A. Tandem-Pore K+ Channels Mediate Inhibition of Orexin Neurons by Glucose. Neuron 2006, 50, 711–722. [Google Scholar] [CrossRef] [PubMed]
- Danguir, J.; Nicolaidis, S. Dependence of Sleep on Nutrients’ Availability. Physiol. Behav. 1979, 22, 735–740. [Google Scholar] [CrossRef]
- Yamanaka, A.; Beuckmann, C.T.; Willie, J.T.; Hara, J.; Tsujino, N.; Mieda, M.; Tominaga, M.; Yagami, K.; Sugiyama, F.; Goto, K.; et al. Hypothalamic Orexin Neurons Regulate Arousal According to Energy Balance in Mice. Neuron 2003, 38, 701–713. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, J.B.; Omori, T.; Guan, Z.; Vgontzas, A.N.; Bixler, E.O.; Fang, J. Sleep Is Increased in Mice with Obesity Induced by High-Fat Food. Physiol. Behav. 2006, 87, 255–262. [Google Scholar] [CrossRef]
- Northeast, R.C.; Vyazovskiy, V.V.; Bechtold, D.A. Eat, Sleep, Repeat: The Role of the Circadian System in Balancing Sleep-Wake Control with Metabolic Need. Curr. Opin. Physiol. 2020, 15, 183–191. [Google Scholar] [CrossRef]
- Lydic, R.; Biebuyck, J.F. Sleep Neurobiology: Relevance for Mechanistic Studies of Anaesthesia. Br. J. Anaesth. 1994, 72, 506–508. [Google Scholar] [CrossRef]
- Mashour, G.A.; Hudetz, A.G. Bottom-Up and Top-Down Mechanisms of General Anesthetics Modulate Different Dimensions of Consciousness. Front. Neural. Circuits 2017, 11, 44. [Google Scholar] [CrossRef] [Green Version]
- Nelson, L.E.; Guo, T.Z.; Lu, J.; Saper, C.B.; Franks, N.P.; Maze, M. The Sedative Component of Anesthesia Is Mediated by GABA(A) Receptors in an Endogenous Sleep Pathway. Nat. Neurosci. 2002, 5, 979–984. [Google Scholar] [CrossRef]
- Nelson, L.E.; Lu, J.; Guo, T.; Saper, C.B.; Franks, N.P.; Maze, M. The Alpha2-Adrenoceptor Agonist Dexmedetomidine Converges on an Endogenous Sleep-Promoting Pathway to Exert Its Sedative Effects. Anesthesiology 2003, 98, 428–436. [Google Scholar] [CrossRef] [Green Version]
- Li, K.Y.; Guan, Y.; Krnjević, K.; Ye, J.H. Propofol Facilitates Glutamatergic Transmission to Neurons of the Ventrolateral Preoptic Nucleus. Anesthesiology 2009, 111, 1271–1278. [Google Scholar] [CrossRef] [Green Version]
- Tung, A.; Szafran, M.J.; Bluhm, B.; Mendelson, W.B. Sleep Deprivation Potentiates the Onset and Duration of Loss of Righting Reflex Induced by Propofol and Isoflurane. Anesthesiology 2002, 97, 906–911. [Google Scholar] [CrossRef] [PubMed]
- Saper, C.B.; Chou, T.C.; Scammell, T.E. The Sleep Switch: Hypothalamic Control of Sleep and Wakefulness. Trends Neurosci. 2001, 24, 726–731. [Google Scholar] [CrossRef]
- Szymusiak, R.; Gvilia, I.; McGinty, D. Hypothalamic Control of Sleep. Sleep Med. 2007, 8, 291–301. [Google Scholar] [CrossRef]
- Steininger, T.L.; Gong, H.; McGinty, D.; Szymusiak, R. Subregional Organization of Preoptic Area/Anterior Hypothalamic Projections to Arousal-Related Monoaminergic Cell Groups. J. Comp. Neurol. 2001, 429, 638–653. [Google Scholar] [CrossRef]
- Scharbarg, E.; Daenens, M.; Lemaitre, F.; Geoffroy, H.; Guille-Collignon, M.; Gallopin, T.; Rancillac, A. Astrocyte-Derived Adenosine Is Central to the Hypnogenic Effect of Glucose. Sci. Rep. 2016, 6, 19107. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, F.; Gómez-Extremera, M.; Bernaola-Galván, P.; Vetrivelan, R.; Saper, C.B.; Scammell, T.E.; Ivanov, P.C. Critical Dynamics and Coupling in Bursts of Cortical Rhythms Indicate Non-Homeostatic Mechanism for Sleep-Stage Transitions and Dual Role of VLPO Neurons in Both Sleep and Wake. J. Neurosci. 2020, 40, 171–190. [Google Scholar] [CrossRef] [PubMed]
- Vetrivelan, R.; Saper, C.B.; Fuller, P.M. Armodafinil-Induced Wakefulness in Animals with Ventrolateral Preoptic Lesions. Nat. Sci. Sleep 2014, 6, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.L.; Cooke, E.K.; Leib, D.E.; Lin, Y.-C.; Daly, G.E.; Zimmerman, C.A.; Knight, Z.A. Warm-Sensitive Neurons That Control Body Temperature. Cell 2016, 167, 47–59.e15. [Google Scholar] [CrossRef] [Green Version]
- Szymusiak, R. Body Temperature and Sleep. Handb. Clin. Neurol. 2018, 156, 341–351. [Google Scholar] [CrossRef]
- Rothhaas, R.; Chung, S. Role of the Preoptic Area in Sleep and Thermoregulation. Front. Neurosci. 2021, 15, 664781. [Google Scholar] [CrossRef]
- Harding, E.C.; Franks, N.P.; Wisden, W. Sleep and Thermoregulation. Curr. Opin. Physiol. 2020, 15, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.L.; Knight, Z.A. Regulation of Body Temperature by the Nervous System. Neuron 2018, 98, 31–48. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Luo, Z.; Zhang, Y.; Zhang, Y.; Wang, Y.; Cao, S.; Fu, B.; Yang, H.; Zhang, L.; Zhou, W.; et al. GABAergic Ventrolateral Pre-optic Nucleus Neurons Are Involved in the Mediation of the Anesthetic Hypnosis Induced by Propofol. Mol. Med. Rep. 2017, 16, 3179–3186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
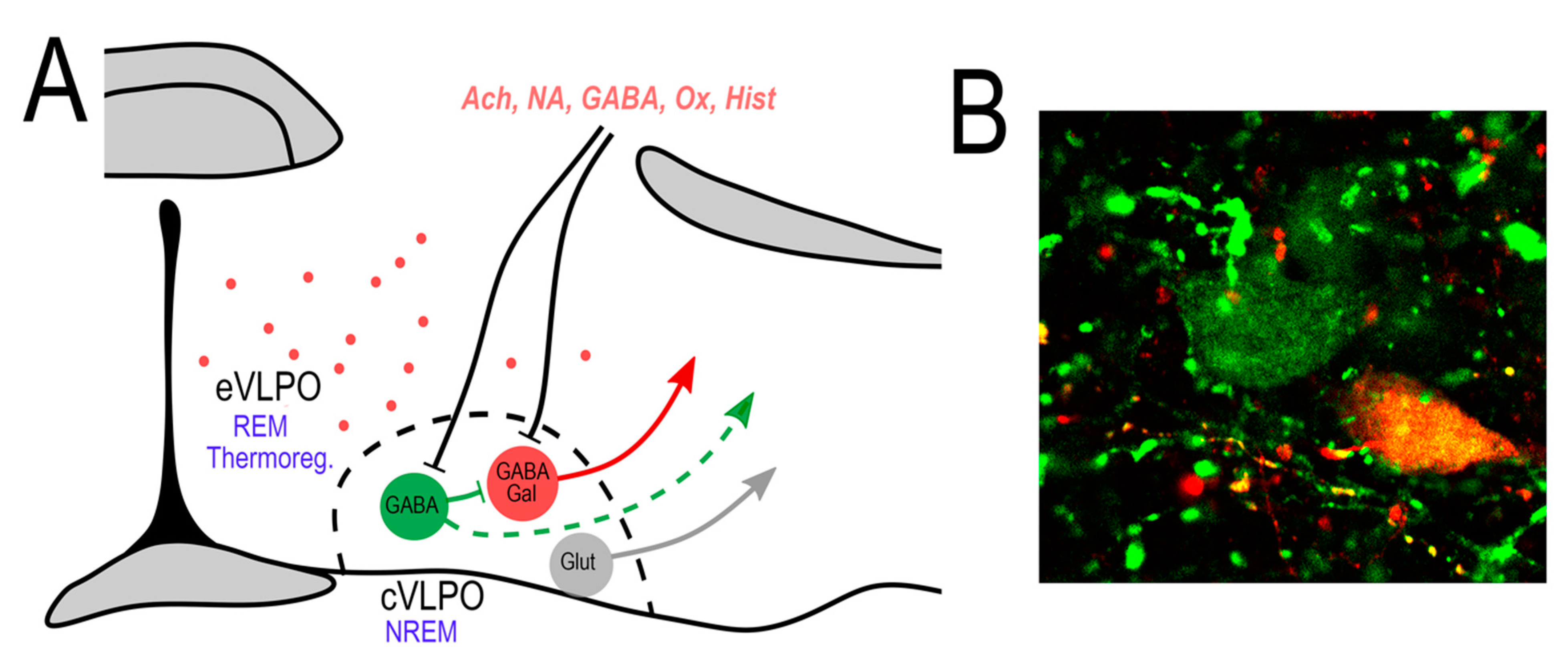
Figure 1.
Cellular anatomy of the VLPO. (A) The VLPO core (cVLPO) contains several distinct neuronal populations including: (1) GABAergic neurons that express galanin (VLPOGal), which are NREM sleep-promoting neurons; (2) GABAergic neurons that do not express galanin (VLPOGABA), which possibly suppress the activity of the VLPOGal neurons; and (3) glutamatergic neurons that presumptively promote arousal. There is also a subgroup of galanin neurons scattered in a region dorsal and medial to the core of VLPO, in the so-called extended VLPO (eVLPO). These neurons are involved in REM sleep and possibly in thermoregulation. Afferents to the cVLPO regulate the VLPOGal sleep-promoting neurons directly and/or indirectly through local VLPOGABA neurons. These VLPOGABA neurons might also project outside of the VLPO core. (B) Confocal image showing VLPOGal neurons (double labeled) and VLPOGABA neurons (in green). In situ hybridization for Vgat in green and galanin in red.
Figure 1.
Cellular anatomy of the VLPO. (A) The VLPO core (cVLPO) contains several distinct neuronal populations including: (1) GABAergic neurons that express galanin (VLPOGal), which are NREM sleep-promoting neurons; (2) GABAergic neurons that do not express galanin (VLPOGABA), which possibly suppress the activity of the VLPOGal neurons; and (3) glutamatergic neurons that presumptively promote arousal. There is also a subgroup of galanin neurons scattered in a region dorsal and medial to the core of VLPO, in the so-called extended VLPO (eVLPO). These neurons are involved in REM sleep and possibly in thermoregulation. Afferents to the cVLPO regulate the VLPOGal sleep-promoting neurons directly and/or indirectly through local VLPOGABA neurons. These VLPOGABA neurons might also project outside of the VLPO core. (B) Confocal image showing VLPOGal neurons (double labeled) and VLPOGABA neurons (in green). In situ hybridization for Vgat in green and galanin in red.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Arrigoni, E.; Fuller, P.M. The Sleep-Promoting Ventrolateral Preoptic Nucleus: What Have We Learned over the Past 25 Years? Int. J. Mol. Sci. 2022, 23, 2905. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23062905
AMA Style
Arrigoni E, Fuller PM. The Sleep-Promoting Ventrolateral Preoptic Nucleus: What Have We Learned over the Past 25 Years? International Journal of Molecular Sciences. 2022; 23(6):2905. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23062905
Chicago/Turabian StyleArrigoni, Elda, and Patrick M. Fuller. 2022. "The Sleep-Promoting Ventrolateral Preoptic Nucleus: What Have We Learned over the Past 25 Years?" International Journal of Molecular Sciences 23, no. 6: 2905. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23062905
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.