
Dual-Function Semaphorin 4D Released by Platelets: Suppression of Osteoblastogenesis and Promotion of Osteoclastogenesis


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
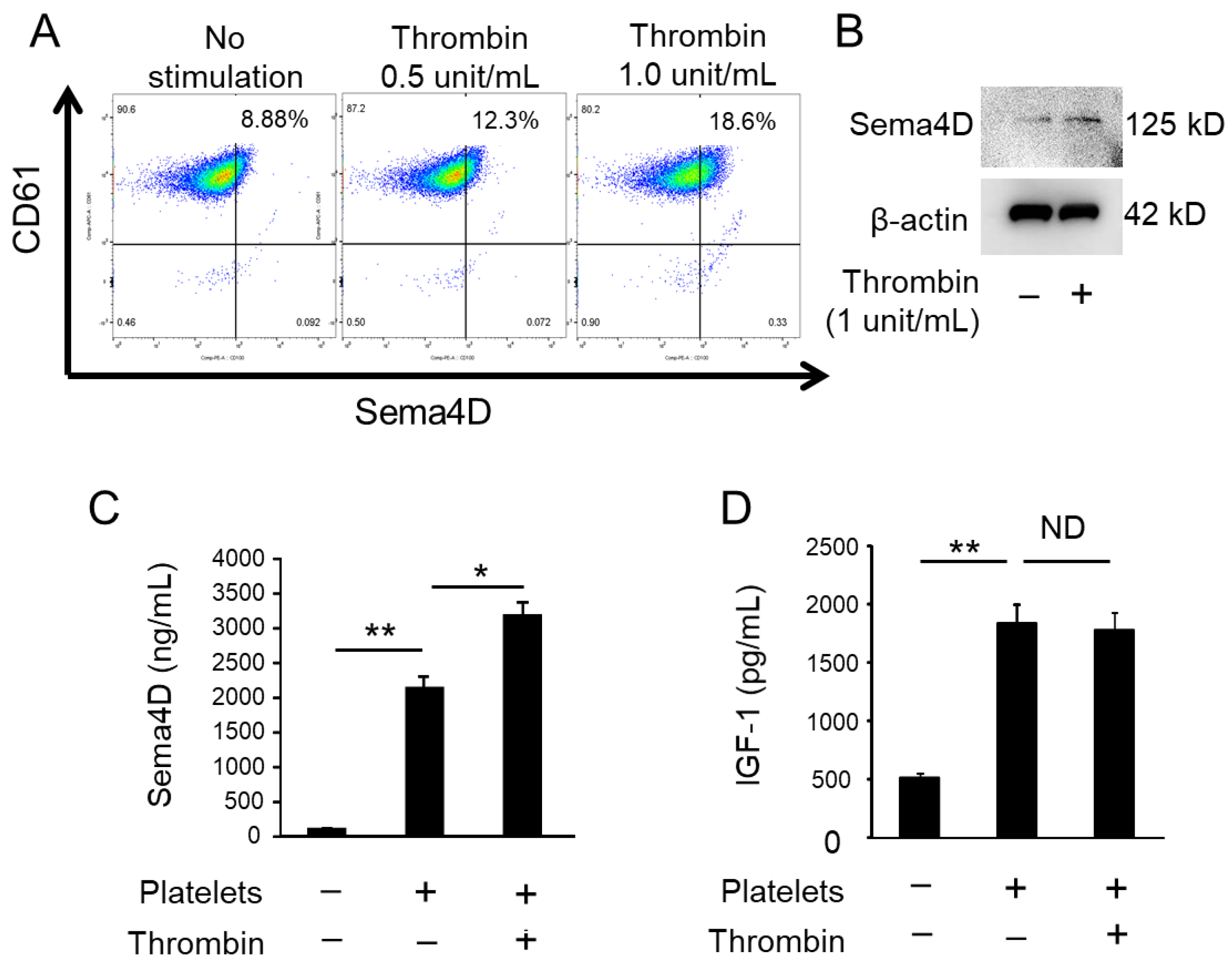
2.1. Activated Platelets Produce Sema4D and IGF-1
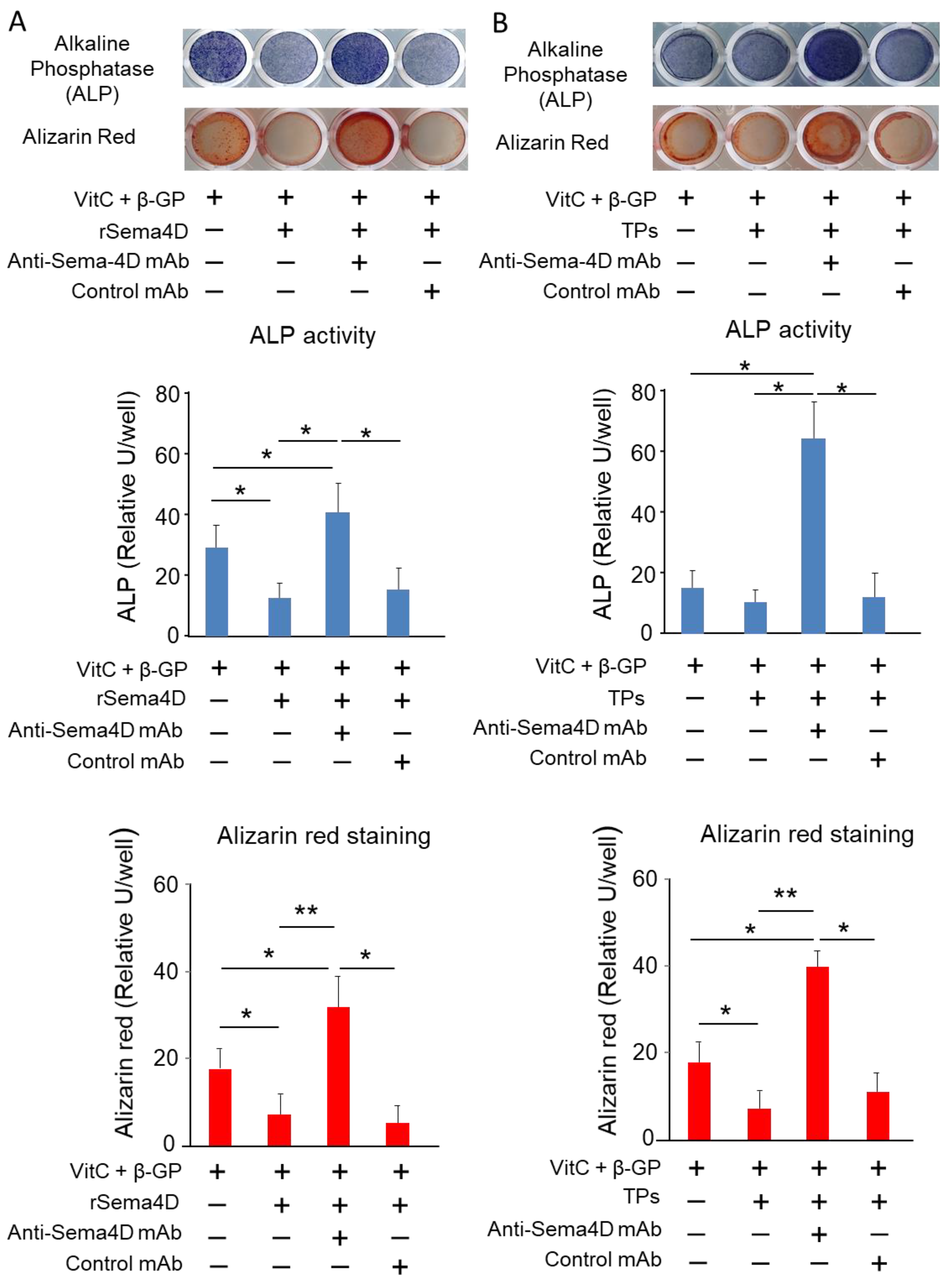
2.2. Sema4D Released from TPs Downmodulates Osteoblastogenesis
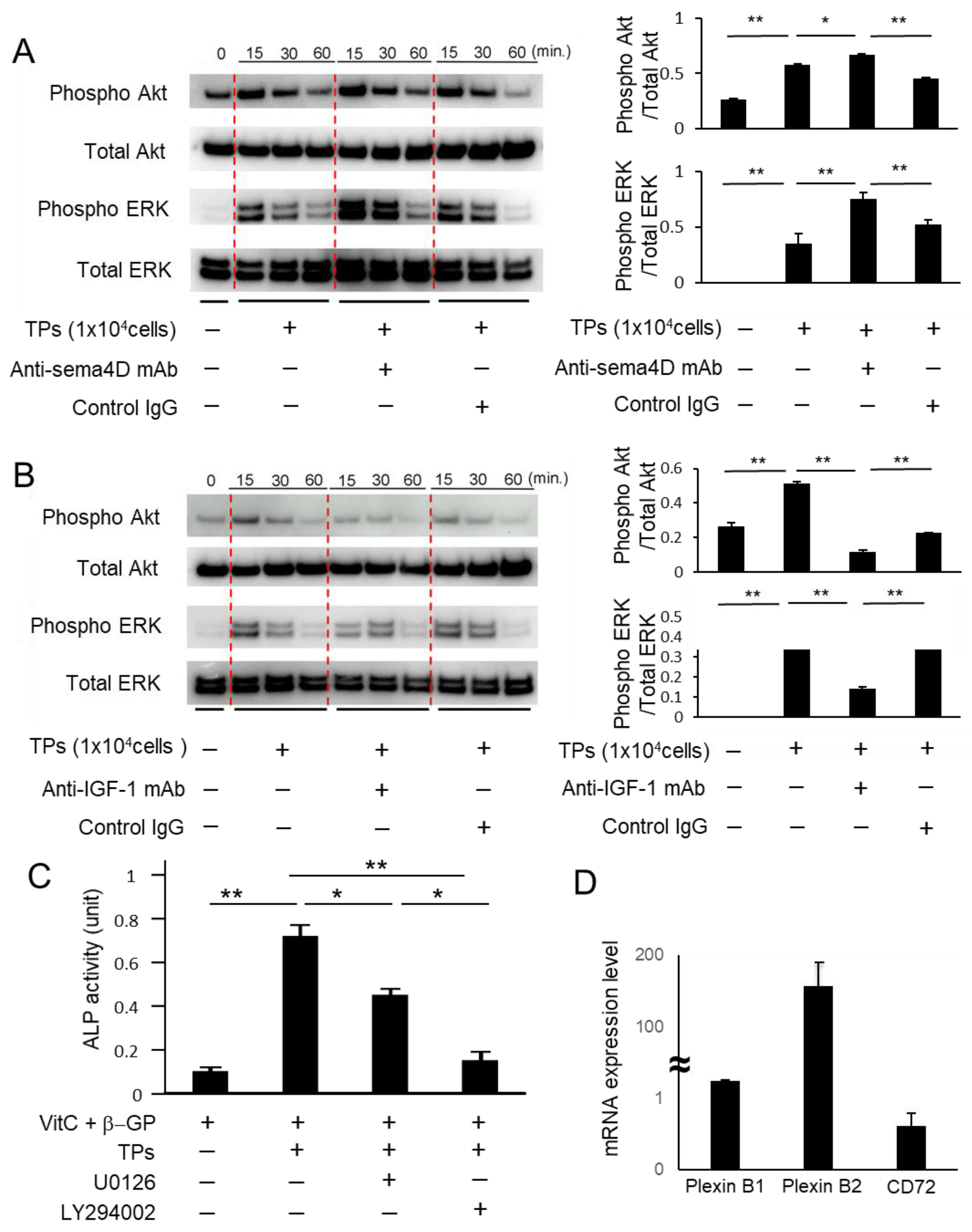
2.3. Sema4D from TPs Suppressed IGF-1-Mediated Osteoblastogenesis via the Akt/ERK Axis
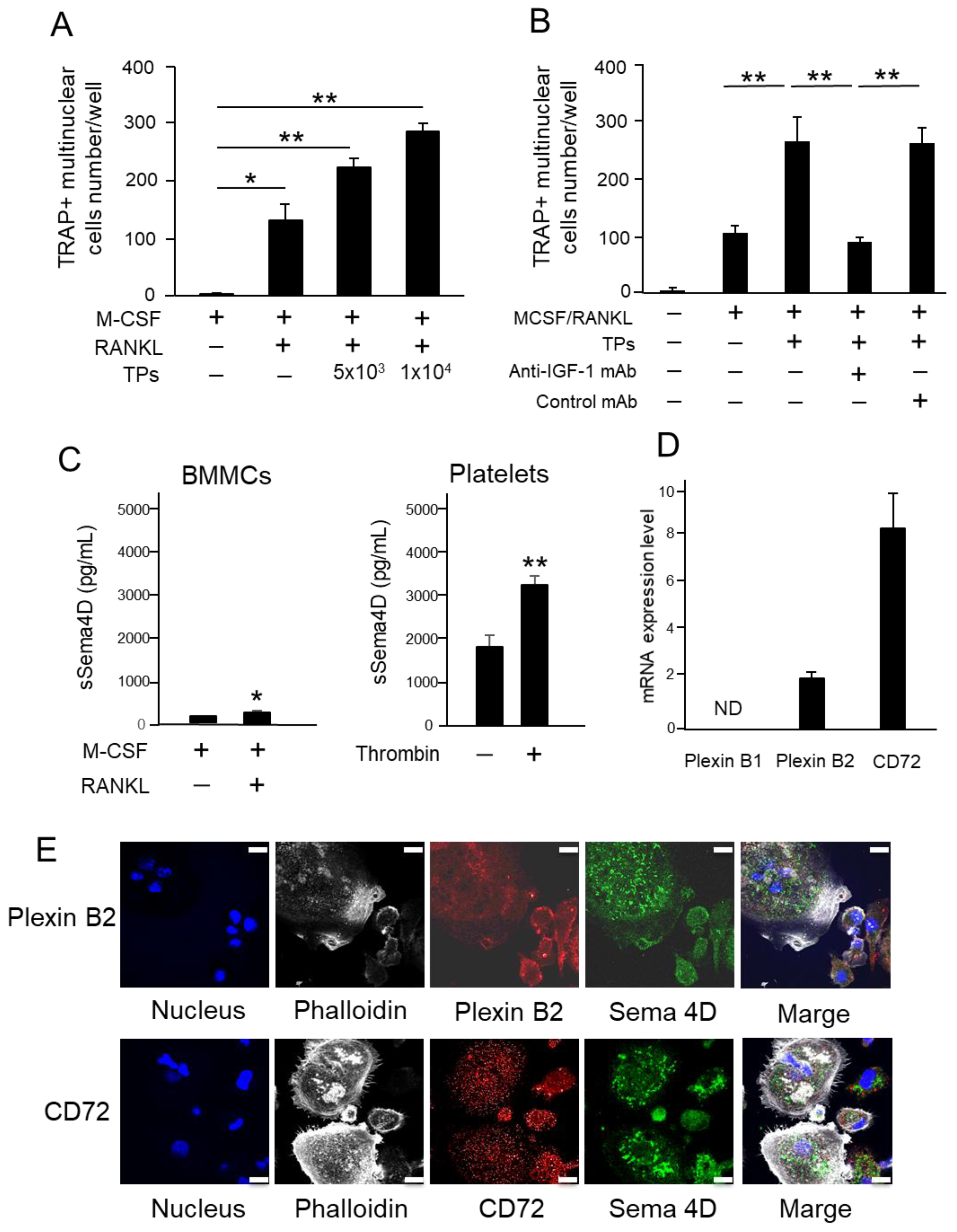
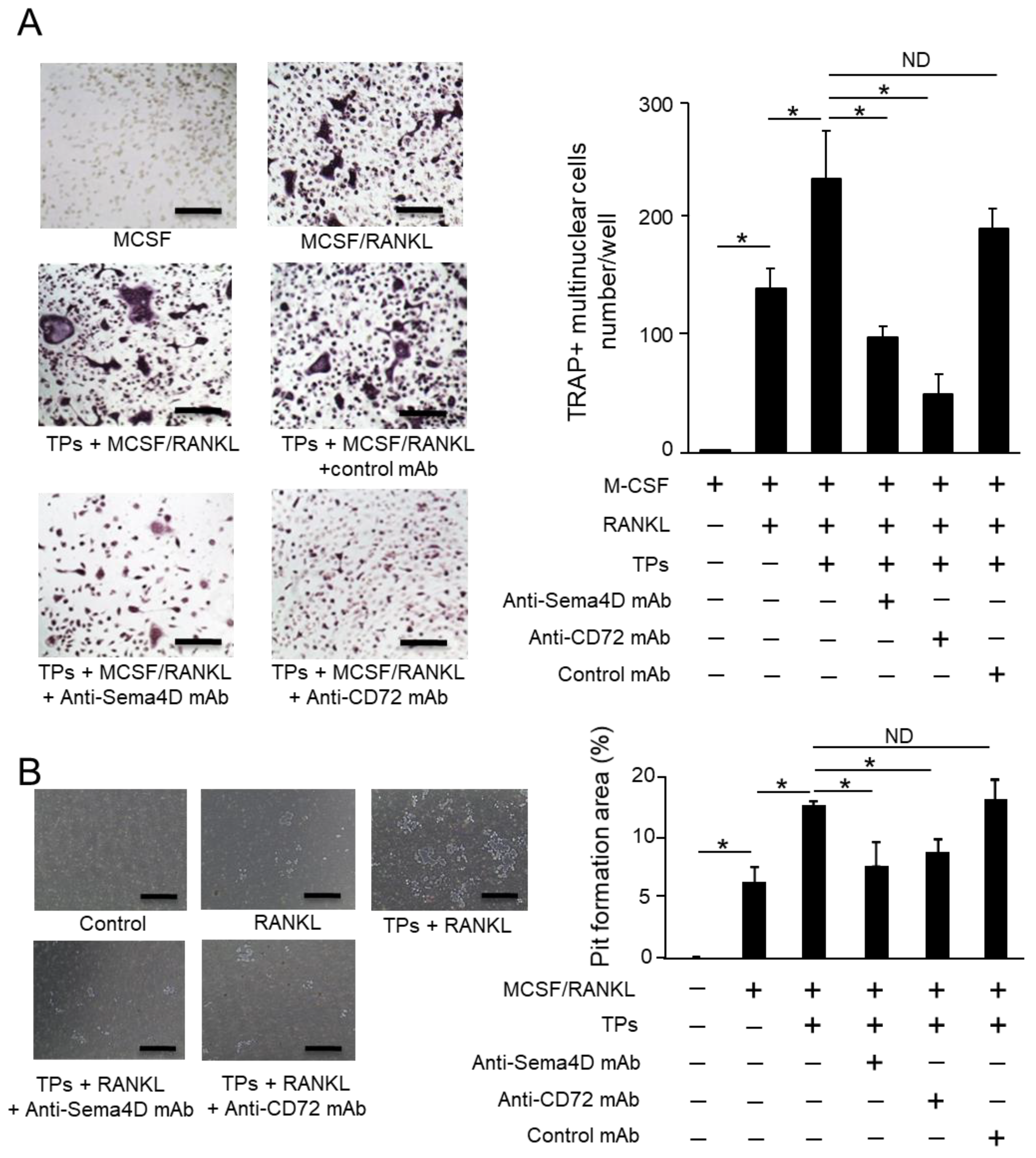
2.4. TPs Increase RANKL-Mediated Osteoclastogenesis and Sema4D Receptor Expression on BMMCs
2.5. Sema4D Released from TPs Increases RANKL-Mediated Osteoclastogenesis from BMMCs
3. Discussion
4. Materials and Methods
4.1. Platelet Isolation from Blood
4.2. Design of Anti-Sema4D-mAb and anti-CD72 mAb
4.3. Animals
4.4. Mouse Bone Marrow Cell Culture and Osteoclast Differentiation
4.5. Pit Formation Assay
4.6. Osteoblastogenesis Assay
4.7. Real-Time RT-PCR
4.8. Flow Cytometry
4.9. Enzyme-Linked Immunosorbent Assay (ELISA)
4.10. Western Blotting
4.11. Histological Analysis
4.12. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Armitage, G.C. Periodontal diagnoses and classification of periodontal diseases. Periodontol. 2000 2004, 34, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Savage, A.; Eaton, K.A.; Moles, D.R.; Needleman, I. A systematic review of definitions of periodontitis and methods that have been used to identify this disease. J. Clin. Periodontol. 2009, 36, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Corbella, S.; Weinstein, R.; Francetti, L.; Taschieri, S.; Del Fabbro, M. Periodontal regeneration in aggressive periodontitis patients: A systematic review of the literature. J. Investig. Clin. Dent. 2017, 8, e12245. [Google Scholar] [CrossRef] [PubMed]
- Terheyden, H.; Stadlinger, B.; Sanz, M.; Garbe, A.I.; Meyle, J. Inflammatory reaction-communication of cells. Clin. Oral Implant. Res. 2013, 25, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, Z.; Hamdan, N.; Ikeda, Y.; Grynpas, M.; Ganss, B.; Glogauer, M. Natural graft tissues and synthetic biomaterials for periodontal and alveolar bone reconstructive applications: A review. Biomater. Res. 2017, 21, 9. [Google Scholar] [CrossRef]
- Shue, L.; Yufeng, Z.; Mony, U. Biomaterials for periodontal regeneration: A review of ceramics and polymers. Biomatter 2012, 2, 271. [Google Scholar] [CrossRef] [Green Version]
- Bottino, M.C.; Thomas, V.; Schmidt, G.; Vohra, Y.K.; Chu, T.-M.G.; Kowolik, M.J.; Janowski, G. Recent advances in the development of GTR/GBR membranes for periodontal regeneration—A materials perspective. Dent. Mater. 2012, 28, 703–721. [Google Scholar] [CrossRef]
- Raja, S.; Byakod, G.; Pudakalkatti, P. Growth factors in periodontal regeneration. Int. J. Dent. Hyg. 2009, 7, 82–89. [Google Scholar] [CrossRef]
- Marx, R.E. Platelet-rich plasma: Evidence to support its use. J. Oral Maxillofac. Surg. 2004, 62, 489–496. [Google Scholar] [CrossRef]
- Tözüm, T.F.; Demiralp, B. Platelet-rich plasma: A promising innovation in dentistry. J.-Can. Dent. Assoc. 2003, 69, 664. [Google Scholar]
- Anitua, E.; Troya, M.; Zalduendo, M.; Tejero, R.; Orive, G. Progress in the Use of Autologous Regenerative Platelet-based Therapies in Implant Dentistry. Curr. Pharm. Biotechnol. 2016, 17, 402–413. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Han, Q.; Chen, W.; Song, J.; Zhao, X.; Ouyang, Y.; Yuan, W.; Fan, C. Platelet-Rich Plasma Derived Growth Factors Contribute to Stem Cell Differentiation in Musculoskeletal Regeneration. Front. Chem. 2017, 5, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anitua, E. Plasma rich in growth factors: Preliminary results of use in the preparation of future sites for implants. Int. J. Oral Maxillofac. Implant. 1999, 14, 529–535. [Google Scholar]
- Lekovic, V.; Camargo, P.M.; Weinlaender, M.; Vasilic, N.; Aleksic, Z.; Kenney, E.B. Effectiveness of a combination of platelet-rich plasma, bovine porous bone mineral and guided tissue regeneration in the treatment of mandibular grade II molar furcations in humans. J. Clin. Periodontol. 2003, 30, 746–751. [Google Scholar] [CrossRef] [PubMed]
- Demir, B.; Sengün, D.; Berberoğlu, A. Clinical evaluation of platelet-rich plasma and bioactive glass in the treatment of intra-bony defects. J. Clin. Periodontol. 2007, 34, 709. [Google Scholar] [CrossRef]
- Raghoebar, G.M.; Schortinghuis, J.; Liem, R.S.B.; Ruben, J.L.; Van Der Wal, J.E.; Vissink, A. Does platelet-rich plasma promote remodeling of autologous bone grafts used for augmentation of the maxillary sinus floor? Clin. Oral Implants Res. 2005, 16, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Thor, A.; Wannfors, K.; Sennerby, L.; Rasmusson, L. Reconstruction of the Severely Resorbed Maxilla with Autogenous Bone, Platelet-Rich Plasma, and Implants: 1 -Year Results of a Controlled Prospective 5-Year Study. Clin. Implant Dent. Relat. Res. 2005, 7, 209–220. [Google Scholar] [CrossRef]
- Monov, G.; Fuerst, G.; Tepper, G.; Watzak, G.; Zechner, W.; Watzek, G. The effect of platelet-rich plasma upon implant stability measured by resonance frequency analysis in the lower anterior mandibles. Clin. Oral Implants Res. 2005, 16, 461–465. [Google Scholar] [CrossRef]
- Gavino Orduna, J.F.; Caviedes-Bucheli, J.; Manzanares Cespedes, M.C.; Berastegui Jimeno, E.; Martin Biedma, B.; Segura-Egea, J.J.; López-López, J. Use of Platelet-rich Plasma in Endodontic Procedures in Adults: Regeneration or Repair? A Report of 3 Cases with 5 Years of Follow-up. J. Endod. 2017, 43, 1294. [Google Scholar] [CrossRef]
- Jalaluddin, M.; Singh, D.K.; Jayanti, I.; Kulkarni, P.; Faizuddin, M.; Tarannum, F. Use of platelet rich plasma in the management of periodontal intra-osseous defects: A clinical study. J. Int. Soc. Prev. Community Dent. 2017, 7, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Al-Hamed, F.S.; Tawfik, M.A.-M.; Abdelfadil, E.; Al-Saleh, M.A. Efficacy of Platelet-Rich Fibrin After Mandibular Third Molar Extraction: A Systematic Review and Meta-Analysis. J. Oral Maxillofac. Surg. 2017, 75, 1124–1135. [Google Scholar] [CrossRef] [PubMed]
- Bougeret, C.; Mansur, I.G.; Dastot, H.; Schmid, M.; Mahouy, G.; Bensussan, A.; Boumsell, L. Increased surface expression of a newly identified 150-kDa dimer early after human T lymphocyte activation. J. Immunol. 1992, 148, 318–323. [Google Scholar] [PubMed]
- Dacquin, R.; Domenget, C.; Kumanogoh, A.; Kikutani, H.; Jurdic, P.; Machuca-Gayet, I. Control of Bone Resorption by Semaphorin 4D Is Dependent on Ovarian Function. PLoS ONE 2011, 6, e26627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negishi-Koga, T.; Shinohara, M.; Komatsu, N.; Bito, H.; Kodama, T.; Friedel, R.H.; Takayanagi, H. Suppression of bone formation by osteoclastic expression of semaphorin 4D. Nat. Med. 2011, 17, 1473–1480. [Google Scholar] [CrossRef]
- Liu, X.L.; Song, J.; Liu, K.J.; Wang, W.P.; Xu, C.; Zhang, Y.Z.; Liu, Y. Role of inhibition of osteogenesis function by Sema4D/Plexin-B1 signaling pathway in skeletal fluorosis in vitro. J. Huazhong Univ. Sci. Technol. Med. Sci. 2015, 35, 712. [Google Scholar] [CrossRef]
- Zhu, L.; Bergmeier, W.; Wu, J.; Jiang, H.; Stalker, T.; Cieslak, M.; Fan, R.; Boumsell, L.; Kumanogoh, A.; Kikutani, H.; et al. Regulated surface expression and shedding support a dual role for semaphorin 4D in platelet responses to vascular injury. Proc. Natl. Acad. Sci. USA 2007, 104, 1621–1626. [Google Scholar] [CrossRef] [Green Version]
- Wannemacher, K.M.; Zhu, L.; Jiang, H.; Fong, K.P.; Stalker, T.J.; Lee, D.; Tran, A.N.; Neeves, K.B.; Maloney, S.; Kumanogoh, A.; et al. Diminished contact-dependent reinforcement of Syk activation underlies impaired thrombus growth in mice lacking Semaphorin 4D. Blood 2010, 116, 5707–5715. [Google Scholar] [CrossRef] [Green Version]
- Movila, A.; Mawardi, H.; Nishimura, K.; Kiyama, T.; Egashira, K.; Kim, J.-Y.; Villa, A.; Sasaki, H.; Woo, S.-B.; Kawai, T. Possible pathogenic engagement of soluble Semaphorin 4D produced by γδT cells in medication-related osteonecrosis of the jaw (MRONJ). Biochem. Biophys. Res. Commun. 2016, 480, 42–47. [Google Scholar] [CrossRef]
- Kim, J.-M.; Yang, Y.-S.; Park, K.H.; Oh, H.; Greenblatt, M.B.; Shim, J.-H. The ERK MAPK Pathway Is Essential for Skeletal Development and Homeostasis. Int. J. Mol. Sci. 2019, 20, 1803. [Google Scholar] [CrossRef] [Green Version]
- Fujita, T.; Azuma, Y.-T.; Fukuyama, R.; Hattori, Y.; Yoshida, C.; Koida, M.; Ogita, K.; Komori, T. Runx2 induces osteoblast and chondrocyte differentiation and enhances their migration by coupling with PI3K-Akt signaling. J. Cell Biol. 2004, 166, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Wannemacher, K.M.; Wang, L.; Zhu, L.; Brass, L.F. The role of semaphorins and their receptors in platelets: Lessons learned from neuronal and immune synapses. Platelets 2011, 22, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Wannemacher, K.M.; Jiang, H.; Hess, P.R.; Shin, Y.; Suzuki-Inoue, K.; Brass, L.F. An expanded role for semaphorin 4D in platelets includes contact-dependent amplification of Clec-2 signaling. J. Thromb. Haemost. 2013, 11, 2190–2193. [Google Scholar] [CrossRef] [Green Version]
- Fréchette, J.-P.; Martineau, I.; Gagnon, G. Platelet-rich Plasmas: Growth Factor Content and Roles in Wound Healing. J. Dent. Res. 2005, 84, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Ohlsson, C.; Mohan, S.; Sjogren, K.; Tivesten, A.; Isgaard, J.; Isaksson, O.; Jansson, J.-O.; Svensson, J. The Role of Liver-Derived Insulin-Like Growth Factor-I. Endocr. Rev. 2009, 30, 494–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, C.J. IGF-I and osteoporosis. Clin. Lab. Med. 2000, 20, 591. [Google Scholar] [CrossRef]
- Weicht, B.; Maitz, P.; Kandler, B.; Fischer, M.B.; Watzek, G.; Gruber, R. Activated platelets positively regulate RANKL-mediated osteoclast differentiation. J. Cell. Biochem. 2007, 102, 1300–1307. [Google Scholar] [CrossRef]
- Li, D.-Q.; Wan, Q.-L.; Pathak, J.L.; Li, Z.-B. Platelet-derived growth factor BB enhances osteoclast formation and osteoclast precursor cell chemotaxis. J. Bone Miner. Metab. 2016, 35, 355–365. [Google Scholar] [CrossRef]
- Fox, S.W.; Evans, K.E.; Lovibond, A.C. Transforming growth factor-beta enables NFATc1 expression during osteoclastogenesis. Biochem. Biophys. Res. Commun. 2008, 366, 123. [Google Scholar] [CrossRef] [Green Version]
- Hemingway, F.; Taylor, R.; Knowles, H.J.; Athanasou, N.A. RANKL-independent human osteoclast formation with APRIL, BAFF, NGF, IGF I and IGF II. Bone 2011, 48, 938. [Google Scholar] [CrossRef]
- Kinoshita, H.; Orita, S.; Inage, K.; Fujimoto, K.; Shiga, Y.; Abe, K.; Inoue, M.; Norimoto, M.; Umimura, T.; Ishii, T.; et al. Freeze-Dried Platelet-Rich Plasma Induces Osteoblast Proliferation via Platelet-Derived Growth Factor Receptor-Mediated Signal Transduction. Asian Spine J. 2020, 14, 1–8. [Google Scholar] [CrossRef]
- De Gorter, D.J.; van Dinther, M.; Korchynskyi, O.; ten Dijke, P. Biphasic effects of transforming growth factor β on bone morphogenetic protein-induced osteoblast differentiation. J. Bone Miner. Res. 2011, 26, 1178. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhang, G.; Guo, C.; Pan, Y. Intracellular Ca2+ signaling mediates IGF-1-induced osteogenic differentiation in bone marrow mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2020, 527, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Tamagnone, L.; Artigiani, S.; Chen, H.; He, Z.; Ming, G.-L.; Song, H.-J.; Chedotal, A.; Winberg, M.L.; Goodman, C.S.; Poo, M.-M.; et al. Plexins Are a Large Family of Receptors for Transmembrane, Secreted, and GPI-Anchored Semaphorins in Vertebrates. Cell 1999, 99, 71–80. [Google Scholar] [CrossRef] [Green Version]
- Kumanogoh, A.; Watanabe, C.; Lee, I.; Wang, X.; Shi, W.; Araki, H.; Hirata, H.; Iwahori, K.; Uchida, J.; Yasui, T.; et al. Identification of CD72 as a Lymphocyte Receptor for the Class IV Semaphorin CD100: A Novel Mechanism for Regulating B Cell Signaling. Immunity 2000, 13, 621–631. [Google Scholar] [CrossRef] [Green Version]
- Masuda, K.; Furuyama, T.; Takahara, M.; Fujioka, S.; Kurinami, H.; Inagaki, S. Sema4D stimulates axonal outgrowth of embryonic DRG sensory neurones. Genes Cells 2004, 9, 821–829. [Google Scholar] [CrossRef]
- Ishida, I.; Kumanogoh, A.; Suzuki, K.; Akahani, S.; Noda, K.; Kikutani, H. Involvement of CD100, a lymphocyte semaphorin, in the activation of the human immune system via CD72: Implications for the regulation of immune and inflammatory responses. Int. Immunol. 2003, 15, 1027–1034. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; O’Sullivan, K.M.; Jones, L.K.; Semple, T.; Kumanogoh, A.; Kikutani, H.; Holdsworth, S.R.; Kitching, A.R. CD100 enhances dendritic cell and CD4+ cell activation leading to pathogenetic humoral responses and immune complex glomerulonephritis. J. Immunol. 2006, 177, 3406–3412. [Google Scholar] [CrossRef] [Green Version]
- Kataoka, T.R.; Kumanogoh, A.; Fukuishi, N.; Ueshima, C.; Hirata, M.; Moriyoshi, K.; Tsuruyama, T.; Haga, H. CD72 negatively regulates mouse mast cell functions and down-regulates the expression of KIT and FcεRIα. Int. Immunol. 2015, 27, 95. [Google Scholar] [CrossRef] [Green Version]
- Pan, C.; Baumgarth, N.; Parnes, J.R. CD72-Deficient Mice Reveal Nonredundant Roles of CD72 in B Cell Development and Activation. Immunity 1999, 11, 495–506. [Google Scholar] [CrossRef] [Green Version]
- Li, D.H.; Tung, J.W.; Tarner, I.H.; Snow, A.L.; Yukinari, T.; Ngernmaneepothong, R.; Martinez, O.M.; Parnes, J.R. CD72 Down-Modulates BCR-Induced Signal Transduction and Diminishes Survival in Primary Mature B Lymphocytes. J. Immunol. 2006, 176, 5321–5328. [Google Scholar] [CrossRef] [Green Version]
- Reche, P.A.; Glutting, J.-P.; Zhang, H.; Reinherz, E.L. Enhancement to the RANKPEP resource for the prediction of peptide binding to MHC molecules using profiles. Immunogenetics 2004, 56, 405–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shindo, S.; Savitri, I.J.; Ishii, T.; Ikeda, A.; Pierrelus, R.; Heidari, A.; Okubo, K.; Nakamura, S.; Kandalam, U.; Rawas-Qalaji, M.; et al. Dual-Function Semaphorin 4D Released by Platelets: Suppression of Osteoblastogenesis and Promotion of Osteoclastogenesis. Int. J. Mol. Sci. 2022, 23, 2938. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23062938
Shindo S, Savitri IJ, Ishii T, Ikeda A, Pierrelus R, Heidari A, Okubo K, Nakamura S, Kandalam U, Rawas-Qalaji M, et al. Dual-Function Semaphorin 4D Released by Platelets: Suppression of Osteoblastogenesis and Promotion of Osteoclastogenesis. International Journal of Molecular Sciences. 2022; 23(6):2938. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23062938
Chicago/Turabian StyleShindo, Satoru, Irma Josefina Savitri, Takenobu Ishii, Atsushi Ikeda, Roodelyne Pierrelus, Alireza Heidari, Keisuke Okubo, Shin Nakamura, Umadevi Kandalam, Mohamad Rawas-Qalaji, and et al. 2022. "Dual-Function Semaphorin 4D Released by Platelets: Suppression of Osteoblastogenesis and Promotion of Osteoclastogenesis" International Journal of Molecular Sciences 23, no. 6: 2938. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23062938