
Biochemical Basis of Xylooligosaccharide Utilisation by Gut Bacteria

 , , , and
, , , and
Abstract
:1. Introduction
2. Results and Discussion
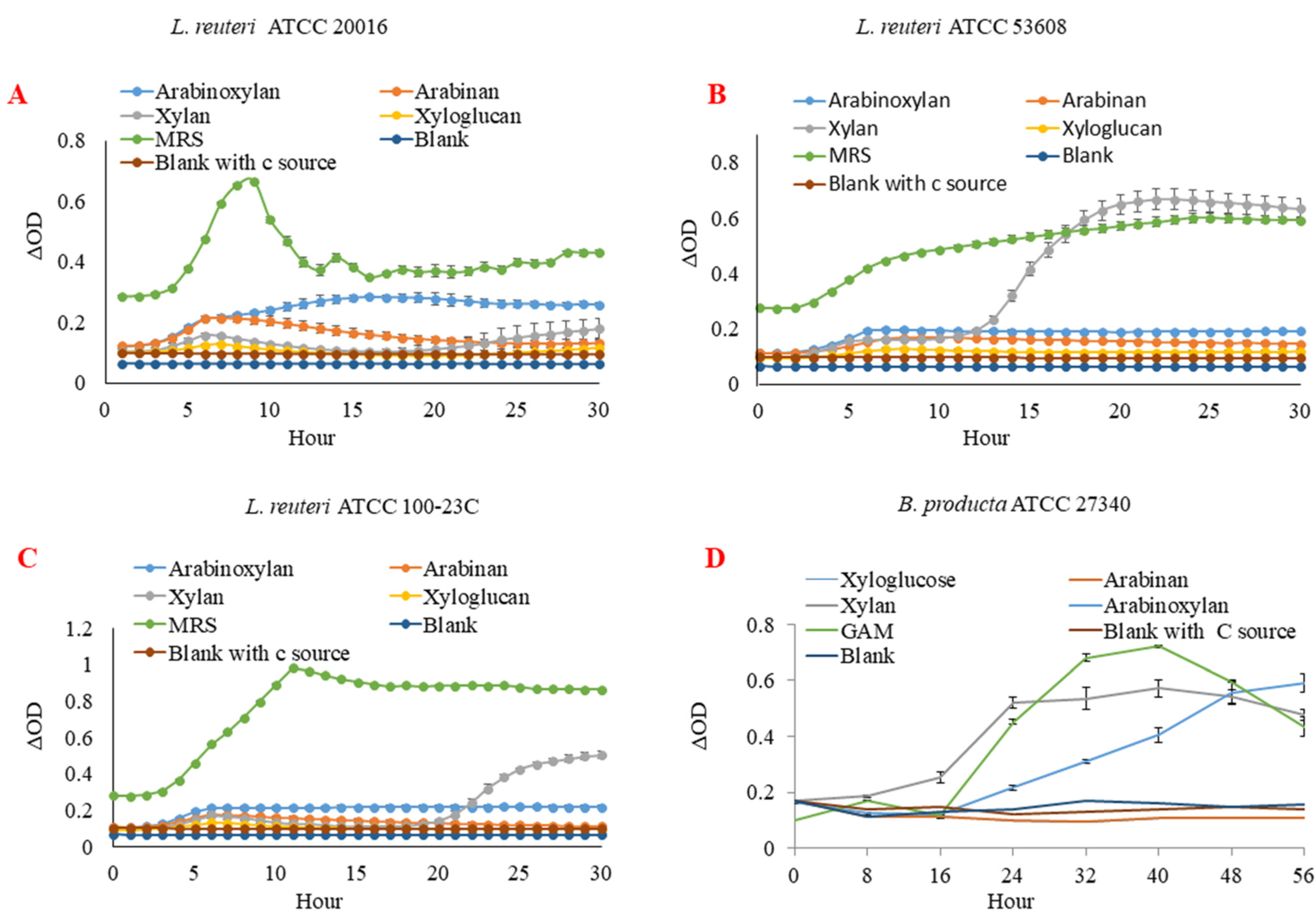
2.1. L. reuteri and B. producta Strains Utilise XOS as Metabolic Substrate
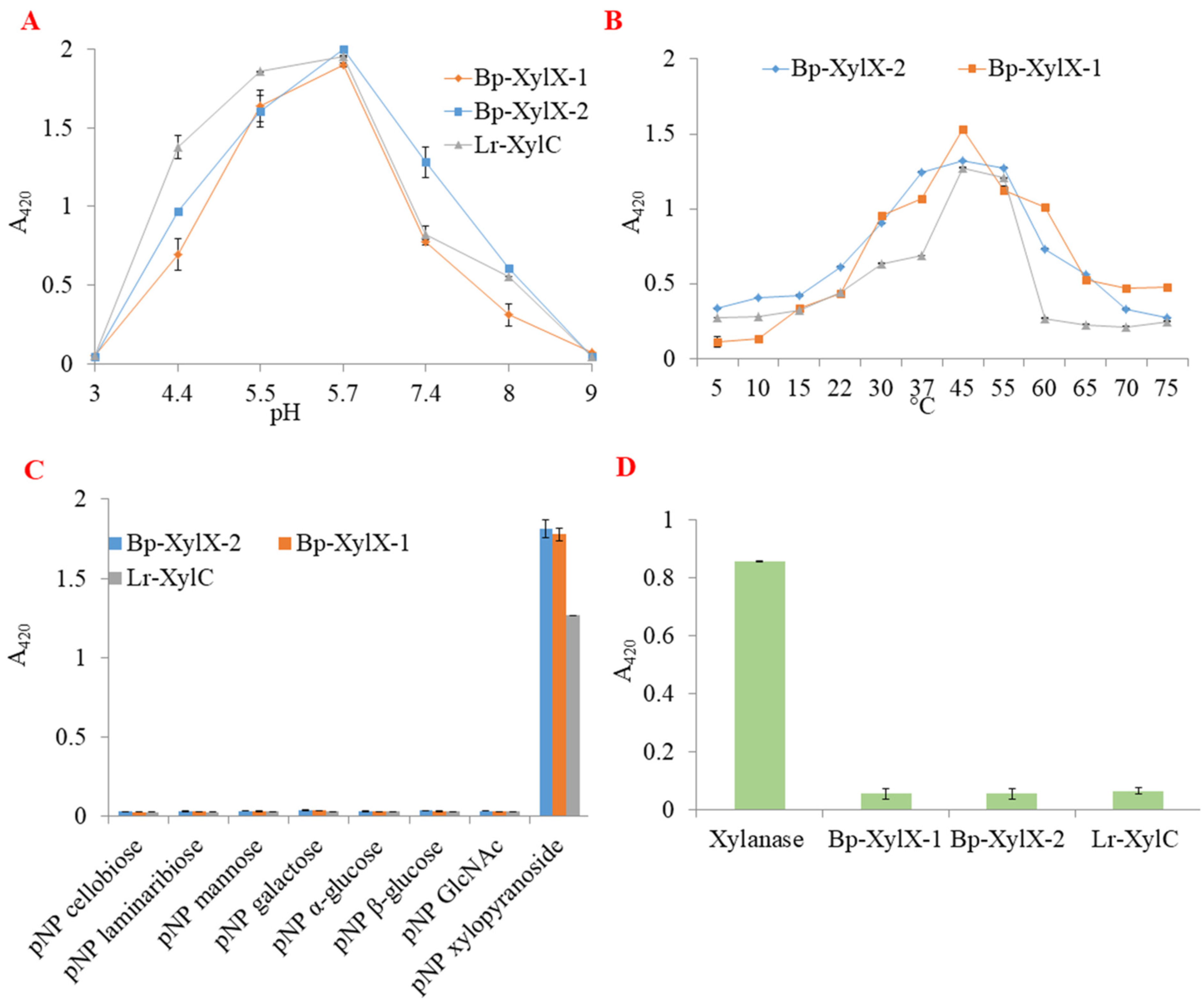
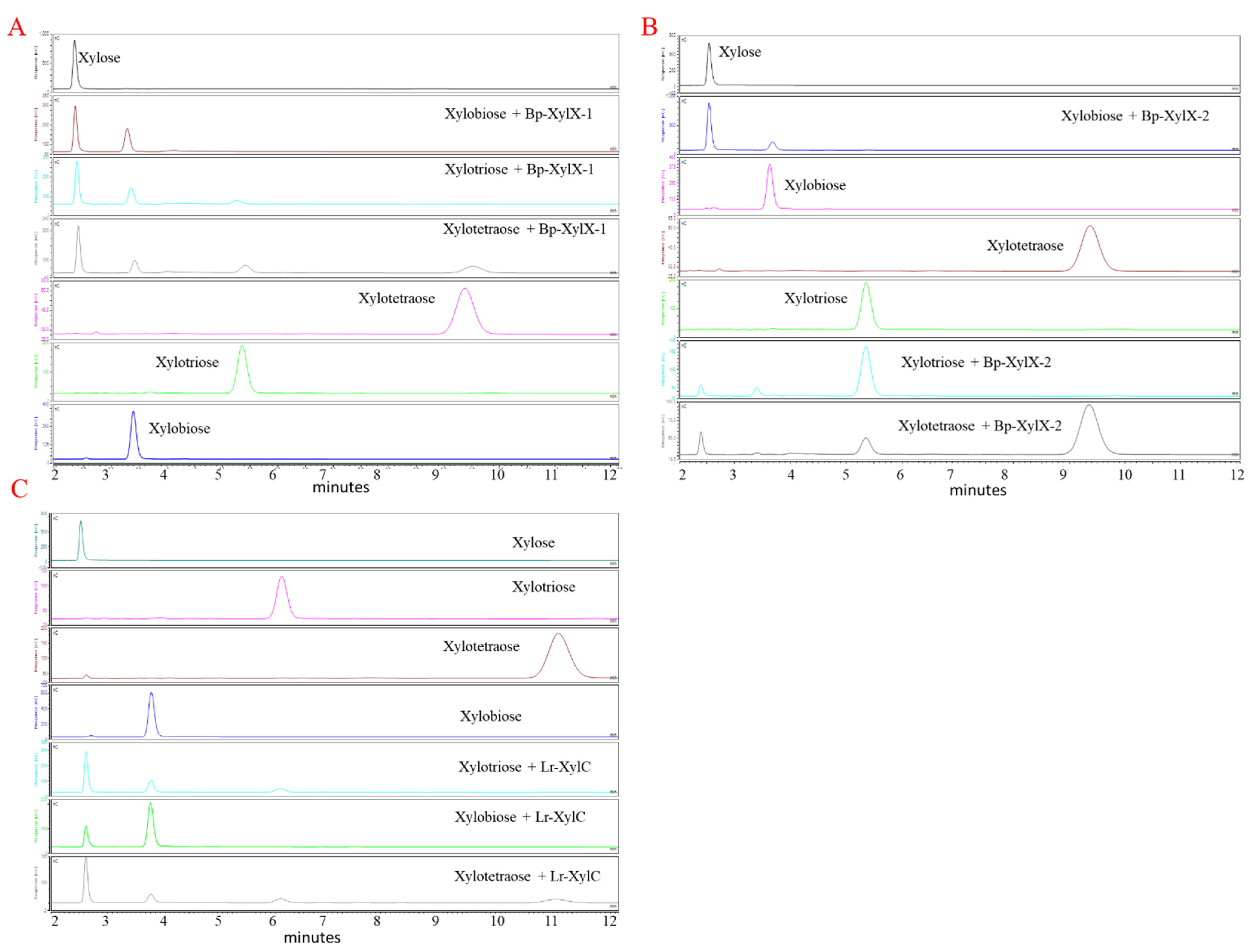
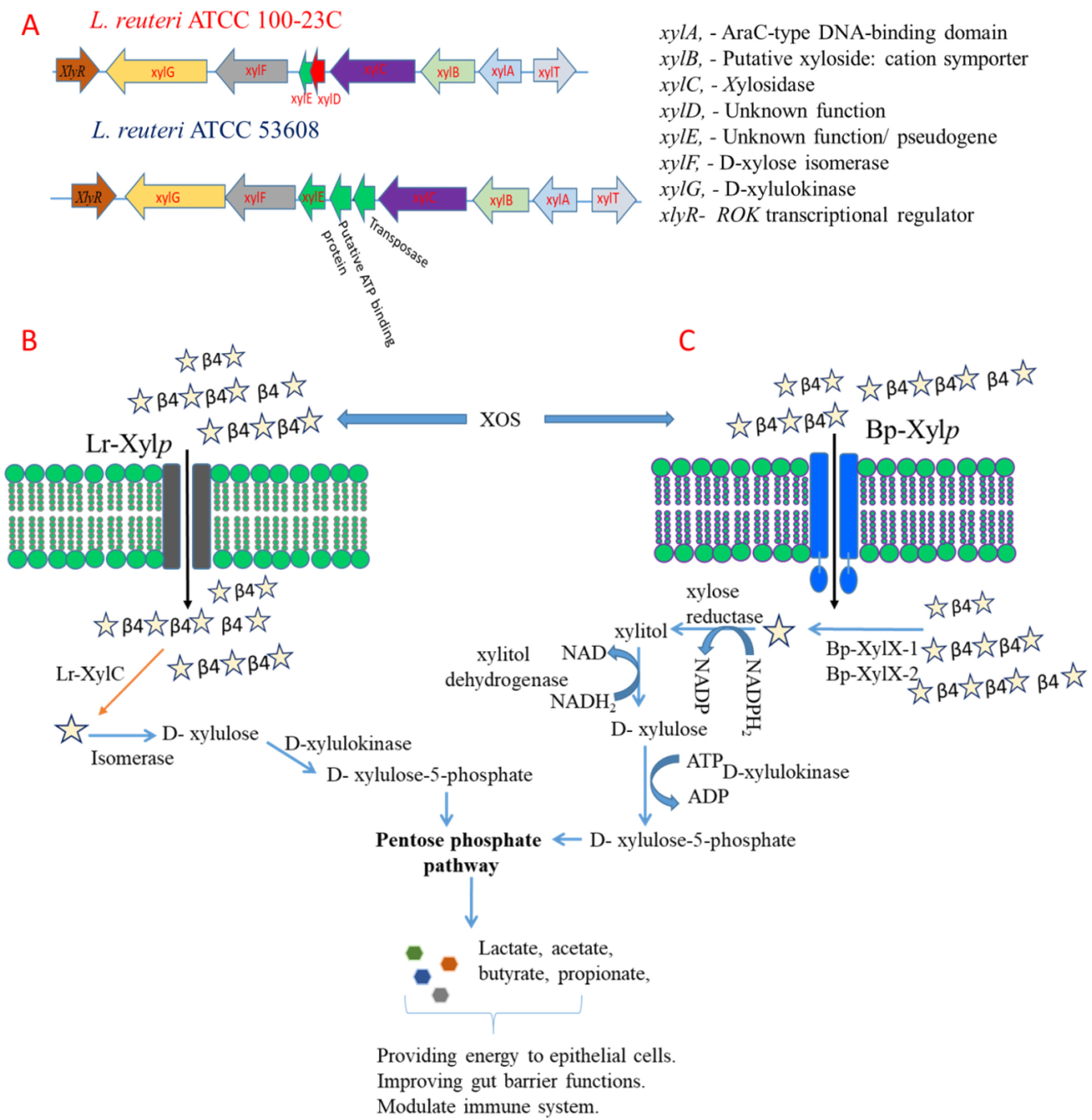
2.2. L. reuteri and B. Producta β-xylosidases Are Involved in XOS Utilisation
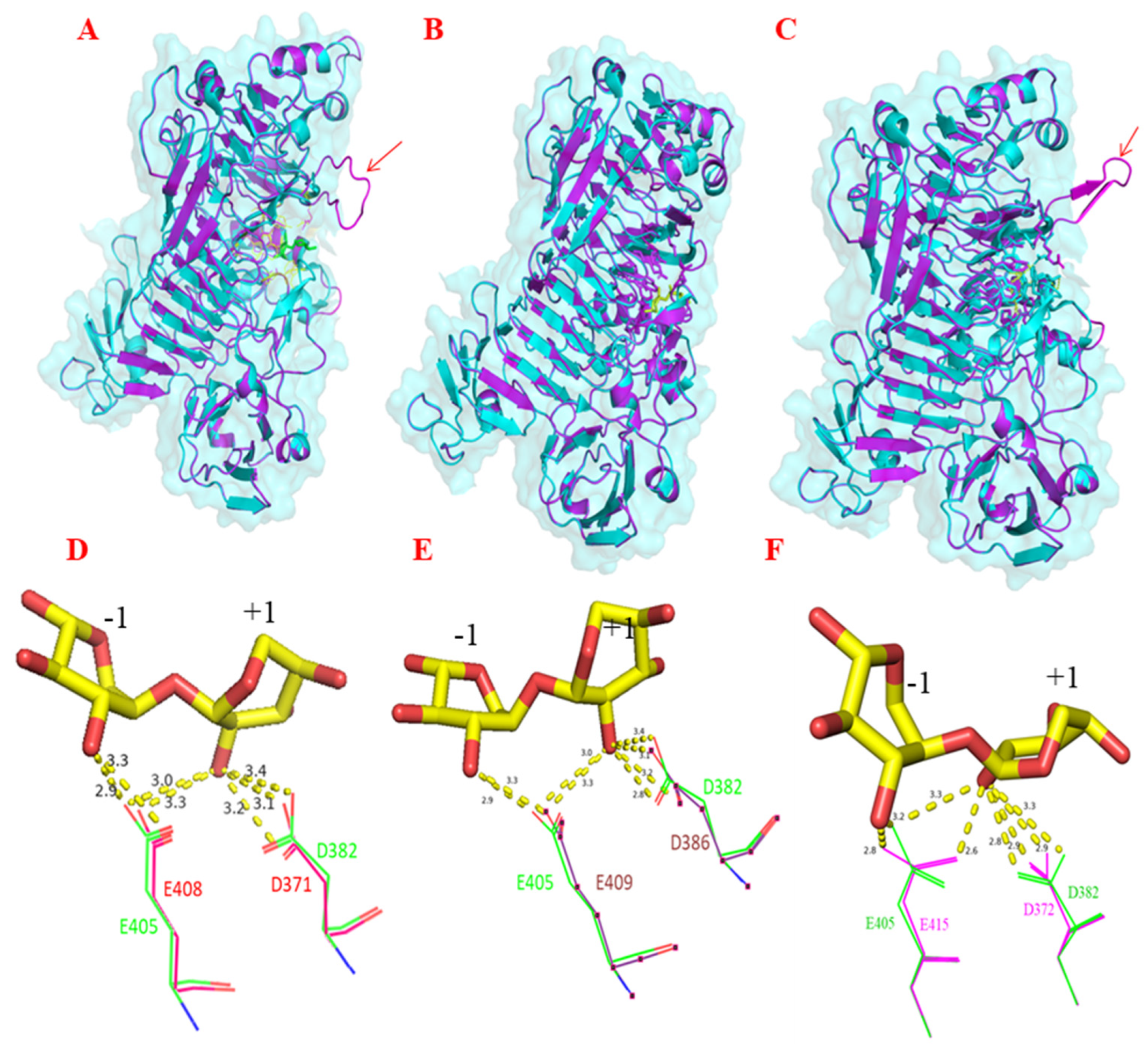
2.3. Proteomics and In Silico Analyses Identified the Complement of Proteins Involved in XOS Utilisation across Lactobacilli and Blautia Strains from Different Hosts
3. Materials and Methods
3.1. Bacterial Strains, Media, Carbon Source, and Growth Assays
3.2. Analysis of Xylooligosaccharides
3.3. Proteomics
3.4. In Silico Analyses
3.5. Cloning, Heterologous Expression, and Purification of Recombinant Enzymes
3.6. Enzymatic Activity Assays
3.7. Fluorophore-Assisted Carbohydrate Electrophoresis (FACE)
3.8. Site-Directed Mutagenesis
3.9. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Flint, H.J. The impact of nutrition on the human microbiome. Nutr. Rev. 2012, 70 (Suppl. 1), S10–S13. [Google Scholar] [CrossRef]
- Almeida, A.; Mitchell, A.L.; Boland, M.; Forster, S.C.; Gloor, G.B.; Tarkowska, A.; Lawley, T.D.; Finn, R.D. A new genomic blueprint of the human gut microbiota. Nature 2019, 568, 499–504. [Google Scholar] [CrossRef] [Green Version]
- Sommer, F.; Backhed, F. The gut microbiota—Masters of host development and physiology. Nat. Rev. Microbiol. 2013, 11, 227–238. [Google Scholar] [CrossRef]
- Belkaid, Y.; Hand, T.W. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [Green Version]
- Muresan, I.A.P.; Pop, L.L.; Dumitrascu, D.L. Lactobacillus reuteri versus triple therapy for the eradication of Helicobacter pylori in functional dyspepsia. Med. Pharm. Rep. 2019, 92, 352–355. [Google Scholar] [CrossRef]
- Servin, A.L. Antagonistic activities of lactobacilli and bifidobacteria against microbial pathogens. FEMS Microbiol. Rev. 2004, 28, 405–440. [Google Scholar] [CrossRef] [Green Version]
- Blaut, M. Gut microbiota and energy balance: Role in obesity. Proc. Nutr. Soc. 2015, 74, 227–234. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Jin, Y.; Zhang, J.; Pan, H.; Wu, L.; Liu, D.; Liu, J.; Hu, J.; Shen, J. Recovery of human gut microbiota genomes with third-generation sequencing. Cell Death Dis. 2021, 12, 569. [Google Scholar] [CrossRef]
- Forster, S.C.; Kumar, N.; Anonye, B.O.; Almeida, A.; Viciani, E.; Stares, M.D.; Dunn, M.; Mkandawire, T.T.; Zhu, A.; Shao, Y.; et al. A human gut bacterial genome and culture collection for improved metagenomic analyses. Nat. Biotechnol. 2019, 37, 186–192. [Google Scholar] [CrossRef]
- Ndeh, D.; Rogowski, A.; Cartmell, A.; Luis, A.S.; Basle, A.; Gray, J.; Venditto, I.; Briggs, J.; Zhang, X.; Labourel, A.; et al. Complex pectin metabolism by gut bacteria reveals novel catalytic functions. Nature 2017, 544, 65–70. [Google Scholar] [CrossRef]
- Jia, B.; Han, X.; Kim, K.H.; Jeon, C.O. Discovery and mining of enzymes from the human gut microbiome. Trends Biotechnol. 2021, 40, 240–254. [Google Scholar] [CrossRef]
- Ndeh, D.; Gilbert, H.J. Biochemistry of complex glycan depolymerisation by the human gut microbiota. FEMS Microbiol. Rev. 2018, 42, 146–164. [Google Scholar] [CrossRef] [Green Version]
- Hamaker, B.R.; Tuncil, Y.E. A perspective on the complexity of dietary fiber structures and their potential effect on the gut microbiota. J. Mol. Biol. 2014, 426, 3838–3850. [Google Scholar] [CrossRef]
- Terrapon, N.; Lombard, V.; Drula, E.; Lapebie, P.; Al-Masaudi, S.; Gilbert, H.J.; Henrissat, B. PULDB: The expanded database of polysaccharide utilization loci. Nucleic Acids Res. 2018, 46, D677–D683. [Google Scholar] [CrossRef]
- Lapebie, P.; Lombard, V.; Drula, E.; Terrapon, N.; Henrissat, B. Bacteroidetes use thousands of enzyme combinations to break down glycans. Nat. Commun. 2019, 10, 2043. [Google Scholar] [CrossRef]
- El Kaoutari, A.; Armougom, F.; Gordon, J.I.; Raoult, D.; Henrissat, B. The abundance and variety of carbohydrate-active enzymes in the human gut microbiota. Nat. Rev. Microbiol. 2013, 11, 497–504. [Google Scholar] [CrossRef]
- Bader Ul Ain, H.; Saeed, F.; Asif Khan, M.; Niaz, B.; Tufail, T.; Anjum, F.M.; Hussain, S.; Rohi, M. Isolation and characterization of cereal cell walls. Int. J. Food Prop. 2019, 22, 130–137. [Google Scholar] [CrossRef]
- Gilbert, H.J. The biochemistry and structural biology of plant cell wall deconstruction. Plant Physiol. 2010, 153, 444–455. [Google Scholar] [CrossRef] [Green Version]
- Dodd, D.; Mackie, R.I.; Cann, I.K. Xylan degradation, a metabolic property shared by rumen and human colonic Bacteroidetes. Mol. Microbiol. 2011, 79, 292–304. [Google Scholar] [CrossRef] [Green Version]
- Martens, E.C.; Lowe, E.C.; Chiang, H.; Pudlo, N.A.; Wu, M.; McNulty, N.P.; Abbott, D.W.; Henrissat, B.; Gilbert, H.J.; Bolam, D.N.; et al. Recognition and degradation of plant cell wall polysaccharides by two human gut symbionts. PLoS Biol. 2011, 9, e1001221. [Google Scholar] [CrossRef]
- Hao, Z.; Wang, X.; Yang, H.; Tu, T.; Zhang, J.; Luo, H.; Huang, H.; Su, X. PUL-mediated plant cell wall polysaccharide utilization in the gut Bacteroidetes. Int. J. Mol. Sci. 2021, 22, 3077. [Google Scholar] [CrossRef]
- Hong, P.Y.; Iakiviak, M.; Dodd, D.; Zhang, M.; Mackie, R.I.; Cann, I. Two new xylanases with different substrate specificities from the human gut bacterium Bacteroides intestinalis DSM 17393. Appl. Environ. Microbiol. 2014, 80, 2084–2093. [Google Scholar] [CrossRef] [Green Version]
- Leth, M.L.; Ejby, M.; Workman, C.; Ewald, D.A.; Pedersen, S.S.; Sternberg, C.; Bahl, M.I.; Licht, T.R.; Aachmann, F.L.; Westereng, B.; et al. Differential bacterial capture and transport preferences facilitate co-growth on dietary xylan in the human gut. Nat. Microbiol. 2018, 3, 570–580. [Google Scholar] [CrossRef]
- Rogowski, A.; Briggs, J.A.; Mortimer, J.C.; Tryfona, T.; Terrapon, N.; Lowe, E.C.; Basle, A.; Morland, C.; Day, A.M.; Zheng, H.; et al. Glycan complexity dictates microbial resource allocation in the large intestine. Nat. Commun. 2015, 6, 7481. [Google Scholar] [CrossRef] [Green Version]
- Broekaert, W.F.; Courtin, C.M.; Verbeke, K.; van de Wiele, T.; Verstraete, W.; Delcour, J.A. Prebiotic and other health-related effects of cereal-derived arabinoxylans, arabinoxylan-oligosaccharides, and xylooligosaccharides. Crit. Rev. Food Sci. Nutr. 2011, 51, 178–194. [Google Scholar] [CrossRef]
- Zeybek, N.; Rastall, R.A.; Buyukkileci, A.O. Utilization of xylan-type polysaccharides in co-culture fermentations of Bifidobacterium and Bacteroides species. Carbohydr. Polym. 2020, 236, 116076. [Google Scholar] [CrossRef]
- Pastell, H.; Westermann, P.; Meyer, A.S.; Tuomainen, P.; Tenkanen, M. In vitro fermentation of arabinoxylan-derived carbohydrates by Bifidobacteria and mixed fecal microbiota. J. Agric. Food Chem. 2009, 57, 8598–8606. [Google Scholar] [CrossRef]
- Odamaki, T.; Bottacini, F.; Kato, K.; Mitsuyama, E.; Yoshida, K.; Horigome, A.; Xiao, J.Z.; van Sinderen, D. Genomic diversity and distribution of Bifidobacterium longum subsp. longum across the human lifespan. Sci. Rep. 2018, 8, 85. [Google Scholar]
- Oh, P.L.; Benson, A.K.; Peterson, D.A.; Patil, P.B.; Moriyama, E.N.; Roos, S.; Walter, J. Diversification of the gut symbiont Lactobacillus reuteri as a result of host-driven evolution. ISME J. 2010, 4, 377–387. [Google Scholar] [CrossRef]
- Frese, S.A.; Benson, A.K.; Tannock, G.W.; Loach, D.M.; Kim, J.; Zhang, M.; Oh, P.L.; Heng, N.C.; Patil, P.B.; Juge, N.; et al. The evolution of host specialization in the vertebrate gut symbiont Lactobacillus reuteri. PLoS Genet. 2011, 7, e1001314. [Google Scholar] [CrossRef] [Green Version]
- Caballero, S.; Kim, S.; Carter, R.A.; Leiner, I.M.; Susac, B.; Miller, L.; Kim, G.J.; Ling, L.; Pamer, E.G. Cooperating commensals restore colonization resistance to vancomycin-resistant Enterococcus faecium. Cell Host Microbe 2017, 21, 592–602. [Google Scholar] [CrossRef]
- Kim, S.G.; Becattini, S.; Moody, T.U.; Shliaha, P.V.; Littmann, E.R.; Seok, R.; Gjonbalaj, M.; Eaton, V.; Fontana, E.; Amoretti, L.; et al. Microbiota-derived lantibiotic restores resistance against vancomycin-resistant Enterococcus. Nature 2019, 572, 665–669. [Google Scholar] [CrossRef]
- Walter, J.; Britton, R.A.; Roos, S. Host-microbial symbiosis in the vertebrate gastrointestinal tract and the Lactobacillus reuteri paradigm. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4645–4652. [Google Scholar] [CrossRef] [Green Version]
- MacKenzie, D.A.; Jeffers, F.; Parker, M.L.; Vibert-Vallet, A.; Bongaerts, R.J.; Roos, S.; Walter, J.; Juge, N. Strain-specific diversity of mucus-binding proteins in the adhesion and aggregation properties of Lactobacillus reuteri. Microbiology 2010, 156, 3368–3378. [Google Scholar] [CrossRef] [Green Version]
- Etzold, S.; Kober, O.I.; Mackenzie, D.A.; Tailford, L.E.; Gunning, A.P.; Walshaw, J.; Hemmings, A.M.; Juge, N. Structural basis for adaptation of lactobacilli to gastrointestinal mucus. Environ. Microbiol. 2014, 16, 888–903. [Google Scholar] [CrossRef]
- Liu, X.; Mao, B.; Gu, J.; Wu, J.; Cui, S.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Blautia-a new functional genus with potential probiotic properties? Gut Microbes 2021, 13, 1–21. [Google Scholar] [CrossRef]
- Xiao, X.; Wen, J.Y.; Wang, Y.Y.; Bian, J.; Li, M.F.; Peng, F.; Sun, R.C. NMR and ESI-MS spectrometry characterization of autohydrolysis xylo-oligosaccharides separated by gel permeation chromatography. Carbohydr. Polym. 2018, 195, 303–310. [Google Scholar] [CrossRef]
- Zhou, A.; Hu, Y.; Li, J.; Wang, W.; Zhang, M.; Guan, S. Characterization of a recombinant β-xylosidase of GH43 family from Bacteroides ovatus strain ATCC 8483. Biocatal. Biotransformation 2020, 38, 46–52. [Google Scholar] [CrossRef]
- Shin, H.Y.; Lee, J.H.; Lee, J.Y.; Han, Y.O.; Han, M.J.; Kim, D.H. Purification and characterization of ginsenoside Ra-hydrolyzing beta-D-xylosidase from Bifidobacterium breve K-110, a human intestinal anaerobic bacterium. Biol. Pharm. Bull. 2003, 26, 1170–1173. [Google Scholar] [CrossRef] [Green Version]
- Viborg, A.H.; Sorensen, K.I.; Gilad, O.; Steen-Jensen, D.B.; Dilokpimol, A.; Jacobsen, S.; Svensson, B. Biochemical and kinetic characterisation of a novel xylooligosaccharide-upregulated GH43 beta-d-xylosidase/alpha-l-arabinofuranosidase (BXA43) from the probiotic Bifidobacterium animalis subsp. lactis BB-12. AMB Express 2013, 3, 56. [Google Scholar] [CrossRef] [Green Version]
- Muzard, M.; Aubry, N.; Plantier-Royon, R.; O’Donohue, M.; Rémond, C. Evaluation of the transglycosylation activities of a GH 39 β-d-xylosidase for the synthesis of xylose-based glycosides. J. Mol. Catal. B Enzym. 2009, 58, 1–5. [Google Scholar] [CrossRef]
- Eneyskaya, E.V.; Brumer, H., 3rd; Backinowsky, L.V.; Ivanen, D.R.; Kulminskaya, A.A.; Shabalin, K.A.; Neustroev, K.N. Enzymatic synthesis of beta-xylanase substrates: Transglycosylation reactions of the beta-xylosidase from Aspergillus sp. Carbohydr. Res. 2003, 338, 313–325. [Google Scholar] [CrossRef]
- Eneyskaya, E.V.; Ivanen, D.R.; Shabalin, K.A.; Kulminskaya, A.A.; Backinowsky, L.V.; Brumer Iii, H.; Neustroev, K.N. Chemo-enzymatic synthesis of 4-methylumbelliferyl beta-(1-->4)-D-xylooligosides: New substrates for beta-D-xylanase assays. Org. Biomol. Chem. 2005, 3, 146–151. [Google Scholar] [CrossRef]
- Shao, W.; Xue, Y.; Wu, A.; Kataeva, I.; Pei, J.; Wu, H.; Wiegel, J. Characterization of a novel beta-xylosidase, XylC, from Thermoanaerobacterium saccharolyticum JW/SL-YS485. Appl. Environ. Microbiol. 2011, 77, 719–726. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Dai, L.; Zhang, W.; Yang, Y.; Wu, Q.; Li, J.; Tang, X.; Zhou, J.; Ding, J.; Han, N.; et al. Characterization of a novel salt-, xylose- and alkali-tolerant GH43 bifunctional beta-xylosidase/alpha-l-arabinofuranosidase from the gut bacterial genome. J. Biosci. Bioeng. 2019, 128, 429–437. [Google Scholar] [CrossRef]
- Li, Q.; Wu, T.; Qi, Z.; Zhao, L.; Pei, J.; Tang, F. Characterization of a novel thermostable and xylose-tolerant GH 39 beta-xylosidase from Dictyoglomus thermophilum. BMC Biotechnol. 2018, 18, 29. [Google Scholar] [CrossRef]
- Zanoelo, F.F.; Polizeli Md Mde, L.; Terenzi, H.F.; Jorge, J.A. Purification and biochemical properties of a thermostable xylose-tolerant beta- D-xylosidase from Scytalidium thermophilum. J. Ind. Microbiol. Biotechnol. 2004, 31, 170–176. [Google Scholar] [CrossRef]
- De Almeida, E.M.; Polizeli, M.d.L.T.M.; Terenzi, H.F.; Atilio Jorge, J. Purification and biochemical characterization of β-xylosidase from Humicola grisea var. thermoidea. FEMS Microbiol. Lett. 1995, 130, 171–175. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Ramón, D. Purification and regulation of the synthesis of a β-xylosidase from Aspergillus nidulans. FEMS Microbiol. Lett. 1996, 135, 287–293. [Google Scholar] [CrossRef]
- Huang, C.H.; Sun, Y.; Ko, T.P.; Chen, C.C.; Zheng, Y.; Chan, H.C.; Pang, X.; Wiegel, J.; Shao, W.; Guo, R.T. The substrate/product-binding modes of a novel GH120 beta-xylosidase (XylC) from Thermoanaerobacterium saccharolyticum JW/SL-YS485. Biochem. J. 2012, 448, 401–407. [Google Scholar] [CrossRef] [Green Version]
- Heuberger, E.H.; Smits, E.; Poolman, B. Xyloside transport by XylP, a member of the galactoside-pentoside-hexuronide family. J. Biol. Chem. 2001, 276, 34465–34472. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Komiyama, A.; Sonomoto, K.; Ishizaki, A.; Hall, S.J.; Stanbury, P.F. Two different pathways for D-xylose metabolism and the effect of xylose concentration on the yield coefficient of L-lactate in mixed-acid fermentation by the lactic acid bacterium Lactococcus lactis IO-1. Appl. Microbiol. Biotechnol. 2002, 60, 160–167. [Google Scholar]
- Veeravalli, S.S.; Mathews, A.P. Continuous fermentation of xylose to short chain fatty acids by Lactobacillus buchneri under low pH conditions. Chem. Eng. J. 2018, 337, 764–771. [Google Scholar] [CrossRef]
- Kontula, P.; von Wright, A.; Mattila-Sandholm, T. Oat bran beta-gluco- and xylo-oligosaccharides as fermentative substrates for lactic acid bacteria. Int. J. Food Microbiol. 1998, 45, 163–169. [Google Scholar] [CrossRef]
- Hsu, C.K.; Liao, J.W.; Chung, Y.C.; Hsieh, C.P.; Chan, Y.C. Xylooligosaccharides and fructooligosaccharides affect the intestinal microbiota and precancerous colonic lesion development in rats. J. Nutr. 2004, 134, 1523–1528. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Yin, J.; Li, L.; Luan, C.; Zhang, J.; Zhao, C.; Li, S. Prebiotic Potential of Xylooligosaccharides Derived from Corn Cobs and Their In Vitro Antioxidant Activity When Combined with Lactobacillus. J. Microbiol. Biotechnol. 2015, 25, 1084–1092. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Li, Q.; Zhang, Y.; Zhou, X.; Yi, R.; Zhao, X. Effects of xylooligosaccharides on lipid metabolism, inflammation, and gut microbiota in C57BL/6J mice fed a high-fat diet. Front. Pharmacol. 2021, 12, 791614. [Google Scholar] [CrossRef]
- Fei, Y.; Wang, Y.; Pang, Y.; Wang, W.; Zhu, D.; Xie, M.; Lan, S.; Wang, Z. Xylooligosaccharide modulates gut microbiota and alleviates colonic inflammation caused by high fat diet induced obesity. Front. Physiol. 2019, 10, 1601. [Google Scholar] [CrossRef]
- Hintikka, J.; Lensu, S.; Makinen, E.; Karvinen, S.; Honkanen, M.; Linden, J.; Garrels, T.; Pekkala, S.; Lahti, L. Xylo-oligosaccharides in prevention of hepatic steatosis and adipose tissue inflammation: Associating taxonomic and metabolomic patterns in fecal microbiomes with biclustering. Int. J. Environ. Res. Public Health 2021, 18, 4049. [Google Scholar] [CrossRef]
- Zhang, C.; Abdulaziz Abbod Abdo, A.; Kaddour, B.; Wu, Q.; Xin, L.; Li, X.; Fan, G.; Teng, C. Xylan-oligosaccharides ameliorate high fat diet induced obesity and glucose intolerance and modulate plasma lipid profile and gut microbiota in mice. J. Funct. Foods 2020, 64, 103622. [Google Scholar] [CrossRef]
- Lim, S.M.; Kim, E.; Shin, J.H.; Seok, P.R.; Jung, S.; Yoo, S.H.; Kim, Y. Xylobiose prevents high-fat diet induced mice obesity by suppressing mesenteric fat deposition and metabolic dysregulation. Molecules 2018, 23, 705. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.P.; Prakash, S.; Bhatia, R.; Negi, M.; Singh, J.; Bishnoi, M.; Kondepudi, K.K. Generation of structurally diverse pectin oligosaccharides having prebiotic attributes. Food Hydrocolloid 2020, 108, 105988. [Google Scholar] [CrossRef]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Comput. Appl. Biosci. CABIOS 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sonderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Yu, N.Y.; Wagner, J.R.; Laird, M.R.; Melli, G.; Rey, S.; Lo, R.; Dao, P.; Sahinalp, S.C.; Ester, M.; Foster, L.J.; et al. PSORTb 3.0: Improved protein subcellular localization prediction with refined localization subcategories and predictive capabilities for all prokaryotes. Bioinformatics 2010, 26, 1608–1615. [Google Scholar] [CrossRef]
- Chen, I.A.; Chu, K.; Palaniappan, K.; Ratner, A.; Huang, J.; Huntemann, M.; Hajek, P.; Ritter, S.; Varghese, N.; Seshadri, R.; et al. The IMG/M data management and analysis system v.6.0: New tools and advanced capabilities. Nucleic Acids Res. 2021, 49, D751–D763. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [Green Version]
- Juncker, A.S.; Willenbrock, H.; von Heijne, G.; Brunak, S.; Nielsen, H.; Krogh, A. Prediction of lipoprotein signal peptides in Gram-negative bacteria. Protein Sci. A Publ. Protein Soc. 2003, 12, 1652–1662. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Km (mM) | kcat (s−1) | kcat/Km (s−1mM−1) |
|---|---|---|---|
| Bp- XylX-1 | 0.19 ± 0.02 | 11.9 ± 0.23 | 63 |
| Bp-XylX-2 | 0.19 ± 0.02 | 10.7 ± 0.2 | 56 |
| Lr-XylC | 0.23 ± 0.02 | 11.4 ± 0.2 | 49 |
| Bp-XylX-1-D371 | 0 | 0 | 0 |
| Bp-XylX-1-E408 | 0 | 0 | 0 |
| Bp-XylX-2-D386 | 0 | 0 | 0 |
| Bp-XylX-2-E409 | 0 | 0 | 0 |
| LR-XylC -D382 | 0 | 0 | 0 |
| LR-XylC -E415 | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, R.P.; Bhaiyya, R.; Thakur, R.; Niharika, J.; Singh, C.; Latousakis, D.; Saalbach, G.; Nepogodiev, S.A.; Singh, P.; Sharma, S.C.; et al. Biochemical Basis of Xylooligosaccharide Utilisation by Gut Bacteria. Int. J. Mol. Sci. 2022, 23, 2992. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23062992
Singh RP, Bhaiyya R, Thakur R, Niharika J, Singh C, Latousakis D, Saalbach G, Nepogodiev SA, Singh P, Sharma SC, et al. Biochemical Basis of Xylooligosaccharide Utilisation by Gut Bacteria. International Journal of Molecular Sciences. 2022; 23(6):2992. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23062992
Chicago/Turabian StyleSingh, Ravindra Pal, Raja Bhaiyya, Raksha Thakur, Jayashree Niharika, Chandrajeet Singh, Dimitrios Latousakis, Gerhard Saalbach, Sergey A. Nepogodiev, Praveen Singh, Sukesh Chander Sharma, and et al. 2022. "Biochemical Basis of Xylooligosaccharide Utilisation by Gut Bacteria" International Journal of Molecular Sciences 23, no. 6: 2992. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23062992