
Pial Vessel-Associated Microglia/Macrophages Increase in Female Dahl-SS/Jr Rats Independent of Pregnancy History

 ,
,
Abstract
:1. Introduction
2. Results
2.1. General Characteristics of Rats
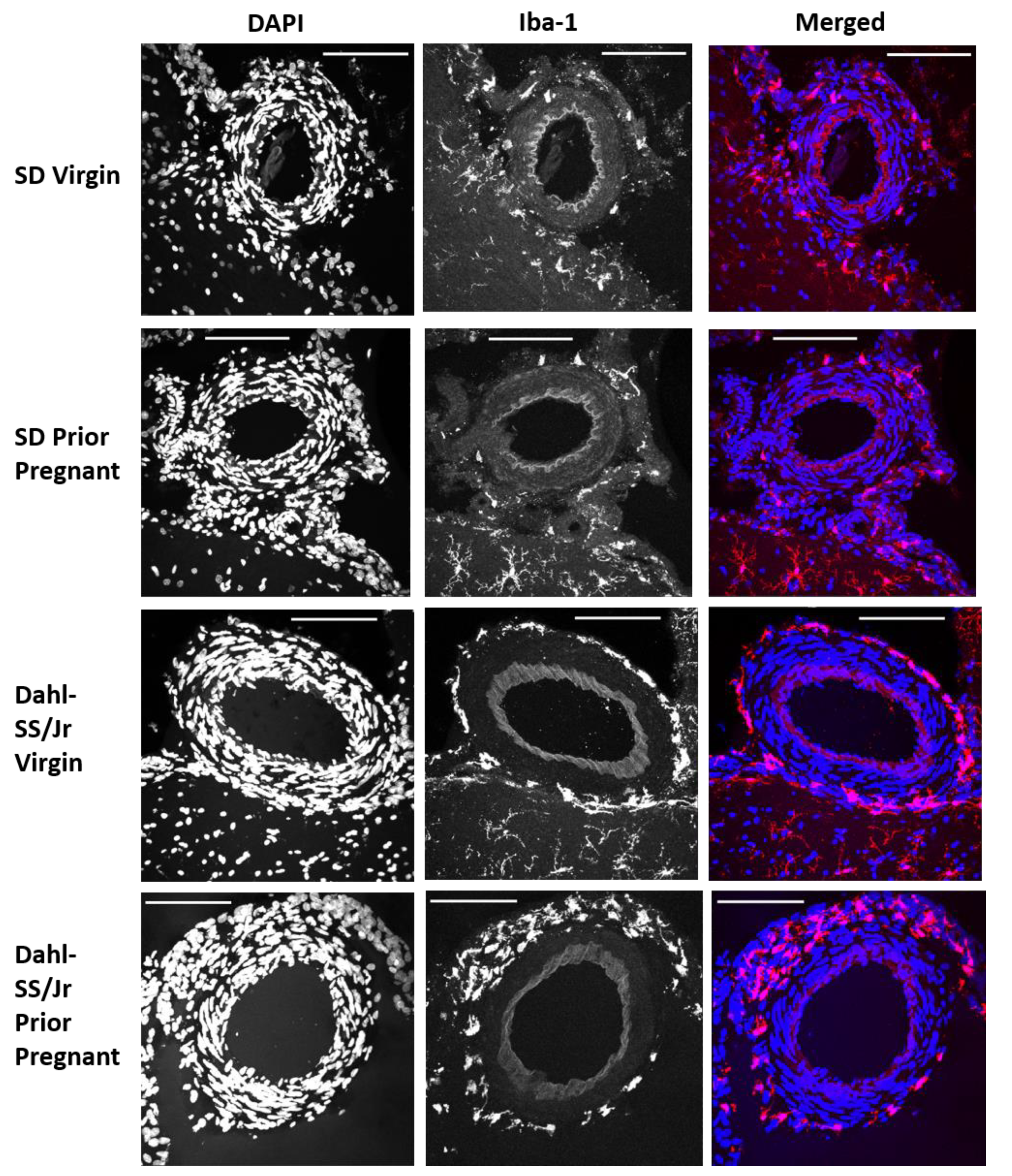
2.2. Perivascular Microglia/Macrophages Associate with Pial Vessels
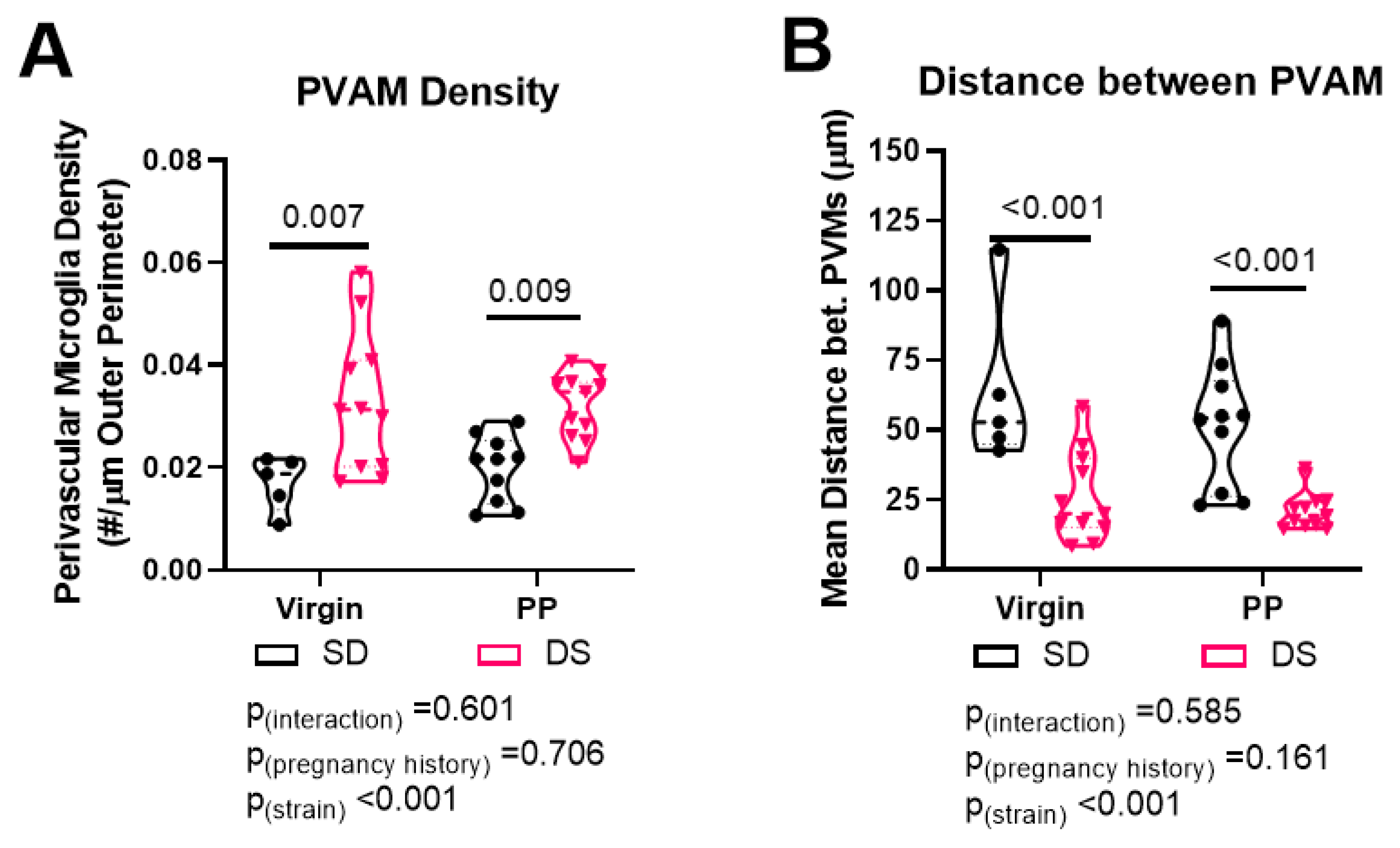
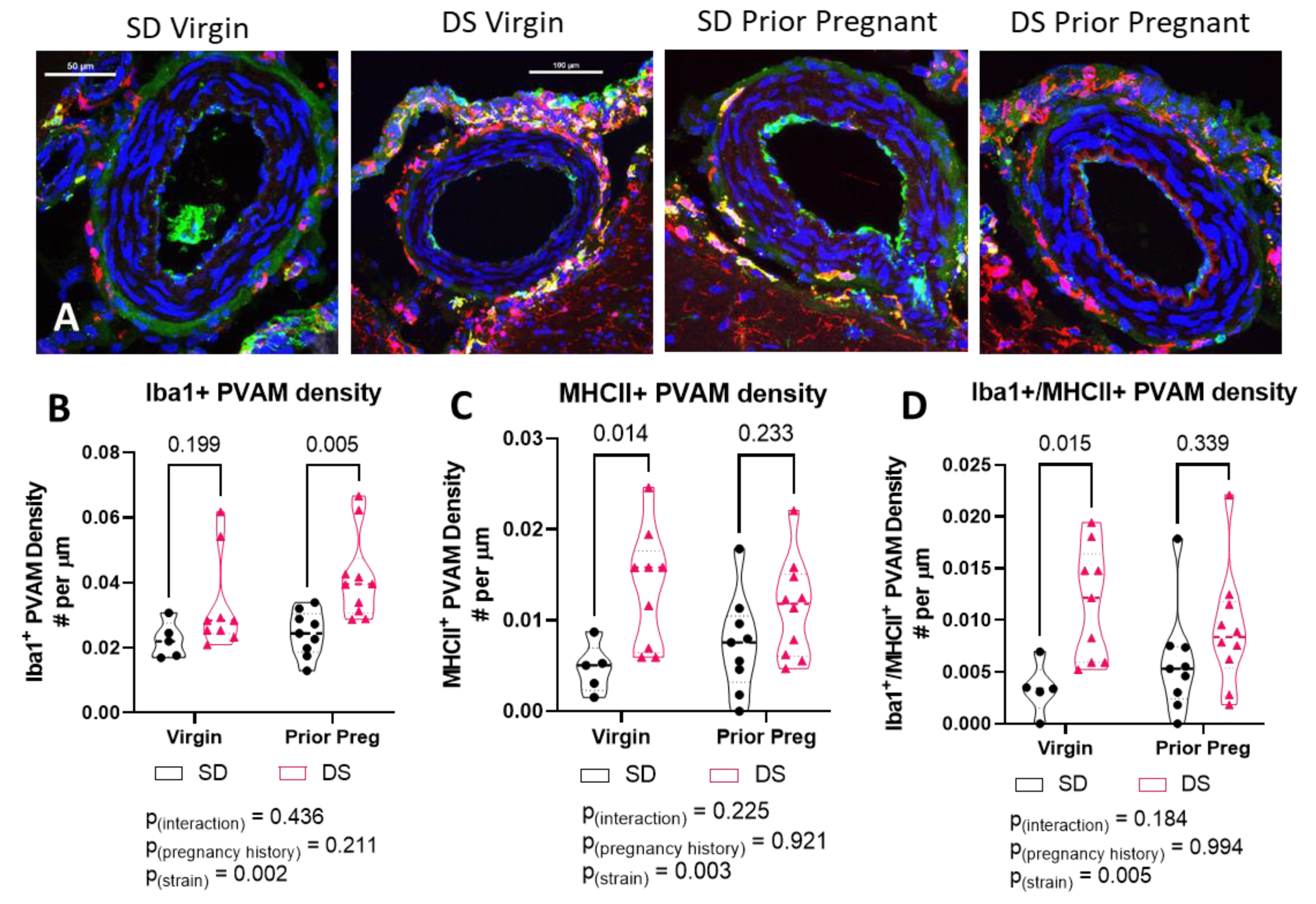
2.3. Dahl-SS/Jr Rats Have Higher Density of and Decreased Distance between PVAMs
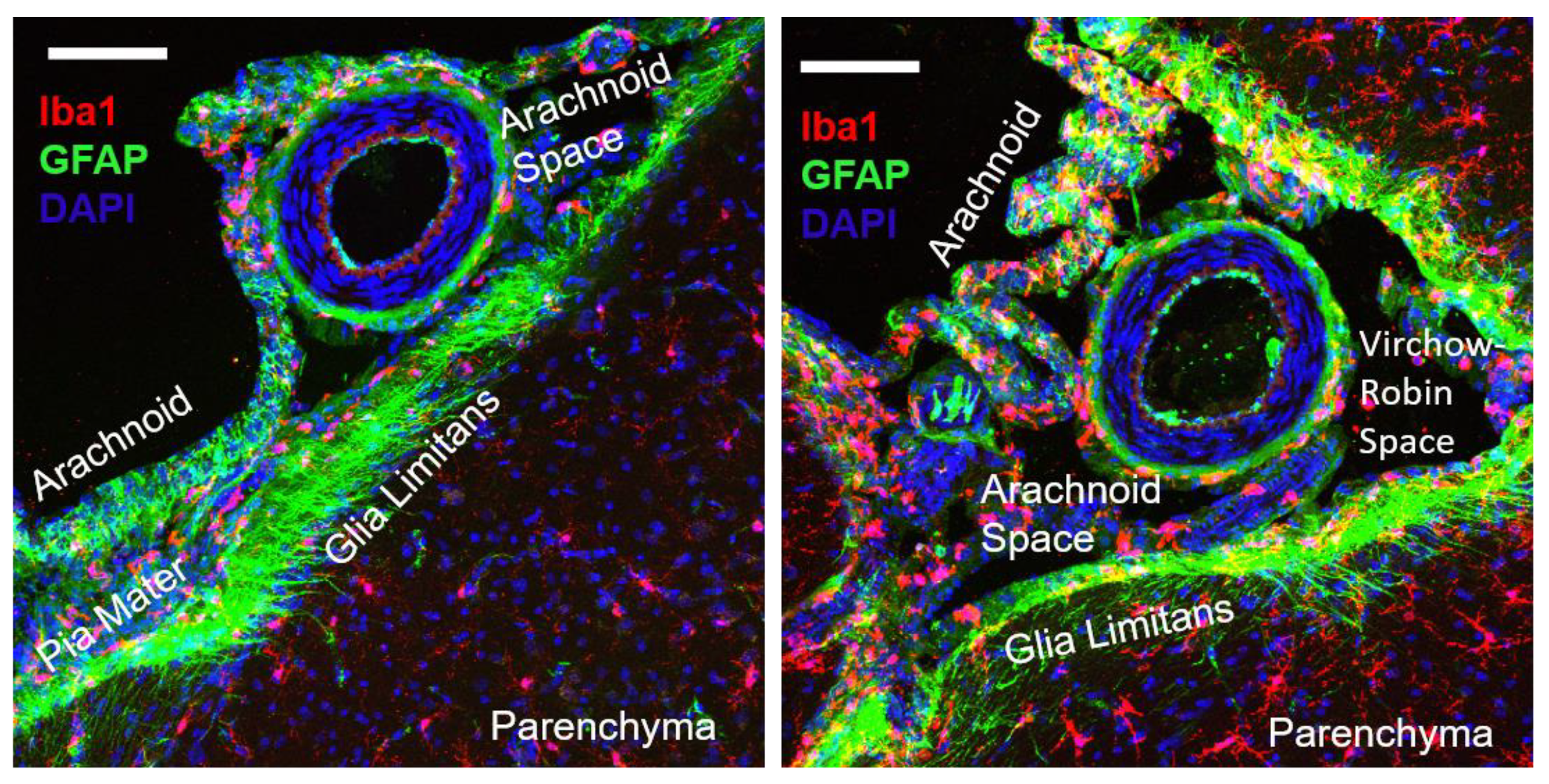
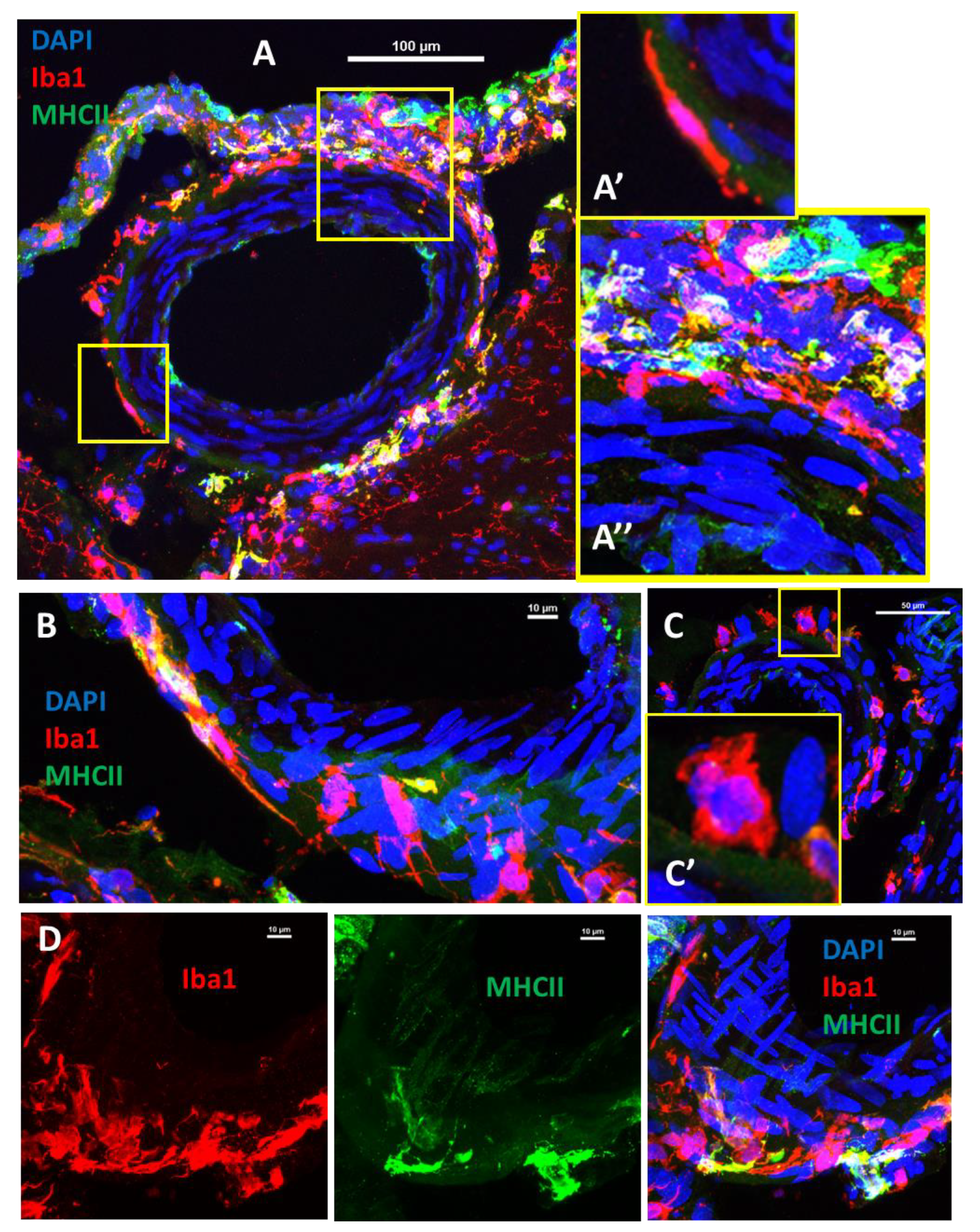
2.4. Pial Perivascular Microglia/Macrophages Are Distinct from the Glia Limitans
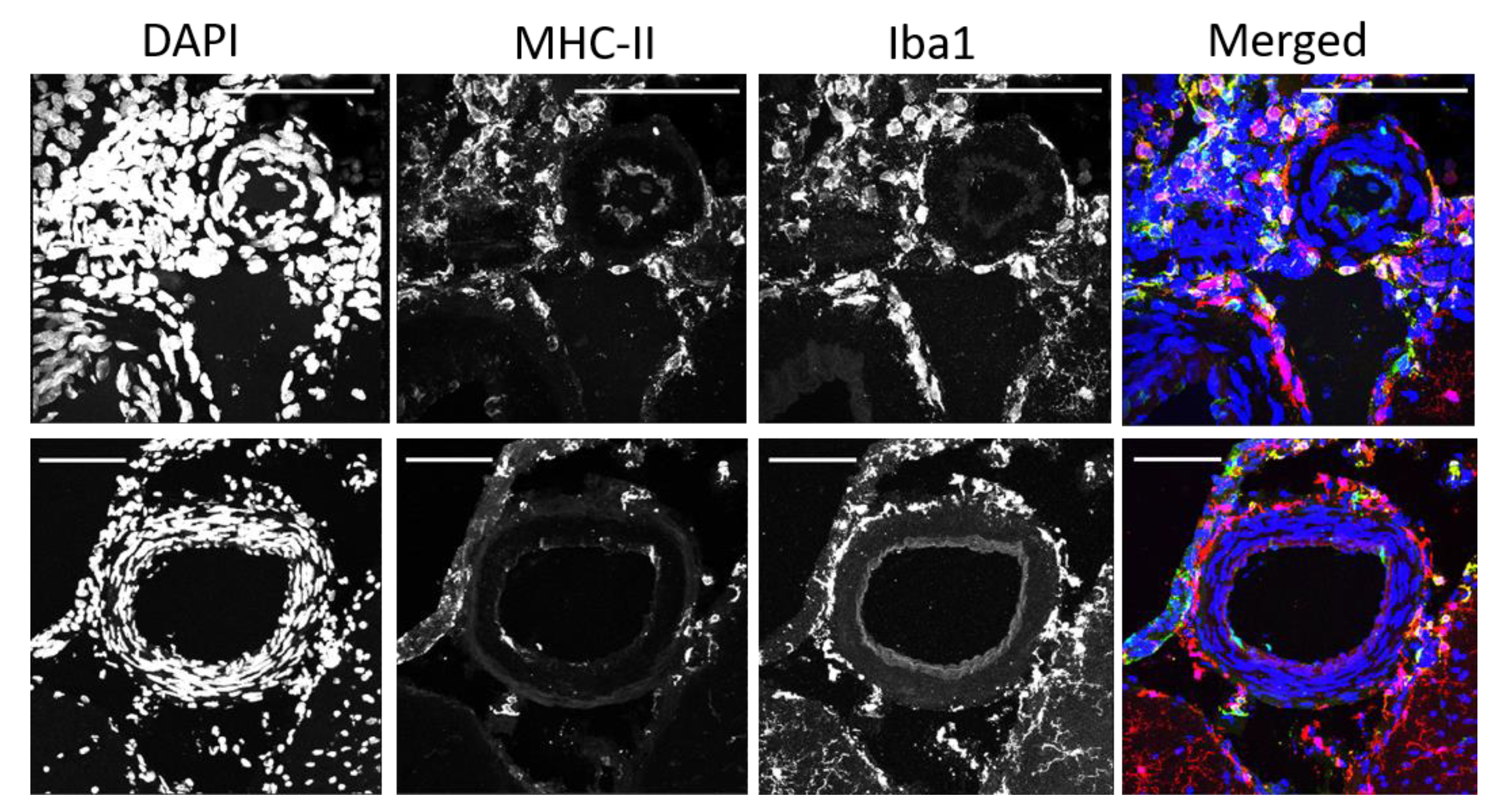
2.5. Some Perivascular Microglia/Macrophages Co-Expressed Major Histocompatibility Complex II (MHC-II)
2.6. The Density of MHCII-Positive PVAM Increased in Dahl-SS/Jr Female Rats
2.7. The Morphology of the PVAMs Differed in Different Brains
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Immunofluorescence Staining
4.3. Confocal Imaging and Analysis
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rua, R.; McGavern, D.B. Advances in Meningeal Immunity. Trends Mol. Med. 2018, 24, 542–559. [Google Scholar] [CrossRef] [PubMed]
- Heagerty, A.M.; Heerkens, E.H.; Izzard, A.S. Small artery structure and function in hypertension. J. Cell. Mol. Med. 2010, 14, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Cipolla, M.J. The adaptation of the cerebral circulation to pregnancy: Mechanisms and consequences. J. Cereb. Blood Flow Metab. 2013, 33, 465–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cipolla, M.J. The Cerebral Circulation. In Integrated Systems Physiology: From Molecule to Function; Morgan & Claypool Life Sciences Publishers: San Rafael, CA, USA, 2009. [Google Scholar]
- Koizumi, T.; Taguchi, K.; Mizuta, I.; Toba, H.; Ohigashi, M.; Onishi, O.; Ikoma, K.; Miyata, S.; Nakata, T.; Tanaka, M.; et al. Transiently proliferating perivascular microglia harbor M1 type and precede cerebrovascular changes in a chronic hypertension model. J. Neuroinflamm. 2019, 16, 79. [Google Scholar] [CrossRef]
- Hickey, W.F.; Kimura, H. Perivascular microglial cells of the CNS are bone marrow-derived and present antigen in vivo. Science 1988, 239, 290–292. [Google Scholar] [CrossRef]
- Graeber, M.B.; Streit, W.J.; Kreutzberg, G.W. Identity of ED2-positive perivascular cells in rat brain. J. Neurosci. Res. 1989, 22, 103–106. [Google Scholar] [CrossRef]
- Koizumi, T.; Kerkhofs, D.; Mizuno, T.; Steinbusch, H.W.M.; Foulquier, S. Vessel-Associated Immune Cells in Cerebrovascular Diseases: From Perivascular Macrophages to Vessel-Associated Microglia. Front. Neurosci. 2019, 13, 1291. [Google Scholar] [CrossRef] [Green Version]
- Gillis, E.E.; Williams, J.M.; Garrett, M.R.; Mooney, J.N.; Sasser, J.M. The Dahl salt-sensitive rat is a spontaneous model of superimposed preeclampsia. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 309, R62–R70. [Google Scholar] [CrossRef]
- American College of Obstetricians and Gynecologists. Hypertension in pregnancy. Report of the American College of Obstetricians and Gynecologists’ Task Force on Hypertension in Pregnancy. Obstet. Gynecol. 2013, 122, 1122–1131. [Google Scholar] [CrossRef]
- Turbeville, H.R.; Taylor, E.B.; Garrett, M.R.; Didion, S.P.; Ryan, M.J.; Sasser, J.M. Superimposed Preeclampsia Exacerbates Postpartum Renal Injury Despite Lack of Long-Term Blood Pressure Difference in the Dahl Salt-Sensitive Rat. Hypertension 2019, 73, 650–658. [Google Scholar] [CrossRef]
- Maeda, K.J.; McClung, D.M.; Showmaker, K.C.; Warrington, J.P.; Ryan, M.J.; Garrett, M.R.; Sasser, J.M. Endothelial cell disruption drives increased blood-brain barrier permeability and cerebral edema in the Dahl SS/jr rat model of superimposed preeclampsia. Am. J. Physiol. Heart Circ. Physiol. 2021, 320, H535–H548. [Google Scholar] [CrossRef] [PubMed]
- Menassa, D.A.; Gomez-Nicola, D. Microglial Dynamics During Human Brain Development. Front. Immunol. 2018, 9, 1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Atalaya, J.P.; Askew, K.E.; Sierra, A.; Gomez-Nicola, D. Development and maintenance of the brain’s immune toolkit: Microglia and non-parenchymal brain macrophages. Dev. Neurobiol. 2018, 78, 561–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Hove, H.; Martens, L.; Scheyltjens, I.; De Vlaminck, K.; Pombo Antunes, A.R.; De Prijck, S.; Vandamme, N.; De Schepper, S.; Van Isterdael, G.; Scott, C.L.; et al. A single-cell atlas of mouse brain macrophages reveals unique transcriptional identities shaped by ontogeny and tissue environment. Nat. Neurosci. 2019, 22, 1021–1035. [Google Scholar] [CrossRef]
- Kida, S.; Steart, P.V.; Zhang, E.T.; Weller, R.O. Perivascular cells act as scavengers in the cerebral perivascular spaces and remain distinct from pericytes, microglia and macrophages. Acta Neuropathol. 1993, 85, 646–652. [Google Scholar] [CrossRef]
- Ito, D.; Imai, Y.; Ohsawa, K.; Nakajima, K.; Fukuuchi, Y.; Kohsaka, S. Microglia-specific localisation of a novel calcium binding protein, Iba1. Brain Res. Mol. Brain Res. 1998, 57, 1–9. [Google Scholar] [CrossRef]
- Morita-Takemura, S.; Nakahara, K.; Hasegawa-Ishii, S.; Isonishi, A.; Tatsumi, K.; Okuda, H.; Tanaka, T.; Kitabatake, M.; Ito, T.; Wanaka, A. Responses of perivascular macrophages to circulating lipopolysaccharides in the subfornical organ with special reference to endotoxin tolerance. J. Neuroinflamm. 2019, 16, 39. [Google Scholar] [CrossRef]
- Faraco, G.; Sugiyama, Y.; Lane, D.; Garcia-Bonilla, L.; Chang, H.; Santisteban, M.M.; Racchumi, G.; Murphy, M.; Van Rooijen, N.; Anrather, J.; et al. Perivascular macrophages mediate the neurovascular and cognitive dysfunction associated with hypertension. J. Clin. Investig. 2016, 126, 4674–4689. [Google Scholar] [CrossRef] [Green Version]
- Park, L.; Uekawa, K.; Garcia-Bonilla, L.; Koizumi, K.; Murphy, M.; Pistik, R.; Younkin, L.; Younkin, S.; Zhou, P.; Carlson, G.; et al. Brain Perivascular Macrophages Initiate the Neurovascular Dysfunction of Alzheimer Abeta Peptides. Circ. Res. 2017, 121, 258–269. [Google Scholar] [CrossRef]
- Pires, P.W.; Girgla, S.S.; McClain, J.L.; Kaminski, N.E.; van Rooijen, N.; Dorrance, A.M. Improvement in middle cerebral artery structure and endothelial function in stroke-prone spontaneously hypertensive rats after macrophage depletion. Microcirculation 2013, 20, 650–661. [Google Scholar] [CrossRef] [Green Version]
- Farhadian, S.F.; Mehta, S.S.; Zografou, C.; Robertson, K.; Price, R.W.; Pappalardo, J.; Chiarella, J.; Hafler, D.A.; Spudich, S.S. Single-cell RNA sequencing reveals microglia-like cells in cerebrospinal fluid during virologically suppressed HIV. JCI Insight 2018, 3, e121718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedragosa, J.; Salas-Perdomo, A.; Gallizioli, M.; Cugota, R.; Miro-Mur, F.; Brianso, F.; Justicia, C.; Perez-Asensio, F.; Marquez-Kisinousky, L.; Urra, X.; et al. CNS-border associated macrophages respond to acute ischemic stroke attracting granulocytes and promoting vascular leakage. Acta Neuropathol. Commun. 2018, 6, 76. [Google Scholar] [CrossRef] [PubMed]
- Kerkhofs, D.; van Hagen, B.T.; Milanova, I.V.; Schell, K.J.; van Essen, H.; Wijnands, E.; Goossens, P.; Blankesteijn, W.M.; Unger, T.; Prickaerts, J.; et al. Pharmacological depletion of microglia and perivascular macrophages prevents Vascular Cognitive Impairment in Ang II-induced hypertension. Theranostics 2020, 10, 9512–9527. [Google Scholar] [CrossRef] [PubMed]
- Nimmerjahn, A.; Kirchhoff, F.; Helmchen, F. Resting microglial cells are highly dynamic surveillants of brain parenchyma in vivo. Science 2005, 308, 1314–1318. [Google Scholar] [CrossRef] [Green Version]
- Bernier, L.P.; Bohlen, C.J.; York, E.M.; Choi, H.B.; Kamyabi, A.; Dissing-Olesen, L.; Hefendehl, J.K.; Collins, H.Y.; Stevens, B.; Barres, B.A.; et al. Nanoscale Surveillance of the Brain by Microglia via cAMP-Regulated Filopodia. Cell Rep. 2019, 27, 2895–2908.e4. [Google Scholar] [CrossRef] [Green Version]
- Zhang, E.T.; Richards, H.K.; Kida, S.; Weller, R.O. Directional and compartmentalised drainage of interstitial fluid and cerebrospinal fluid from the rat brain. Acta Neuropathol. 1992, 83, 233–239. [Google Scholar] [CrossRef]
- Serrats, J.; Schiltz, J.C.; García-Bueno, B.; van Rooijen, N.; Reyes, T.M.; Sawchenko, P.E. Dual roles for perivascular macrophages in immune-to-brain signaling. Neuron 2010, 65, 94–106. [Google Scholar] [CrossRef] [Green Version]
- Mato, M.; Ookawara, S.; Mato, T.K.; Namiki, T. An attempt to differentiate further between microglia and fluorescent granular perithelial (FGP) cells by their capacity to incorporate exogenous protein. Am. J. Anat. 1985, 172, 125–140. [Google Scholar] [CrossRef]
- Bechmann, I.; Nitsch, R. Identification of phagocytic glial cells after lesion-induced anterograde degeneration using double-fluorescence labeling: Combination of axonal tracing and lectin or immunostaining. Histochem. Cell Biol. 1997, 107, 391–397. [Google Scholar] [CrossRef]
- Bechmann, I.; Nitsch, R. Astrocytes and microglial cells incorporate degenerating fibers following entorhinal lesion: A light, confocal, and electron microscopical study using a phagocytosis-dependent labeling technique. Glia 1997, 20, 145–154. [Google Scholar] [CrossRef]
- Aukes, A.M.; Vitullo, L.; Zeeman, G.G.; Cipolla, M.J. Pregnancy prevents hypertensive remodeling and decreases myogenic reactivity in posterior cerebral arteries from Dahl salt-sensitive rats: A role in eclampsia? Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H1071–H1076. [Google Scholar] [CrossRef]
- Kettenmann, H.; Hanisch, U.K.; Noda, M.; Verkhratsky, A. Physiology of microglia. Physiol. Rev. 2011, 91, 461–553. [Google Scholar] [CrossRef] [PubMed]
- Bechmann, I.; Priller, J.; Kovac, A.; Bontert, M.; Wehner, T.; Klett, F.F.; Bohsung, J.; Stuschke, M.; Dirnagl, U.; Nitsch, R. Immune surveillance of mouse brain perivascular spaces by blood-borne macrophages. Eur. J. Neurosci. 2001, 14, 1651–1658. [Google Scholar] [CrossRef] [PubMed]
- Albini, T.A.; Wang, R.C.; Reiser, B.; Zamir, E.; Wu, G.S.; Rao, N.A. Microglial stability and repopulation in the retina. Br. J. Ophthalmol. 2005, 89, 901–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Askew, K.; Li, K.; Olmos-Alonso, A.; Garcia-Moreno, F.; Liang, Y.; Richardson, P.; Tipton, T.; Chapman, M.A.; Riecken, K.; Beccari, S.; et al. Coupled Proliferation and Apoptosis Maintain the Rapid Turnover of Microglia in the Adult Brain. Cell Rep. 2017, 18, 391–405. [Google Scholar] [CrossRef] [Green Version]
- Hung, S.K.; Lee, M.S.; Lin, H.Y.; Chen, L.C.; Chuang, C.J.; Chew, C.H.; Yu, B.H.; Yang, H.J.; Hsu, F.C.; Chiou, W.Y. Impact of Hypertensive Disorders of Pregnancy on the Risk of Stroke Stratified by Subtypes and Follow-Up Time. Stroke 2022, 53, 338–344. [Google Scholar] [CrossRef]
- Basit, S.; Wohlfahrt, J.; Boyd, H.A. Pre-eclampsia and risk of dementia later in life: Nationwide cohort study. BMJ 2018, 363, k4109. [Google Scholar] [CrossRef] [Green Version]
- Clayton, A.M.; Shao, Q.; Paauw, N.D.; Giambrone, A.B.; Granger, J.P.; Warrington, J.P. Postpartum increases in cerebral edema and inflammation in response to placental ischemia during pregnancy. Brain Behav. Immun. 2018, 70, 376–389. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measurement | SD Virgin | SD Prior Pregnant | Dahl-SS Virgin | Dahl-SS Prior Pregnant |
|---|---|---|---|---|
| Body Weight (g) | 291 ± 10 (5) | 294 ± 6 (5) | 254 ± 12 (3) | 307 ± 39 (3) |
| Hematocrit (%) | Not assessed | 44 ± 3 (5) | 49 ± 5 (9) | 48 ± 3 (9) |
| Mean Arterial Pressure (mmHg) | Not assessed | 144 ± 22 (4) | 157 ± 14 (8) | 156 ± 11 (7) |
| Pial Vessel Parameters | ||||
| Outer Perimeter (µm) | 587± 107 (5) | 527 ± 109 (10) | 622 ± 122 (11) | 769 ± 253 (11) ‡ |
| Wall Thickness (µm) | 49 ± 15 (5) | 43 ± 6 (10) | 45 ± 6 (11) | 48 ± 5 (11) |
| No. of PVAM | 10 ± 3 (5) | 11 ± 5 (10) | 19 ± 9 (11) ¥ | 24 ± 6 (11) ‡ |
| Cell Type | Markers Used | Location | Method | Main Findings | Ref |
|---|---|---|---|---|---|
| Perivascular Cells | ED2 | Parenchyma | Rotring Indian Ink + Light Microscopy + IHC + EM | ED2+ cells in perivascular spaces took up the ink. No uptake by pericytes, macrophages, or microglia. Perivascular cells act as scavengers | [16] |
| Perivascular Macrophages | Iba1low | Subfornical Organ | IF, LPS injection | Iba1+ perivascular macrophages expressed IL-1β. May contribute to tolerance of endotoxin | [18] |
| Perivascular Macrophages | CD206 Lyve1 Iba1low | Parenchyma | IF, AngII infusion | Increased production of ROS, BBB leakage, cognitive impairments. Depletion of PVMs prevented these | [19] |
| Perivascular Macrophages | CD206 CD36 | Parenchyma | IF | PVMs express Aβ receptor, CD36, and may contribute to the pathophysiology of Alzheimer’s disease | [20] |
| Perivascular Macrophages | CD163 | MCA | IF | Depletion using Clodronate reduced perivascular macrophages in spontaneously hypertensive rats | [21] |
| Myeloid-2 Cells | TREM2, APOE | CSF | ScRNA-Seq | ↑ Transcripts of myeloid cells in CSF from HIV+ participants compared to control and blood | [22] |
| Border-Associated Macrophages | CD163 | Leptomenigeal spaces and Choroid plexus | scRNA-Seq Ischemia/reperfusion | Granulocyte recruitment, ↑ VEGF expression, ↑ pial and cortical vessel permeability, ↑ neurological dysfunction post-ischemia/reperfusion acutely | [23] |
| Border-Associated Macrophages | N/A | Dura mater, subdural meninges, choroid plexus | scRNA-Seq | Distinct tissue-specific transcriptional profiles. Unique microglial subset on apical surface of choroid plexus epithelium | [15] |
| Perivascular Macrophages/ Microglia | Cx3Cr1 | Parenchyma | PLX5622, AngII | Depletion prevents AngII-induced short-term memory impairment | [24] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Warrington, J.P.; Shao, Q.; Clayton, A.M.; Maeda, K.J.; Beckett, A.G.; Garrett, M.R.; Sasser, J.M. Pial Vessel-Associated Microglia/Macrophages Increase in Female Dahl-SS/Jr Rats Independent of Pregnancy History. Int. J. Mol. Sci. 2022, 23, 3384. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23063384
Warrington JP, Shao Q, Clayton AM, Maeda KJ, Beckett AG, Garrett MR, Sasser JM. Pial Vessel-Associated Microglia/Macrophages Increase in Female Dahl-SS/Jr Rats Independent of Pregnancy History. International Journal of Molecular Sciences. 2022; 23(6):3384. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23063384
Chicago/Turabian StyleWarrington, Junie P., Qingmei Shao, Ahsia M. Clayton, Kenji J. Maeda, Ashtin G. Beckett, Michael R. Garrett, and Jennifer M. Sasser. 2022. "Pial Vessel-Associated Microglia/Macrophages Increase in Female Dahl-SS/Jr Rats Independent of Pregnancy History" International Journal of Molecular Sciences 23, no. 6: 3384. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23063384