
How miRNAs Regulate Schwann Cells during Peripheral Nerve Regeneration—A Systemic Review

 , , , , and
, , , , and
Abstract
:1. Introduction
1.1. Wallerian Degeneration
1.2. miRNA
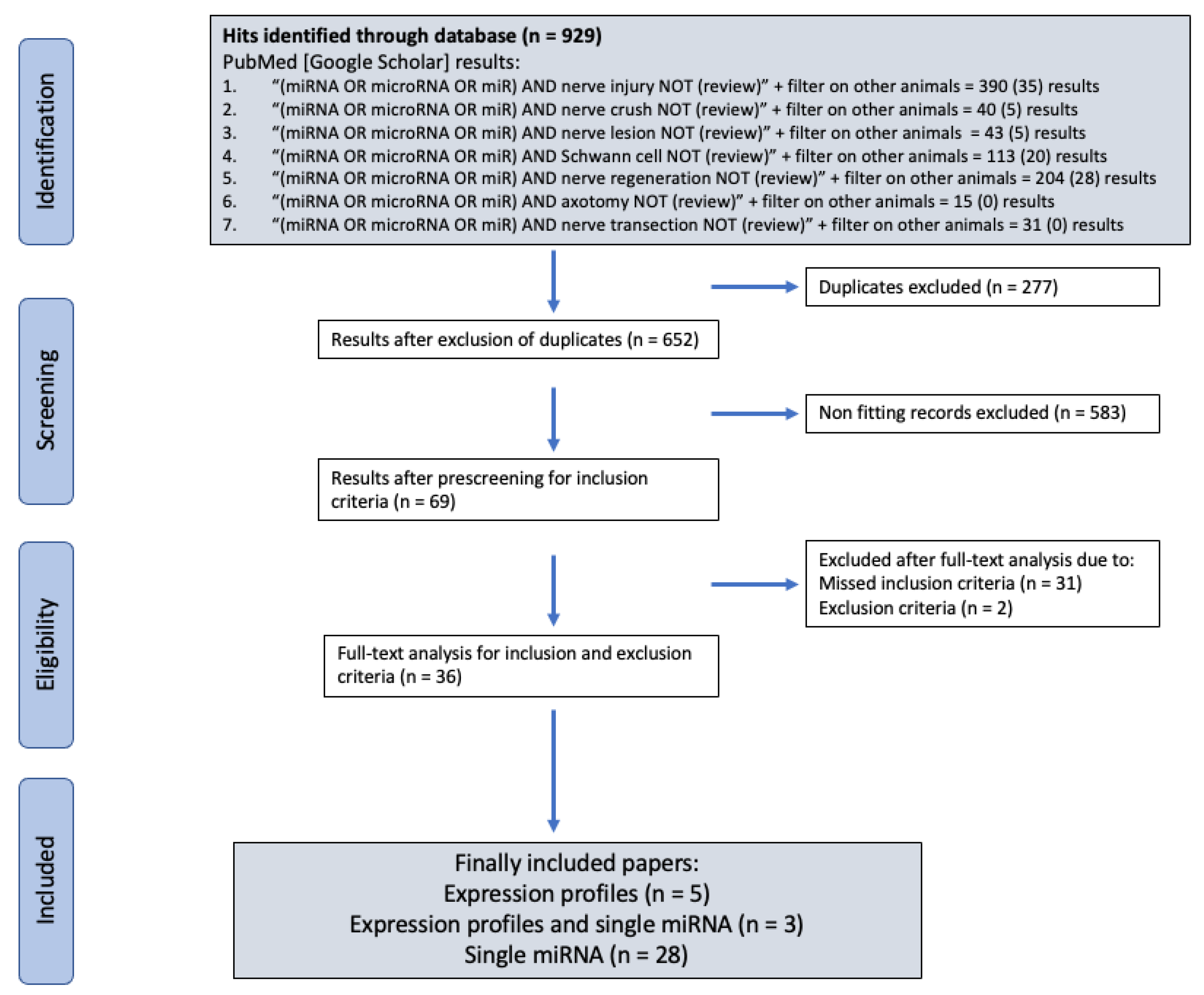
2. Materials and Methods
2.1. Inclusion Criteria
- (a)
- articles investigating a single miRNA, its targets and function in SC cultures;
- (b)
- publications exploring overall expression of miRNAs in acute/traumatic peripheral nerve lesion models (crush or transection) with at least with a microarray or full sequencing approach.
2.2. Exclusion Criteria
2.3. Search Algorithm Results
2.4. Data Extraction
3. Results
3.1. Expression Profiles of miRNA upon Peripheral Nerve Injuries
3.2. Single miRNAs Influence Certain Nerve Regeneration Incrementally
3.2.1. Inflammation
miR-182
miR-340
3.2.2. Proliferation
miR-1
miR-21 and miR-124
miR-221/222
miR-192-5p
miR-3099
miR-146b
miR-210
3.2.3. Migration
miR-9
miR-138-5p
miR-129
miR-34a, miR-132, and miR-3075
3.2.4. Myelination
miR-34a and let-7 family
miR-140
miR-29a and miR-29b
miR-485-5p
miR-30c
4. Discussion
4.1. Expression Profiles and Function
4.2. Controversial miRNAs
4.3. miRNAs in Therapeutic Approaches
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DRG | dorsal root ganglion |
| EV | extracellular vesicles |
| MSC | mesenchymal stem cell |
| SC | Schwann cell |
References
- Lowe, J.B.; Sen, S.K.; Mackinnon, S.E. Current Approach to Radial Nerve Paralysis. Plast. Reconstr. Surg. 2002, 110, 1099–1112. [Google Scholar] [CrossRef] [PubMed]
- Rosberg, H.E.; Carlsson, K.S.; Höjgård, S.; Lindgren, B.; Lundborg, G.; Dahlin, L.B. Injury to the human median and ulnar nerves in the forearm—Analysis of costs for treatment and rehabilitation of 69 patients in southern sweden. J. Hand Surg. 2005, 30, 35–39. [Google Scholar] [CrossRef]
- Seddon, R.J. Surgical experiences with peripheral nerve injuries. Q. Bull. Northwestern Univ. (Evanst. Ill.) Med. Sch. 1947, 21, 201–210. [Google Scholar]
- Sunderland, S. A classification of peripheral nerve injuries producing loss of function. Brain 1951, 74, 491–516. [Google Scholar] [CrossRef] [PubMed]
- Mackinnon, S.E. Surgical Approach to the Radial Nerve. Tech. Hand Up. Extrem. Surg. 1999, 3, 87–98. [Google Scholar] [CrossRef]
- Waller, A. Experiments on the Section of the Glosso-Pharyngeal and Hypoglossal Nerves of the Frog, and Observations of the Alterations Produced Thereby in the Structure of Their Primitive Fibres. Edinb. Med. Surg. J. 1851, 76, 369–376. [Google Scholar]
- Conforti, L.; Gilley, J.; Coleman, M.P. Wallerian degeneration: An emerging axon death pathway linking injury and disease. Nat. Rev. Neurosci. 2014, 15, 394–409. [Google Scholar] [CrossRef]
- Stoll, G.; Griffin, J.W.; Li, C.Y.; Trapp, B.D. Wallerian degeneration in the peripheral nervous system: Participation of both Schwann cells and macrophages in myelin degradation. J. Neurocytol. 1989, 18, 671–683. [Google Scholar] [CrossRef]
- Gaudet, A.D.; Popovich, P.G.; Ramer, M.S. Wallerian degeneration: Gaining perspective on inflammatory events after peripheral nerve injury. J. Neuroinflamm. 2011, 8, 110. [Google Scholar] [CrossRef] [Green Version]
- Terzis, J.K.; Konofaos, P. Radial Nerve Injuries and Outcomes: Our Experience. Plast. Reconstr. Surg. 2011, 127, 739–751. [Google Scholar] [CrossRef]
- Pan, C.H.; Chuang, D.C.C.; Rodríguez-Lorenzo, A. Outcomes of Nerve Reconstruction for Radial Nerve Injuries Based on the Level of Injury in 244 Operative Cases. J. Hand Surg. 2010, 35, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Ruijs, A.C.J.; Jaquet, J.-B.; Kalmijn, S.; Giele, H.; Hovius, S.E.R. Median and Ulnar Nerve Injuries: A Meta-Analysis of Predictors of Motor and Sensory Recovery after Modern Microsurgical Nerve Repair. Plast. Reconstr. Surg. 2005, 116, 484–494. [Google Scholar] [CrossRef] [Green Version]
- Best, T.J.; Mackinnon, S.E.; Evans, P.J.; Hunter, D.; Midha, R. Peripheral Nerve Revascularization: Histomorphometric Study of Small-and Large-Caliber Grafts. J. Reconstr. Microsurg. 1999, 15, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Xiao, G.; Zhang, K.; Ying, G.; Xu, H.; De Melo, B.A.G.; Li, S.; Liu, F.; Yetisen, A.K.; Jiang, N. Tacrolimus- and Nerve Growth Factor-Treated Allografts for Neural Tissue Regeneration. ACS Chem. Neurosci. 2019, 10, 1411–1419. [Google Scholar] [CrossRef]
- Masgutov, R.; Masgutova, G.; Mukhametova, L.; Garanina, E.; Arkhipova, S.S.; Zakirova, E.; Mukhamedshina, Y.O.; Margarita, Z.; Gilazieva, Z.; Syromiatnikova, V.; et al. Allogenic Adipose Derived Stem Cells Transplantation Improved Sciatic Nerve Regeneration in Rats: Autologous Nerve Graft Model. Front. Pharmacol. 2018, 9, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kornfeld, T.; Vogt, P.M.; Bucan, V.; Peck, C.-T.; Reimers, K.; Radtke, C. Characterization and Schwann Cell Seeding of up to 15.0 cm Long Spider Silk Nerve Conduits for Reconstruction of Peripheral Nerve Defects. J. Funct. Biomater. 2016, 7, 30. [Google Scholar] [CrossRef] [Green Version]
- Xia, B.; Gao, J.; Li, S.; Huang, L.; Ma, T.; Zhao, L.; Yang, Y.; Huang, J.; Luo, Z. Extracellular Vesicles Derived From Olfactory Ensheathing Cells Promote Peripheral Nerve Regeneration in Rats. Front. Cell. Neurosci. 2019, 13, 548. [Google Scholar] [CrossRef] [Green Version]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved Seed Pairing, Often Flanked by Adenosines, Indicates that Thousands of Human Genes are MicroRNA Targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Jinek, M.; Doudna, J.A. A three-dimensional view of the molecular machinery of RNA interference. Nature 2009, 457, 405–412. [Google Scholar] [CrossRef]
- Siomi, H.; Siomi, M.C. On the road to reading the RNA-interference code. Nature 2009, 457, 396–404. [Google Scholar] [CrossRef]
- Zeng, Y. Structural requirements for pre-microRNA binding and nuclear export by Exportin 5. Nucleic Acids Res. 2004, 32, 4776–4785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebert, L.F.R.; Macrae, I.J. Regulation of microRNA function in animals. Nat. Rev. Mol. Cell Biol. 2019, 20, 21–37. [Google Scholar] [CrossRef]
- Pong, S.K.; Gullerova, M. Noncanonical functions of micro RNA pathway enzymes—Drosha, DGCR 8, Dicer and Ago proteins. FEBS Lett. 2018, 592, 2973–2986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filipowicz, W.; Bhattacharyya, S.N.; Sonenberg, N. Mechanisms of post-transcriptional regulation by microRNAs: Are the answers in sight? Nat. Rev. Genet. 2008, 9, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Sohn, E.J.; Park, H.T. MicroRNA Mediated Regulation of Schwann Cell Migration and Proliferation in Peripheral Nerve Injury. BioMed Res. Int. 2018, 2018, 8198365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, P.T.; Kiriakidou, M.; Mourelatos, Z.; Tan, G.S.; Jennings, M.H.; Xie, K.; Wang, W.-X. High-throughput experimental studies to identify miRNA targets directly, with special focus on the mammalian brain. Brain Res. 2010, 1338, 122–130. [Google Scholar] [CrossRef] [Green Version]
- Ambros, V.; Bartel, B.; Bartel, D.P.; Burge, C.B.; Carrington, J.C.; Chen, X.; Dreyfuss, G.; Eddy, S.R.; Griffiths-Jones, S.; Marshall, M.; et al. A uniform system for microRNA annotation. RNA 2003, 9, 277–279. [Google Scholar] [CrossRef] [Green Version]
- Griffiths-Jones, S.; Grocock, R.J.; Van Dongen, S.; Bateman, A.; Enright, A.J. miRBase: microRNA sequences, targets and gene nomenclature. Nucleic Acids Res. 2006, 34, D140–D144. [Google Scholar] [CrossRef]
- Mishra, S.; Yadav, T.; Rani, V. Exploring miRNA based approaches in cancer diagnostics and therapeutics. Crit. Rev. Oncol. 2015, 98, 12–23. [Google Scholar] [CrossRef]
- Mycko, M.P.; E Baranzini, S. microRNA and exosome profiling in multiple sclerosis. Mult. Scler. J. 2020, 26, 599–604. [Google Scholar] [CrossRef]
- Darden, D.B.; Stortz, J.A.; Hollen, M.K.; Cox, M.C.; Apple, C.G.; Hawkins, R.B.; Rincon, J.C.; Lopez, M.-C.; Wang, Z.; Navarro, E.; et al. Identification of Unique mRNA and miRNA Expression Patterns in Bone Marrow Hematopoietic Stem and Progenitor Cells After Trauma in Older Adults. Front. Immunol. 2020, 11, 1289. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, D.E.; Duque, E.; Hadjiargyrou, M. MicroRNAs and fracture healing: Pre-clinical studies. Bone 2021, 143, 115758. [Google Scholar] [CrossRef]
- Zhou, S.; Shen, D.; Wang, Y.; Gong, L.; Tang, X.; Yu, B.; Gu, X.; Ding, F. microRNA-222 Targeting PTEN Promotes Neurite Outgrowth from Adult Dorsal Root Ganglion Neurons following Sciatic Nerve Transection. PLoS ONE 2012, 7, e44768. [Google Scholar] [CrossRef]
- Arthur-Farraj, P.J.; Morgan, C.C.; Adamowicz, M.; Gomez-Sanchez, J.A.; Fazal, S.V.; Beucher, A.; Razzaghi, B.; Mirsky, R.; Jessen, K.R.; Aitman, T.J. Changes in the Coding and Non-coding Transcriptome and DNA Methylome that Define the Schwann Cell Repair Phenotype after Nerve Injury. Cell Rep. 2017, 20, 2719–2734. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.-L.; Wang, H.-C.; Chunag, Y.-T.; Chou, C.-W.; Lin, I.-L.; Lai, C.-S.; Chang, L.-L.; Cheng, K.-I. miRNA Expression Change in Dorsal Root Ganglia After Peripheral Nerve Injury. J. Mol. Neurosci. 2016, 61, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. PLoS Med. 2021, 18, e1003583. [Google Scholar] [CrossRef]
- Li, S.; Zhang, R.; Yuan, Y.; Yi, S.; Chen, Q.; Gong, L.; Liu, J.; Ding, F.; Cao, Z.; Gu, X. MiR-340 Regulates Fibrinolysis and Axon Regrowth Following Sciatic Nerve Injury. Mol. Neurobiol. 2017, 54, 4379–4389. [Google Scholar] [CrossRef]
- Yu, B.; Qian, T.; Wang, Y.; Zhou, S.; Ding, G.; Ding, F.; Gu, X. miR-182 inhibits Schwann cell proliferation and migration by targeting FGF9 and NTM, respectively at an early stage following sciatic nerve injury. Nucleic Acids Res. 2012, 40, 10356–10365. [Google Scholar] [CrossRef] [Green Version]
- Yi, S.; Yuan, Y.; Chen, Q.; Wang, X.; Gong, L.; Liu, J.; Gu, X.; Li, S. Regulation of Schwann cell proliferation and migration by miR-1 targeting brain-derived neurotrophic factor after peripheral nerve injury. Sci. Rep. 2016, 6, 29121. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-P.; Xu, P.; Guo, C.-X.; Luo, Z.-R.; Zhu, J.; Mou, F.-F.; Cai, H.; Wang, C.; Ye, X.-C.; Shao, S.-J.; et al. miR-1b overexpression suppressed proliferation and migration of RSC96 and increased cell apoptosis. Neurosci. Lett. 2018, 687, 137–145. [Google Scholar] [CrossRef]
- Li, X.; Yuan, L.; Wang, J.; Zhang, Z.; Fu, S.; Wang, S.; Li, X. MiR-1b up-regulation inhibits rat neuron proliferation and regeneration yet promotes apoptosis via targeting KLF7. Folia Neuropathol. 2021, 59, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Nagata, K.; Hama, I.; Kiryu-Seo, S.; Kiyama, H. microRNA-124 is down regulated in nerve-injured motor neurons and it potentially targets mRNAs for KLF6 and STAT3. Neuroscience 2014, 256, 426–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ning, X.-J.; Lu, X.-H.; Luo, J.-C.; Chen, C.; Gao, Q.; Li, Z.-Y.; Wang, H. Molecular mechanism of microRNA-21 promoting Schwann cell proliferation and axon regeneration during injured nerve repair. RNA Biol. 2020, 17, 1508–1519. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Leal, R.; Díaz-Viraqué, F.; Catalan, R.J.; Saquel, C.; Enright, A.; Iraola, G.; Court, F.A. Schwann cell reprogramming into repair cells increases exosome-loaded miRNA-21 promoting axonal growth. J. Cell Sci. 2020, 133, jcs239004. [Google Scholar] [CrossRef]
- Yu, B.; Zhou, S.; Wang, Y.; Qian, T.; Ding, G.; Ding, F.; Gu, X. miR-221/222 promote Schwann cell proliferation and migration by targeting LASS2 following sciatic nerve injury. J. Cell Sci. 2012, 125, 2675–2683. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Yuan, Y.; Li, P.; Pan, J.; Qin, J.; Liu, Y.; Zhang, Y.; Tian, F.; Yu, B.; Zhou, S. miR-221-3p Inhibits Schwann Cell Myelination. Neuroscience 2018, 379, 239–245. [Google Scholar] [CrossRef]
- Liu, X.; Cui, X.; Guan, G.; Dong, Y.; Zhang, Z. microRNA-192-5p is involved in nerve repair in rats with peripheral nerve injury by regulating XIAP. Cell Cycle 2020, 19, 326–338. [Google Scholar] [CrossRef]
- Qian, T.-M.; Liu, Q.-Y.; Miao, Y.; Wang, X.-H.; Wang, P.; Cheng, Z.-C. Increased levels of miR-3099 induced by peripheral nerve injury promote Schwann cell proliferation and migration. Neural Regen. Res. 2019, 14, 525–531. [Google Scholar] [CrossRef]
- Li, W.-Y.; Zhang, W.-T.; Cheng, Y.-X.; Liu, Y.-C.; Zhai, F.-G.; Sun, P.; Li, H.-T.; Deng, L.-X.; Zhu, X.-F.; Wang, Y. Inhibition of KLF7-Targeting MicroRNA 146b Promotes Sciatic Nerve Regeneration. Neurosci. Bull. 2018, 34, 419–437. [Google Scholar] [CrossRef]
- Zhang, X.; Gong, X.; Qiu, J.; Zhang, Y.; Gong, F. MicroRNA-210 contributes to peripheral nerve regeneration through promoting the proliferation and migration of Schwann cells. Exp. Ther. Med. 2017, 14, 2809–2816. [Google Scholar] [CrossRef]
- Zhou, S.; Gao, R.; Hu, W.; Qian, T.; Wang, N.; Ding, G.; Ding, F.; Yu, B.; Gu, X. miR-9 inhibits Schwann cell migration by targeting CTHRC1 following sciatic nerve injury. J. Cell Sci. 2014, 127, 967–976. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Chen, Q.; Yi, S.; Liu, Q.; Zhang, R.; Wang, P.; Qian, T.; Li, S. The microRNAs let-7 and miR-9 down-regulate the axon-guidance genes Ntn1 and Dcc during peripheral nerve regeneration. J. Biol. Chem. 2019, 294, 3489–3500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rieger-Christ, K.M.; Sullivan, T.B.; Robert, L.C.; A Teebagy, P.; E Morgan, S.; Beatty, E.W.; Cicuto, B.J.; Nowd, P.K.; Bryan, D.J. Spatiotemporal microRNA profile in peripheral nerve regeneration: miR-138 targets vimentin and inhibits Schwann cell migration and proliferation. Neural Regen. Res. 2018, 13, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Xue, C.; Yao, M.; Wang, H.; Zhang, P.; Qian, T.; Zhou, S.; Li, S.; Yu, B.; Wang, Y.; et al. miR-129 controls axonal regeneration via regulating insulin-like growth factor-1 in peripheral nerve injury. Cell Death Dis. 2018, 9, 720. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.; Shi, X.; Zhang, Z.; Zhou, S.; Qian, T.; Wang, Y.; Ding, F.; Gu, X.; Yu, B. Hypoxia-Induced Upregulation of miR-132 Promotes Schwann Cell Migration After Sciatic Nerve Injury by Targeting PRKAG3. Mol. Neurobiol. 2016, 53, 5129–5139. [Google Scholar] [CrossRef]
- Wang, P.; He, J.; Wang, S.; Wang, X.; Liu, Q.; Peng, W.; Qian, T. miR-3075 Inhibited the Migration of Schwann Cells by Targeting Cntn2. Neurochem. Res. 2018, 43, 1879–1886. [Google Scholar] [CrossRef]
- Zou, D.; Zhou, X.; Liu, J.; Zhao, Y.; Jiang, X. MiR-34a regulates Schwann cell proliferation and migration by targeting CNTN2. NeuroReport 2020, 31, 1180–1188. [Google Scholar] [CrossRef]
- Viader, A.; Chang, L.-W.; Fahrner, T.; Nagarajan, R.; Milbrandt, J. MicroRNAs Modulate Schwann Cell Response to Nerve Injury by Reinforcing Transcriptional Silencing of Dedifferentiation-Related Genes. J. Neurosci. 2011, 31, 17358–17369. [Google Scholar] [CrossRef] [Green Version]
- Gökbuget, D.; Pereira, J.A.; Bachofner, S.; Marchais, A.; Ciaudo, C.; Stoffel, M.; Schulte, J.H.; Suter, U. The Lin28/let-7 axis is critical for myelination in the peripheral nervous system. Nat. Commun. 2015, 6, 8584. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Wang, X.; Gu, Y.; Chen, C.; Wang, Y.; Liu, J.; Hu, W.; Yu, B.; Wang, Y.; Ding, F.; et al. Let-7 microRNAs Regenerate Peripheral Nerve Regeneration by Targeting Nerve Growth Factor. Mol. Ther. 2015, 23, 423–433. [Google Scholar] [CrossRef] [Green Version]
- Verrier, J.D.; Lau, P.; Hudson, L.; Murashov, A.K.; Renne, R.; Notterpek, L. Peripheral myelin protein 22 is regulated post-transcriptionally by miRNA-29a. Glia 2009, 57, 1265–1279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Li, X.; Li, A.; Wu, G. miR-485-5p suppresses Schwann cell proliferation and myelination by targeting cdc42 and Rac1. Exp. Cell Res. 2020, 388, 111803. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.-L.; Yi, S.; Wang, Q.-H.; Zhao, L.-L.; Qin, J.; Wang, Y.-X.; Yu, B. miR-30c promotes Schwann cell remyelination following peripheral nerve injury. Neural Regen. Res. 2017, 12, 1708–1715. [Google Scholar] [CrossRef]
- Wu, D.; Raafat, M.; Pak, E.; Hammond, S.; Murashov, A. MicroRNA machinery responds to peripheral nerve lesion in an injury-regulated pattern. Neuroscience 2011, 190, 386–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adilakshmi, T.; Sudol, I.; Tapinos, N. Combinatorial Action of miRNAs Regulates Transcriptional and Post-Transcriptional Gene Silencing following in vivo PNS Injury. PLoS ONE 2012, 7, e39674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, B.; Zhou, S.; Wang, Y.; Ding, G.; Ding, F.; Gu, X. Profile of MicroRNAs following Rat Sciatic Nerve Injury by Deep Sequencing: Implication for Mechanisms of Nerve Regeneration. PLoS ONE 2011, 6, e24612. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Yu, B.; Wang, Y.; Yao, D.; Zhang, Z.; Gu, X. Identification and functional annotation of novel microRNAs in the proximal sciatic nerve after sciatic nerve transection. Sci. China Life Sci. 2011, 54, 806–812. [Google Scholar] [CrossRef] [Green Version]
- Bitetti, A.; Mallory, A.C.; Golini, E.; Carrieri, C.; Gutiérrez, H.C.; Perlas, E.; Pérez-Rico, Y.A.; Tocchini-Valentini, G.P.; Enright, A.J.; Norton, W.; et al. MicroRNA degradation by a conserved target RNA regulates animal behavior. Nat. Struct. Mol. Biol. 2018, 25, 244–251. [Google Scholar] [CrossRef]
- Deng, B.; Lv, W.; Duan, W.; Liu, Y.; Li, Z.; Song, X.; Cui, C.; Qi, X.; Wang, X.; Li, C. FGF9 modulates Schwann cell myelination in developing nerves and induces a pro-inflammatory environment during injury. J. Cell. Biochem. 2018, 119, 8643–8658. [Google Scholar] [CrossRef]
- Lv, W.; Deng, B.; Duan, W.; Li, Y.; Song, X.; Ji, Y.; Li, Z.; Liu, Y.; Wang, X.; Li, C.; et al. FGF9 alters the Wallerian degeneration process by inhibiting Schwann cell transformation and accelerating macrophage infiltration. Brain Res. Bull. 2019, 152, 285–296. [Google Scholar] [CrossRef]
- Bachelin, C.; Zujovic, V.; Buchet, D.; Mallet, J.; Evercooren, A.B.-V. Ectopic expression of polysialylated neural cell adhesion molecule in adult macaque Schwann cells promotes their migration and remyelination potential in the central nervous system. Brain 2010, 133, 406–420. [Google Scholar] [CrossRef] [PubMed]
- Bosse, F.; Hasenpusch-Theil, K.; Küry, P.; Müller, H.W. Gene expression profiling reveals that peripheral nerve regeneration is a consequence of both novel injury-dependent and reactivated developmental processes. J. Neurochem. 2006, 96, 1441–1457. [Google Scholar] [CrossRef] [PubMed]
- Weiss, T.; Taschner-Mandl, S.; Bileck, A.; Slany, A.; Kromp, F.; Rifatbegovic, F.; Frech, C.; Windhager, R.; Kitzinger, H.B.; Tzou, C.-H.J.; et al. Proteomics and transcriptomics of peripheral nerve tissue and cells unravel new aspects of the human Schwann cell repair phenotype: Deciphering the Repair Phenotype of Human Schwann Cells. Glia 2016, 64, 2133–2153. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, M.B.; Laranjeira, S.G.; Eriksson, T.M.; Jessen, K.R.; Mirsky, R.; Quick, T.J.; Phillips, J.B. Characterising cellular and molecular features of human peripheral nerve degeneration. Acta Neuropathol. Commun. 2020, 8, 51. [Google Scholar] [CrossRef] [PubMed]
- Braun, S.; Croizat, B.; Lagrange, M.-C.; Warter, J.-M.; Poindron, P. Neurotrophins increase motoneurons’ ability to innervate skeletal muscle fibers in rat spinal cord-human muscle cocultures. J. Neurol. Sci. 1996, 136, 17–23. [Google Scholar] [CrossRef]
- Wang, W.; Li, Y.; Li, Y.; Hong, A.; Wang, J.; Lin, B.; Li, R. NDRG3 is an androgen regulated and prostate enriched gene that promotesin vitroandin vivoprostate cancer cell growth. Int. J. Cancer 2009, 124, 521–530. [Google Scholar] [CrossRef]
- Zhou, L.; Too, H.-P. Mitochondrial Localized STAT3 Is Involved in NGF Induced Neurite Outgrowth. PLoS ONE 2011, 6, e21680. [Google Scholar] [CrossRef]
- Montalban, E.; Mattugini, N.; Ciarapica, R.; Provenzano, C.; Savino, M.; Scagnoli, F.; Prosperini, G.; Carissimi, C.; Fulci, V.; Matrone, C.; et al. MiR-21 is an Ngf-modulated microRNA that supports Ngf signaling and regulates neuronal degeneration in PC12 cells. NeuroMolecular Med. 2014, 16, 415–430. [Google Scholar] [CrossRef] [Green Version]
- Strickland, I.T.; Richards, L.; Holmes, F.E.; Wynick, D.; Uney, J.B.; Wong, L.-F. Axotomy-Induced miR-21 Promotes Axon Growth in Adult Dorsal Root Ganglion Neurons. PLoS ONE 2011, 6, e23423. [Google Scholar] [CrossRef] [Green Version]
- Zhong, L.; Xiao, W.; Wang, F.; Liu, J.; Zhi, L. miR-21-5p inhibits neuropathic pain development via directly targeting C-C motif ligand 1 and tissue inhibitor of metalloproteinase-3. J. Cell. Biochem. 2019, 120, 16614–16623. [Google Scholar] [CrossRef]
- Fan, D.; Kassiri, Z. Biology of Tissue Inhibitor of Metalloproteinase 3 (TIMP3), and Its Therapeutic Implications in Cardiovascular Pathology. Front. Physiol. 2020, 11, 661. [Google Scholar] [CrossRef]
- Jamsuwan, S.; Klimaschewski, L.; Hausott, B. Simultaneous Knockdown of Sprouty2 and PTEN Promotes Axon Elongation of Adult Sensory Neurons. Front. Cell. Neurosci. 2020, 13, 583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCauslin, C.S.; Heath, V.; Colangelo, A.M.; Malik, R.; Lee, S.; Mallei, A.; Mocchetti, I.; Johnson, P.F. CAAT/Enhancer-binding Protein δ and cAMP-response Element-binding Protein Mediate Inducible Expression of the Nerve Growth Factor Gene in the Central Nervous System. J. Biol. Chem. 2006, 281, 17681–17688. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Zhang, S.; Wang, Y.; Yi, S.; Zhao, L.; Tang, X.; Yu, B.; Gu, X.; Ding, F. miR-21 and miR-222 inhibit apoptosis of adult dorsal root ganglion neurons by repressing TIMP3 following sciatic nerve injury. Neurosci. Lett. 2015, 586, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Jin, B.; Li, D.-W.; Shen, B.; Gong, N.; Zhang, T.-Z.; Dong, P. Effect of laminin-binding BDNF on induction of recurrent laryngeal nerve regeneration by miR-222 activation of mTOR signal pathway. Am. J. Transl. Res. 2015, 7, 1071–1080. [Google Scholar] [PubMed]
- Wang, H.; Zuo, Y.; Ding, M.; Ke, C.; Yan, R.; Zhan, H.; Liu, J.; Wang, W.; Li, N.; Wang, J. LASS2 inhibits growth and invasion of bladder cancer by regulating ATPase activity. Oncol. Lett. 2017, 13, 661–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, N.; Nagarajan, R.; Wang, J.Y.T.; Svaren, J.; LaPash, C.; Araki, T.; E Schmidt, R.; Milbrandt, J. Nab proteins are essential for peripheral nervous system myelination. Nat. Neurosci. 2005, 8, 932–940. [Google Scholar] [CrossRef]
- Ayachi, O.; Barlin, M.; Broxtermann, P.N.; Kashkar, H.; Mauch, C.; Zigrino, P. The X-linked inhibitor of apoptosis protein (XIAP) is involved in melanoma invasion by regulating cell migration and survival. Cell. Oncol. 2019, 42, 319–329. [Google Scholar] [CrossRef]
- Abidin, S.Z.; Fam, S.-Z.; Chong, C.-E.; Abdullah, S.; Cheah, P.-S.; Nordin, N.; Ling, K.-H. miR-3099 promotes neurogenesis and inhibits astrogliogenesis during murine neural development. Gene 2019, 697, 201–212. [Google Scholar] [CrossRef]
- Blackmore, M.G.; Wang, Z.; Lerch, J.K.; Motti, D.; Zhang, Y.P.; Shields, C.B.; Lee, J.K.; Goldberg, J.L.; Lemmon, V.P.; Bixby, J.L. Krüppel-like Factor 7 engineered for transcriptional activation promotes axon regeneration in the adult corticospinal tract. Proc. Natl. Acad. Sci. USA 2012, 109, 7517–7522. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, W.-Y.; Jia, H.; Zhai, F.-G.; Qu, W.-R.; Cheng, Y.-X.; Liu, Y.-C.; Deng, L.-X.; Guo, S.-F.; Jin, Z.-S. KLF7-transfected Schwann cell graft transplantation promotes sciatic nerve regeneration. Neuroscience 2017, 340, 319–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.-W.; Jiang, J.-J.; Gao, Y.; Wang, R.-Y.; Tu, G.-J. MicroRNA-210 promotes sensory axon regeneration of adult mice in vivo and in vitro. Neurosci. Lett. 2016, 622, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yin, B.; Wang, B.; Ma, Z.; Liu, W.; Lv, G. MicroRNA-210 Promotes Proliferation and Invasion of Peripheral Nerve Sheath Tumor Cells Targeting EFNA3. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2014, 21, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.-P.; Oksuz, I.; Svaren, J.; Awatramani, R. Egr2-dependent microRNA-138 is dispensable for peripheral nerve myelination. Sci. Rep. 2018, 8, 3817. [Google Scholar] [CrossRef] [Green Version]
- Tan, D.; Wen, J.; Li, L.; Wang, X.; Qian, C.; Pan, M.; Lai, M.; Deng, J.; Hu, X.; Zhang, H.; et al. Inhibition of RhoA-Subfamily GTPases Suppresses Schwann Cell Proliferation Through Regulating AKT Pathway Rather Than ROCK Pathway. Front. Cell. Neurosci. 2018, 12, 437. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.-Y.; Zan, C.-F.; Li, J.-A.; Zhao, C.-F.; Li, S.-J.; Zhang, J.; Li, Z.-H.; Zhang, Q. Modified insulin-like growth factor 1 containing collagen-binding domain for nerve regeneration. Neural Regen. Res. 2018, 13, 298–303. [Google Scholar] [CrossRef]
- Jessen, K.R.; Mirsky, R. Negative regulation of myelination: Relevance for development, injury, and demyelinating disease. Glia 2008, 56, 1552–1565. [Google Scholar] [CrossRef]
- Martinez-Moreno, M.; O’Shea, T.M.; Zepecki, J.P.; Olaru, A.; Ness, J.K.; Langer, R.; Tapinos, N. Regulation of Peripheral Myelination through Transcriptional Buffering of Egr2 by an Antisense Long Non-coding RNA. Cell Rep. 2017, 20, 1950–1963. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Moon, C.; Zheng, Y.; Ratner, N. Cdc42 regulates schwann cell radial sorting and myelin sheath folding through NF2/merlin-dependent and independent signaling. Glia 2013, 61, 1906–1921. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Moon, C.; Niehaus, K.; Zheng, Y.; Ratner, N. Rac1 Controls Schwann Cell Myelination through cAMP and NF2/merlin. J. Neurosci. 2012, 32, 17251–17261. [Google Scholar] [CrossRef] [Green Version]
- Cai, W.; Zhao, Q.; Shao, J.; Zhang, J.; Li, L.; Ren, X.; Su, S.; Bai, Q.; Li, M.; Chen, X.; et al. MicroRNA-182 Alleviates Neuropathic Pain by Regulating Nav1.7 Following Spared Nerve Injury in Rats. Sci. Rep. 2018, 8, 16750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, J.; Dang, Y.; Zhang, W.; Zhao, X.; Zhang, C.; Hou, Z.; Jin, Y.; McNutt, M.A.; Marks, A.R.; Yin, Y. PTEN arginine methylation by PRMT6 suppresses PI3K–AKT signaling and modulates pre-mRNA splicing. Proc. Natl. Acad. Sci. USA 2019, 116, 6868–6877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cioffi, J.A.; Yue, W.Y.; Mendolia-Loffredo, S.; Hansen, K.R.; Wackym, P.A.; Hansen, M. MicroRNA-21 Overexpression Contributes to Vestibular Schwannoma Cell Proliferation and Survival. Otol. Neurotol. 2010, 31, 1455–1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradtmöller, M.; Hartmann, C.; Zietsch, J.; Jäschke, S.; Mautner, V.-F.; Kurtz, A.; Park, S.-J.; Baier, M.; Harder, A.; Reuss, D.; et al. Impaired Pten Expression in Human Malignant Peripheral Nerve Sheath Tumours. PLoS ONE 2012, 7, e47595. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| miR | Function | Effect | Target | Author, Year, Reference |
|---|---|---|---|---|
| miR-340 | Inflammation | Impair | TPA | Li et al. (2017), [37] |
| miR-182 | Inflammation, Migration | Impair | FGF9, NTM | Yu et al. (2012), [38] |
| miR-1 | Proliferation | Impair | BDNF | Yi et al. (2016), [39] |
| miR-1b | Proliferation, Migration | Impair | NDRG3 | Liu et al. (2018), [40] |
| miR-1b | Proliferation | Impair | KLF7 | Li et al. (2021), [41] |
| miR-124 | Proliferation, Migration | Impair | STAT 3 | Nagata et al. (2014), [42] |
| miR-21 | Proliferation | Promote | TGFb1, EPHA4, TIMP3 | Ning et al. (2020), [43] |
| miR-21 | Proliferation | Promote | PTEN | Lopez-Leal et al. (2020), [44] |
| miR-221/222 | Proliferation | Promote | LASS2 | Yu et al. (2012), [45] |
| miR-221 | Myelination | Impair | NAB1 | Zhao et al. (2018), [46] |
| miR-192 | Proliferation | Impair | XIAP | Liu et al. (2020), [47] |
| miR-3099 | Proliferation, Migration | Promote | None identified | Liu et al. (2019), [48] |
| miR-146b | Proliferation | Impair | KLF7 | Li et al. (2018), [49] |
| miR-210 | Proliferation, Migration, Myelination | Promote proliferation and migration, impair myelination | None identified | Zhang et al. (2017), [50] |
| miR-9 | Migration | Impair | CTHRC1 | Zhou et al. (2014), [51] |
| miR-9/let-7 family | Migration | Impair | NTN1, DCC | Wang et al. (2019), [52] |
| miR-138 | Migration | Impair | Vimentin | Sullivan et al. (2018), [53] |
| miR-129 | Migration | Impair | IGF9 | Zhu et al. (2018), [54] |
| miR-132 | Migration | Promote | PRKAG3 | Yao et al. (2016), [55] |
| miR-3075 | Migration | Impair | CNTN2 | Wang et al. (2018), [56] |
| miR-34a | Migration, Proliferation | Impair | CNTN2 | Zou et al. (2020), [57] |
| miR-34a | Myelination, Proliferation | Promote myelination, Impair proliferation | NOTCH1, CCND1 | Viader et al. (2011), [58] |
| miR-140 | Myelination | Impair | EGR2 | Viader et al. (2011), [58] |
| Lin28/let-7-axis | Myelination | Promote | NOTCH1 | Gökbuget et al. (2018), [59] |
| let-7 | Proliferation | Impair | NGF | Li et al. (2015), [60] |
| miR-29a/miR-29b | Myelination | Impair | PMP22 | Verrier et al. (2009), [61] |
| miR-485-5p | Myelination | Impair | CDC42, RAC1 | Zhang et al. (2020), [62] |
| miR-30c | Myelination | Impair | MBP | Yi et al. (2017), [63] |
| Author, Year, Reference | Method | Species | Gender | Lesion Model | Time Points (Postinjury) |
|---|---|---|---|---|---|
| Viader et al. (2011), [58] | Microarray | Mice | Both | Crush | 4 d, 14 d |
| Wu et al. (2011), [64] | Microarray | Mice | Nd | Crush | 4 d, 7 d |
| Adilakshimi et al. (2012), [65] | Microarray | Mice | Female | Cut | 6 h, 24 h |
| Yu et al. (2012), [45] | Microarray | Rat | Male | Cut | 1 d, 4 d, 7 d, 14 d |
| Sullivan et al. (2018), [53] | Microarray | Rat | Male | Cut | 30 d, 60 d, 70 d |
| Arthur-Farraj et al. (2017), [34] | Illumina sequencing | Mice | Male | Cut | 3 d, 7 d |
| Yu et al. (2011), [66] | Solexa sequencing | Rat | Male | Cut | 1 d, 4 d, 7 d, 14 d |
| Li et al. (2011), [67] | Solexa sequencing | Rat | Male | Cut | 1 d, 4 d, 7 d, 14 d |
| Acute Phase | Late | |||||||
|---|---|---|---|---|---|---|---|---|
| miR | 6 h | 1 d | 4 d | 7 d | 14 d | 30 d | 60 d | 90 d |
| 182 | UP | DOWN | DOWN | NOT CLEAR | DOWN | DOWN | DOWN | |
| 340 | DOWN | DOWN | DOWN | NOT CLEAR | UP | UP | UP | |
| 132 | UP | UP | UP | UP | ||||
| 3075 | UP | UP | UP | UP | ||||
| 9 | DOWN | DOWN | DOWN | DOWN | DOWN | DOWN | DOWN | |
| 129 | NOT CLEAR | DOWN | DOWN | DOWN | UP | UP | UP | |
| 138 | DOWN | DOWN | DOWN | DOWN | DOWN | DOWN | DOWN | DOWN |
| 29a | NOT CLEAR | UP | UP | UP | ||||
| 29b | UP | UP | UP | UP | ||||
| 485 | UP | UP | NOT CLEAR | NOT CLEAR | ||||
| 30c | DOWN | DOWN | DOWN | DOWN | UP | |||
| 140 | DOWN | DOWN | DOWN | NOT CLEAR | UP | UP | UP | NORMAL |
| 34a | DOWN | DOWN | DOWN | DOWN | NOT CLEAR | |||
| let-7-a | DOWN | DOWN | DOWN | UP | NORMAL | UP | UP | |
| let-7-b | DOWN | DOWN | DOWN | UP | NORMAL | NORMAL | UP | |
| let-7-c | NORMAL | DOWN | DOWN | UP | ||||
| let-7-d | UP | DOWN | DOWN | UP | UP | UP | UP | |
| let-7-e | UP | DOWN | DOWN | UP | NORMAL | UP | UP | |
| let-7-f | NOT CLEAR | DOWN | DOWN | NORMAL | DOWN | NORMAL | UP | |
| 1 | DOWN | DOWN | DOWN | DOWN | DOWN | DOWN | NORMAL | |
| 124 | NOT CLEAR | DOWN | DOWN | DOWN | DOWN | DOWN | DOWN | |
| 192 | DOWN | DOWN | DOWN | NOT CLEAR | ||||
| 210 | DOWN | DOWN | DOWN | UP | ||||
| 146b | UP | UP | UP | UP | NOT CLEAR | UP | UP | UP |
| 21 | UP | UP | UP | UP | UP | UP | UP | |
| 221 | UP | UP | UP | UP | UP | UP | UP | |
| 222 | UP | DOWN | UP | UP | UP | UP | UP | |
| 3099 | UP | UP | NOT CLEAR | UP | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borger, A.; Stadlmayr, S.; Haertinger, M.; Semmler, L.; Supper, P.; Millesi, F.; Radtke, C. How miRNAs Regulate Schwann Cells during Peripheral Nerve Regeneration—A Systemic Review. Int. J. Mol. Sci. 2022, 23, 3440. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23073440
Borger A, Stadlmayr S, Haertinger M, Semmler L, Supper P, Millesi F, Radtke C. How miRNAs Regulate Schwann Cells during Peripheral Nerve Regeneration—A Systemic Review. International Journal of Molecular Sciences. 2022; 23(7):3440. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23073440
Chicago/Turabian StyleBorger, Anton, Sarah Stadlmayr, Maximilian Haertinger, Lorenz Semmler, Paul Supper, Flavia Millesi, and Christine Radtke. 2022. "How miRNAs Regulate Schwann Cells during Peripheral Nerve Regeneration—A Systemic Review" International Journal of Molecular Sciences 23, no. 7: 3440. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23073440