
Melatonin Positively Regulates Both Dark- and Age-Induced Leaf Senescence by Reducing ROS Accumulation and Modulating Abscisic Acid and Auxin Biosynthesis in Cucumber Plants

Abstract
:1. Introduction
2. Results
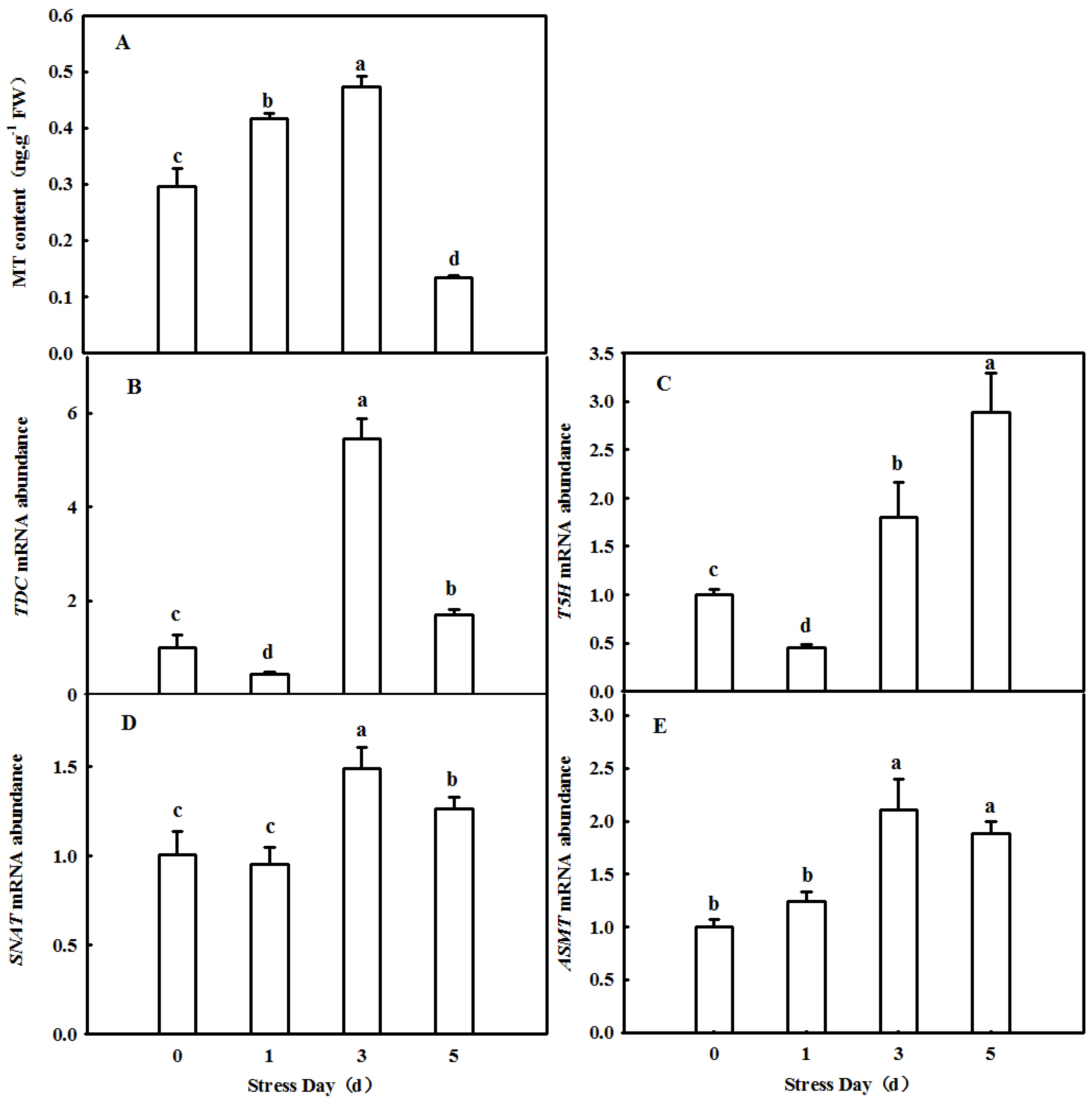
2.1. Dynamic Changes in Endogenous MT in Cucumber Leaves in Which Senescence Was Induced by Darkness
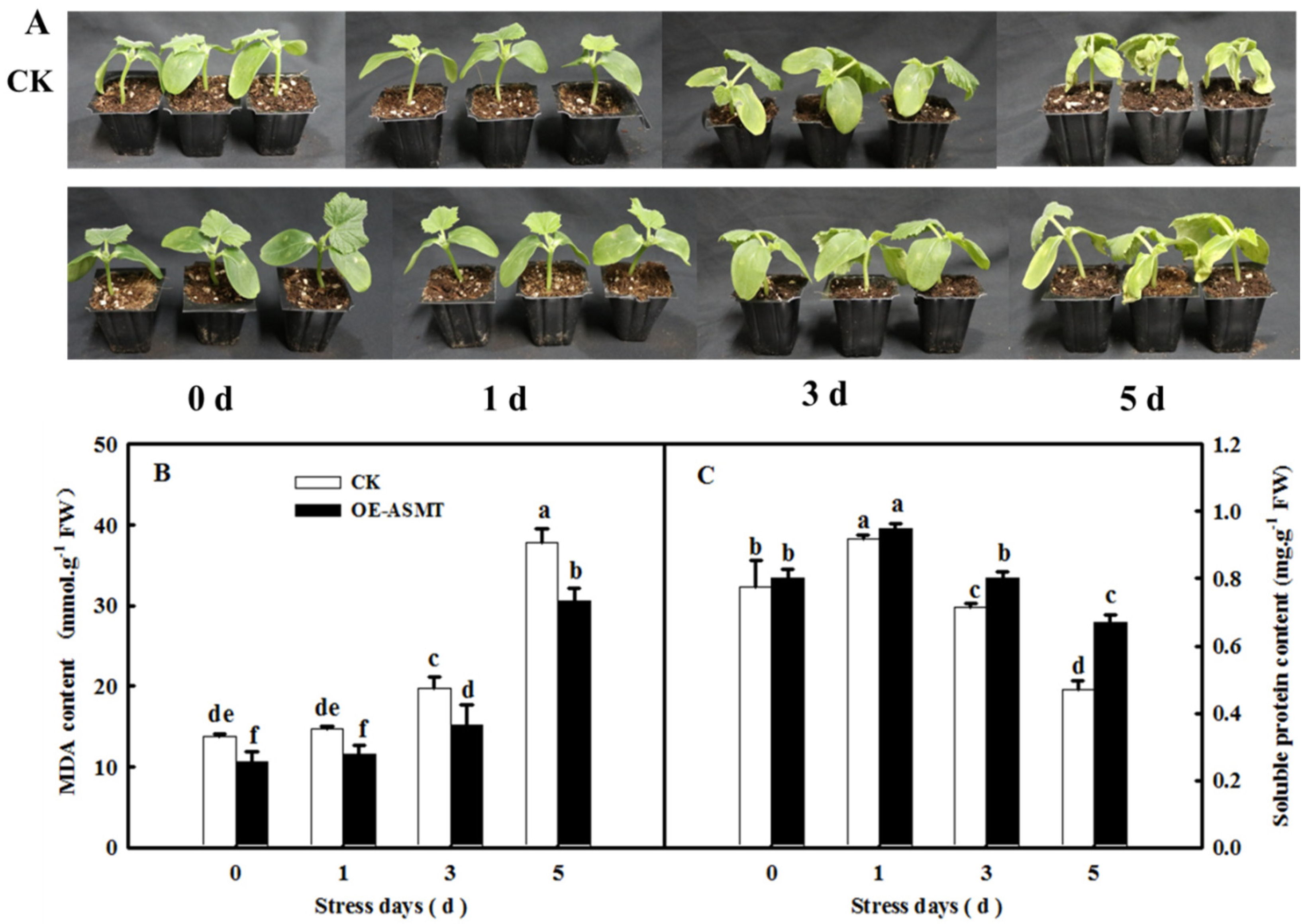
2.2. Effects of ASMT Overexpression on Dark-Induced Cucumber Leaf Senescence
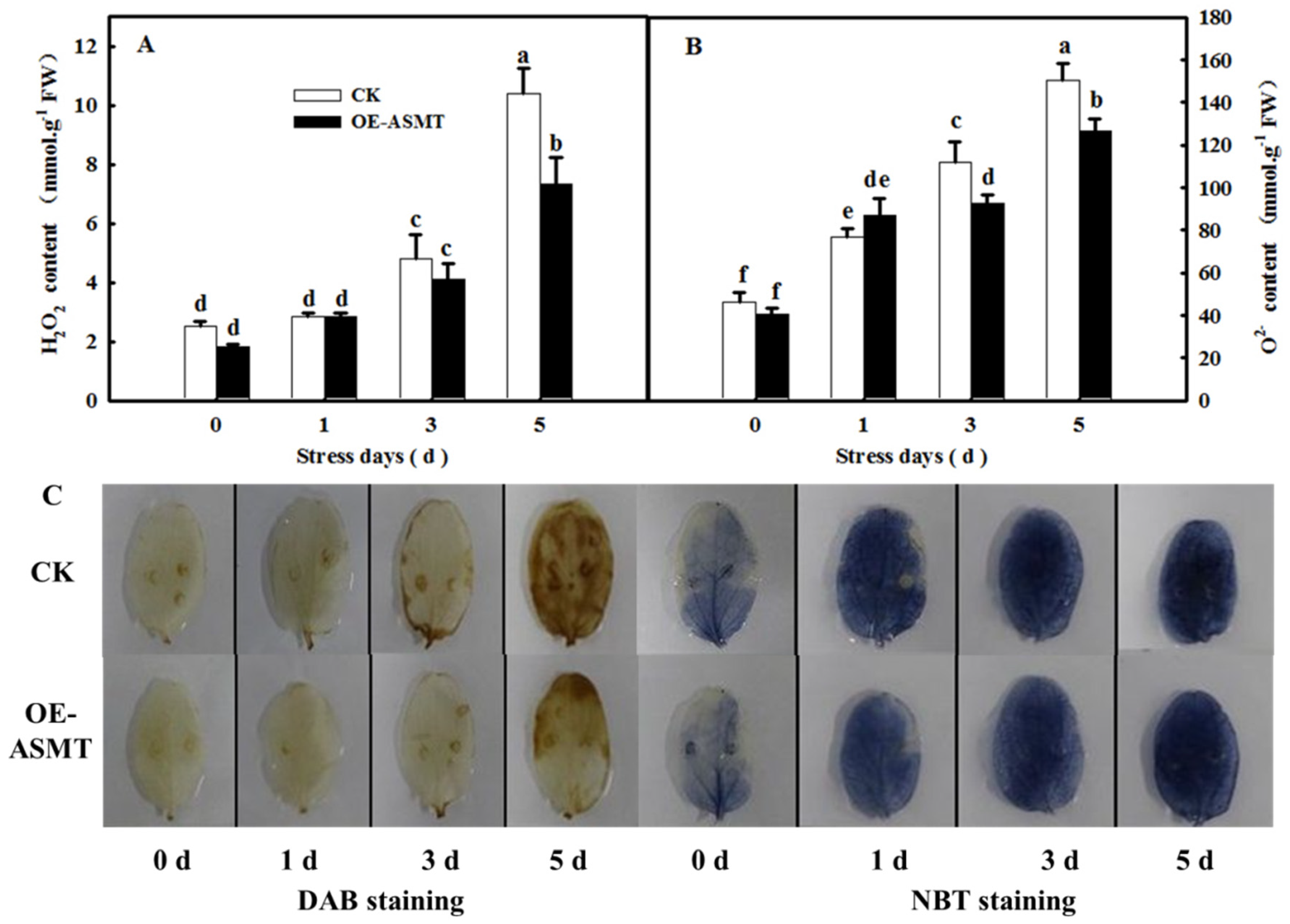
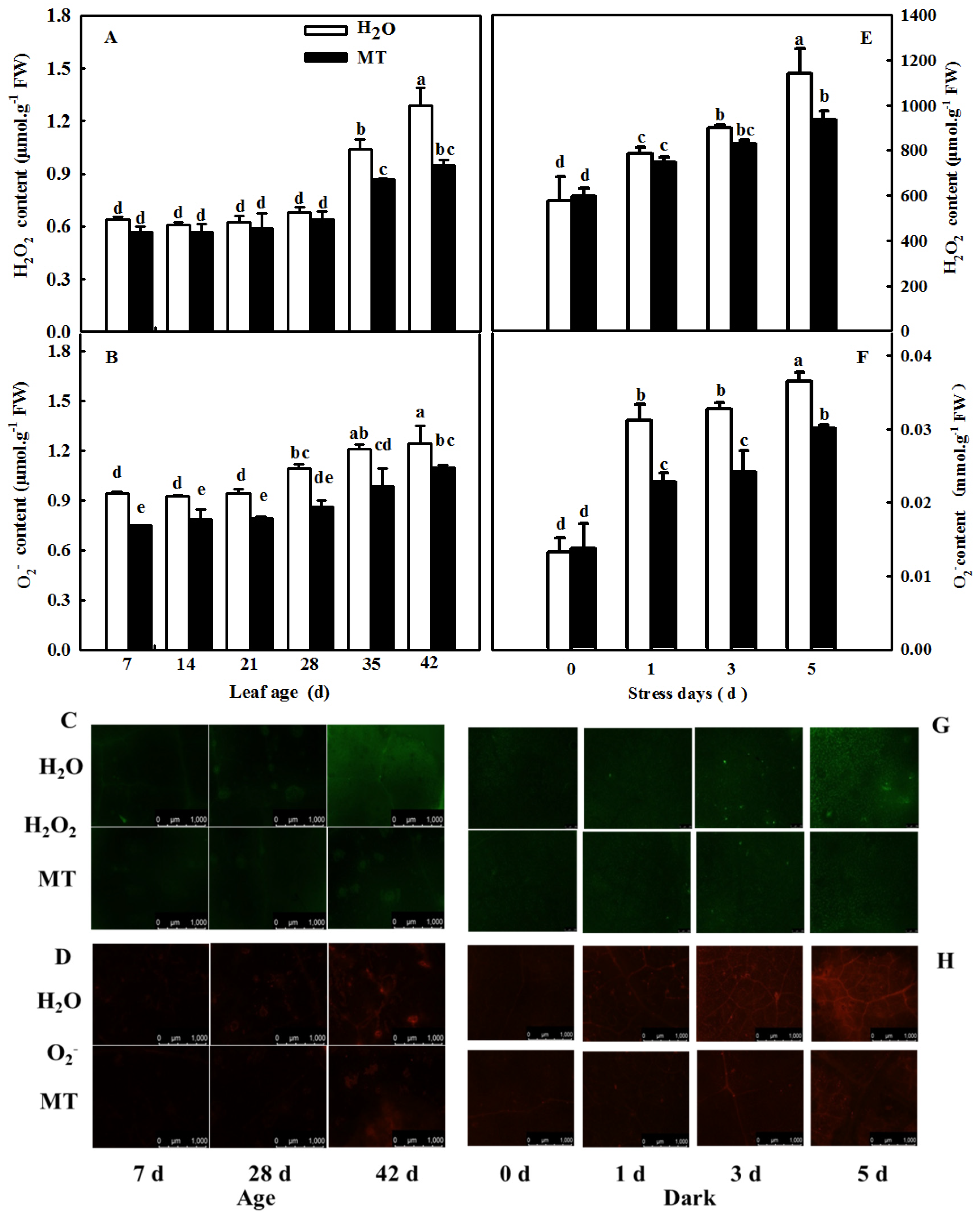
2.3. Effects of ASMT Overexpression on ROS Content in Cucumber Leaves in Which Senescence Was Induced by Darkness
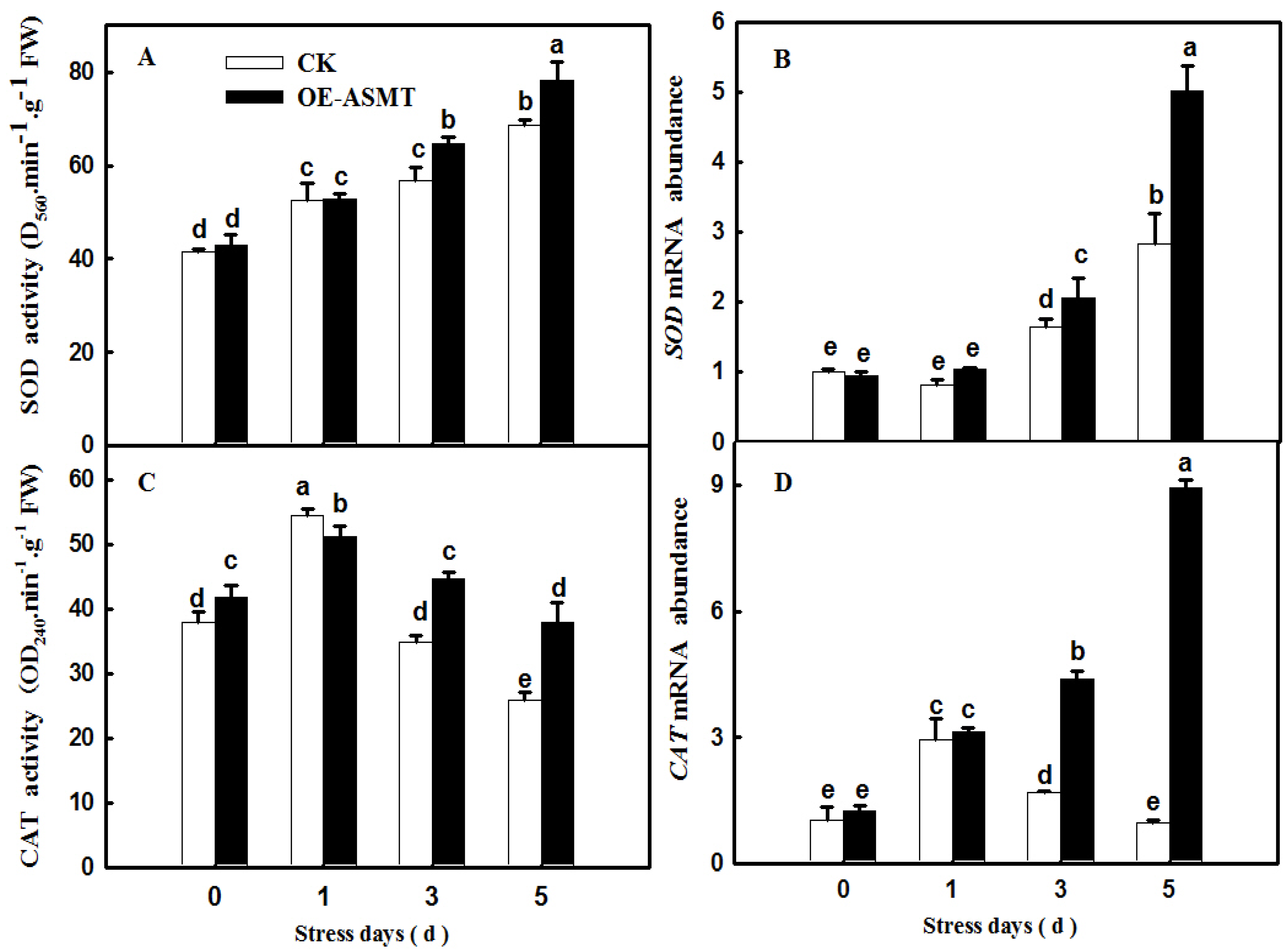
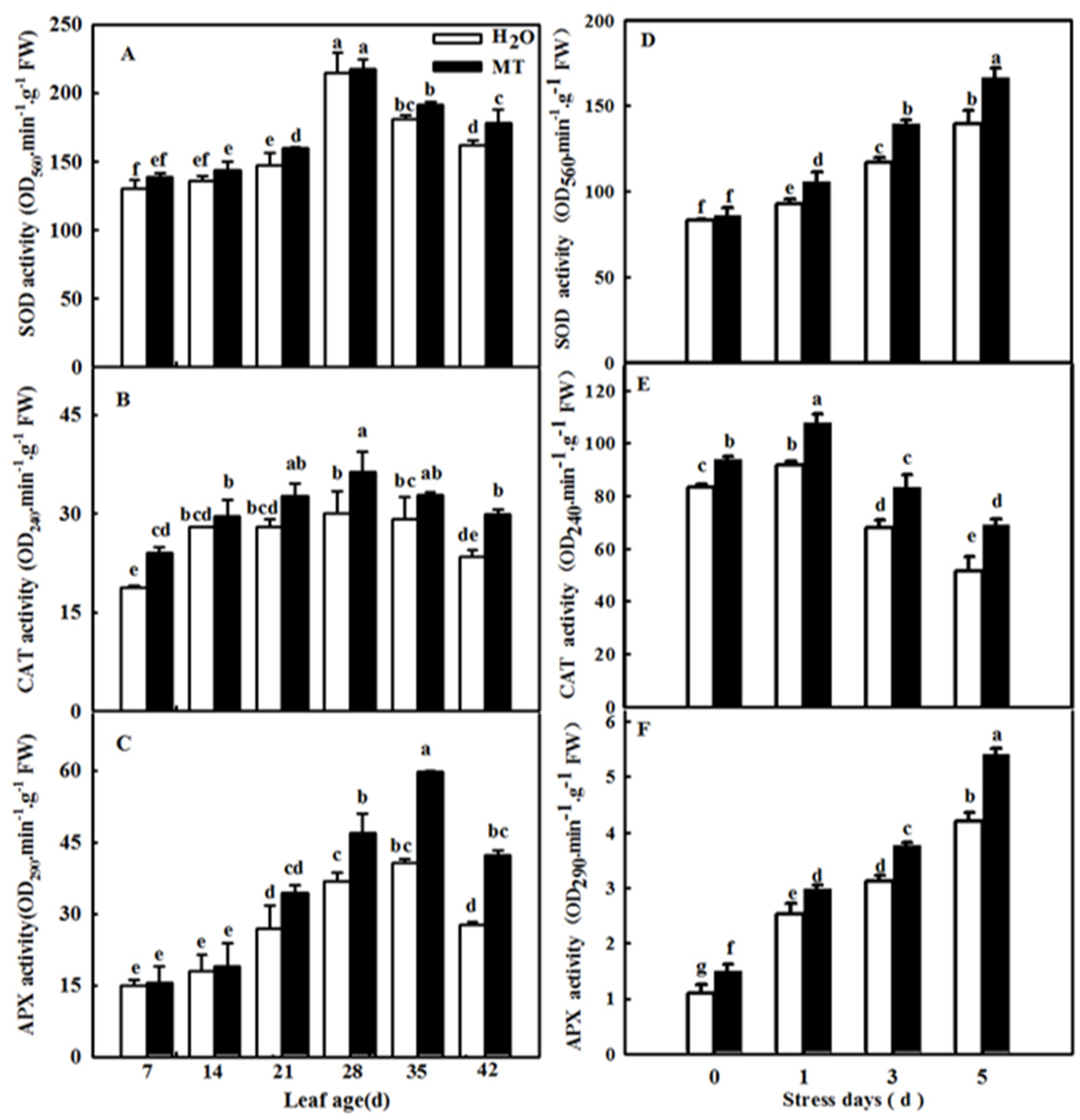
2.4. Effects of ASMT Overexpression on Antioxidant Enzyme Activity in Cucumber Leaves in Which Senescence Was Induced by Darkness
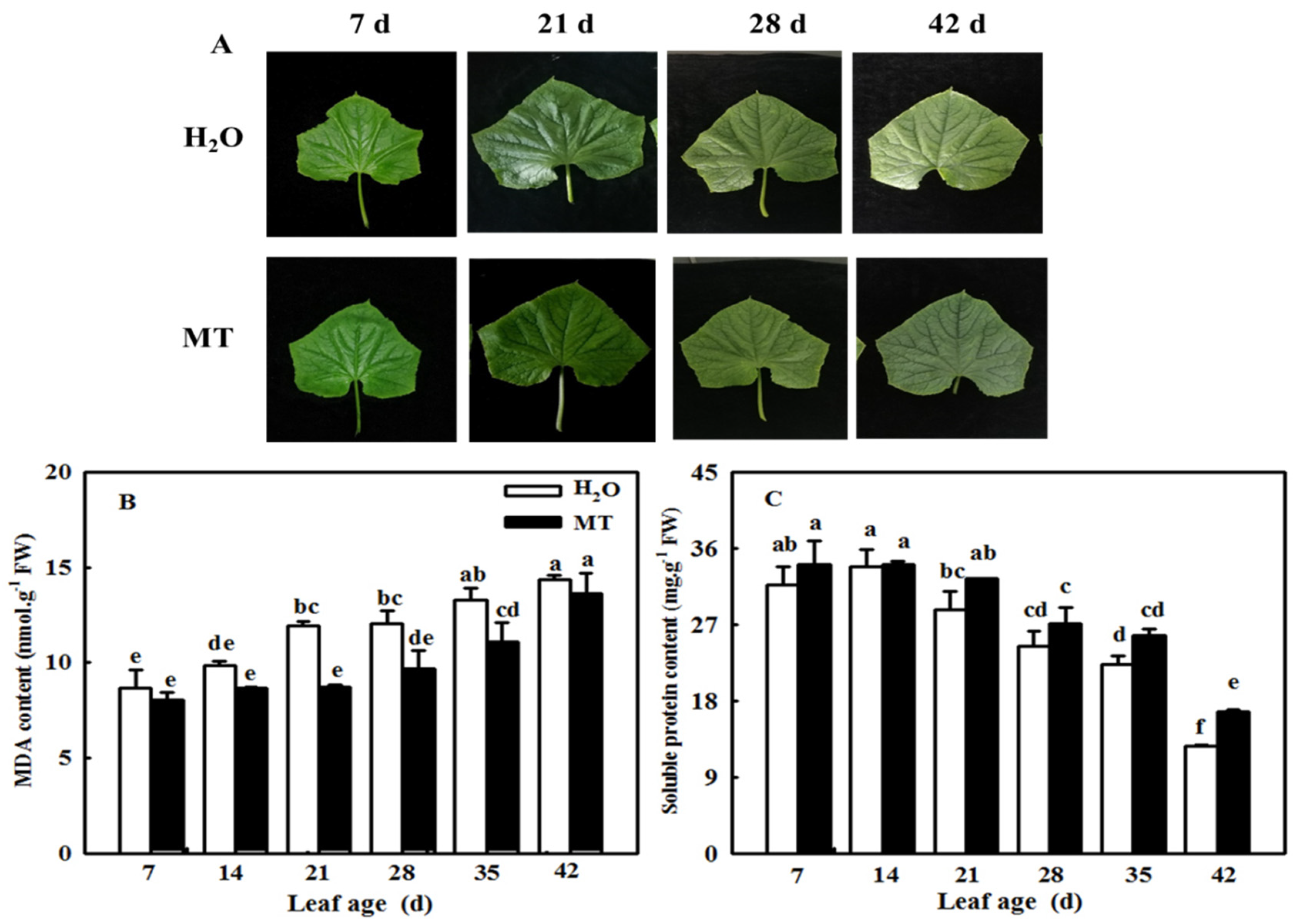
2.5. MT Delays the Cucumber Leaf Senescence Induced by Age
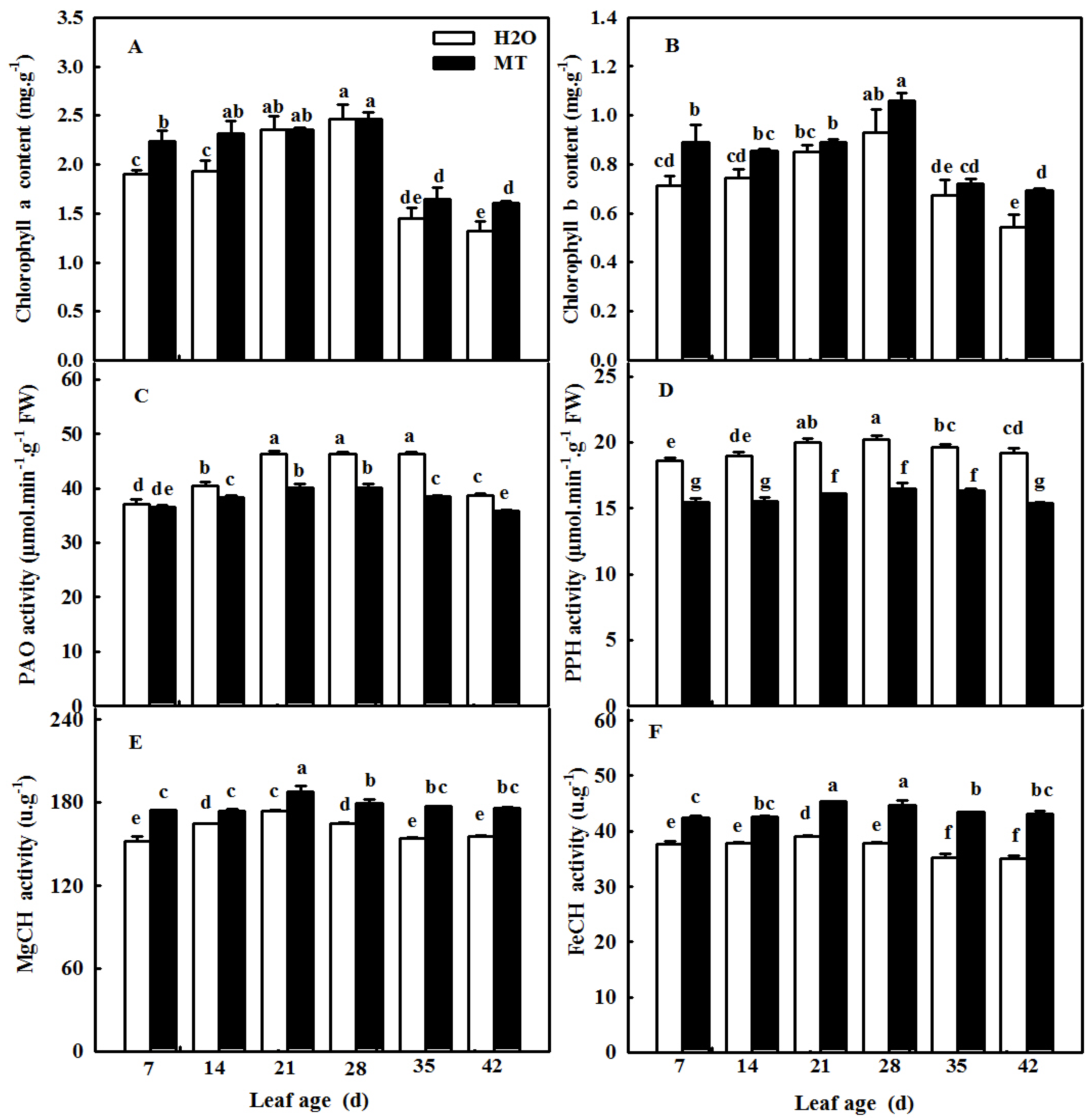
2.6. MT Regulates Chlorophyll Metabolism Enzyme Activity to Maintain Chlorophyll Content during Cucumber Leaf Senescence
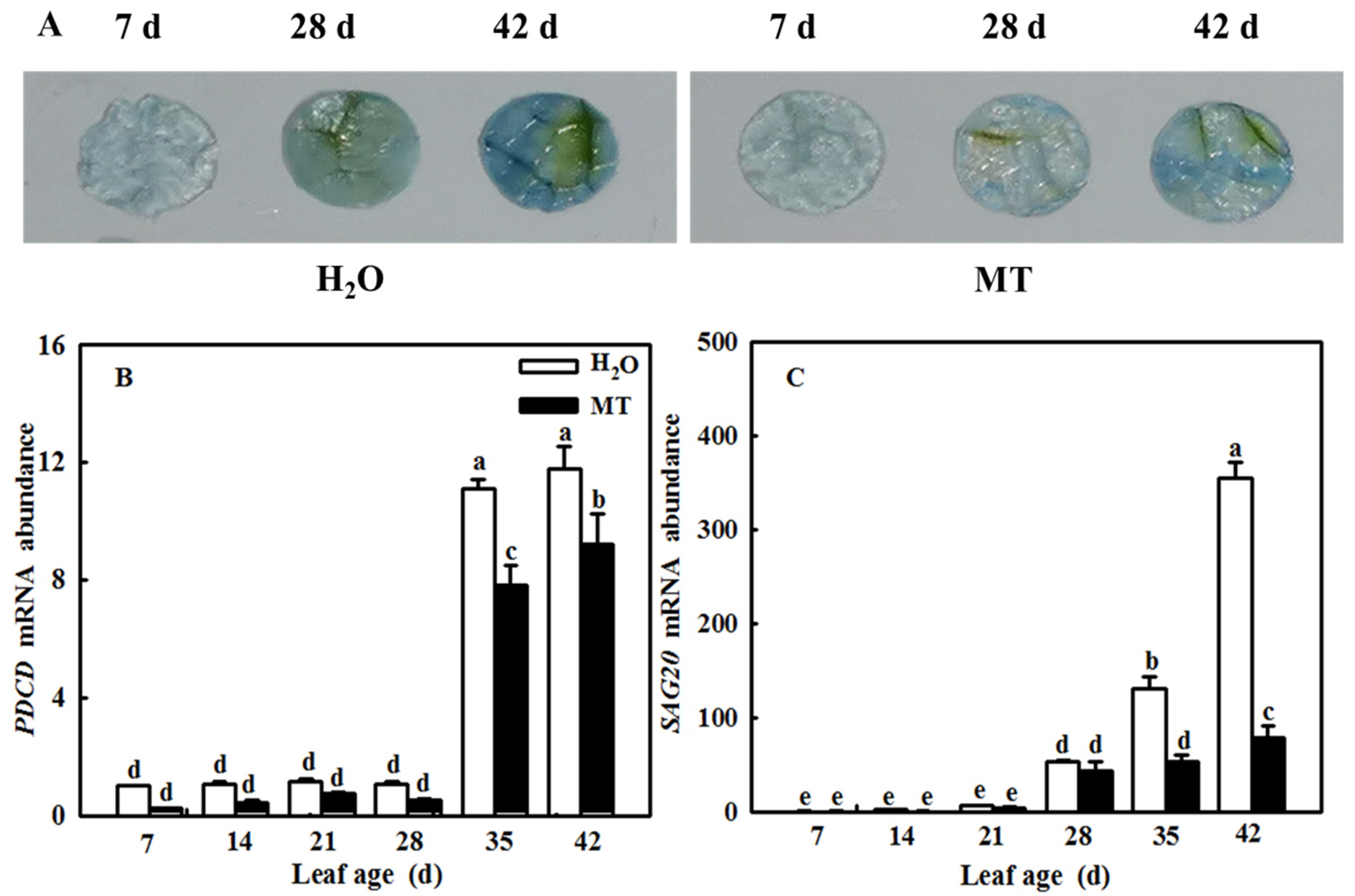
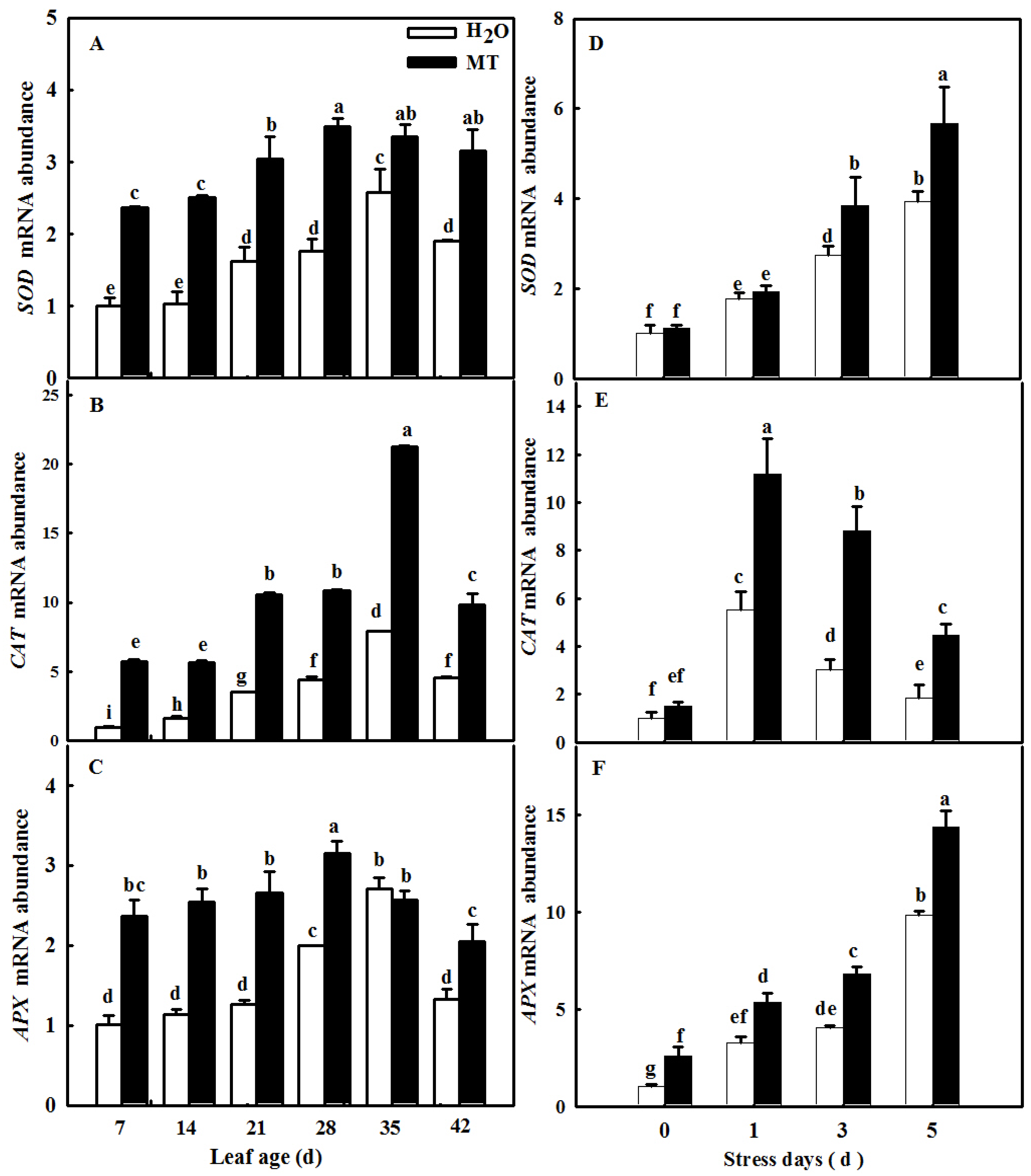
2.7. MT Downregulates Cell-Death- and Senescence-Related Gene mRNA Abundance to Delay Leaf Senescence Induced by Age
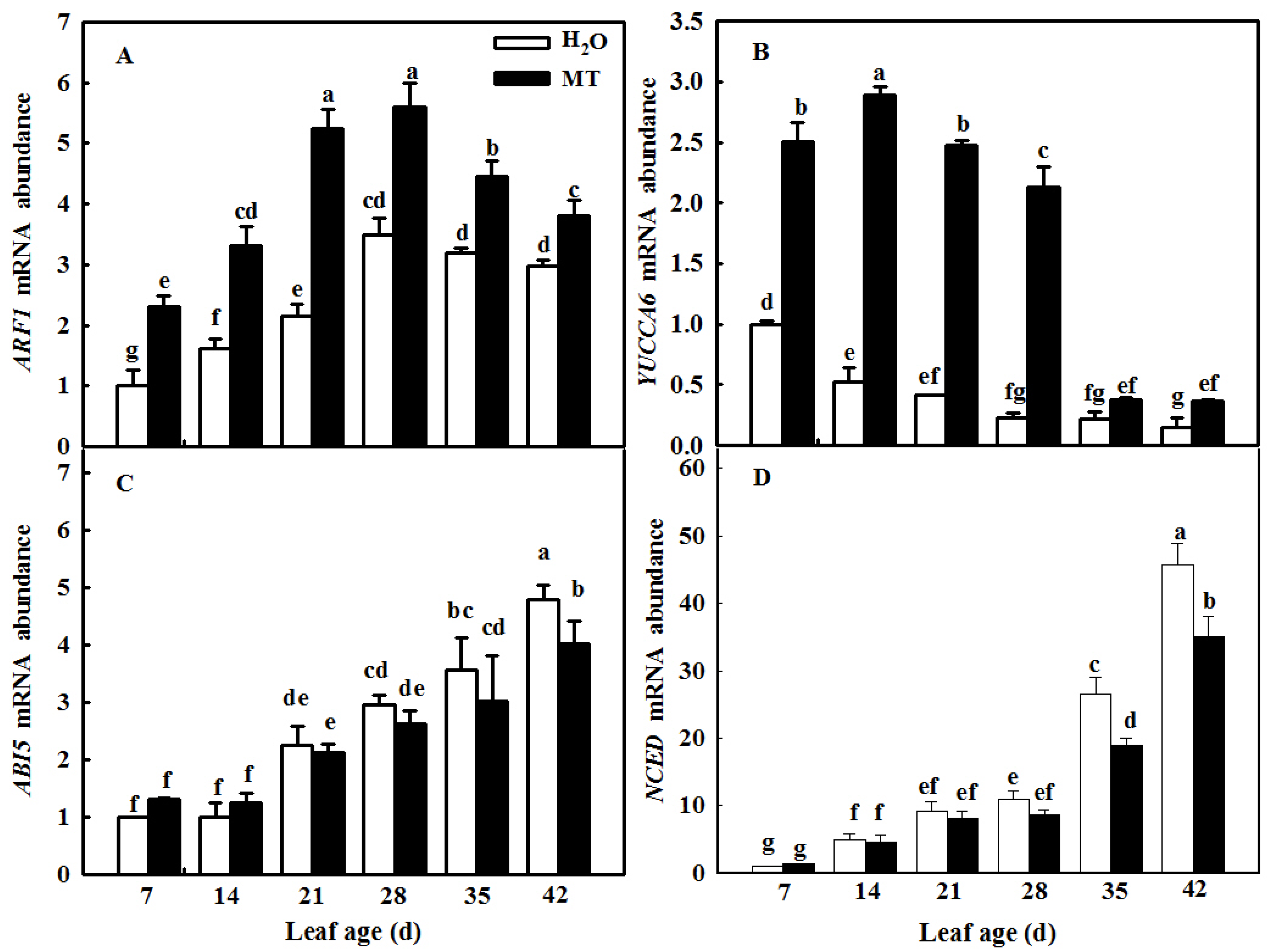
2.8. MT Modulates IAA and ABA Signals and Synthesis to Delay Leaf Senescence Induced by Age
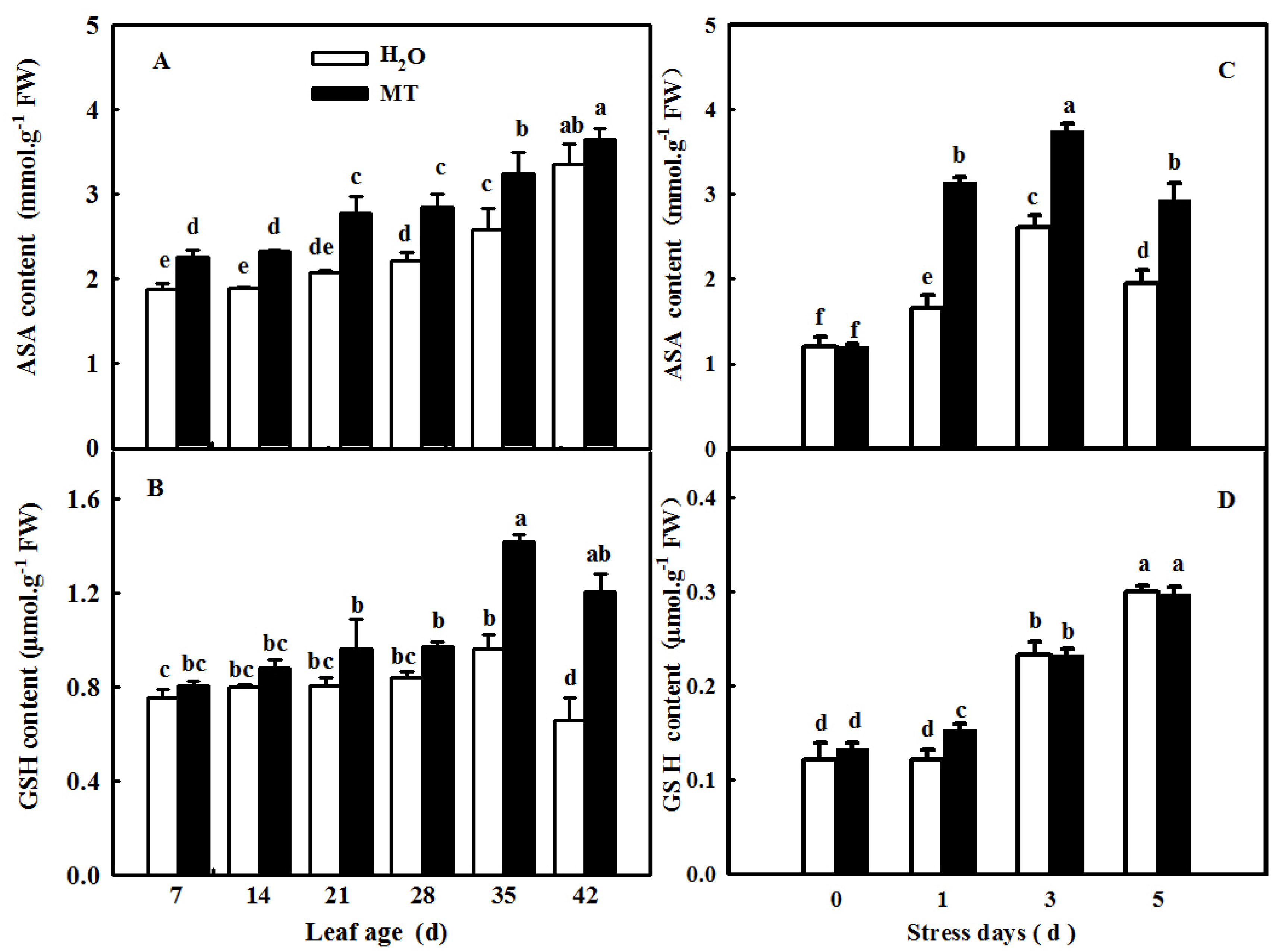
2.9. MT-Delayed Leaf Senescence Is Associated with Increased ROS-Scavenging Activity
3. Discussion
4. Materials and Methods
4.1. Cucumber Transient Transformation
4.2. Measurement of MDA, Soluble Protein, and Chlorophyll Contents
4.3. Key Chlorophyll Degradation Enzyme Activities
4.4. Measurement of ROS Content
4.5. NBT and DAB Staining
4.6. Antioxidant Enzyme Activity Assay
4.7. Determination of the Redox Substance Contents
4.8. Trypan Blue Staining for Cell Death
4.9. Measurements of MT Content
4.10. RNA Extraction and Gene Expression Analysis
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sun, J.L.; Sui, X.L.; Huang, H.Y.; Wang, S.H.; Wei, Y.X.; Zhang, Z.X. Low Light Stress Down-Regulated Rubisco Gene Expression and Photosynthetic Capacity During Cucumber (Cucumis sativus L.) Leaf Development. J. Integr. Agric. 2014, 13, 997–1007. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Gao, Y.; Gao, C.; Liu, S.; Zhang, J.; Chen, G.; Zhang, S.; Wu, F. Study of the physiological mechanism of delaying cucumber senescence by wheat intercropping pattern. J. Plant Physiol. 2019, 234, 154–166. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.Y.; Dong, F.Q.; Liu, L.A.; Shi, L.; Zhang, W.F.; Jiang, C.D. Dorsoventral variation in photosynthesis during leaf senescence probed by chlorophyll a fluorescence induction kinetics in cucumber and maize plants. Photosynthetica 2020, 58, 479–487. [Google Scholar] [CrossRef] [Green Version]
- Betania, F.Q.; Yoo-Sun, N.; Edward, H.; Richard, M.A. Molecular aspects of leaf senescence. Trends Plant Sci. 2000, 5, 278–282. [Google Scholar]
- Nooden, L.D.; Penny, J.P. Correlative controls of senescence and plant death in Arabidopsis thaliana Brassicaceae. J. Exp. Bot. 2001, 52, 2151–2159. [Google Scholar] [CrossRef]
- Thakur, N.; Sharma, V.; Kishore, K. Leaf senescence, an overview. Indian J. Plant Physiol. 2019, 21, 225–238. [Google Scholar]
- Zhang, J.; Shi, Y.; Zhang, X.; Du, H.; Xu, B.; Huang, B. Melatonin suppression of heat-induced leaf senescence involves changes in abscisic acid and cytokinin biosynthesis and signaling pathways in perennial ryegrass (Lolium perenne L.). Environ. Exp. Bot. 2017, 138, 36–45. [Google Scholar]
- Ma, X.; Zhang, J.; Burgess, P.; Rossi, S.; Huang, B. Interactive effects of melatonin and cytokinin on alleviating drought induced leaf senescence in creeping bentgrass (Agrostis stolonifera). Environ. Exp. Bot. 2018, 145, 1–11. [Google Scholar] [CrossRef]
- Petrov, V.; Hille, J.; Mueller-Roeber, B.; Gechev, T.S. ROS-mediated abiotic stress-induced programmed cell death in plants. Front. Plant Sci. 2015, 6, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Liu, J.; Zhou, L.; Pan, G.; Li, Z.; Zaidi, S.; Cheng, F. Senescence-specific change in ROS scavenging enzyme activities and regulation of various SOD isozymes to ROS levels in psf mutant rice leaves. Plant Physiol. Biochem. 2016, 109, 248–261. [Google Scholar] [CrossRef]
- Tan, X.L.; Zhao, Y.T.; Shan, W.; Kuang, J.F.; Chen, J.Y. Melatonin delays leaf senescence of postharvest Chinese flowering cabbage through ROS homeostasis. Food Res. Int. 2020, 138, 109790. [Google Scholar]
- Zhang, K.; Xia, X.; Zhang, Y.; Gan, S.S. An ABA-regulated and golgi-localized protein phosphatase controls water loss during leaf senescence in Arabidopsis. Plant J. 2012, 69, 667–678. [Google Scholar] [PubMed]
- Zhang, Y.; Ji, T.; Li, T.; Tian, Y.; Wang, L.; Liu, W. Jasmonic acid promotes leaf senescence through MYC2-mediated repression of CATALASE2 expression in Arabidopsis. Plant Sci. 2020, 299, 110–604. [Google Scholar]
- Yu, X.; Xu, Y.; Yan, S. Salicylic acid and ethylene coordinately promote leaf Senescence. J. Integr. Plant Biol. 2021, 63, 823–827. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.I.; Murphy, A.S.; Baek, D.; Lee, S.W.; Narasimhan, M.L. YUCCA6 over-expression demonstrates auxin function in delaying leaf senescence in Arabidopsis thaliana. J. Exp. Bot. 2011, 62, 3981–3992. [Google Scholar] [CrossRef] [PubMed]
- Oshry, M.; Evyatar, S.; Štěpán, K.; Petr, T.; Asaph, A.; Rivka, E. Silicon promotes cytokinin biosynthesis and delayssenescence in Arabidopsis and Sorghum. Plant Cell Environ. 2017, 40, 1189–1196. [Google Scholar]
- Lerner, A.B.; Case, J.D.; Takahashi, Y.; Lee, T.H.; Mori, W. Isolation of melatonin, the pineal gland factor that lightens melanocytes. J. Am. Chem. Soc. 1958, 80, 2587. [Google Scholar] [CrossRef]
- Serkan, E. Melatonin promotes plant growth by maintaining integration and coordination between carbon and nitrogen metabolisms. Plant Cell Rep. 2019, 38, 1001–1012. [Google Scholar]
- Shi, H.; Reiter, R.J.; Tan, D.X.; Chan, Z. INDOLE-3-ACETIC ACID INDUCIBLE 17 positively modulates natural leaf senescence through melatonin-mediated pathway in Arabidopsis. J. Pineal Res. 2015, 58, 26–33. [Google Scholar]
- Arnao, M.B.; Hernández-Ruiz, J. Protective effect of melatonin against chlorophyll degradation during the senescence of barley leaves. J. Pineal Res. 2009, 46, 58–63. [Google Scholar]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin, plant growth regulator and/or biostimulator during stress? Trend Plant Sci. 2014, 19, 789–797. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Functions of melatonin in plants, a review. J. Pineal Res. 2015, 59, 133–150. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Sun, Q.; Li, H.; Li, X.; Cao, Y.; Zhang, H.; Guo, Y.D. Melatonin improved anthocyanin accumulation by regulating gene expressions and resulted in high reactive oxygen species scavenging capacity in cabbage. Front. Plant Sci. 2016, 7, 197. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.Y.; Xu, S.S.; Mu, D.; Sadeghnezhad, E.; Li, Q.; Ma, Z.H.; Zhao, L.X.; Zhang, Q.D.; Wang, L.X. Exogenous Melatonin Delays Dark-Induced Grape Leaf Senescence by Regulation of Antioxidant System and Senescence Associated Genes (SAGs). Plants 2019, 8, 366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, B.; Shi, Q. Review of Melatonin in Horticultural Crops. Sci. Agric. Sin. 2017, 50, 2326–2337. [Google Scholar]
- Tan, X.L.; Fan, Z.Q.; Kuang, J.F.; Lu, W.J.; Reiter, R.J.; Lakshmanan, P.; Su, X.G.; Zhou, J.; Chen, J.Y.; Shan, W. Melatonin delays leaf senescence of Chinese flowering cabbage by suppressing ABFs-mediated abscisic acid biosynthesis and chlorophyll degradation. J. Pineal Res. 2019, 67, e12570. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.Q.; Fu, X.; Han, L.J.; Xu, C.X.; Liu, C.Y.; Bi, H.G.; Ai, X.Z. Nitric Oxide Functions as a Downstream Signal for Melatonin-Induced Cold Tolerance in Cucumber Seedlings. Front. Plant Sci. 2021, 12, 686545. [Google Scholar] [CrossRef]
- Liang, C.Z.; Zheng, G.Y.; Li, W.Z.; Wang, Y.; Hu, B.; Wang, H.; Wu, H.; Qian, Y.; Zhu, X.-G.; Tan, D.-X.; et al. Melatonin delays leaf senescence and enhances salt stress tolerance in rice. J. Pineal Res. 2015, 59, 91–101. [Google Scholar] [CrossRef]
- Park, W.J. Melatonin as an endogenous plant regulatory signal: Debates and perspectives. J. Pineal Res. 2011, 54, 143–149. [Google Scholar] [CrossRef]
- Xu, W.; Cai, S.Y.; Zhang, Y.; Wang, Y.; Ahammed, G.J.; Xia, X.J.; Shi, K.; Zhou, Y.H.; Yu, J.Q.; Reiter, R.J.; et al. Melatonin enhances thermotolerance by promoting cellular protein protection in tomato plants. J. Pineal Res. 2016, 61, 457–469. [Google Scholar] [CrossRef]
- Li, M.Q.; Hasan, M.K.; Li, C.X.; Ahammed, G.J.; Xia, X.J.; Shi, K.; Zhou, Y.H.; Reiter, R.J.; Yu, J.Q.; Xu, M.X.; et al. Melatonin mediates selenium-induced tolerance to cadmium stress in tomato plants. J. Pineal Res. 2016, 61, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Sun, S.; Zhao, N.; Yang, W.; Shi, Q.; Gong, B. COMT1 overexpression resulting in increased melatonin biosynthesis contributes to the alleviation of carbendazim phytotoxicity and residues in tomato plants. Environ. Pollut. 2019, 252, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Weaver, L.M.; Gan, S.; Quirino, B.; Amasino, R.M. A comparison of the expression patterns of several senescence-associated genes in response to stress and hormone treatment. Plant Mol. Biol. 1998, 37, 455–469. [Google Scholar] [CrossRef]
- Yu, J.; Zhang, Y.; Di, C.; Zhang, Q.; Zhang, K.; Wang, C.; You, Q.; Yan, H.; Dai, S.Y.; Yuan, J.S.; et al. JAZ7 negatively regulates dark-induced leaf senescence in Arabidopsis. J. Exp. Bot. 2016, 67, 751–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Liu, C.; Guo, Y. Wheat Transcription Factor TaSNAC11-4B Positively Regulates Leaf Senescence through Promoting ROS Production in Transgenic Arabidopsis. Int. J. Mol. Sci. 2020, 21, 7672. [Google Scholar] [CrossRef]
- Niu, F.; Cui, X.; Zhao, P.; Sun, M.; Yang, B.; Deyholos, M.K.; Li, Y.; Zhao, X.; Jiang, Y. WRKY42 transcription factor positively regulates leaf senescence through modulating SA and ROS synthesis in Arabidopsis thaliana. Plant J. 2020, 104, 171–184. [Google Scholar] [CrossRef]
- Asad, M.A.U.; Wang, F.B.; Ye, Y.; Guan, X.Y.; Zhou, L.J.; Han, Z.Y.; Pan, G.; Cheng, F.M. Contribution of ABA metabolism and ROS generation to sugar starvation-induced senescence of rice leaves. Plant Growth Regul. 2021, 95, 241–257. [Google Scholar] [CrossRef]
- Ribeiro, C.W.; Korbes, A.P.; Garighan, J.A.; Jardim-Messeder, D.; Carvalho, F.E.; Sousa, R.H.; Caverzan, A.; Teixeira, F.K.; Silveira, J.A.G.; Margis-Pinheiro, M. Rice peroxisomal ascorbate peroxidase knockdown affects ros signaling and triggers early leaf senescence. Plant Sci. 2017, 263, 55–65. [Google Scholar] [CrossRef]
- Choudhary, A.; Kumar, A.; Kaur, N. ROS and oxidative burst: Roots in plant development. Plant Divers. 2020, 42, 33–43. [Google Scholar] [CrossRef]
- Zhao, Y.-Q.; Zhang, Z.-W.; Chen, Y.-E.; Ding, C.-B.; Yuan, S.; Reiter, R.J.; Yuan, M. Melatonin: A potential agent in delaying leaf senescence. Crit. Rev. Plant Sci. 2021, 40, 1–22. [Google Scholar] [CrossRef]
- Krasensky, J.; Jonak, C. Drought, salt and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maricruz, R.; Donald, J.H.; Eduardo, V.C. Abiotic stress triggers ROS-mediated programmed cell death in banana (Musa sp., AAA group, Cavendish sub-group) fruit. Sci. Hortic. 2022, 293, 110748. [Google Scholar]
- Chen, Y.; Yamori, W.; Tanaka, A.; Tanaka, R.; Ito, H. Degradation of the photosystem II core complex is independent of chlorophyll degradation mediated by Stay-Green Mg2+ dechelatase in Arabidopsis. Plant Sci. 2021, 307, 110–902. [Google Scholar] [CrossRef] [PubMed]
- Takamiya, K.I.; Tsuchiya, T.; Ohta, H. Degradation pathway(s) of chlorophyll, what has gene cloning revealed? J. Trends Plant Sci. 2000, 5, 426–431. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, T.Q.; Ren, G.D.; Hörtensteiner, S.; Zhou, Y.M.; Cahoon, E.B.; Zhang, C.Y. Chlorophyll degradation, the tocopherol biosynthesis-related phytol hydrolase in Arabidopsis seeds is still missing. Plant Physiol. 2014, 166, 70–79. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Li, H.X.; Zeng, H.L.; Cai, Q.S.; Zhou, X.; Yin, C.X. Exogenous jasmonic acid and cytokinin antagonistically regulate rice flag leaf senescence by mediating chlorophyll degradation, membrane deterioration, and senescence-associated genes expression. J. Plant Growth Regul. 2016, 35, 366–376. [Google Scholar] [CrossRef]
- Yu, G.H.; Xie, Z.N.; Zhang, J.; Lei, S.S.; Lin, W.J.; Xu, B.; Huang, B.R. NOL-mediated functional stay-green traits in perennial ryegrass (Lolium perenne L.) involving multifaceted molecular factors and metabolic pathways regulating leaf senescence. Plant J. 2021, 106, 1219–1232. [Google Scholar] [CrossRef]
- Rossi, S.; Chapman, C.; Huang, B. Suppression of heat-induced leaf senescence by γ-aminobutyric acid, proline, and ammonium nitrate through regulation of chlorophyll degradation in creeping bentgrass. Environ. Exp. Bot. 2020, 177, 104–116. [Google Scholar] [CrossRef]
- Zhang, J.; Li, H.; Xu, B.; Li, J.; Huang, B. Exogenous Melatonin Suppresses Dark-Induced Leaf Senescence by Activating the Superoxide Dismutase-Catalase Antioxidant Pathway and Down-Regulating Chlorophyll Degradation in Excised Leaves of Perennial Ryegrass (Lolium perenne L.). Front. Plant Sci. 2016, 7, 1500. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zhang, T.; Ding, F. Exogenous Melatonin Delays Methyl Jasmonate-Triggered Senescence in Tomato Leaves. Agronomy 2019, 9, 795. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Li, R.; Ge, J.; Liu, J.; Wang, W.; Xu, M.; Zhang, R.; Hussain, S.; Wei, H.; Dai, Q. Exogenous melatonin confers enhanced salinity tolerance in rice by blocking the ROS burst and improving Na+/K+ homeostasis. Environ. Exp. Bot. 2021, 189, 104–530. [Google Scholar] [CrossRef]
- Fan, J.; Lou, Y.; Shi, H.; Chen, L.; Cao, L. Transcriptomic analysis of dark-induced senescence in bermudagrass (Cynodon dactylon). Plants 2019, 8, 614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernd, M.-R.; Salma, B. Auxin and Its Role in Plant Senescence. J. Plant Growth Regul. 2014, 33, 21–33. [Google Scholar]
- Goren-Saglam, N.; Harrison, E.; Breeze, E.; Oz, G.; Buchanan-Wollaston, V. Analysis of the impact of indole-3-acetic acid (IAA) on gene expression during leaf senescence in Arabidopsis thaliana. Physiol. Mol. Biol. Plants 2020, 26, 733–745. [Google Scholar] [CrossRef]
- Feng, G.; Xu, Q.; Wang, Z.; Zhuoma, Q. AINTEGUMENTA negatively regulates age-dependent leaf senescence downstream of AUXIN RESPONSE FACTOR 2 in Arabidopsis thaliana. Plant Biotechnol. 2016, 33, 71–76. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.C.; Hong, S.W.; Whang, S.S.; Lim, P.O.; Nam, H.G.; Koo, J.C. Age-Dependent Action of an ABA-Inducible Receptor Kinase, RPK1, as a Positive Regulator of Senescence in Arabidopsis Leaves. Plant Cell Physiol. 2011, 52, 651–662. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Chan, Z.; Gao, J.; Xing, L.; Cao, M.; Yu, C.; Hu, Y.; You, J.; Shi, H.; Zhu, Y.; et al. ABA receptor PYL9 promotes drought resistance and leaf senescence. Proc. Natl. Acad. Sci. USA 2016, 113, 1949–1954. [Google Scholar] [CrossRef] [Green Version]
- Cha, J.-Y.; Kim, M.R.; Jung, I.J.; Kang, S.B.; Park, H.J.; Kim, M.G.; Dae-Jin, Y.; Kim, W.-Y. The thiol reductase activity of YUCCA6 mediates delayed leaf senescence by regulating genes involved in auxin redistribution. Front. Plant Sci. 2016, 7, 626. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.N.; Yu, Y.; Zhao, J.Y.; Cui, N.; Song, T.F.; Yang, Y.; Fan, H.Y. The two translationally controlled tumor protein genes, CsTCTP1 and CsTCTP2, are negative modulators in the Cucumis sativus defense response to Sphaerotheca fuliginea. Front. Plant Sci. 2018, 9, 544. [Google Scholar] [CrossRef] [Green Version]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Zhao, S.J.; Shi, G.A.; Dong, X.C. Experimental Technique of Plant Physiology; Agricultural Science and Technology Press of China: Beijing, China, 2002; pp. 55–57. [Google Scholar]
- Wang, A.G. Quantitative relation between the reaction of hydroxylamine and superoxide anion radicals in plants. Plant Physiol. Commun. 1990, 26, 55–57. [Google Scholar]
- Liu, F.; Zhang, X.; Cai, B.; Pan, D.; Fu, X.; Bi, H.; Ai, X. Physiological response and transcription profiling analysis reveal the role of glutathione in H2S-induced chilling stress tolerance of cucumber seedlings. Plant Sci. 2020, 291, 110363. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Feng, Y.; Jing, T.; Liu, X.; Ai, X.; Bi, H. Melatonin Promotes the Chilling Tolerance of Cucumber Seedlings by Regulating Antioxidant System and Relieving Photoinhibition. Front. Plant Sci. 2021, 12, 789617. [Google Scholar] [CrossRef]
- Jabs, T.; Dietrich, R.A.; Dangl, J.L. Initiation of runaway cell death in an Arabidopsis mutant by extracellular superoxide. Science 1996, 27, 1853–1856. [Google Scholar] [CrossRef]
- Thordal-Christensen, H.; Zhang, Z.; Wei, Y.; Collinge, D.B. Subcellular localization of H2O2 in plants: H2O2 accumulation in papillae and hypersensitive response during the barley-powdery mildew interaction. Plant J. 1997, 11, 1187–1194. [Google Scholar] [CrossRef]
- Beyer, W.F.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A.C. Assay of catalase and peroxidases. Methods Enzym. 1995, 2, 764–775. [Google Scholar]
- Nakano, Y.; Asada, K. Purification of ascorbate peroxidase in spinach chloroplasts; its inactivation in ascorbate-depleted medium and reactivation by monodehydroascorbate radical. Plant Cell Physiol. 1987, 28, 131–140. [Google Scholar]
- Li, H.S.; Sun, Q.; Zhao, S.J.; Zhang, W.H. Experimental Principles and Techniques of Plant Physiology and Biochemistry; Higher Education Press: Beijing, China, 2000. [Google Scholar]
- Li, Y.-M.; He, X.-R.; Li, Q.-M.; Liu, B.-B.; Li, S.-H.; Ai, X.-Z.; Wei, M.; Zhang, D.-L. Effect of CO2 enrichment on antioxidant system in cucumber seedling root system under drought stress. Plant Physiol. J. 2019, 55, 1011–1019. [Google Scholar]
- Yin, Z.C.; Chen, J.; Zeng, L.R.; Goh, M.; Leung, H.; Khush, G.S.; Wang, G.L. Characterizing rice lesion mimic mutants and identifying a mutant with broad-spectrum resistance to rice blast and bacterial blight. Mol. Plant-Microbe Interact. 2000, 13, 869–876. [Google Scholar] [CrossRef] [Green Version]
- Bian, F.; Xiao, Q.; Hao, G.; Sun, Y.; Lu, W.; Zhai, H. Effect of root-applied melatonin on endogenous melatonin and chlorophyll fluorescence characteristics in grapevine under NaCl stress. Sci. Agric. Sin. 2018, 51, 952–963. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Accession Numbers | Primer Pairs (5′–3′) |
|---|---|---|
| ASMT | XM_004144879 | ATTGGAAGTTTAGTTGATGTGGGA |
| AGCATCAGCCTTGGGAATGGAAT | ||
| TDC | XM_004135488 | ATAAATGGTTCTTCTCGGCGCCAG |
| GTTAATCATATTCGACTTCTGGT | ||
| T5H | XM_004140201 | AGCTTGTGCAGGCTACCAACT |
| GAACGTTGGAACAAACTTGTG | ||
| SNAT | XM_011655429 | AGTCCCCTGTTTCAGAGGAGAAT |
| AGATTCCGATAAAACTCTACCAC | ||
| SAG20 | XM_004149882 | CAGACCTGGAGTGGTGGTTC |
| GCCGGAGATCTGTCACAACA | ||
| PDCD | XM_011661791 | AGATGATGATGACGACGATG- |
| CAGCCTTGCTTGGAAATAG | ||
| PAO | XM_031884976 | GGGCATTGAAAACTGGAAGA |
| TTACTTGGCGATCAAAAATGG | ||
| PPH | XM_011661125 | GCAATGTGACGCCCTTAACT |
| CATCGAACAGGTCATTGGTG | ||
| SOD | XM_011660217 | GGAAAGATGTGAAGGCTGTGG |
| GCACCATGTTGTTTTCCAGCAG | ||
| CAT | XM_001308916 | AATGGCCGGAGGATGTGA |
| CCAACGACATAGAGAAAGCCAAG | ||
| APX | XM_001280706 | GTGCTACCCTGTTGTGAGTG |
| AACAGCGATGTCAAGGCCAT | ||
| ARF1 | XM_011656213 | CCAGATCCTCCCCTTCCTGA |
| GTCATCCGCATGCCTCCTAA | ||
| YUCCA6 | XM_004150231 | GGGACACTGCAAGATTCGGA |
| GCTTGACGTTTCAGCCGTTT | ||
| NCED | XM_004147720 | TGGTGAACCGAAATCTACTTG |
| CGAAGGCTAAGATGTGGC | ||
| ABI5 | XM_011651278 | GGAATTGCTTTTCAGCGGCA |
| ACTCCATTGGCATTCAGCGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jing, T.; Liu, K.; Wang, Y.; Ai, X.; Bi, H. Melatonin Positively Regulates Both Dark- and Age-Induced Leaf Senescence by Reducing ROS Accumulation and Modulating Abscisic Acid and Auxin Biosynthesis in Cucumber Plants. Int. J. Mol. Sci. 2022, 23, 3576. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23073576
Jing T, Liu K, Wang Y, Ai X, Bi H. Melatonin Positively Regulates Both Dark- and Age-Induced Leaf Senescence by Reducing ROS Accumulation and Modulating Abscisic Acid and Auxin Biosynthesis in Cucumber Plants. International Journal of Molecular Sciences. 2022; 23(7):3576. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23073576
Chicago/Turabian StyleJing, Tongtong, Kun Liu, Yanan Wang, Xizhen Ai, and Huangai Bi. 2022. "Melatonin Positively Regulates Both Dark- and Age-Induced Leaf Senescence by Reducing ROS Accumulation and Modulating Abscisic Acid and Auxin Biosynthesis in Cucumber Plants" International Journal of Molecular Sciences 23, no. 7: 3576. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23073576