
Identification, Isolation, and Characterization of Melanocyte Precursor Cells in the Human Limbal Stroma

 and
and
Abstract
:1. Introduction
2. Results
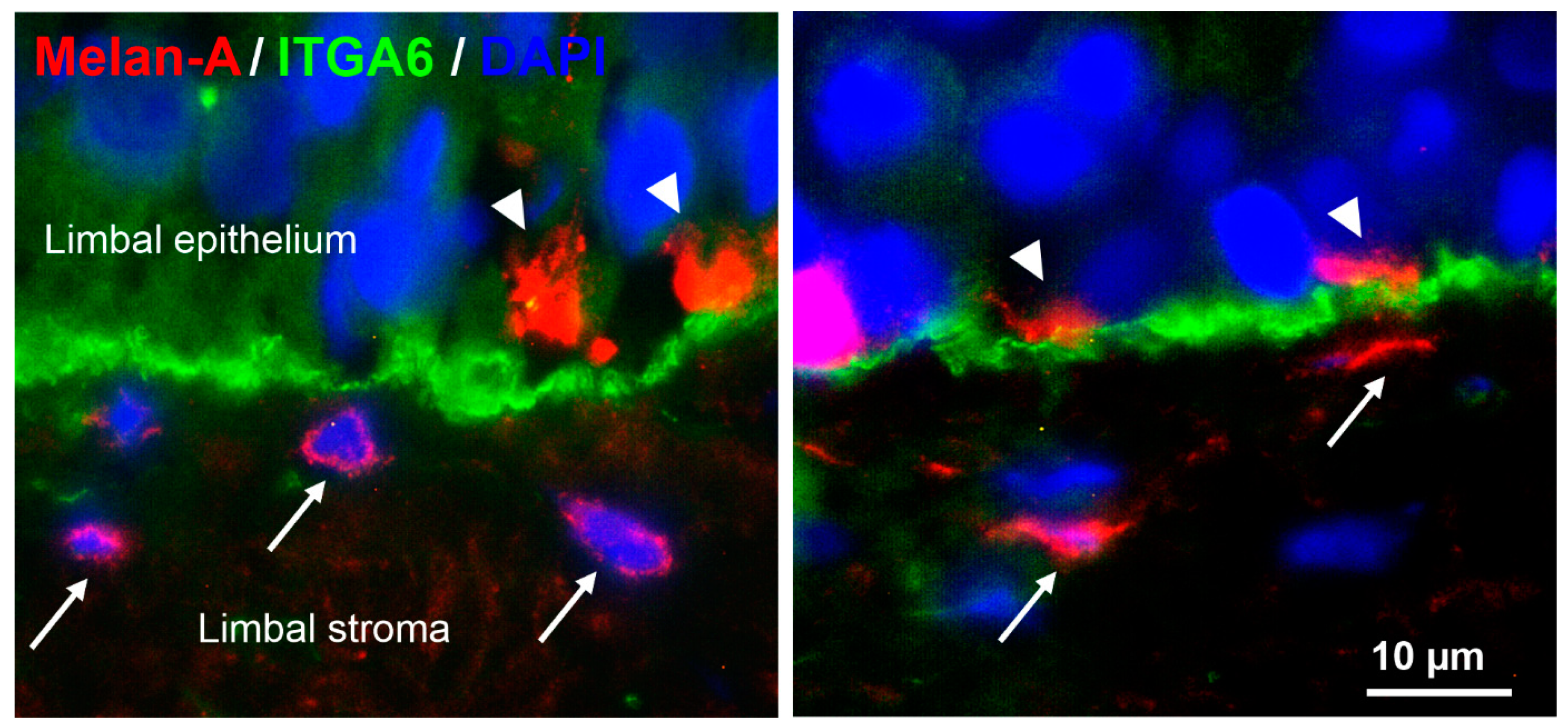
2.1. Localization of Two Melanocyte Populations at the Human Limbus In Situ
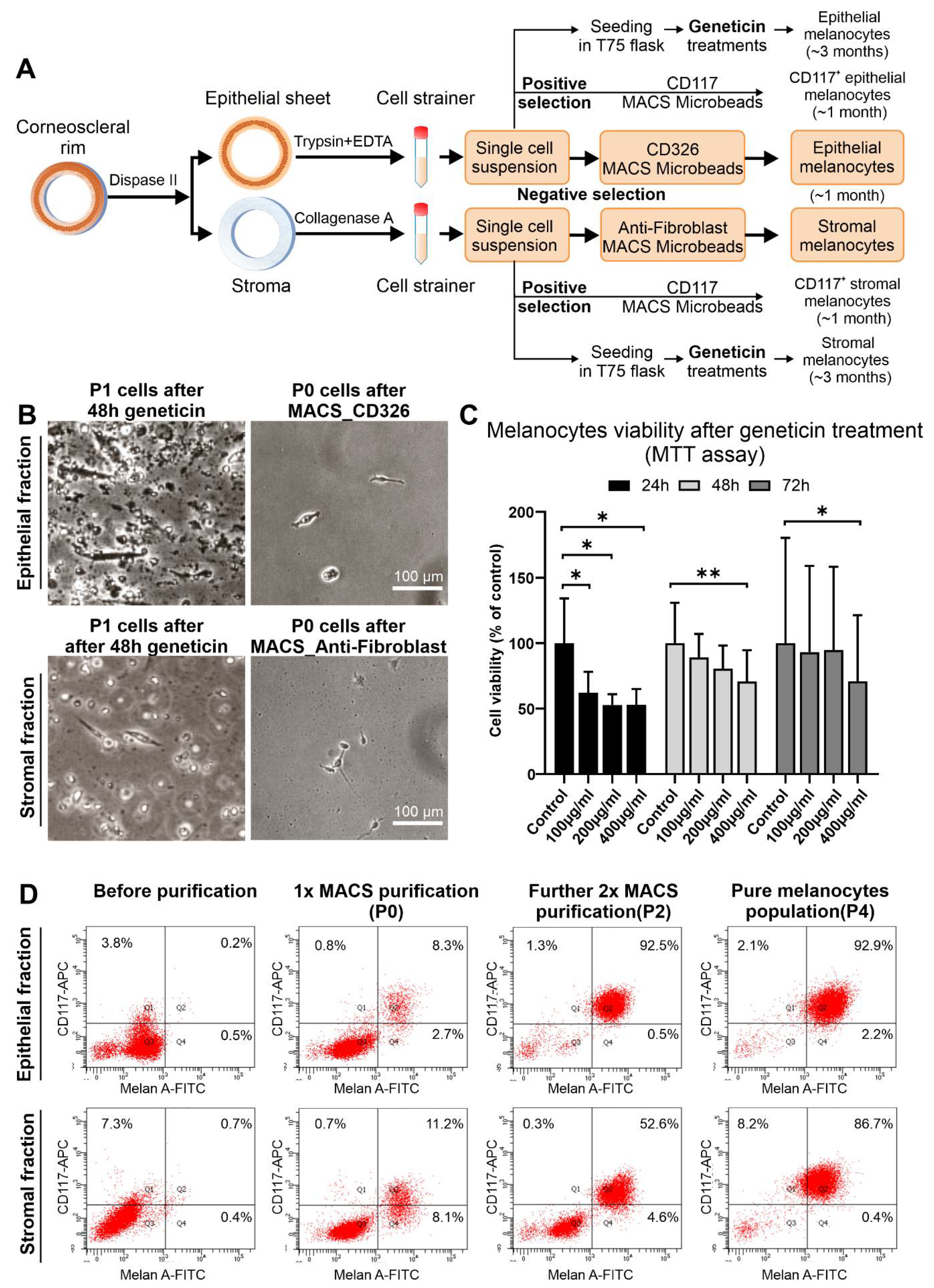
2.2. Isolation and Enrichment of Limbal Epithelial and Stromal Melanocytes
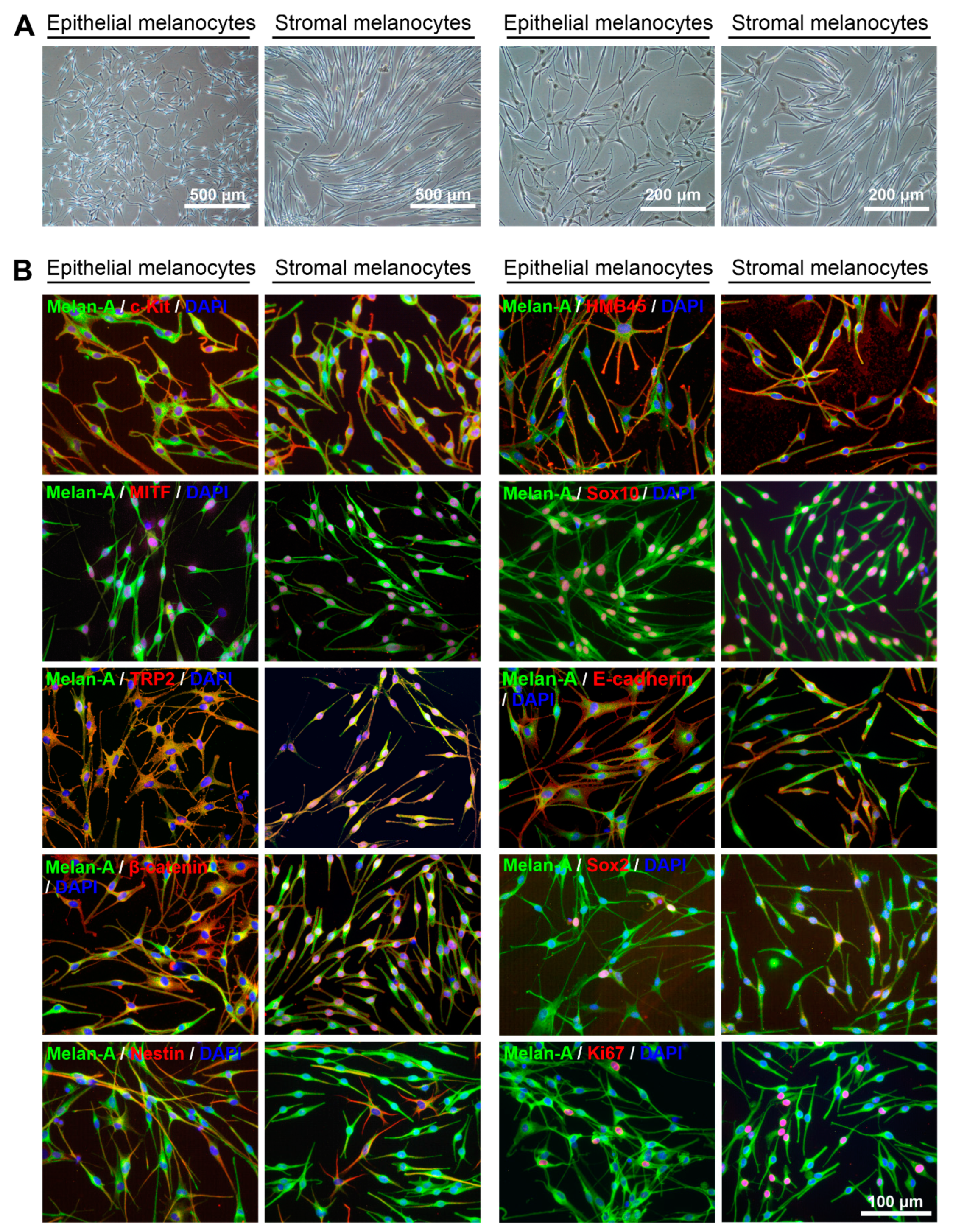
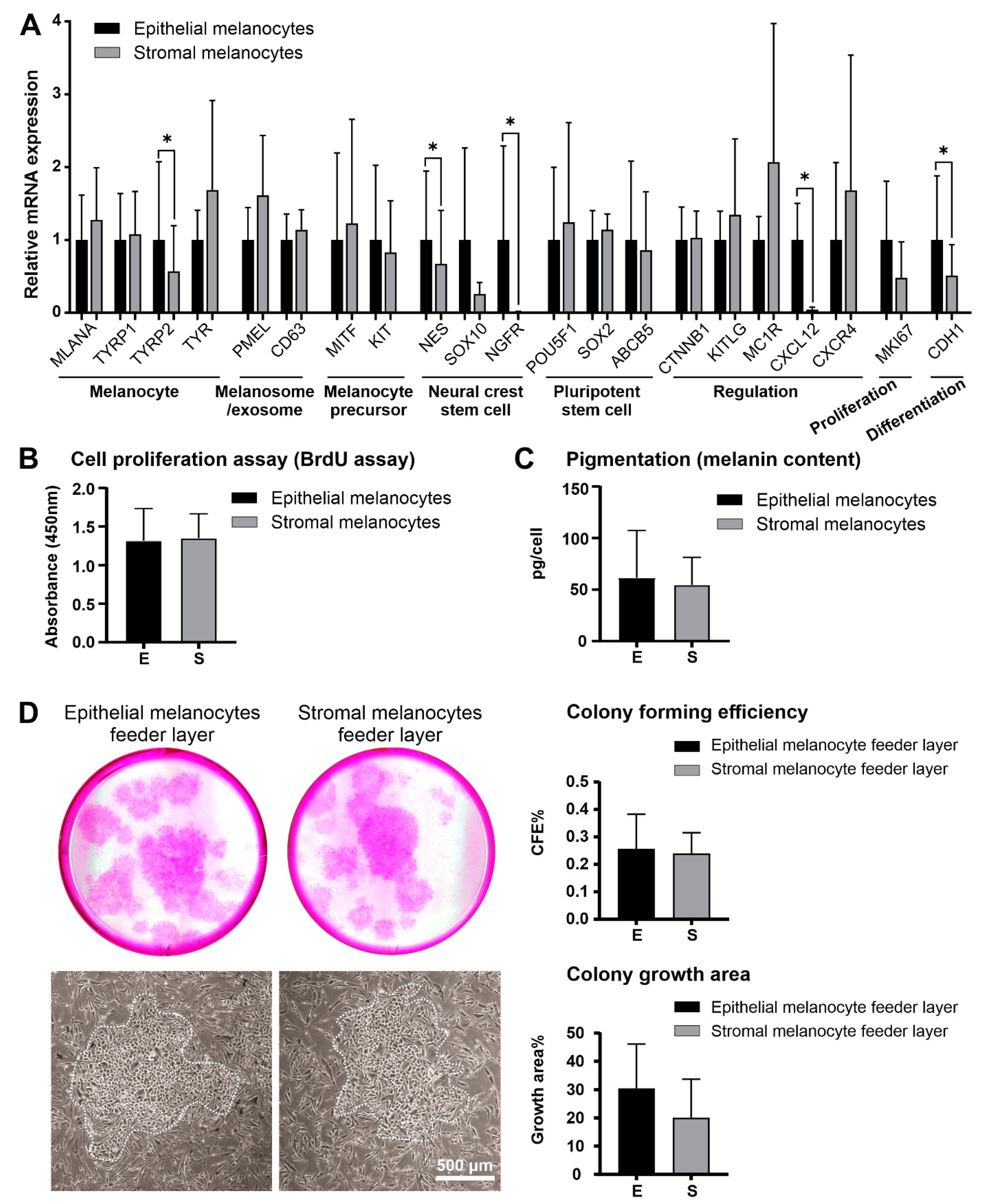
2.3. Comparative Characterization of Limbal Epithelial and Stromal Melanocytes
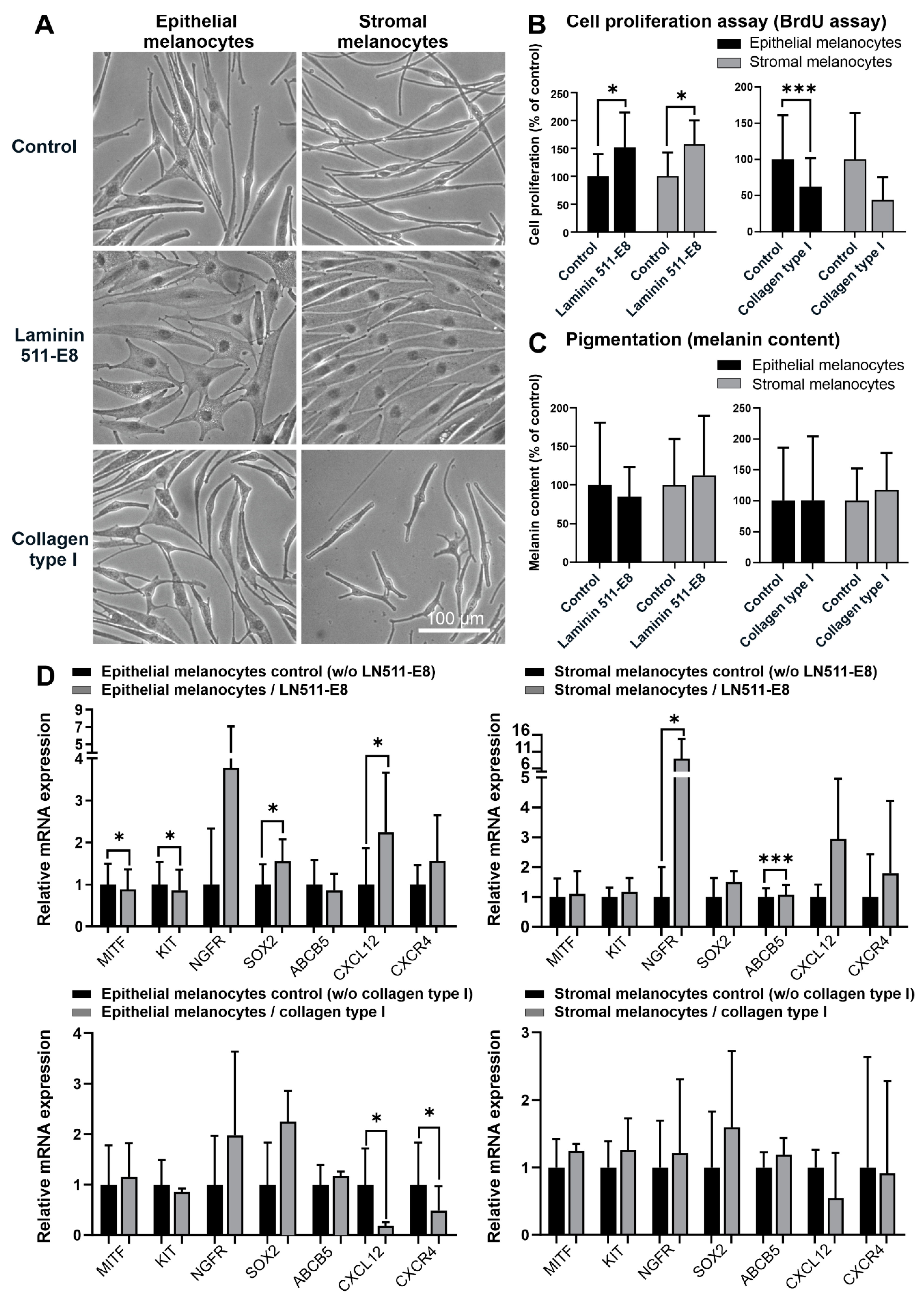
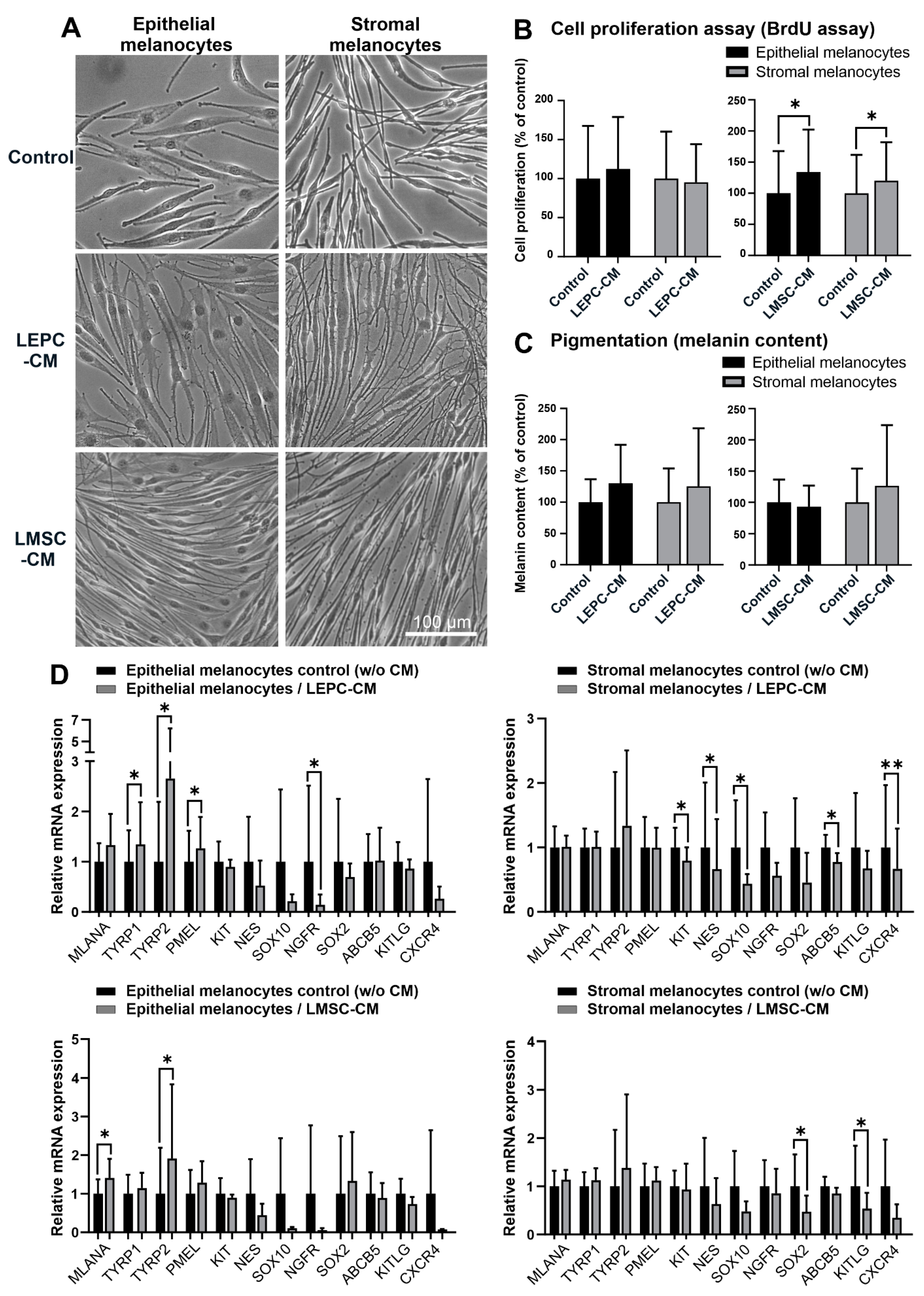
2.4. Influence of Limbal Niche Factors on Melanocyte Phenotype
3. Discussion
4. Materials and Methods
4.1. Human Tissues and Study Approval
4.2. Melanocyte Isolation by Magnetic-Activated Cell Sorting (MACS)
4.3. Melanocyte Isolation by Geneticin Treatment
4.4. Melanocyte Cultivation on Extracellular Matrix and in Conditioned Media
4.5. Cell Viability Assay (MTT)
4.6. Flow Cytometry
4.7. Immunohistochemistry and Immunocytochemistry
4.8. Quantitative RT-PCR
4.9. Cell Proliferation Assay
4.10. Pigmentation Assay
4.11. Colony Forming Efficiency Assay
4.12. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bonnet, C.; González, S.; Roberts, J.S.; Robertson, S.Y.; Ruiz, M.; Zheng, J.; Deng, S.X. Human limbal epithelial stem cell regulation, bioengineering and function. Prog. Retin. Eye Res. 2021, 85, 100956. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, G.; Sasamoto, Y.; Ksander, B.R.; Frank, M.; Frank, N.Y. Limbal stem cells: Identity, developmental origin, and therapeutic potential. Wiley Interdiscip. Rev. Dev. Biol. 2017, 7, e303. [Google Scholar] [CrossRef]
- Shortt, A.J.; Secker, G.A.; Munro, P.M.; Khaw, P.T.; Tuft, S.J.; Daniels, J.T. Characterization of the Limbal Epithelial Stem Cell Niche: Novel Imaging Techniques Permit In Vivo Observation and Targeted Biopsy of Limbal Epithelial Stem Cells. Stem Cells 2007, 25, 1402–1409. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Hayashida, Y.; Chen, Y.-T.; Tseng, S.C.G. Niche regulation of corneal epithelial stem cells at the limbus. Cell Res. 2007, 17, 26–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ordonez, P.; Di Girolamo, N. Limbal Epithelial Stem Cells: Role of the Niche Microenvironment. Stem Cells 2012, 30, 100–107. [Google Scholar] [CrossRef]
- Mei, H.; Gonzalez, S.; Deng, S.X. Extracellular Matrix is an Important Component of Limbal Stem Cell Niche. J. Funct. Biomater. 2012, 3, 879–894. [Google Scholar] [CrossRef] [Green Version]
- Dziasko, M.A.; Daniels, J.T. Anatomical Features and Cell-Cell Interactions in the Human Limbal Epithelial Stem Cell Niche. Ocul. Surf. 2016, 14, 322–330. [Google Scholar] [CrossRef]
- Polisetti, N.; Zenkel, M.; Menzel-Severing, J.; Kruse, F.E.; Schlötzer-Schrehardt, U. Cell adhesion mole-cules and stem cell-niche-interactions in the limbal stem cell niche. Stem Cells 2016, 34, 203–219. [Google Scholar] [CrossRef]
- Higa, K.; Shimmura, S.; Miyashita, H.; Shimazaki, J.; Tsubota, K. Melanocytes in the corneal limbus in-teract with K19-positive basal epithelial cells. Exp. Eye Res. 2005, 81, 218–223. [Google Scholar] [CrossRef]
- Shimmura, S.; Kawakita, T. Accessory Cell Populations in the Cornea. Ocul. Surf. 2006, 4, 74–80. [Google Scholar] [CrossRef]
- Hayashi, R.; Yamato, M.; Sugiyama, H.; Sumide, T.; Yang, J.; Okano, T.; Tano, Y.; Nishida, K. N-Cadherin Is Expressed by Putative Stem/Progenitor Cells and Melanocytes in the Human Limbal Epithelial Stem Cell Niche. Stem Cells 2006, 25, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Rohrbach, J.M.; Süsskind, D.; Grüb, M. Der Melanozyt und das Auge: Eine Übersicht unter besonderer Berücksichtigung der Kornea [The melanocyte and the eye: A review with special emphasis on the cornea]. Klin. Monbl. Augenheilkd. 2012, 229, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Polisetti, N.; Gießl, A.; Zenkel, M.; Heger, L.; Dudziak, D.; Naschberger, E.; Stich, L.; Steinkasserer, A.; Kruse, F.E.; Schlötzer-Schrehardt, U. Melanocytes as emerging key players in niche regulation of limbal epithelial stem cells. Ocul. Surf. 2021, 22, 172–189. [Google Scholar] [CrossRef] [PubMed]
- Dziasko, M.A.; Tuft, S.J.; Daniels, J.T. Limbal melanocytes support limbal epithelial stem cells in 2D and 3D microenvironments. Exp. Eye Res. 2015, 138, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Eisinger, M.; Marko, O. Selective proliferation of normal human melanocytes in vitro in the presence of phorbol ester and cholera toxin. Proc. Natl. Acad. Sci. USA 1982, 79, 2018–2022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, D.N.; McCormick, S.A.; Ritch, R.; Pelton-Henrion, K. Studies of human uveal melanocytes in vitro: Isolation, purification and cultivation of human uveal melanocytes. Investig. Ophthalmol. Vis. Sci. 1993, 34, 2210–2219. [Google Scholar]
- Hu, D.-N.; McCormick, S.A.; Seedor, J.A.; Ritterband, D.; Shah, M.K. Isolation, purification and cultivation of conjunctival melanocytes. Exp. Eye Res. 2007, 84, 655–662. [Google Scholar] [CrossRef]
- Hsu, M.-Y.; Li, L.; Herlyn, M. Cultivation of Normal Human Epidermal Melanocytes in the Absence of Phorbol Esters. Hum. Cell Cult. Protoc. 2005, 107, 13–28. [Google Scholar] [CrossRef]
- Halaban, R.; Alfano, F.D. Selective elimination of fibroblasts from cultures of normal human melanocytes. In Vitro 1984, 20, 447–450. [Google Scholar] [CrossRef]
- Horikawa, T.; Norris, D.A.; Zekman, T.; Morelli, J.G. Effective Elimination of Fibroblasts in Cultures of Melanocytes by Lowering Calcium Concentration in TPA Depleted Medium Following Geneticin Treatment. Pigment Cell Res. 1996, 9, 58–62. [Google Scholar] [CrossRef]
- Polisetti, N.; Gießl, A.; Li, S.; Sorokin, L.; Kruse, F.E.; Schlötzer-Schrehardt, U. Laminin-511-E8 promotes efficient in vitro expansion of human limbal melanocytes. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Willemsen, M.; Luiten, R.M.; Teunissen, M.B.M. Instant isolation of highly purified human melanocytes from freshly prepared epidermal cell suspensions. Pigment Cell Melanoma Res. 2020, 33, 763–766. [Google Scholar] [CrossRef]
- Polisetti, N.; Schlötzer-Schrehardt, U.; Reinhard, T.; Schlunck, G. Isolation and enrichment of melanocytes from human corneal limbus using CD117 (c-Kit) as selection marker. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Grützkau, A.; Radbruch, A. Small but mighty: How the MACS®-technology based on nanosized superparamagnetic particles has helped to analyze the immune system within the last 20 years. Cytom. Part A 2010, 77, 643–647. [Google Scholar] [CrossRef] [PubMed]
- Bacon, K.; Lavoie, A.; Rao, B.M.; Daniele, M.; Menegatti, S. Past, Present, and Future of Affinity-based Cell Separation Technologies. Acta Biomater. 2020, 112, 29–51. [Google Scholar] [CrossRef] [PubMed]
- Tobin, D.; Bystryn, J.-C. Different Populations of Melanocytes Are Present in Hair Follicles and Epidermis. Pigment Cell Res. 1996, 9, 304–310. [Google Scholar] [CrossRef]
- Nishimura, E.K. Melanocyte stem cells: A melanocyte reservoir in hair follicles for hair and skin pigmenta-tion. Pigment Cell Melanoma Res. 2011, 24, 401–410. [Google Scholar] [CrossRef]
- Hoerter, J.D.; Bradley, P.; Casillas, A.; Chambers, D.; Denholm, C.; Johnson, K.; Weiswasser, B. Extrafollicular Dermal Melanocyte Stem Cells and Melanoma. Stem Cells Int. 2012, 2012, 407079. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Fukunaga-Kalabis, M.; Yu, H.; Xu, X.; Kong, J.; Lee, J.T.; Herlyn, M. Human dermal stem cells differentiate into functional epidermal melanocytes. J. Cell Sci. 2010, 123, 853–860. [Google Scholar] [CrossRef] [Green Version]
- Yamada, T.; Hasegawa, S.; Inoue, Y.; Date, Y.; Yamamoto, N.; Mizutani, H.; Nakata, S.; Matsunaga, K.; Akamatsu, H. Wnt/β-Catenin and Kit Signaling Sequentially Regulate Melanocyte Stem Cell Differentiation in UVB-Induced Epidermal Pigmentation. J. Investig. Dermatol. 2013, 133, 2753–2762. [Google Scholar] [CrossRef] [Green Version]
- Pincelli, C. p75 Neurotrophin Receptor in the Skin: Beyond Its Neurotrophic Function. Front. Med. 2017, 4, 22. [Google Scholar] [CrossRef] [Green Version]
- Yaar, M.; Grossman, K.; Eller, M.; Gilchrest, B.A. Evidence for nerve growth factor-mediated paracrine effects in human epidermis. J. Cell Biol. 1991, 115, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.T.; Chen, S.Y.; Li, G.G.; Tseng, S.C. Limbal epithelial stem/progenitor cells attract stromal niche cells by SDF-1/CXCR4 signaling to prevent differentiation. Stem Cells 2011, 29, 1874–1885. [Google Scholar] [CrossRef]
- Rezk, A.F.; Kemp, D.M.; El-Domyati, M.; El-Din, W.H.; Lee, J.B.; Uitto, J.; Igoucheva, O.; Alexeev, V. Misbalanced CXCL12 and CCL5 Chemotactic Signals in Vitiligo Onset and Progression. J. Investig. Dermatol. 2017, 137, 1126–1134. [Google Scholar] [CrossRef] [Green Version]
- Yamada, T.; Hasegawa, S.; Hasebe, Y.; Hotta, M.; Arima, M.; Iwata, Y.; Kobayashi, T.; Numata, S.; Yamamoto, N.; Nakata, S.; et al. CXCL12 regulates differentiation of human immature melanocyte precursors as well as their migration. Arch. Dermatol. Res. 2018, 311, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Üstün, Y.; Reibetanz, M.; Brachvogel, B.; Nischt, R.; Eckes, B.; Zigrino, P.; Krieg, T. Dual role of laminin-511 in regulating melanocyte migration and differentiation. Matrix Biol. 2018, 80, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, P.R.; Ho, T.; Abdel-Malek, Z.A. Participation of keratinocyte- and fibroblast-derived factors in melanocyte homeostasis, the response to UV, and pigmentary disorders. Pigment Cell Melanoma Res. 2021, 34, 762–776. [Google Scholar] [CrossRef]
- Tian, X.; Liu, Z.; Niu, B.; Zhang, J.; Tan, T.K.; Lee, S.R.; Zhao, Y.; Harris, D.C.H.; Zheng, G. E-Cadherin/β-Catenin Complex and the Epithelial Barrier. J. Biomed. Biotechnol. 2011, 2011, 567305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, B.; Chen, S.-Y.; Zhu, Y.-T.; Tseng, S.C. Integration of BMP/Wnt signaling to control clonal growth of limbal epithelial progenitor cells by niche cells. Stem Cell Res. 2014, 12, 562–573. [Google Scholar] [CrossRef] [Green Version]
- Menzel-Severing, J.; Zenkel, M.; Polisetti, N.; Sock, E.; Wegner, M.; Kruse, F.E.; Schlötzer-Schrehardt, U. Transcription factor profiling identifies Sox9 as regulator of proliferation and differentiation in corneal epithelial stem/progenitor cells. Sci. Rep. 2018, 8, 1–18. [Google Scholar] [CrossRef]
- Polisetti, N.; Schmid, A.; Schlötzer-Schrehardt, U.; Maier, P.; Lang, S.J.; Steinberg, T.; Schlunck, G.; Reinhard, T. A decellularized human corneal scaffold for anterior corneal surface reconstruction. Sci. Rep. 2021, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Means, T.L.; Geroski, D.H.; L‘Hernault, N.; Grossniklaus, H.E.; Kim, T.; Edelhauser, H.F. The corneal epithelium after optisol-GS storage. Cornea 1996, 15, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Soni, N.G.; Hoover, C.K.; Da Silva, H.; Jeng, B.H. Preservation of the Corneal Epithelium in Different Corneal Storage Media. Cornea 2015, 34, 1400–1403. [Google Scholar] [CrossRef] [PubMed]
- Kureshi, A.K.; Funderburgh, J.L.; Daniels, J.T. Human Corneal Stromal Stem Cells Exhibit Survival Capacity Following Isolation From Stored Organ–Culture Corneas. Investig. Opthalmol. Vis. Sci. 2014, 55, 7583–7588. [Google Scholar] [CrossRef] [Green Version]
- Miyashita, H.; Niwano, H.; Yoshida, S.; Hatou, S.; Inagaki, E.; Tsubota, K.; Shimmura, S. Long-term homeostasis and wound healing in an in vitro epithelial stem cell niche model. Sci. Rep. 2017, 7, 43557. [Google Scholar] [CrossRef] [Green Version]
- Cario, M.; Taieb, A. Isolation and Culture of Epidermal Melanocytes. Ski. Tissue Eng. 2019, 1993, 33–46. [Google Scholar] [CrossRef]
- Grichnik, J.M.; Burch, J.A.; Burchette, J.; Shea, C.R. The SCF/KIT Pathway Plays a Critical Role in the Control of Normal Human Melanocyte Homeostasis. J. Investig. Dermatol. 1998, 111, 233–238. [Google Scholar] [CrossRef] [Green Version]
- Ordonez, P.; Chow, S.; Wakefield, D.; Di Girolamo, N. Human limbal epithelial progenitor cells express αvβ5-integrin and the interferon-inducible chemokine CXCL10/IP-10. Stem Cell Res. 2013, 11, 888–901. [Google Scholar] [CrossRef] [Green Version]
- Nakatsu, M.N.; Deng, S.X. Enrichment of Human Corneal Epithelial Stem/Progenitor Cells by Magnetic Bead Sorting Using SSEA4 as a Negative Marker. Corneal Regen. Med. 2013, 1014, 71–77. [Google Scholar] [CrossRef]
- Schlötzer-Schrehardt, U.; Latta, L.; Gießl, A.; Zenkel, M.; Fries, F.N.; Käsmann-Kellner, B.; Kruse, F.E.; Seitz, B. Dysfunction of the limbal epithelial stem cell niche in aniridia-associated keratopathy. Ocul. Surf. 2021, 21, 160–173. [Google Scholar] [CrossRef]
- Zenkel, M.; Krysta, A.; Pasutto, F.; Juenemann, A.; Kruse, F.E.; Schlötzer-Schrehardt, U. Regulation of Lysyl Oxidase-like 1 (LOXL1) and Elastin-Related Genes by Pathogenic Factors Associated with Pseudoexfoliation Syndrome. Investig. Opthalmol. Vis. Sci. 2011, 52, 8488–8495. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody (Clone) | Host Species | Source | Concentration | Application |
|---|---|---|---|---|
| Isotype control antibody, APC | mouse | Miltenyi Biotec | 1:50 | Flow cytometry analysis |
| Isotype control antibody, FITC | mouse | Miltenyi Biotec | 1:50 | Flow cytometry analysis |
| CD117 antibody, anti-human, APC | mouse | Miltenyi Biotec | 1:20 | Flow cytometry analysis |
| Melan-A/MART-1 antibody, FITC | mouse | Novus Biologicals | 1:133 | Flow cytometry analysis |
| Melan-A | rabbit | Abcam | 1:500 | Immunofluorescence |
| Melan-A (A103) | mouse | DAKO | 1:25 | Immunofluorescence |
| Integrin α6 (ITGA6) (GoH3) | rat | Millipore | 1:100 | Immunofluorescence |
| CD117/c-Kit (Ab81) | mouse | Cell signaling | 1:200 | Immunofluorescence |
| Sox10 (BC34) | mouse | Abcam | 1:50 | Immunofluorescence |
| TRP2/DCT | rabbit | Invitrogen | 1:100 | Immunofluorescence |
| β-catenin (15B8) | mouse | Thermo Fisher | 1:100 | Immunofluorescence |
| E-cadherin (36) | mouse | BD | 1:100 | Immunofluorescence |
| Nestin (10C2) | mouse | Abcam | 1:100 | Immunofluorescence |
| P75 NGFR (NGFR5) | mouse | Abcam | 1:100 | Immunofluorescence |
| Ki67 (MIB-1) | mouse | DAKO | 1:100 | Immunofluorescence |
| MITF (C5) | mouse | Abcam | 1:150 | Immunofluorescence |
| Sox2 | rabbit | Cell signaling | 1:100 | Immunofluorescence |
| HMB45 (HMB45) | mouse | DAKO | 1:50 | Immunofluorescence |
| TRP1 (TA99) | mouse | Abcam | 1:50 | Immunofluorescence |
| Gene Symbol | Accession Number | Sequence | |
|---|---|---|---|
| ABCB5 | NM_001163941.2 | 5 | CAGCAAGGGAAGCAAATGCGTA |
| 3 | ATCCTCTGTTTCTGCCCTCCAC | ||
| ACTB | NM_001101.3 | 5 | GCGTACAGGTCTTTGCGGATG |
| 3 | TGAGGCACTCTTCCAGCCTTC | ||
| CD63 | NM_001780.6 | 5 | TACGTCCTCCTGCTGGCCTTTT |
| 3 | ATGACCACTGGCAACAGAGAGC | ||
| CDH1 | NM_001317184.2 | 5 | CAGGATGGCTGAAGGTGACAGA |
| 3 | AGAGCACCTTCCATGACAGACC | ||
| CTNNB1 | NM_001904.4 | 5 | GGTCTGAGGAGCAGCTTCAGT |
| 3 | GGCCATGTCCAACTCCATCAAA | ||
| CXCL12 | NM_000609.7 | 5 | CATGCCGATTCTTCGAAAGCCA |
| 3 | CTGTTGTTGTTCTTCAGCCGGG | ||
| CXCR4 | NM_001008540.2 | 5 | GACCGCTTCTACCCCAATGACT |
| 3 | TACCAGGCAGGATAAGGCCAAC | ||
| GAPDH | NM_002046.2 | 5 | AAGGTCGGAGTCAACGGATTTGG |
| 3 | ATGACAAGCTTCCCGTTCTCAGC | ||
| KIT | NM_000222.3 | 5 | AAGCACAATGGCACGGTTGAAT |
| 3 | ACAGGGTGTGGGGATGGATTTG | ||
| KITLG | NM_003994.5 | 5 | ACCTTGTGGAGTGCGTGAAAGA |
| 3 | AGTAAAGAGCCTGGGTTCTGGG | ||
| MC1R | NM_002386.4 | 5 | CATCTCTGACGGGCTCTTCCTC |
| 3 | AGGCAGCAGATGAAGCAGTACA | ||
| MITF | NM_198159.3 | 5 | CGGGCTCTGTTCTCACTTTCCA |
| 3 | GAGCTTATCGGAGGCTTGGAGG | ||
| MKI67 | NM_002417.5 | 5 | TACGGATTATACCTGGCCTTCCC |
| 3 | ACAACAGGAAGCTGGATACGGA | ||
| MLANA | NM_005511.2 | 5 | TTACTGCTCATCGGCTGTTGGT |
| 3 | AGACACTTTGCTGTCCCGATGA | ||
| NES | NM_006617.2 | 5 | CAGCGTTGGAACAGAGGTTGGA |
| 3 | TCTGTAGGCCCTGTTTCTCCTG | ||
| NGFR | NM_002507.4 | 5 | CACCGACAACCTCATCCCTGT |
| 3 | GCTGTTGGCTCCTTGCTTGTTC | ||
| PMEL | NM_001200053.1 | 5 | GTCAGCACCCAGCTTATCATGC |
| 3 | CTGCTATGTGGCAACTGGGGTA | ||
| POU5F1 | NM_002701.6 | 5 | GGAGAAGGAGAAGCTGGAGCAA |
| 3 | GCAGATGGTCGTTTGGCTGAAT | ||
| SOX2 | NM_003106.4 | 5 | GAAGGATAAGTACACGCTGCCC |
| 3 | CGTTCATGTGCGCGTAACTGTC | ||
| SOX10 | NM_006941.4 | 5 | CATCCAGGCCCACTACAAGAGC |
| 3 | TCTGTCTTCGGGGTGGTTGGAG | ||
| TYR | NM_000372.5 | 5 | TCATCCAAAGATCTGGGCTATGAC |
| 3 | GACGACACAGCAAGCTCACA | ||
| TYRP1 | NM_000550.3 | 5 | GTCGCTCAGTGCTTGGAAGTTG |
| 3 | GTCATACTTTCCCGTGGGGTCA | ||
| TYRP2/DCT | NM_001129889.2 Variante 2 | 5 | GGACAAACGCTTTGCCACATTC |
| 3 | GAAGGGAGTTCCTTGGTCGCTT | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Zenkel, M.; Kruse, F.E.; Gießl, A.; Schlötzer-Schrehardt, U. Identification, Isolation, and Characterization of Melanocyte Precursor Cells in the Human Limbal Stroma. Int. J. Mol. Sci. 2022, 23, 3756. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23073756
Li S, Zenkel M, Kruse FE, Gießl A, Schlötzer-Schrehardt U. Identification, Isolation, and Characterization of Melanocyte Precursor Cells in the Human Limbal Stroma. International Journal of Molecular Sciences. 2022; 23(7):3756. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23073756
Chicago/Turabian StyleLi, Shen, Matthias Zenkel, Friedrich E. Kruse, Andreas Gießl, and Ursula Schlötzer-Schrehardt. 2022. "Identification, Isolation, and Characterization of Melanocyte Precursor Cells in the Human Limbal Stroma" International Journal of Molecular Sciences 23, no. 7: 3756. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23073756