
Plasma Proteomic Profile of Patients with Tick-Borne Encephalitis and Co-Infections

 , , ,
, , , 
Abstract
:1. Introduction
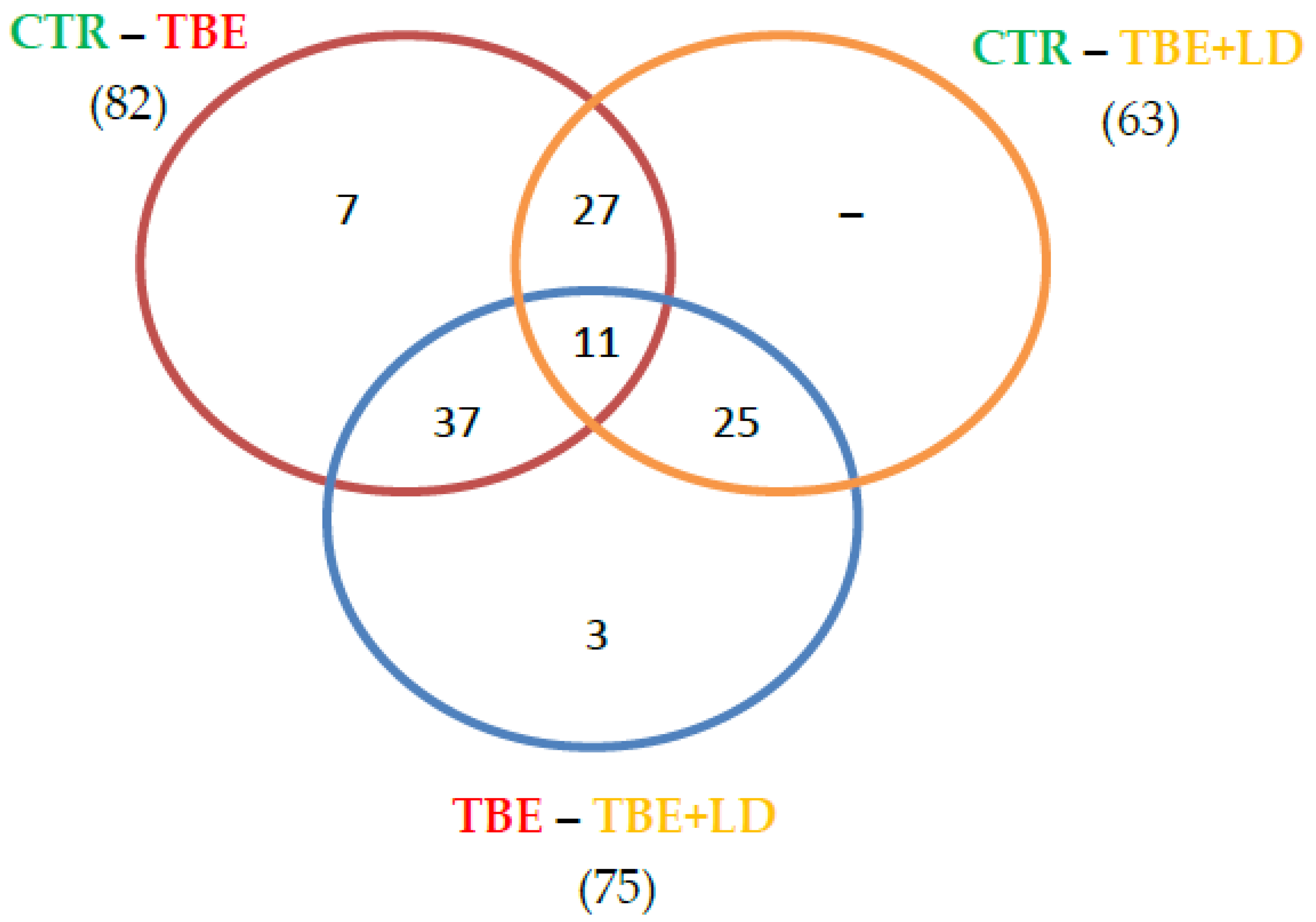
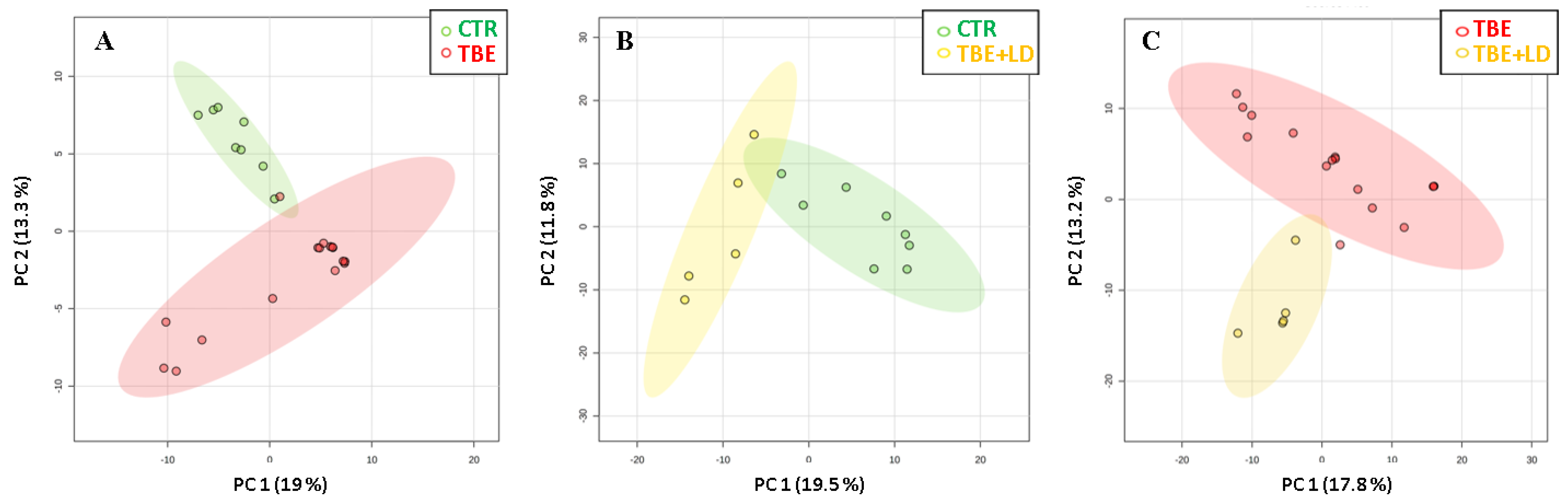
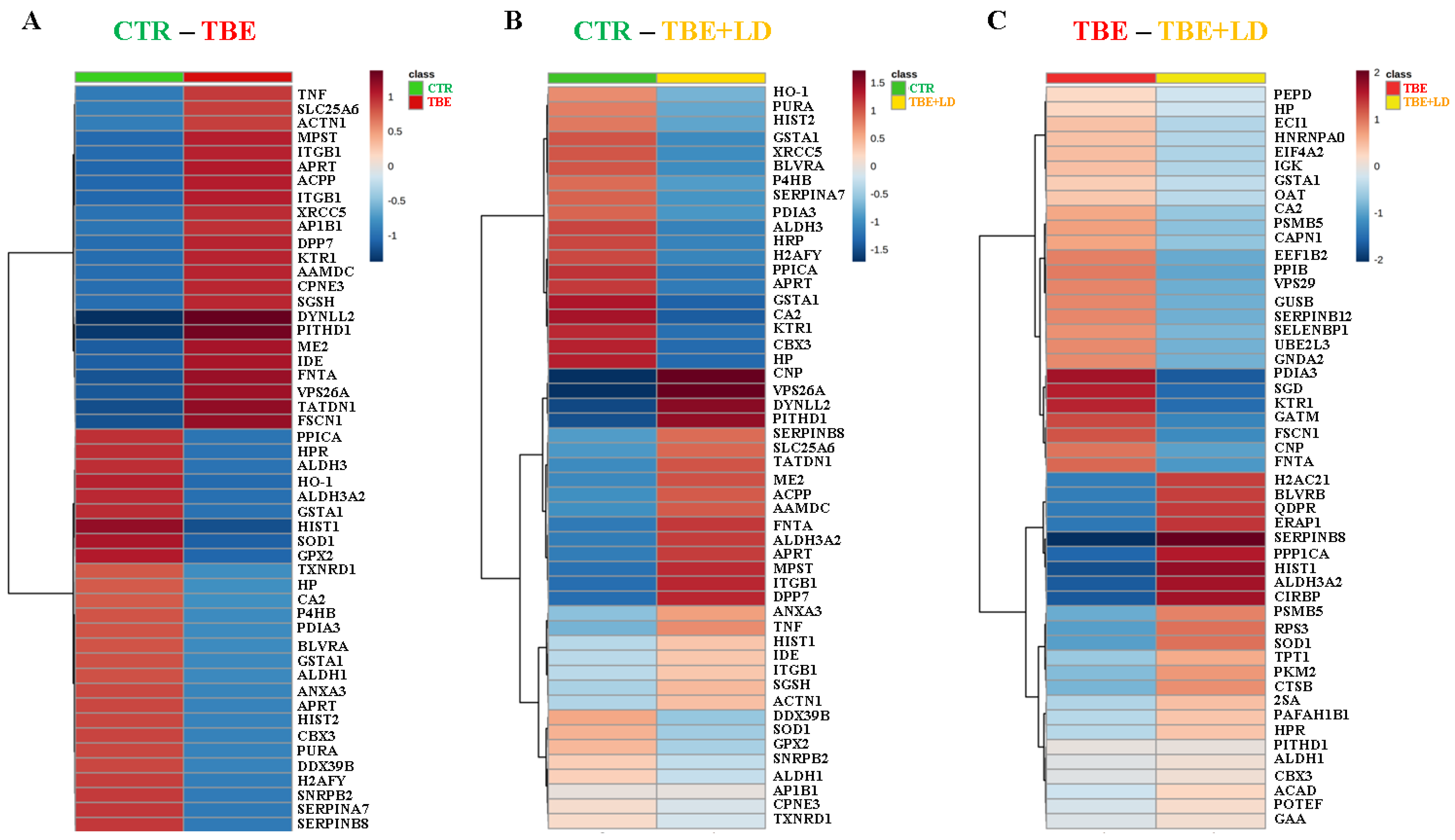
2. Results
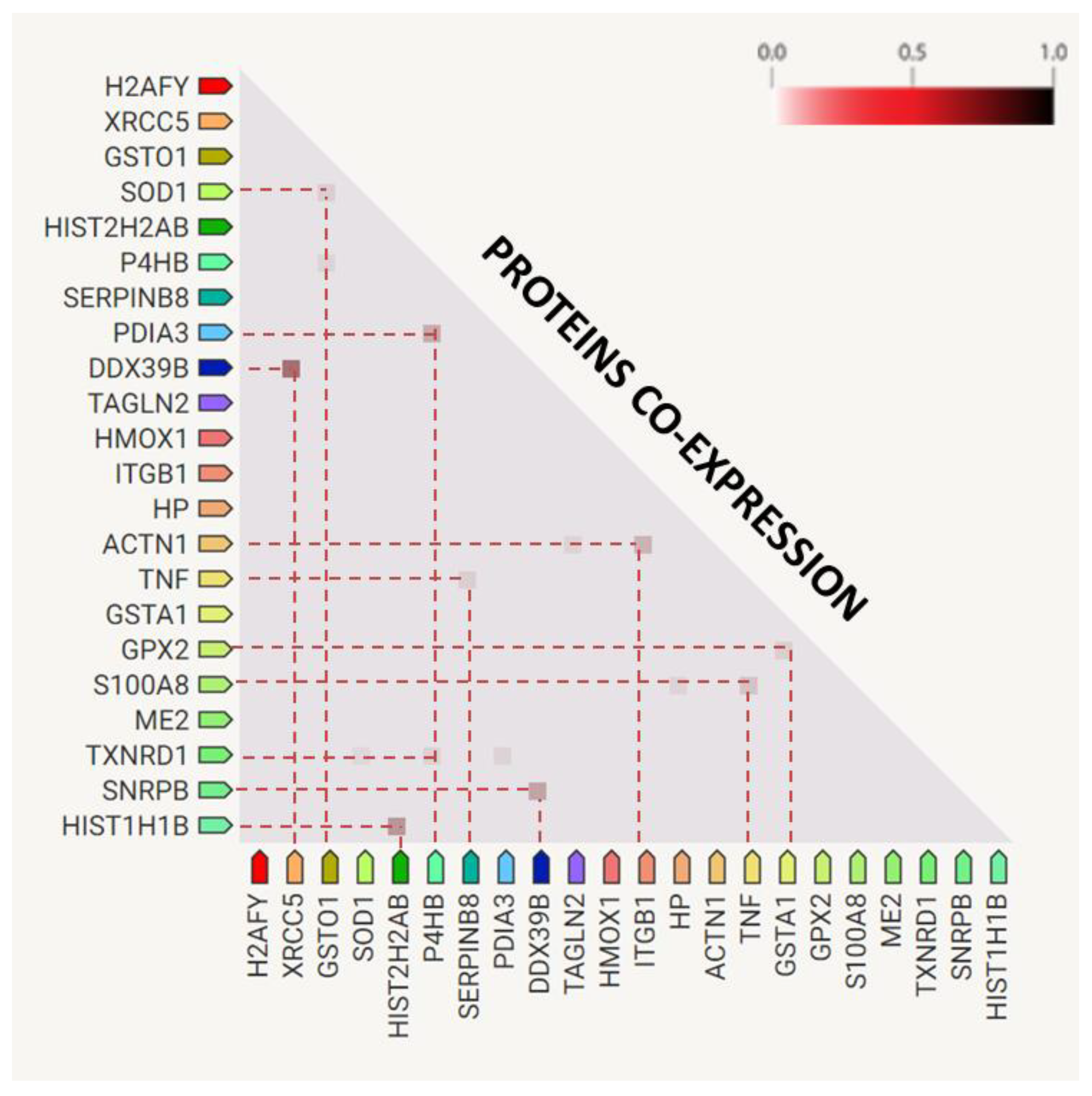
2.1. Changes in Plasma Proteome of TBE and Co-Infected Patients
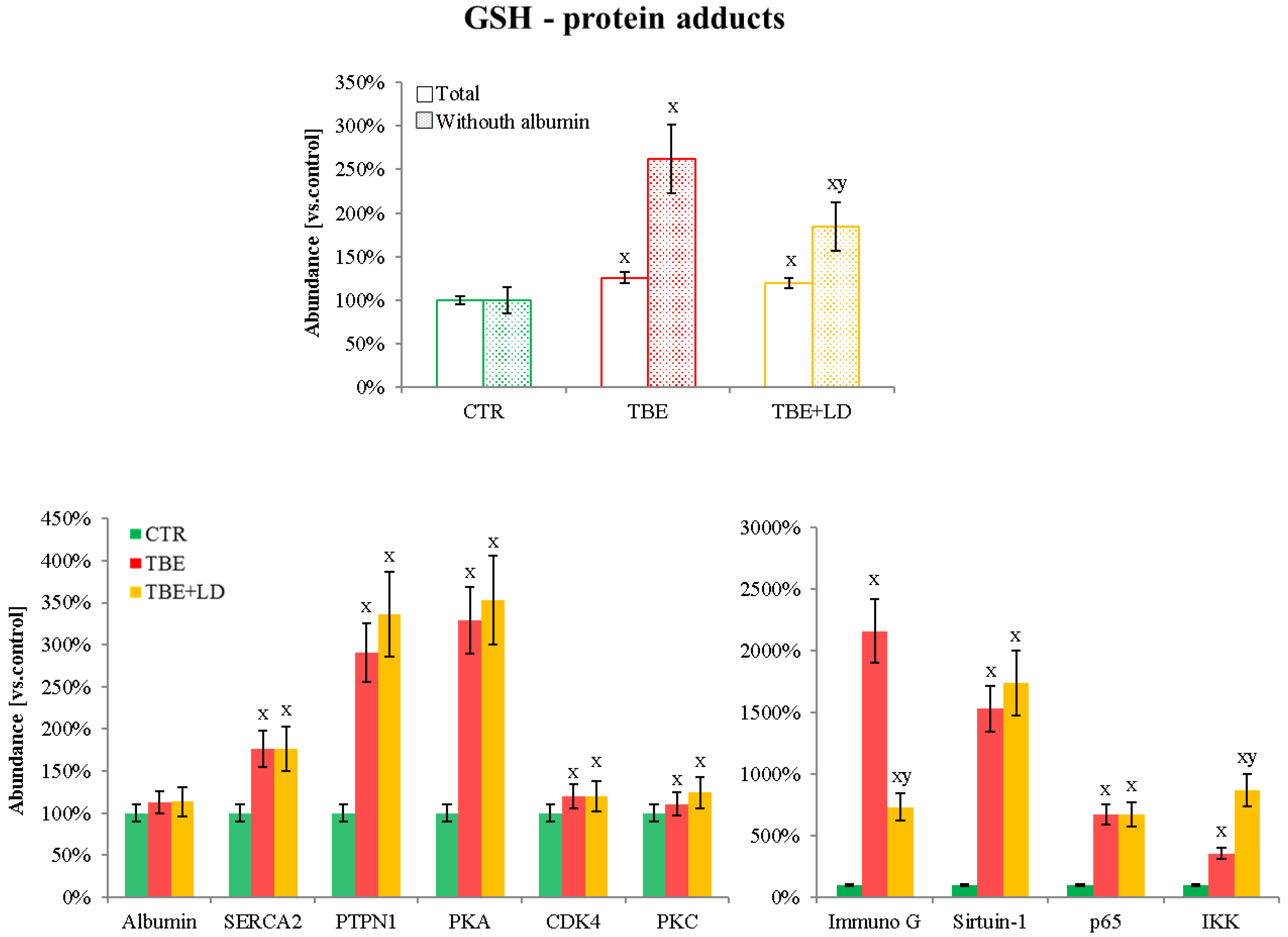
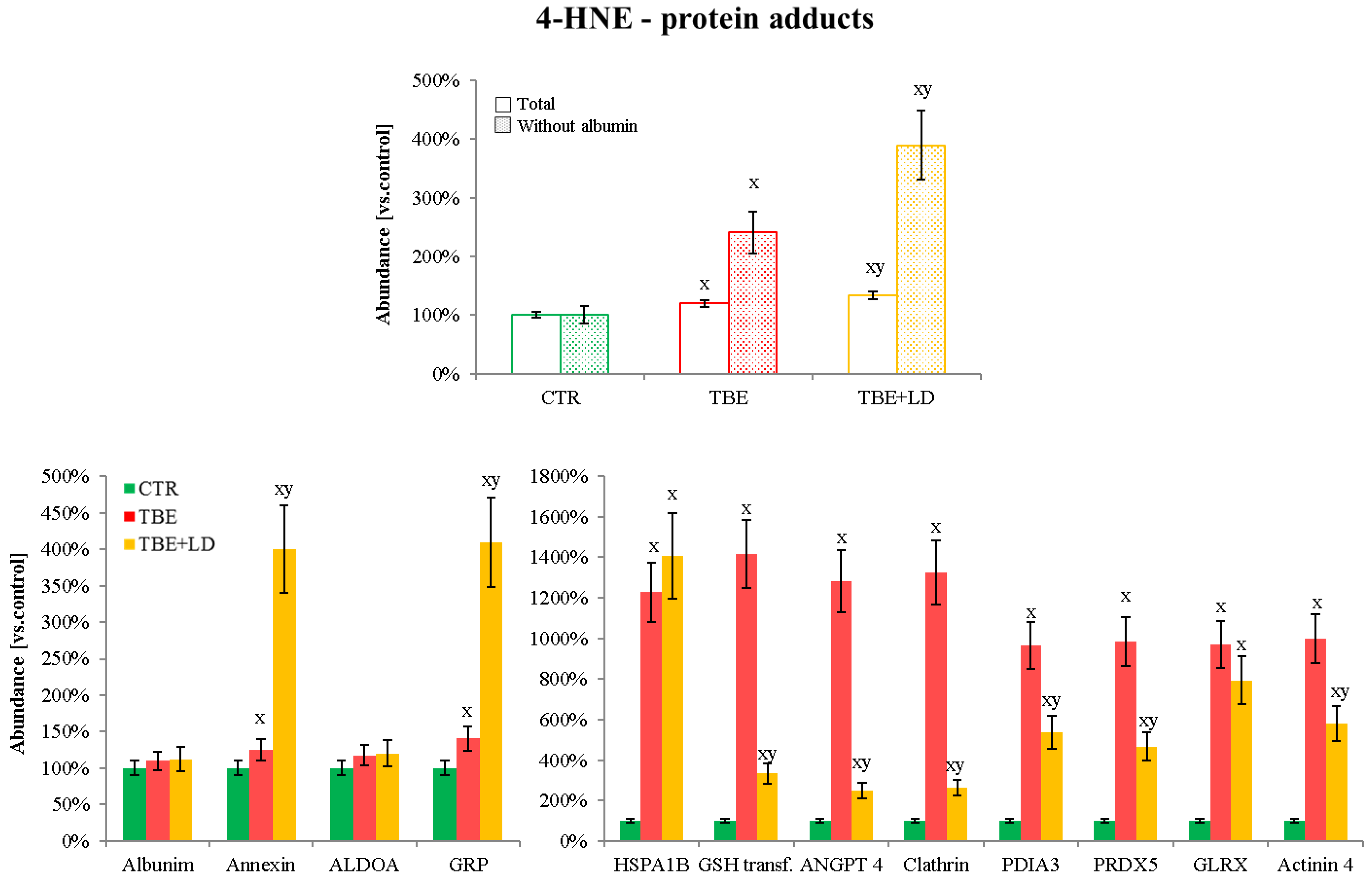
2.2. Protein Adducts Formation as an Effect of TBE and TBE+LD Infections
3. Discussion
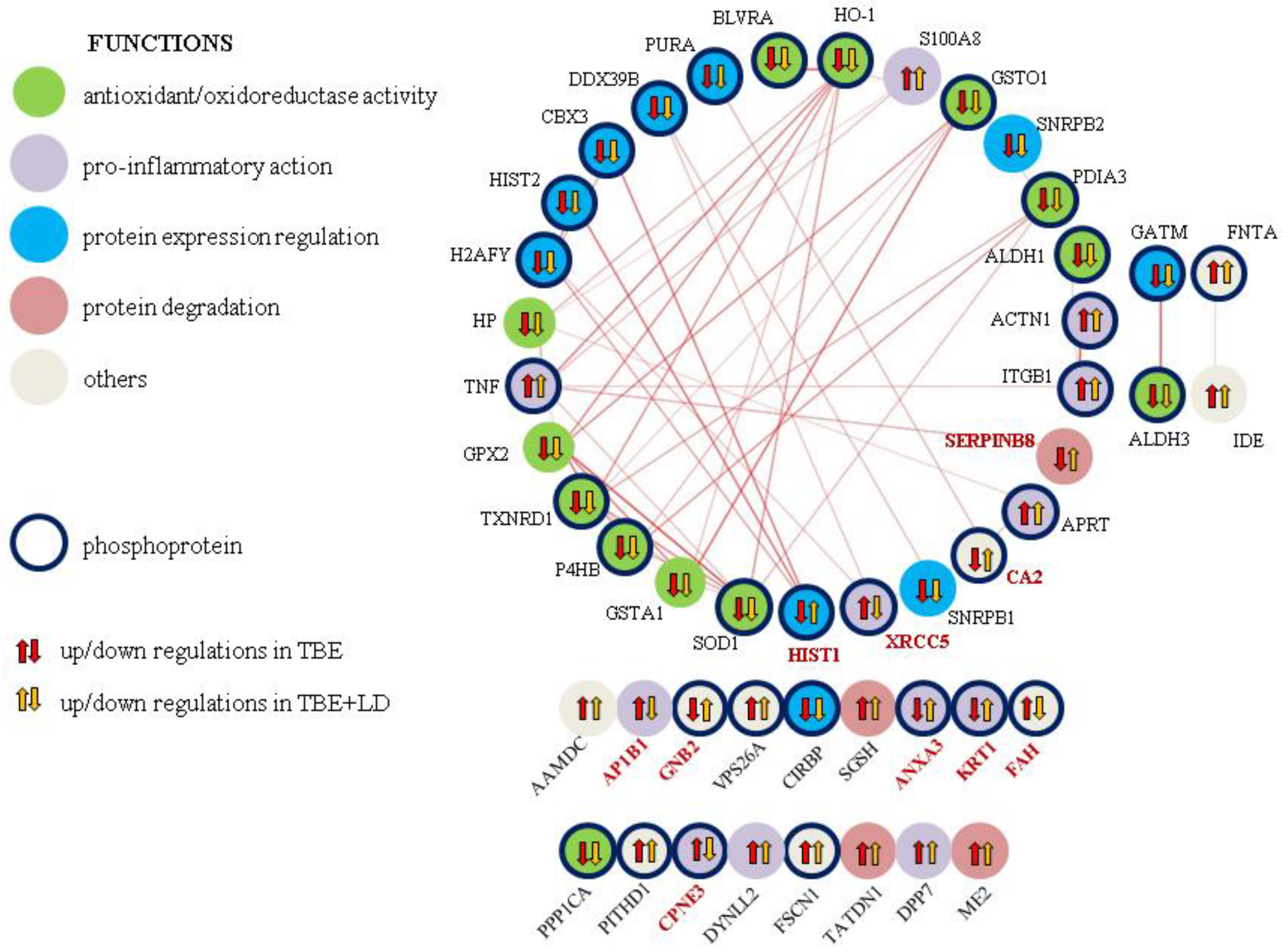
3.1. Changes in Plasma Proteome of TBE and Co-Infected Patients
3.1.1. Molecules Involved in Protein Expression Regulation
3.1.2. Expression of Proteins Involved in Antioxidant Capacity
3.1.3. Expression of Proteins Responsible for Inflammation
3.1.4. Proteins Differentiating TBE from Co-Infected Patients
3.2. Effect of GSH and 4-HNE Action on Protein Adducts Formation
3.2.1. GSH–Protein Adducts in Plasma of Patients Infected with Tick-Borne Diseases
3.2.2. 4-HNE–Protein Adducts in Plasma of Patients Infected with Tick-Borne Diseases
4. Materials and Methods
4.1. Samples Collection
- Clinical presentation: A tick-borne illness characterized by acute onset of fever and one or more of the following symptoms or signs: headache, myalgia, malaise, anemia, leukopenia, thrombocytopenia, or elevated hepatic transaminases.
- Exposure: History of having been in potential tick habitat in the 14 days prior to the onset of illness or history of tick bite.
- Laboratory criteria for diagnosis: Detection of A. phagocytophilum DNA in a clinical specimen via amplification of a specific target by polymerase chain reaction (PCR) assay (a nested PCR directed to a 546 bp fragment of the 16S rRNA gene of A. phagocytophilum was performed (Blirt-DNA Gdańsk, Poland) in a SensoQuest LabCycler (SensoQuest, Göttingen, Germany)).
4.2. Isolation of GSH–Protein Adducts
4.3. Protein Digestion and Proteomic Analysis
4.4. Protein Identification and Label-Free Quantification
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wikel, S.K. Ticks and tick-borne infections: Complex ecology, agents, and host interactions. Vet. Sci. 2018, 5, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulanger, N.; Boyer, P.; Talagrand-Reboul, E.; Hansmann, Y. Ticks and tick-borne diseases. Med. Mal. Infect. 2019, 49, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Madison-Antenucci, S.; Kramer, L.D.; Gebhardt, L.L.; Kauffman, E. Emerging tick-borne diseases. Clin. Microbiol. Rev. 2020, 33, e00083-18. [Google Scholar] [CrossRef] [PubMed]
- Tickborne Encephalitis—Chapter 4—2020 Yellow Book|Travelers’ Health|CDC. (n.d.). Available online: https://wwwnc.cdc.gov/travel/yellowbook/2020/travel-related-infectious-diseases/tickborne-encephalitis (accessed on 3 April 2022).
- Bogovic, P.; Strle, F. Tick-borne encephalitis: A review of epidemiology, clinical characteristics, and management. World J. Clin. Cases 2015, 3, 430–441. [Google Scholar] [CrossRef]
- Czupryna, P.; Grygorczuk, S.; Krawczuk, K.; Pancewicz, S.; Zajkowska, J.; Dunaj, J.; Matosek, A.; Kondrusik, M.; Moniuszko-Malinowska, A. Sequelae of tick-borne encephalitis in retrospective analysis of 1072 patients. Epidemiol. Infect. 2018, 146, 1663–1670. [Google Scholar] [CrossRef] [Green Version]
- Ruzek, D.; Avšič Županc, T.; Borde, J.; Chrdle, A.; Eyer, L.; Karganova, G.; Kholodilov, I.; Knap, N.; Kozlovskaya, L.; Matveev, A.; et al. Tick-borne encephalitis in Europe and Russia: Review of pathogenesis, clinical features, therapy, and vaccines. Antivir. Res. 2019, 164, 23–51. [Google Scholar] [CrossRef]
- Krawczuk, K.; Czupryna, P.; Pancewicz, S.; Ołdak, E.; Moniuszko-Malinowska, A. Comparison of tick-borne encephalitis between children and adults-analysis of 669 patients. J. Neurovirol. 2020, 26, 565–571. [Google Scholar] [CrossRef]
- Marques, A.R.; Strle, F.; Wormser, G.P. Comparison of Lyme Disease in the United States and Europe. Emerg. Infect. Dis. 2021, 27, 2017–2024. [Google Scholar] [CrossRef]
- Bobe, J.R.; Jutras, B.L.; Horn, E.J.; Embers, M.E.; Bailey, A.; Moritz, R.L.; Zhang, Y.; Soloski, M.J.; Ostfeld, R.S.; Marconi, R.T.; et al. Recent Progress in Lyme Disease and Remaining Challenges. Front. Med. 2021, 18, 666554. [Google Scholar] [CrossRef]
- Feder, H.M.; Johnson, B.J.; O’Connell, S.; Shapiro, E.D.; Steere, A.C.; Wormser, G.P.; Agger, W.A.; Artsob, H.; Auwaerter, P.; Dumler, J.S.; et al. A critical appraisal of “chronic Lyme disease”. N. Engl. J. Med. 2007, 357, 1422–1430. [Google Scholar] [CrossRef] [Green Version]
- Ismail, N.; McBride, J.W. Tick-Borne Emerging Infections: Ehrlichiosis and Anaplasmosis. Clin. Lab. Med. 2017, 37, 317–340. [Google Scholar] [CrossRef] [PubMed]
- Kuzmenko, Y.V.; Smirnova, O.A.; Ivanov, A.V.; Starodubova, E.S.; Karpov, V.L. Nonstructural Protein 1 of Tick-Borne Encephalitis Virus Induces Oxidative Stress and Activates Antioxidant Defense by the Nrf2/ARE Pathway. Intervirology 2016, 59, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.R.; Dennis, E.A. Borrelia burgdorferi infection induces lipid mediatorproduction during Lyme arthritis. Biochimie 2017, 141, 86. [Google Scholar] [CrossRef] [PubMed]
- Łuczaj, W.; Domingues, P.; Domingues, M.R.; Pancewicz, S.; Skrzydlewska, E. Phospholipidomic Analysis Reveals Changes in Sphingomyelin and Lysophosphatidylcholine Profiles in Plasma from Patients with Neuroborreliosis. Lipids 2017, 52, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Łuczaj, W.; Moniuszko, A.; Jarocka-Karpowicz, I.; Pancewicz, S.; Andrisic, L.; Zarkovic, N.; Skrzydlewska, E. Tick-borne encephalitis-lipid peroxidation and its consequences. Scand. J. Clin. Lab. Investig. 2016, 76, 1–9. [Google Scholar] [CrossRef]
- Du, Y.; Mi, Z.; Xie, Y.; Lu, D.; Zheng, H.; Sun, H.; Zhang, M.; Niu, Y. Insights into the molecular basis of tick-borne encephalitis from multiplatform metabolomics. PLoS Negl. Trop. Dis. 2021, 15, e0009172. [Google Scholar] [CrossRef]
- Adamu, L.; Turaki, U.A.; Bukar-Kolo, Y.M.; Husainy, A.Y.; Dauda, I.; Gulani, I.A.; Abadam, F.A.; Mani, A.U. Current updates on diagnostic methodologies for tick-borne hemoparasitic diseases in equids: A review. J. Adv. Vet. Anim. Res. 2016, 3, 84–91. [Google Scholar] [CrossRef]
- Villar, M.; Ayllón, N.; Alberdi, P.; Moreno, A.; Moreno, M.; Tobes, R.; Mateos-Hernández, L.; Weisheit, S.; Bell-Sakyi, L.; de la Fuente, J. Integrated Metabolomics, Transcriptomics and Proteomics Identifies Metabolic Pathways Affected by Anaplasma phagocytophilum Infection in Tick Cells. Mol. Cell. Proteom. 2015, 14, 3154. [Google Scholar] [CrossRef] [Green Version]
- Grillon, A.; Westermann, B.; Cantero, P.; Jaulhac, B.; Voordouw, M.J.; Kapps, D.; Collin, E.; Barthel, C.; Ehret-Sabatier, L.; Boulanger, N. Identification of Borrelia protein candidates in mouse skin for potential diagnosis of disseminated Lyme borreliosis. Sci. Rep. 2017, 7, 16719. [Google Scholar] [CrossRef] [Green Version]
- Rusanov, A.L.; Stepanov, A.A.; Zgoda, V.G.; Kaysheva, A.L.; Selinger, M.; Maskova, H.; Loginov, D.; Sterba, J.; Grubhoffer, L.; Luzgina, N.G. Proteome dataset of mouse macrophage cell line infected with tick-borne encephalitis virus. Data Br. 2020, 28, 105029. [Google Scholar] [CrossRef]
- Gal, A.; Harrus, S.; Arcoh, I.; Lavy, E.; Aizenberg, I.; Mekuzas-Yisaschar, Y.; Baneth, G. Coinfection with multiple tick-borne and intestinal parasites in a 6-week-old dog. Can. Vet. J. 2007, 48, 619. [Google Scholar] [PubMed]
- Chen, Z.; Liu, Q.; Liu, J.-Q.; Xu, B.-L.; Lv, S.; Xia, S.; Zhou, X.-N. Tick-borne pathogens and associated co-infections in ticks collected from domestic animals in central China. Parasites Vectors 2014, 7, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diuk-Wasser, M.A.; Vannier, E.; Krause, P.J. Coinfection by Ixodes Tick-Borne Pathogens: Ecological, Epidemiological, and Clinical Consequences. Trends Parasitol. 2016, 32, 30–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moniuszko, A.; Dunaj, J.; Święcicka, I.; Zambrowski, G.; Chmielewska-Badora, J.; Żukiewicz-Sobczak, W.; Zajkowska, J.; Czupryna, P.; Kondrusik, M.; Grygorczuk, S.; et al. Co-infections with Borrelia species, Anaplasma phagocytophilum and Babesia spp. in patients with tick-borne encephalitis. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 1835–1841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheeseman, K.; Jannot, G.; Lourenço, N.; Villares, M.; Berthelet, J.; Calegari-Silva, T.; Hamroune, J.; Letourneur, F.; Rodrigues-Lima, F.; Weitzman, J.B. Dynamic methylation of histone H3K18 in differentiating Theileria parasites. Nat. Commun. 2021, 12, 3221. [Google Scholar] [CrossRef] [PubMed]
- Cabezas-Cruz, A.; Alberdi, P.; Ayllón, N.; Valdés, J.J.; Pierce, R.; Villar, M.; de la Fuente, J. Anaplasma phagocytophilum increases the levels of histone modifying enzymes to inhibit cell apoptosis and facilitate pathogen infection in the tick vector Ixodes scapularis. Epigenetics 2016, 11, 303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.T.; Xue, J.H.; Zhang, Z.W.; Kong, H.B.; Liu, A.J.; Li, S.C.; Xu, D.G. Cold-inducible RNA-binding protein inhibits neuron apoptosis through the suppression of mitochondrial apoptosis. Brain Res. 2015, 1622, 474–483. [Google Scholar] [CrossRef]
- Barkhash, A.V.; Yurchenko, A.A.; Yudin, N.S.; Kozlova, I.V.; Borishchuk, I.A.; Smolnikova, M.V.; Zaitseva, O.I.; Pozdnyakova, L.L.; Voevoda, M.I.; Romaschenko, A.G. Association of ABCB9 and COL22A1 Gene Polymorphism with Human Predisposition to Severe Forms of Tick-Borne Encephalitis. Russ. J. Genet. 2019, 55, 368–377. [Google Scholar] [CrossRef]
- Katoh, H.; Mori, Y.; Kambara, H.; Abe, T.; Fukuhara, T.; Morita, E.; Moriishi, K.; Kamitani, W.; Matsuura, Y. Heterogeneous Nuclear Ribonucleoprotein A2 Participates in the Replication of Japanese Encephalitis Virus through an Interaction with Viral Proteins and RNA. J. Virol. 2011, 85, 10976–10988. [Google Scholar] [CrossRef] [Green Version]
- Dobrzyńska, M.; Moniuszko-Malinowska, A.; Jarocka-Karpowicz, I.; Czupryna, P.; Groth, M.; Skrzydlewska, E. Metabolic Response to Tick-Borne Encephalitis Virus Infection and Bacterial Co-Infections. Pathogens 2022, 11, 384. [Google Scholar] [CrossRef]
- Nazifi, S.; Razavi, S.M.; Mansourian, M.; Nikahval, B.; Moghaddam, M. Studies on correlations among parasitaemia and some hemolytic indices in two tropical diseases (theileriosis and anaplasmosis) in Fars province of Iran. Trop. Anim. Health Prod. 2007, 40, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Łuczaj, W.; Moniuszko, A.; Rusak, M.; Pancewicz, S.; Zajkowska, J.; Skrzydlewska, E. Lipid peroxidation products as potential bioindicators of Lyme arthritis. Eur. J. Clin. Microbiol. Infect. Dis. 2010, 30, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Moniuszko-Malinowska, A.; Łuczaj, W.; Jarocka-Karpowicz, I.; Pancewicz, S.; Zajkowska, J.; Andrisic, L.; Zarkovic, N.; Skrzydlewska, E. Lipid peroxidation in the pathogenesis of neuroborreliosis. Free Radic. Biol. Med. 2016, 96, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, A.P.; Holmgren, A. Glutaredoxins: Glutathione-Dependent Redox Enzymes with Functions Far Beyond a Simple Thioredoxin Backup System. Antioxid. Redox Signal. 2004, 6, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Conrad, M.; Schick, J.; Angeli, J.P.F. Glutathione and thioredoxin dependent systems in neurodegenerative disease: What can be learned from reverse genetics in mice. Neurochem. Int. 2013, 62, 738–749. [Google Scholar] [CrossRef]
- Jastrząb, A.; Jarocka-Karpowicz, I.; Markowska, A.; Wroński, A.; Gȩgotek, A.; Skrzydlewska, E. Antioxidant and Anti-inflammatory Effect of Cannabidiol Contributes to the Decreased Lipid Peroxidation of Keratinocytes of Rat Skin Exposed to UV Radiation. Oxid. Med. Cell. Longev. 2021, 2021, 6647222. [Google Scholar] [CrossRef]
- Wu, C.; Parrott, A.M.; Fu, C.; Liu, T.; Marino, S.M.; Gladyshev, V.N.; Jain, M.R.; Baykal, A.T.; Li, Q.; Oka, S.; et al. Thioredoxin 1-Mediated Post-Translational Modifications: Reduction, Transnitrosylation, Denitrosylation, and Related Proteomics Methodologies. Antioxid. Redox Signal. 2011, 15, 2565–2604. Available online: https://home.liebertpub.com/ars (accessed on 20 December 2021). [CrossRef] [Green Version]
- Luz, N.F.; DeSouza-Vieira, T.; De Castro, W.; Vivarini, A.C.; Pereira, L.; França, R.R.; Silveira-Mattos, P.S.; Costa, D.L.; Teixeira, C.; Meneses, C.; et al. Lutzomyia longipalpis Saliva Induces Heme Oxygenase-1 Expression at Bite Sites. Front. Immunol. 2018, 9, 2779. [Google Scholar] [CrossRef]
- Roth, W.; Kumar, V.; Beer, H.D.; Richter, M.; Wohlenberg, C.; Reuter, U.; Thiering, S.; Staratschek-Jox, A.; Hofmann, A.; Kreusch, F.; et al. Keratin 1 maintains skin integrity and participates in an inflammatory network in skin through interleukin-18. J. Cell Sci. 2012, 125, 5269–5279. [Google Scholar]
- Herbein, G.; Khan, K.A. Is HIV infection a TNF receptor signalling-driven disease? Trends Immunol. 2008, 29, 61–67. [Google Scholar] [CrossRef]
- Wang, S.; Song, R.; Wang, Z.; Jing, Z.; Wang, S.; Ma, J. S100A8/A9 in inflammation. Front. Immunol. 2018, 9, 1298. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Pan, Q.Z.; Zhong, A.L.; Hu, H.; Zhao, J.J.; Tang, Y.; Hu, W.M.; Li, M.; Weng, D.S.; Chen, M.Y.; et al. Annexin A3 upregulates the infiltrated neutrophil-lymphocyte ratio to remodel the immune microenvironment in hepatocellular carcinoma. Int. Immunopharmacol. 2020, 89, 107139. [Google Scholar] [CrossRef] [PubMed]
- Ólafsson, E.B.; Barragan, A. The unicellular eukaryotic parasite Toxoplasma gondii hijacks the migration machinery of mononuclear phagocytes to promote its dissemination. Biol. Cell 2020, 112, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Cao, J.; Zhou, Y.; Zhang, H.; Zhou, J. Isolation and characterization of two novel serpins from the tick Rhipicephalus haemaphysaloides. Ticks Tick. Borne. Dis. 2013, 4, 297–303. [Google Scholar] [CrossRef]
- Li, Y.; Liu, S.; Qin, Z.; Yao, J.; Jiang, C.; Song, L.; Dunham, R.; Liu, Z. The serpin superfamily in channel catfish: Identification, phylogenetic analysis and expression profiling in mucosal tissues after bacterial infections. Dev. Comp. Immunol. 2015, 49, 267–277. [Google Scholar] [CrossRef]
- Bao, J.; Pan, G.; Poncz, M.; Wei, J.; Ran, M.; Zhou, Z. Serpin functions in host-pathogen interactions. PeerJ 2018, 6, e4557. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Zheng, D.; Abbott, J.; Liu, L.; Bartee, M.Y.; Long, M.; Davids, J.; Williams, J.; Feldmann, H.; Strong, J.; et al. Myxomavirus-derived serpin prolongs survival and reduces inflammation and hemorrhage in an unrelated lethal mouse viral infection. Antimicrob. Agents Chemother. 2013, 57, 4114–4127. [Google Scholar] [CrossRef] [Green Version]
- Torina, A.; Villari, S.; Blanda, V.; Vullo, S.; La Manna, M.P.; Azgomi, M.S.; Di Liberto, D.; de la Fuente, J.; Sireci, G. Innate Immune Response to Tick-Borne Pathogens: Cellular and Molecular Mechanisms Induced in the Hosts. Int. J. Mol. Sci. 2020, 21, 5437. [Google Scholar] [CrossRef]
- Determination of Erythrocyte Zinc and Copper Concentration and Carbonic Anhydrase Enzyme Activities in Sheep with Babesiosis. Available online: https://www.cabdirect.org/cabdirect/abstract/20113074002 (accessed on 31 August 2021).
- Grab, D.J.; Nyarko, E.; Barat, N.C.; Nikolskaia, O.V.; Dumler, J.S. Anaplasma phagocytophilum-Borrelia burgdorferi coinfection enhances chemokine, cytokine, and matrix metalloprotease expression by human brain microvascular endothelial cells. Clin. Vaccine Immunol. 2007, 14, 1420–1424. [Google Scholar] [CrossRef] [Green Version]
- Barbet, A.F.; Blentlinger, R.; Yi, J.; Lundgren, A.M.; Blouin, E.F.; Kocan, K.M. Comparison of surface proteins of Anaplasma marginale grown in tick cell culture, tick salivary glands, and cattle. Infect. Immun. 1999, 67, 102–107. [Google Scholar] [CrossRef] [Green Version]
- Wagner, E.F. AP-1—Introductory remarks. Oncogene 2001, 20, 2334–2335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, B.; Li, Y.; Zhou, Y.; Ng, T.K.; Zhao, C.; Gan, Q.; Gu, X.; Xiang, J. Circulating exosomal CPNE3 as a diagnostic and prognostic biomarker for colorectal cancer. J. Cell. Physiol. 2019, 234, 1416–1425. [Google Scholar] [CrossRef] [PubMed]
- Grabowski, J.M.; Gulia-Nuss, M.; Kuhn, R.J.; Hill, C.A. RNAi reveals proteins for metabolism and protein processing associated with Langat virus infection in Ixodes scapularis (black-legged tick) ISE6 cells. Parasites Vectors 2017, 10, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebicki, J.M.; Nauser, T.; Domazou, A.; Steinmann, D.; Bounds, P.L.; Koppenol, W.H. Reduction of protein radicals by GSH and ascorbate: Potential biological significance. Amino Acids 2010, 39, 1131–1137. [Google Scholar] [CrossRef]
- Checconi, P.; Limongi, D.; Baldelli, S.; Ciriolo, M.R.; Nencioni, L.; Palamara, A.T. Role of Glutathionylation in Infection and Inflammation. Nutrients 2019, 11, 1952. [Google Scholar] [CrossRef] [Green Version]
- Kerstholt, M.; Vrijmoeth, H.; Lachmandas, E.; Oosting, M.; Lupse, M.; Flonta, M.; Dinarello, C.A.; Netea, M.G.; Joosten, L.A.B. Role of glutathione metabolism in host defense against Borrelia burgdorferi infection. Proc. Natl. Acad. Sci. USA 2018, 115, E2320–E2328. [Google Scholar] [CrossRef] [Green Version]
- Ghezzi, P. Protein glutathionylation in health and disease. Biochim. Biophys. Acta-Gen. Subj. 2013, 1830, 3165–3172. [Google Scholar] [CrossRef]
- Shelton, M.D.; Mieyal, J.J. Regulation by Reversible S-Glutathionylation: Molecular Targets Implicated in Inflammatory Diseases. Mol. Cells 2008, 25, 332. [Google Scholar]
- Humphries, K.M.; Juliano, C.; Taylor, S.S. Regulation of cAMP-dependent Protein Kinase Activity by Glutathionylation*. J. Biol. Chem. 2002, 277, 43505–43511. [Google Scholar] [CrossRef] [Green Version]
- Reddy, S.; Jones, A.D.; Cross, C.E.; Wong, P.S.-Y.; Van Der Vliet, A. Inactivation of creatine kinase by S-glutathionylation of the active-site cysteine residue. Biochem. J. 2000, 347, 821–827. [Google Scholar] [CrossRef]
- Kambe, T.; Song, T.; Takata, T.; Hatano, N.; Miyamoto, Y.; Nozaki, N.; Naito, Y.; Tokumitsu, H.; Watanabe, Y. Inactivation of Ca2+/calmodulin-dependent protein kinase I by S-glutathionylation of the active-site cysteine residue. FEBS Lett. 2010, 584, 2478–2484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anselmo, A.N.; Cobb, M.H. Protein kinase function and glutathionylation. Biochem. J. 2004, 381, e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popov, D. Protein S-glutathionylation: From current basics to targeted modifications. Arch. Physiol. Biochem. 2014, 120, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Hoxhaj, G.; Manning, B.D. The PI3K–AKT network at the interface of oncogenic signalling and cancer metabolism. Nat. Rev. Cancer 2019, 20, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Lieskovska, J.; Kopecky, J. Effect of tick saliva on signalling pathways activated by TLR-2 ligand and Borrelia afzelii in dendritic cells. Parasite Immunol. 2012, 34, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Carroll, L.; Rasmussen, L.M.; Davies, M.J. Oxidation of protein disulfide bonds by singlet oxygen gives rise to glutathionylated proteins. Redox Biol. 2021, 38, 101822. [Google Scholar] [CrossRef] [PubMed]
- Manevich, Y.; Ye, Z.-W.; Zhang, J.; Ancrum, T.; Townsend, D.M.; Tew, K.D. S-Glutathionylation of Endoplasmic Reticulum Proteins Impacts Unfolded Protein Response Sensitivity. Antioxid. Redox Signal. 2017, 26, 247–261. [Google Scholar]
- Łuczaj, W.; Gęgotek, A.; Skrzydlewska, E. Antioxidants and HNE in redox homeostasis. Free Radic. Biol. Med. 2017, 111, 87–101. [Google Scholar] [CrossRef]
- D’Acunto, C.W.; Gbelcova, H.; Festa, M.; Ruml, T. The complex understanding of Annexin A1 phosphorylation. Cell. Signal. 2014, 26, 173–178. [Google Scholar] [CrossRef] [Green Version]
- Gęgotek, A.; Skrzydlewska, E. Biological effect of protein modifications by lipid peroxidation products. Chem. Phys. Lipids 2019, 221, 46–52. [Google Scholar] [CrossRef]
- Ibrahim, I.M.; Abdelmalek, D.H.; Elfiky, A.A. GRP78: A cell’s response to stress. Life Sci. 2019, 226, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Royle, J.; Ramírez-Santana, C.; Akpunarlieva, S.; Donald, C.L.; Gestuveo, R.J.; Anaya, J.-M.; Merits, A.; Burchmore, R.; Kohl, A.; Varjak, M. Glucose-Regulated Protein 78 Interacts with Zika Virus Envelope Protein and Contributes to a Productive Infection. Viruses 2020, 12, 524. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Liu, Q.; Han, K.; Wang, H.; Yang, J.; Bi, K.; Liu, Y.; Liu, N.; Tian, Y.; Li, Y. Identification of Glucose-Regulated Protein 78 (GRP78) as a Receptor in BHK-21 Cells for Duck Tembusu Virus Infection. Front. Microbiol. 2018, 9, 694. [Google Scholar] [CrossRef] [PubMed]
- Alam, J.; Kim, Y.C.; Choi, Y. Potential Role of Bacterial Infection in Autoimmune Diseases: A New Aspect of Molecular Mimicry. Immune Netw. 2014, 14, 7–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galligan, J.J.; Fritz, K.S.; Backos, D.S.; Shearn, C.T.; Smathers, R.L.; Jiang, H.; Maclean, K.N.; Reigan, P.R.; Petersen, D.R. Oxidative stress-mediated aldehyde adduction of GRP78 in a mouse model of alcoholic liver disease: Functional independence of ATPase activity and chaperone function. Free Radic. Biol. Med. 2014, 73, 411–420. [Google Scholar] [CrossRef] [Green Version]
- Chavez, J.; Chung, W.-G.; Miranda, C.L.; Singhal, M.; Stevens, J.F.; Maier, C.S. Site-Specific Protein Adducts of 4-Hydroxy-2(E)-Nonenal in Human THP-1 Monocytic Cells: Protein Carbonylation Is Diminished by Ascorbic Acid. Chem. Res. Toxicol. 2009, 23, 37–47. [Google Scholar] [CrossRef] [Green Version]
- EUR-Lex-32012D0506-EN-EUR-Lex. Available online: https://eur-lex.europa.eu/eli/dec_impl/2012/506/oj (accessed on 23 September 2021).
- Stanek, G.; Fingerle, V.; Hunfeld, K.P.; Jaulhac, B.; Kaiser, R.; Krause, A.; Kristoferitsch, W.; O’Connell, S.; Ornstein, K.; Strle, F.; et al. Lyme borreliosis: Clinical case definitions for diagnosis and management in Europe. Clin. Microbiol. Infect. 2011, 17, 69–79. [Google Scholar] [CrossRef] [Green Version]
- Mygland, Å.; Ljøstad, U.; Fingerle, V.; Rupprecht, T.; Schmutzhard, E.; Steiner, I. EFNS guidelines on the diagnosis and management of European Lyme neuroborreliosis. Eur. J. Neurol. 2010, 17, 8-e4. [Google Scholar] [CrossRef]
- Bradford, M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Medzihradszky, K.F. In-solution digestion of proteins for mass spectrometry. Methods Enzymol. 2005, 405, 50–65. [Google Scholar]
- Gęgotek, A.; Domingues, P.; Wroński, A.; Wójcik, P.; Skrzydlewska, E. Proteomic plasma profile of psoriatic patients. J. Pharm. Biomed. Anal. 2018, 155, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Domingues, M.; Fedorova, M.; Domingues, P. Mass spectrometry detection of protein modification by cross-reaction with lipid peroxidation products. React. Oxyg. Species Lipid Peroxidation Protein Oxid. 2015, 3, 61–86. [Google Scholar]
- Vanhecke, G.C.; Abeywardana, M.Y.; Ahn, Y.H. Proteomic Identification of Protein Glutathionylation in Cardiomyocytes. J. Proteome Res. 2019, 18, 1806–1818. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for comprehensive and integrative metabolomics data analysis. Curr. Protoc. Bioinf. 2019, 68, e86. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.; Wang, J.; Ramage, D.; Amin, A.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CTR | TBE | TBE+LD | |
|---|---|---|---|
| Age (years) | 40.25 ± 7.42 | 42.75 ± 9.33 | 42.5 ± 15.24 |
| Sex, female/male | 4/8 female (50%) 4/8 male (50%) | 7/16 female (43%) 9/16 male (57%) | 3/5 female (60%) 2/5 male (40%) |
| Place of residence (urban/rural area) | 7/16 urban (44%) 10/16 rural (56%) | 3/5 urban (60%) 2/5 rural (40%) | |
| Noticeable tick bite | 0/8 (0%) | 9/16 (56%) | 5/5 (100%) |
| Time since tick bite (days) | 25 ± 16.19 | 17.5 ± 4.95 | |
| Duration of hospitalization (days) | 12.56 ± 2.13 | 12.6 ± 2.07 | |
| Duration of symptoms (days) | 8.53 ± 8.08 | 4.33 ± 3.21 | |
| Clinical form | |||
| Meningitis | 0/8 (0%) | 10/16 (62%) | 4/5 (83%) |
| Meningoencephalitis | 0/8 (0%) | 6/16 (38%) | 1/5 (17%) |
| Meningoencephalomyelitis | 0/8 (0%) | 0/16 (0%) | 0/5 (0%) |
| Clinical presentation | |||
| Headache | 0/8 (0%) | 16/16 (100%) | 5/5 (100%) |
| Fever | 0/8 (0%) | 15/16 (94%) | 4/5 (80%) |
| Neck stiffness | 0/8 (0%) | 13/16 (81%) | 3/5 (60%) |
| Kernig’s sign | 0/8 (0%) | 5/16 (31%) | 1/5 (20%) |
| Vertigo | 0/8 (0%) | 7/16 (44%) | 0/5 (0%) |
| Nausea | 0/8 (0%) | 7/16 (44%) | 2/5 (40%) |
| Vomiting | 0/8 (0%) | 6/16 (38%) | 0/5 (0%) |
| Ataxia | 0/8 (0%) | 5/16 (31%) | 0/5 (0%) |
| Tremor | 0/8 (0%) | 4/16 (25%) | 0/5 (0%) |
| Hyperesthesia | 0/8 (0%) | 2/16 (13%) | 1/5 (20%) |
| Muscle pain | 0/8 (0%) | 1/16 (6%) | 0/5 (0%) |
| Joint pain | 0/8 (0%) | 1/16 (6%) | 0/5 (0%) |
| Consciousness disturbances | 0/8 (0%) | 1/16 (6%) | 0/5 (0%) |
| Skin lesion | 0/8 (0%) | 0/16 (0%) | 1/5 (20%) |
| Muscle weakness | 0/8 (0%) | 0/16 (0%) | 1/5 (20%) |
| CTR | TBE | TBE+LD | |
|---|---|---|---|
| Complete blood count | |||
| WBC [103/μL] | 4.00–10.00 | 10.35 ± 2.27 | 7.41 ± 1.47 |
| Neutrophils [%] | 40.0–72.0 | 70.56 ± 10.21 | 61.2 ± 7.54 |
| Lymphocytes [%] | 18.00–48.00 | 18.8 ± 9.27 | 25.58 ± 4.77 |
| Monocytes [%] | 2.50–10.00 | 9.49 ± 2.68 | 10.86 ± 2.18 |
| RBC [106/μL] | 4.00–5.50 | 4.29 ± 0.33 | 4.33 ± 0.61 |
| HGB [g/dL] | 12.00–16.00 | 12.91 ± 1.08 | 12.74 ± 1.39 |
| PLT [103/μL] | 130–350 | 251 ± 49.17 | 280 ± 57.92 |
| CRP [mg/L] | 0.00–5.00 | 12.16 ± 18.48 | 1.56 ± 1.28 |
| Glucose [mg/dL] | 70–110 | 96 ± 10.47 | 92.67 ± 8.5 |
| Creatinine [mg/dL] | 0.50–0.90 | 0.89 ± 0.17 | 0.79 ± 0.09 |
| ALT [U/I] | 0–31 | 25 ± 27.3 | 17.75 ± 11.27 |
| AST [U/I] | 0–32 | 16.2 ± 7.01 | 17 ± 3.67 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gęgotek, A.; Moniuszko-Malinowska, A.; Groth, M.; Pancewicz, S.; Czupryna, P.; Dunaj, J.; Atalay, S.; Radziwon, P.; Skrzydlewska, E. Plasma Proteomic Profile of Patients with Tick-Borne Encephalitis and Co-Infections. Int. J. Mol. Sci. 2022, 23, 4374. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23084374
Gęgotek A, Moniuszko-Malinowska A, Groth M, Pancewicz S, Czupryna P, Dunaj J, Atalay S, Radziwon P, Skrzydlewska E. Plasma Proteomic Profile of Patients with Tick-Borne Encephalitis and Co-Infections. International Journal of Molecular Sciences. 2022; 23(8):4374. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23084374
Chicago/Turabian StyleGęgotek, Agnieszka, Anna Moniuszko-Malinowska, Monika Groth, Sławomir Pancewicz, Piotr Czupryna, Justyna Dunaj, Sinemyiz Atalay, Piotr Radziwon, and Elżbieta Skrzydlewska. 2022. "Plasma Proteomic Profile of Patients with Tick-Borne Encephalitis and Co-Infections" International Journal of Molecular Sciences 23, no. 8: 4374. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23084374