
Scaffold Pore Curvature Influences ΜSC Fate through Differential Cellular Organization and YAP/TAZ Activity

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
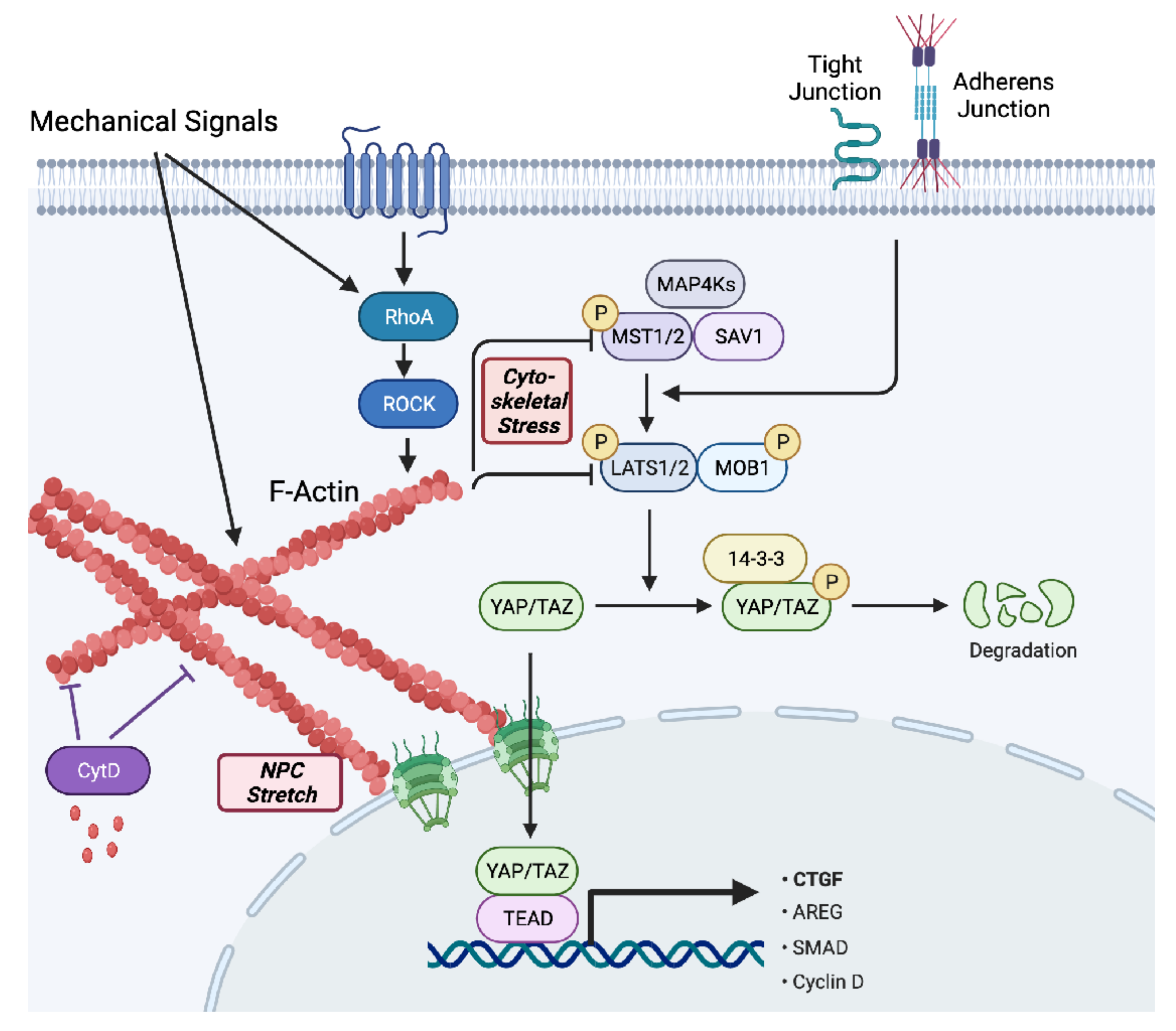{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Discretely Controlled Macropore Diameters Modulate Principle Curvature in Nanofibrous Scaffolds
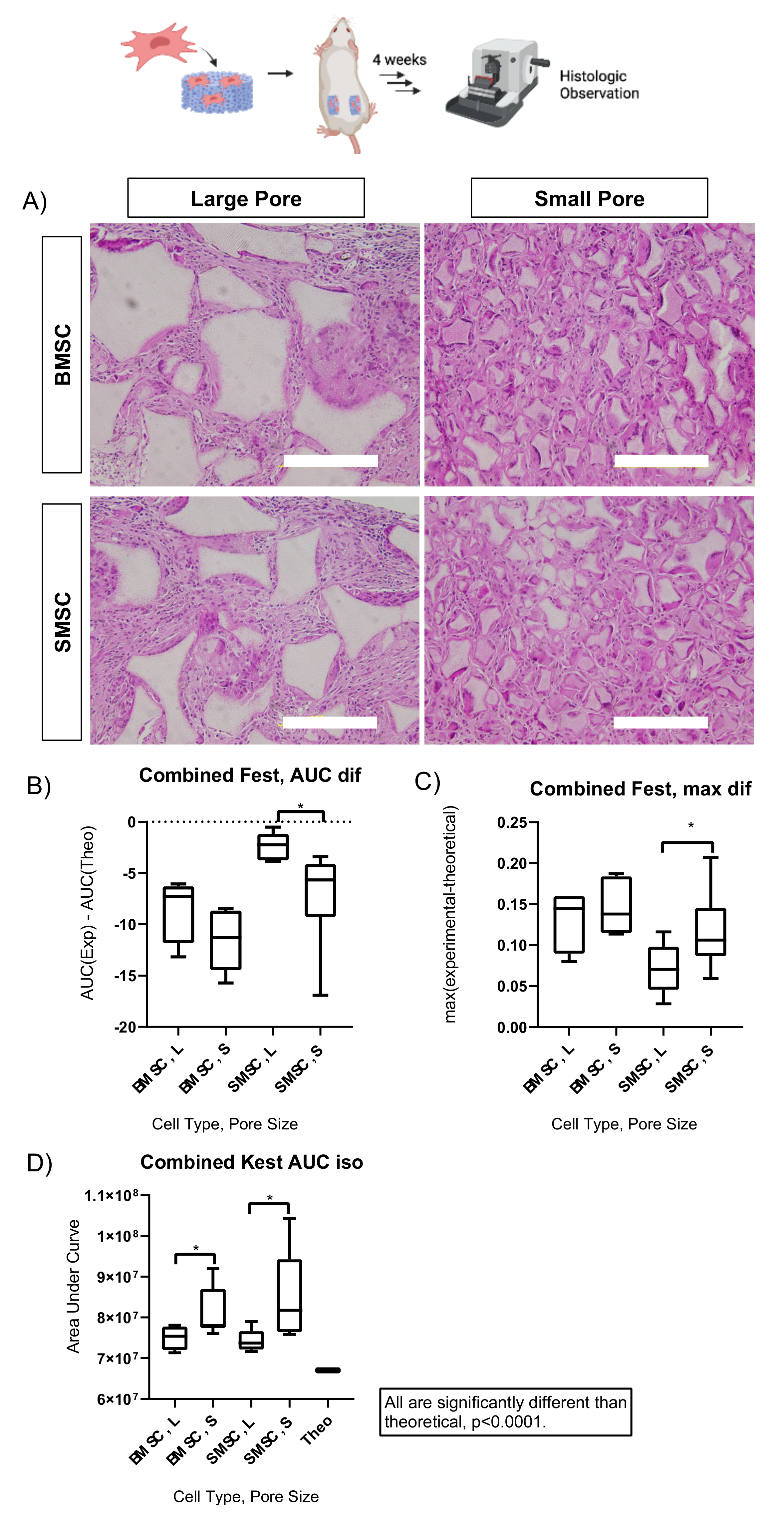
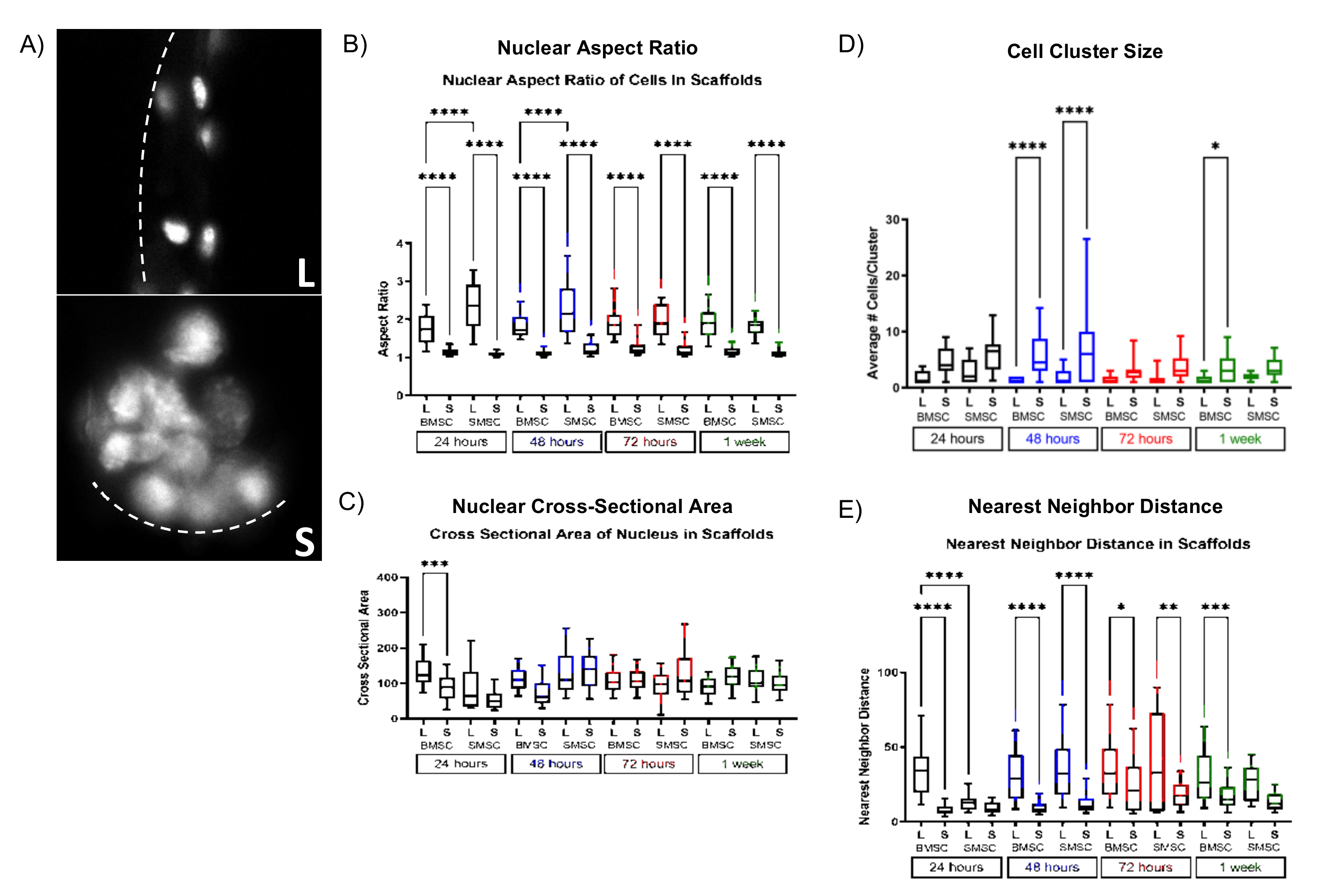
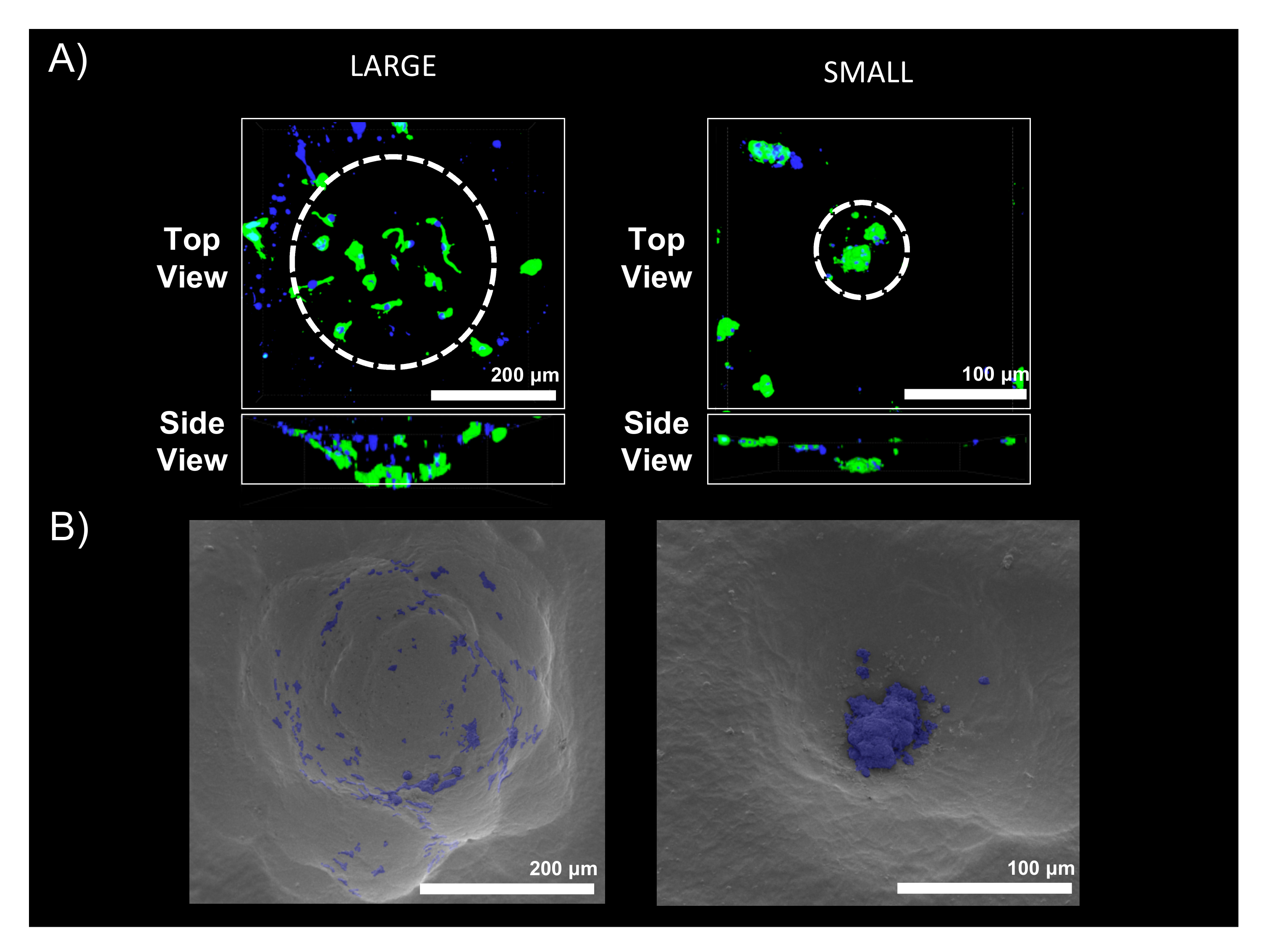
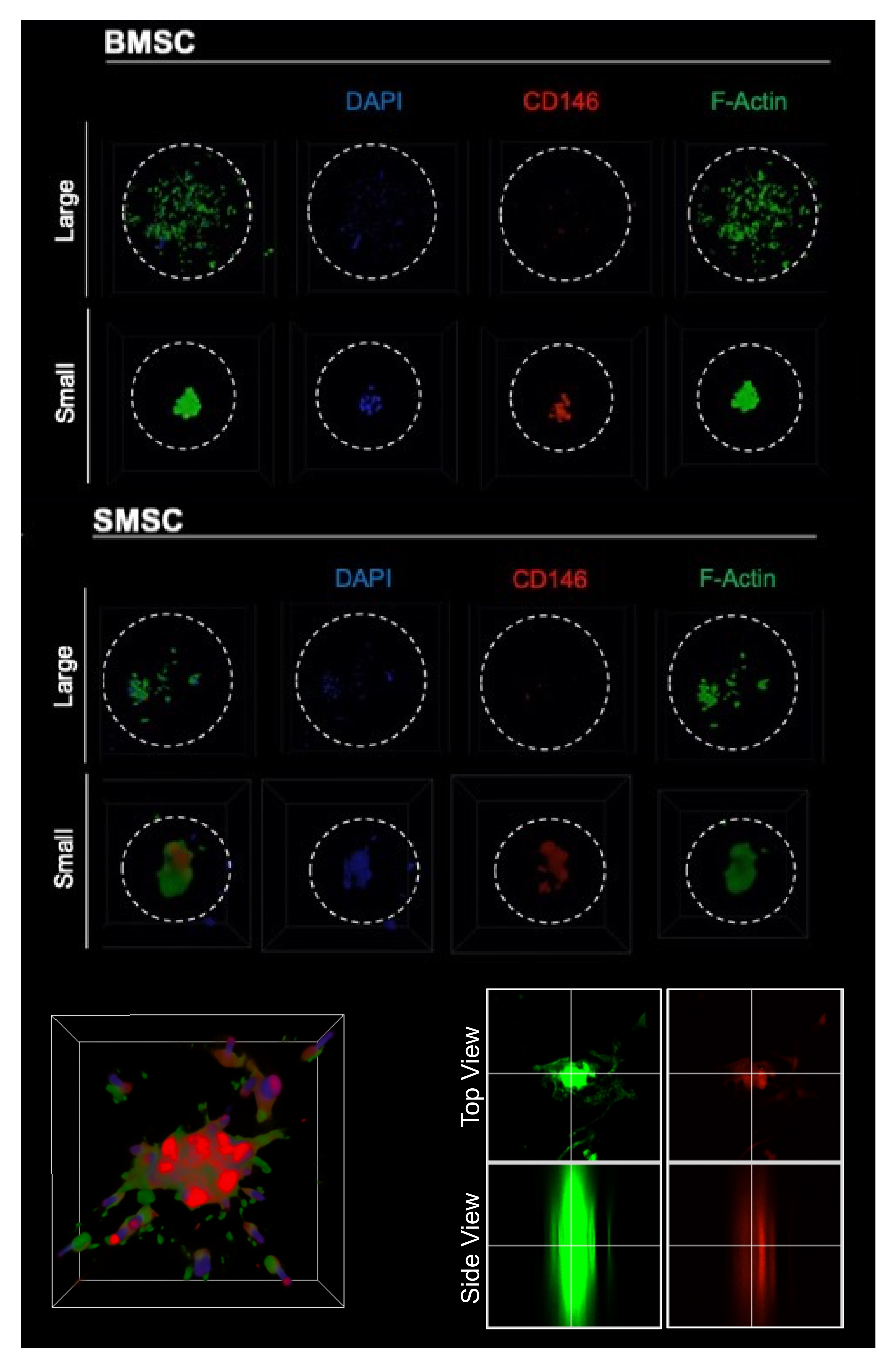
2.2. Scaffold Pore Size Facilitates Cell Organization In Vivo and In Vitro
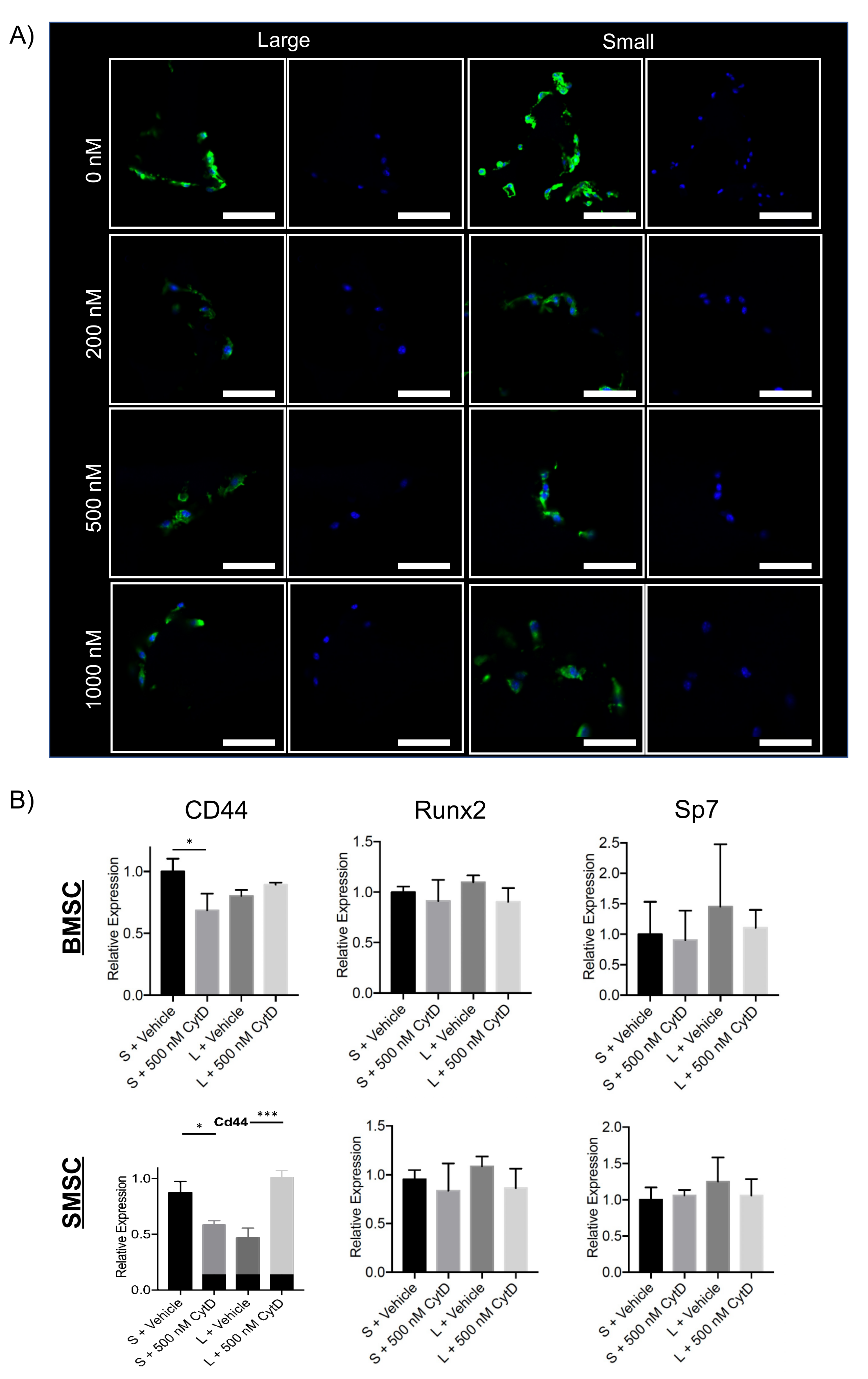
2.3. Cytoskeletal Disruption in Scaffold Macropores Alters Cell Morphology and Gene Expression
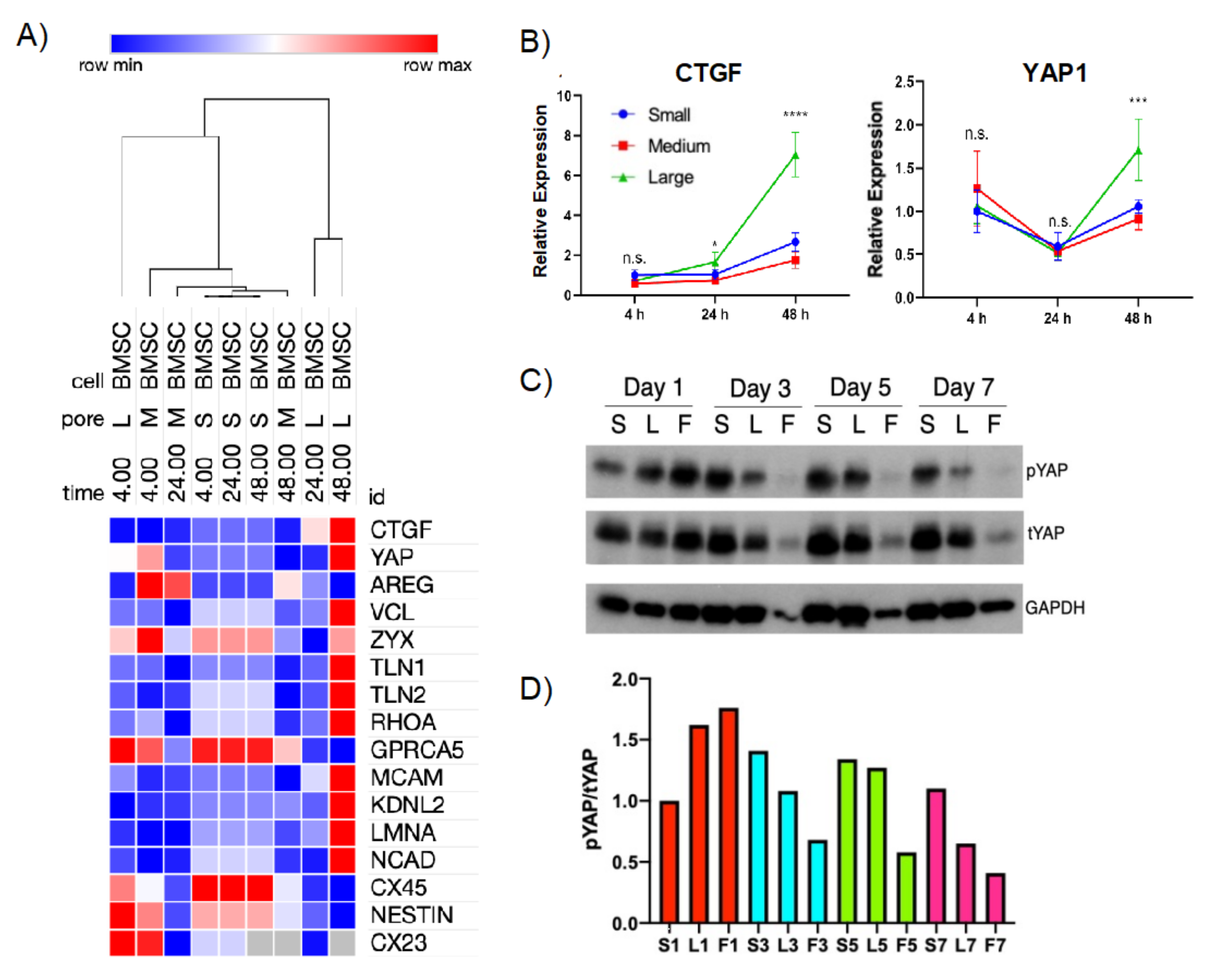
2.4. Curvature-Mediated Cytoskeletal Strain May Differentially Regulate YAP/TAZ Signaling in Cell–Scaffold Constructs
2.5. Reproducing Curvature-Mediated Cell Organization with a Macropore-Mimetic Hemisphere Platform to Assess Differential YAP Regulation
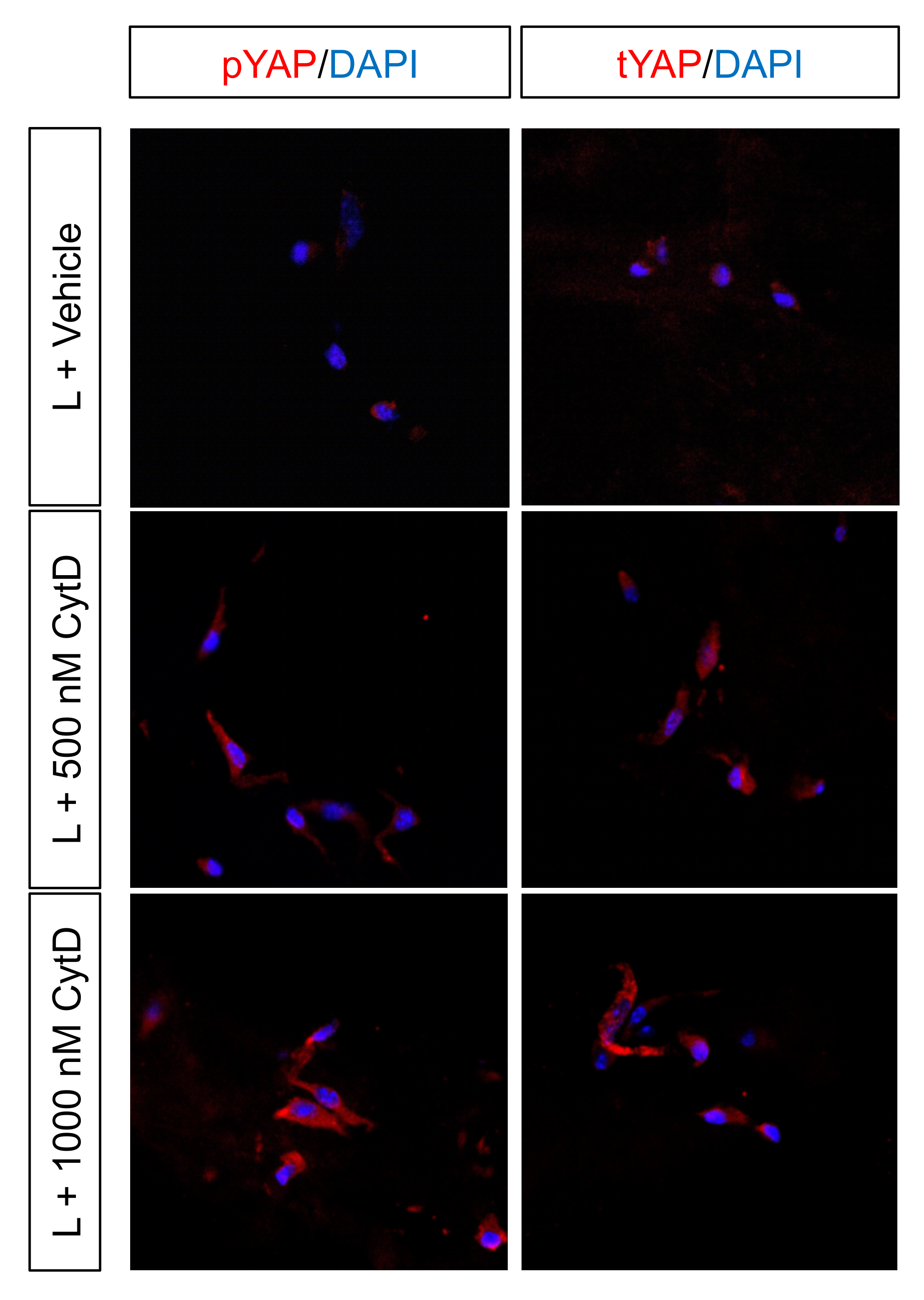
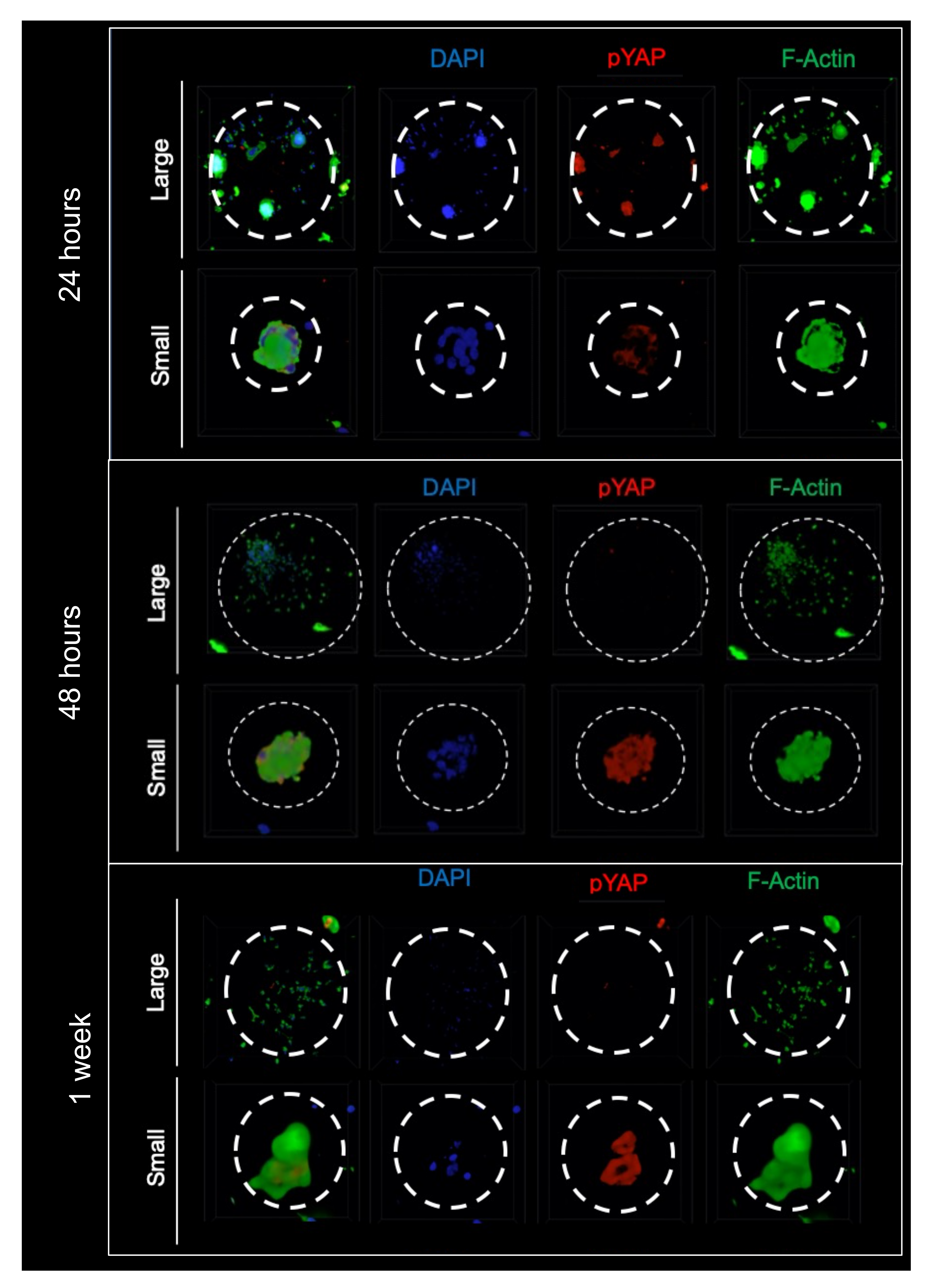
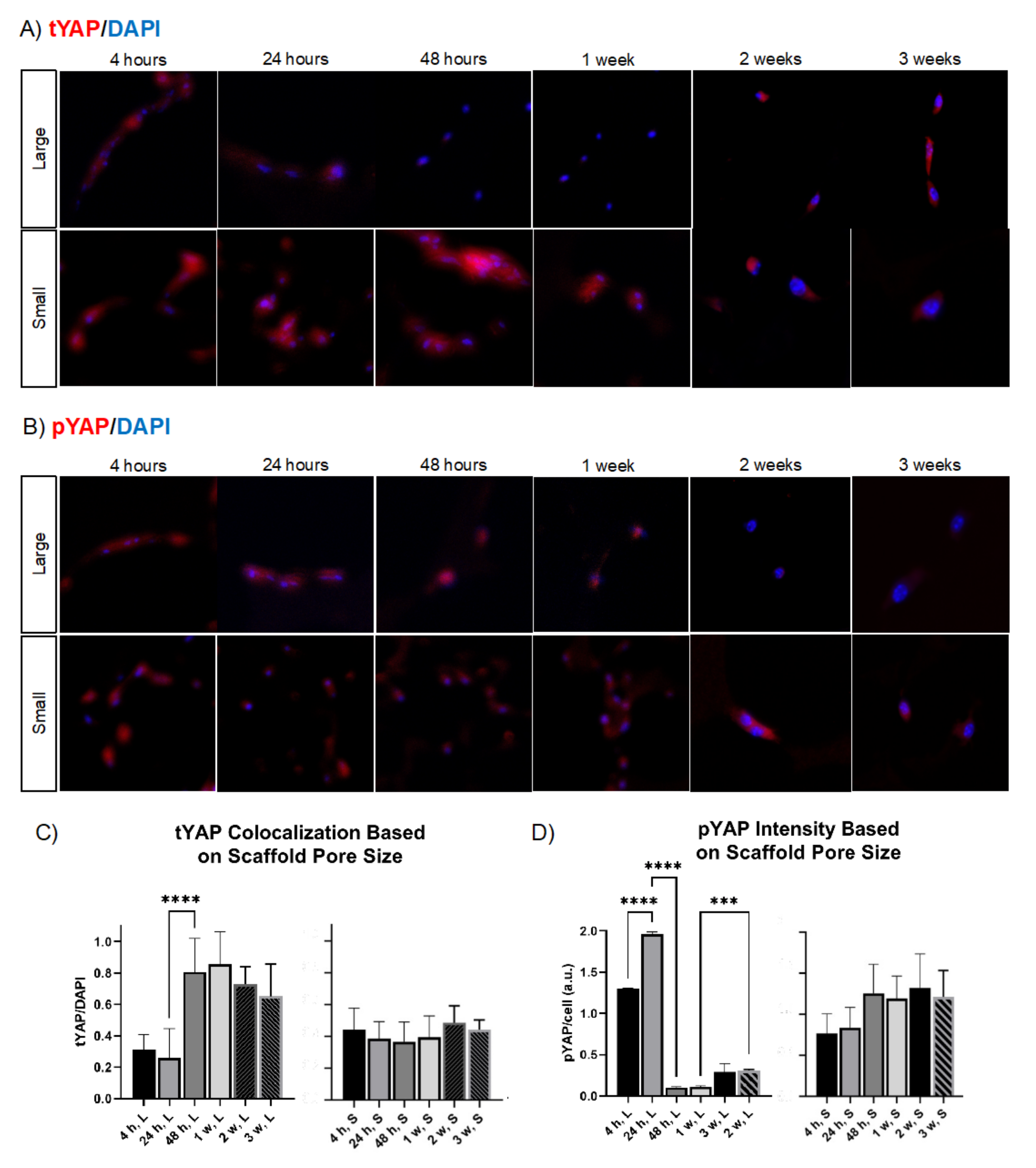
2.6. Curvature Differentially Regulates YAP Nuclear Translocation and Its Cytosolic Phosphorylation In Vitro in Cell-Scaffold Constructs
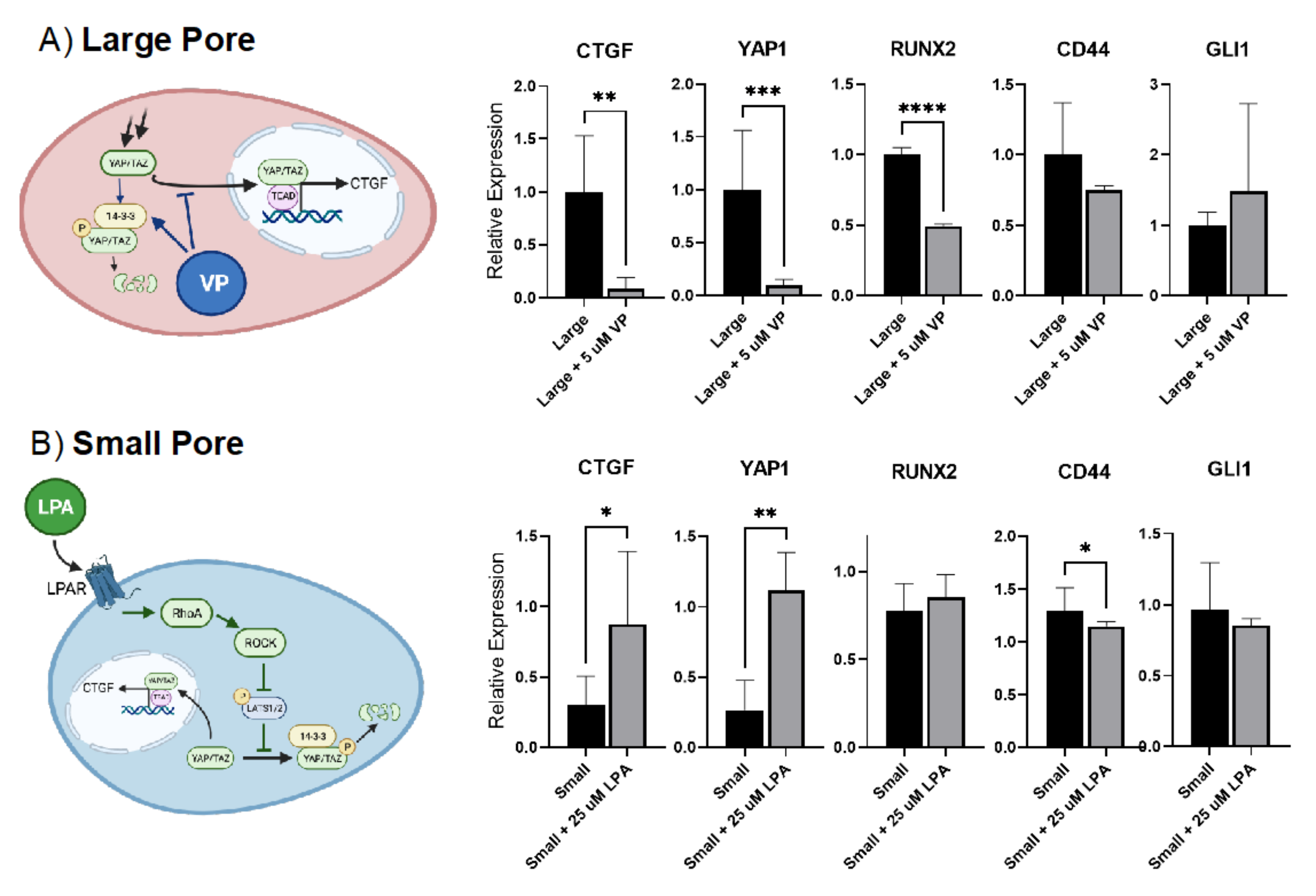
2.7. Pharmacologic Intervention Highlights the Role of YAP/TAZ Signaling to Regulate Gene Expression Where Curvature-Mediated Cytoskeletal Strain Is the Major Driver of Regenerative Fate
3. Discussion
4. Materials and Methods
4.1. Materials
4.1.1. Pharmacologic Reagents
4.1.2. Antibodies and Immunofluorescence Reagents
4.1.3. Cell Culture Reagents
4.1.4. Gene Expression Analysis Primer Sequences
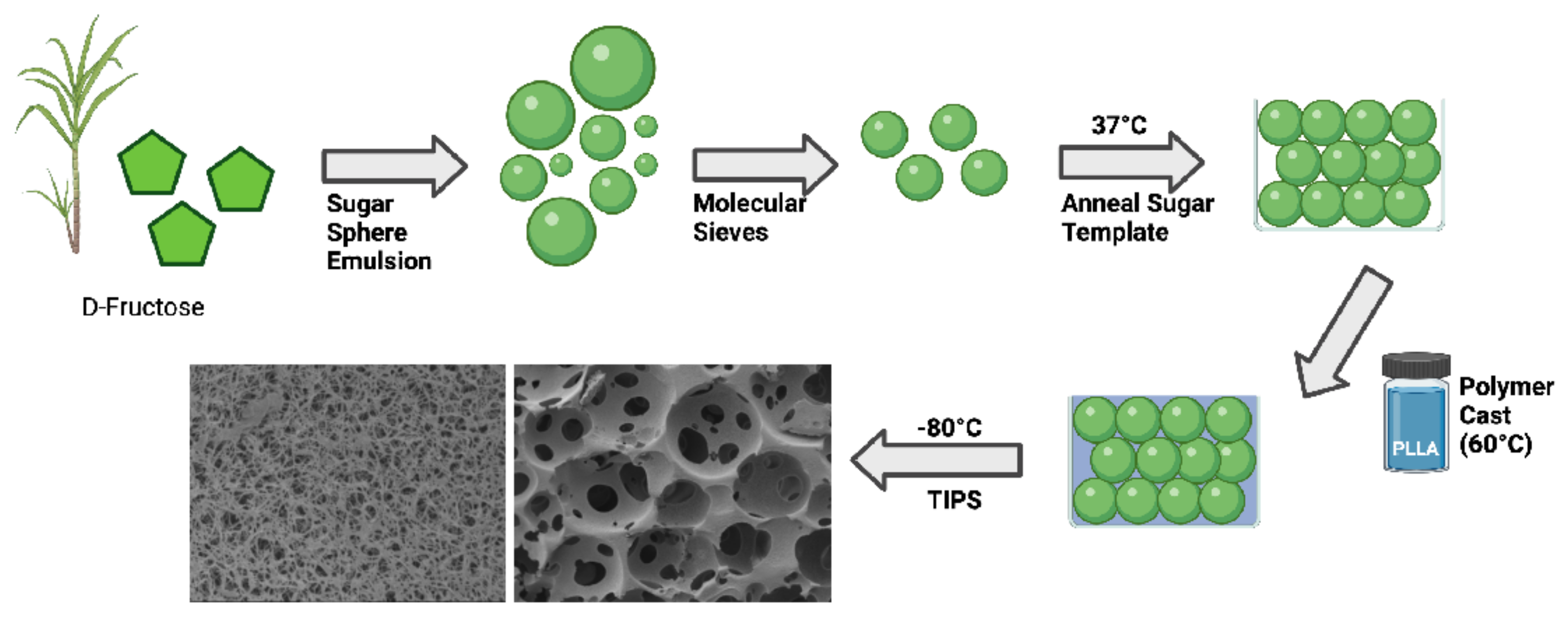
4.2. Fabrication of Nanofibrous, Macroporous Tissue Engineering Scaffolds
4.3. Scanning Electron Microscopy (SEM)
4.4. Cell Isolation
4.4.1. Bone Marrow Mesenchymal Stem Cells
4.4.2. Suture Mesenchymal Stem Cells
4.5. Cell Culture on Scaffolds
4.5.1. Cell Seeding
4.5.2. Cell Culture
4.5.3. Administration of Cytochalasin D
4.5.4. Administration of Verteporfin
4.5.5. Administration of Lysophosphatidic Acid
4.6. Subcutaneous Implantation in Mice
4.7. Histologic Preparation
4.8. Quantitative Real Time Polymerase Chain Reaction (Gene Expression Assay)
4.9. Pore-Mimetic Imaging Platform for Three-Dimensional Visualization
4.10. Immunohistochemistry
4.10.1. Immunohistochemistry for Fluorescence Microscopy
4.10.2. Western Blot
4.11. Confocal Laser Microscopy
4.12. Image Analysis
4.12.1. Analysis of Macropores from SEM Images
4.12.2. Spatial Point Pattern Analysis
4.12.3. Nuclear Morphology in Scaffold Sections
4.12.4. Quantification of Cell Clustering in Scaffold Sections
4.12.5. Quantification of YAP-Nuclear Colocalization
4.12.6. Quantification of YAP Phosphorylation
4.13. Statistical Analysis
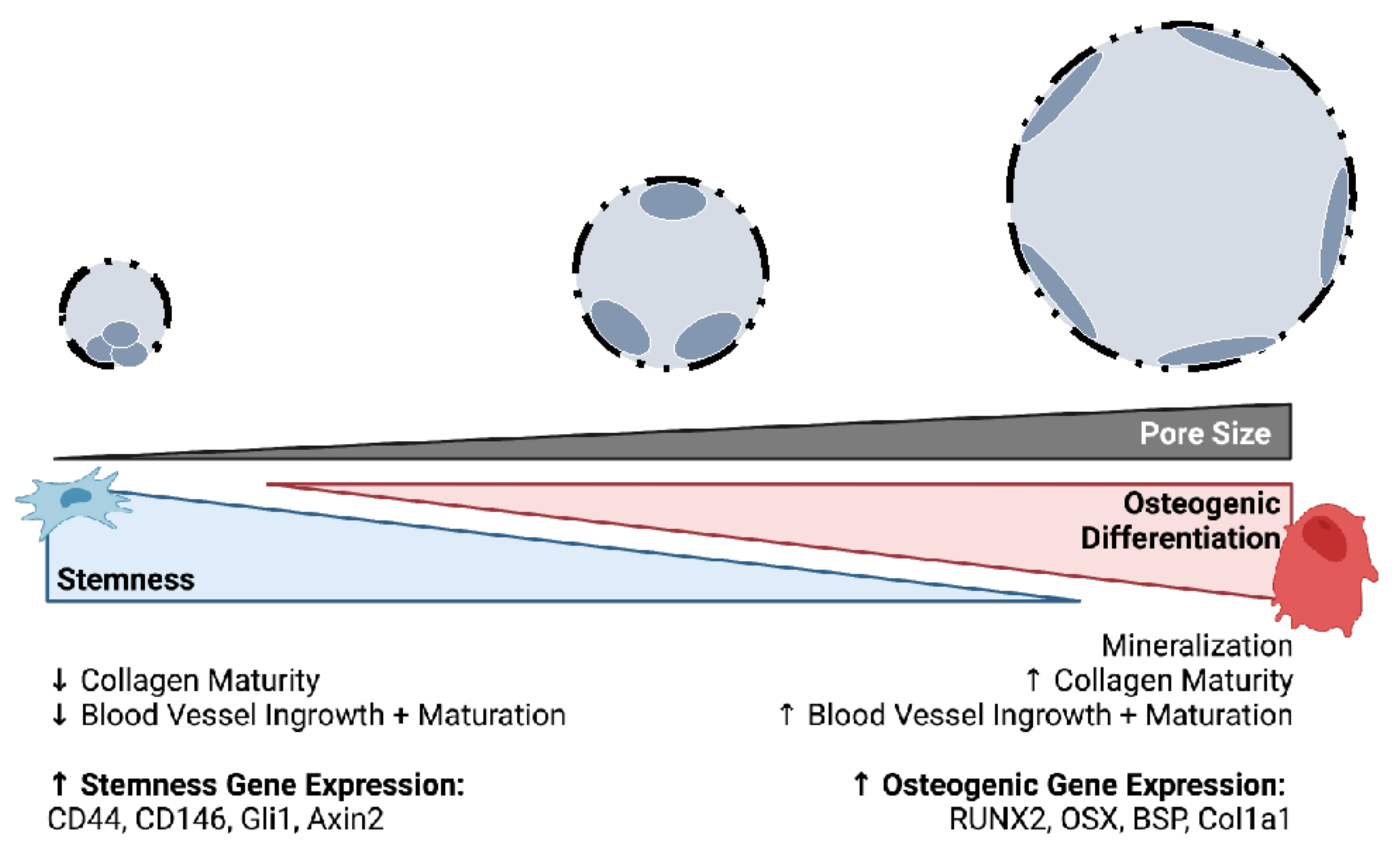
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Langer, R.; Vacanti, J.P. Tissue engineering. Science 1993, 260, 920–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langer, R.; Vacanti, J. Advances in tissue engineering. J. Pediatr. Surg. 2016, 51, 8–12. [Google Scholar] [CrossRef] [Green Version]
- Ma, P.X. Biomimetic materials for tissue engineering. Adv. Drug Deliv. Rev. 2008, 60, 184–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swanson, W.B.; Ma, P.X. Nanofibrous and Porous Biomaterials. In Biomaterials Science, An Introduction to Materials in Medicine; Zhang, G., Ed.; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Zhang, R.Y.; Ma, P.X. Porous poly(L-lactic acid)/apatite composites created by biomimetic process. J. Biomed. Mater. Res. 1999, 45, 285–293. [Google Scholar] [CrossRef]
- Woo, K.M.; Chen, V.J.; Ma, P.X. Nano-fibrous scaffolding architecture selectively enhances protein adsorption contributing to cell attachment. J. Biomed. Mater. Res. A 2003, 67, 531–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, L.A.; Liu, X.; Hu, J.; Ma, P.X. The Enhancement of human embryonic stem cell osteogenic differentiation with nano-fibrous scaffolding. Biomaterials 2010, 31, 5526–5535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Ma, H.; Jin, X.; Hu, J.; Liu, X.; Ni, L.; Ma, P.X. The effect of scaffold architecture on odontogenic differentiation of human dental pulp stem cells. Biomaterials 2011, 32, 7822–7830. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Smith, L.A.; Hu, J.; Ma, P.X. Biomimetic nanofibrous gelatin/apatite composite scaffolds for bone tissue engineering. Biomaterials 2009, 30, 2252–2258. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.A.; Liu, X.; Hu, J.; Wang, P.; Ma, P.X. Enhancing Osteogenic Differentiation of Mouse Embryonic Stem Cells by Nanofibers. Tissue Eng. Part A 2009, 15, 1855–1864. [Google Scholar] [CrossRef]
- Boyan, B.D.; Sylvia, V.L.; Liu, Y.; Sagun, R.; Cochran, D.L.; Lohmann, C.H.; Dean, D.D.; Schwartz, Z. Surface roughness mediates its effects on osteoblasts via protein kinase A and phospholipase A2. Biomaterials 1999, 20, 2305–2310. [Google Scholar] [CrossRef]
- Ito, Y. Surface micropatterning to regulate cell functions. Biomaterials 1999, 20, 2333–2342. [Google Scholar] [CrossRef]
- Nunes, C.R.; Simske, S.J.; Sachdeva, R.; Wolford, L.M. Long-term ingrowth and apposition of porous hydroxylapatite implants. J. Biomed. Mater. Res. 1997, 36, 560–563. [Google Scholar] [CrossRef]
- Chalasani, R.; Poole-Warren, L.; Conway, R.M.; Ben-Nissan, B. Porous Orbital Implants in Enucleation: A Systematic Review. Surv. Ophthalmol. 2007, 52, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.X. Tissue Engineering. In Encyclopedia of Polymer Science and Technology, 3rd ed.; Kroschwitz, J.I., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2005; Volume 12, pp. 261–291. [Google Scholar]
- Alberts, B. Molecular Biology of the Cell, 4th ed.; Garland Science: New York, NY, USA, 2002; p. xxxiv. 1548p. [Google Scholar]
- Lu, X.; Ding, Z.; Xu, F.; Lu, Q.; Kaplan, D.L. Subtle Regulation of Scaffold Stiffness for the Optimized Control of Cell Behavior. ACS Appl. Bio Mater. 2019, 2, 3108–3119. [Google Scholar] [CrossRef]
- VijayaVenkataRaman, S.; Shuo, Z.; Fuh, J.Y.H.; Lu, W.F. Design of Three-Dimensional Scaffolds with Tunable Matrix Stiffness for Directing Stem Cell Lineage Specification: An In Silico Study. Bioengineering 2017, 4, 66. [Google Scholar] [CrossRef] [Green Version]
- Swanson, W.B.; Gong, T.; Zhang, Z.; Eberle, M.; Niemann, D.; Dong, R.; Rambhia, K.J.; Ma, P.X. Controlled release of odontogenic exosomes from a biodegradable vehicle mediates dentinogenesis as a novel biomimetic pulp capping therapy. J. Control. Release 2020, 324, 679–694. [Google Scholar] [CrossRef]
- Swanson, W.B.; Zhang, Z.; Xiu, K.; Gong, T.; Eberle, M.; Wang, Z.; Ma, P.X. Scaffolds with controlled release of pro-mineralization exosomes to promote craniofacial bone healing without cell transplantation. Acta Biomater. 2020, 118, 215–232. [Google Scholar] [CrossRef]
- Zhuo, S.; Zhang, F.; Yu, J.; Zhang, X.; Yang, G.; Liu, X. pH-Sensitive Biomaterials for Drug Delivery. Molecules 2020, 25, 5649. [Google Scholar] [CrossRef]
- Zou, Y.; Zhang, L.; Yang, L.; Zhu, F.; Ding, M.; Lin, F.; Wang, Z.; Li, Y. “Click” chemistry in polymeric scaffolds: Bioactive materials for tissue engineering. J. Control. Release 2018, 273, 160–179. [Google Scholar] [CrossRef]
- Scantlebury, T.; Ambruster, J. The Development of Guided Regeneration: Making the Impossible Possible and the Unpredictable Predictable. J. Évid.-Based Dent. Pract. 2012, 12, 101–117. [Google Scholar] [CrossRef]
- Ayala, R.; Zhang, C.; Yang, D.; Hwang, Y.; Aung, A.; Shroff, S.S.; Arce, F.T.; Lal, R.; Arya, G.; Varghese, S. Engineering the cell–material interface for controlling stem cell adhesion, migration, and differentiation. Biomaterials 2011, 32, 3700–3711. [Google Scholar] [CrossRef]
- Changqing, Z.; Guan, J.; Zhang, C. Mesenchymal stem cells: Mechanisms and role in bone regeneration. Postgrad. Med. J. 2014, 90, 643–647. [Google Scholar] [CrossRef]
- Zhao, H.; Feng, J.; Ho, T.-V.; Grimes, W.; Urata, M.; Chai, Y. The suture provides a niche for mesenchymal stem cells of craniofacial bones. Nat. Cell Biol. 2015, 17, 386–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupte, M.J.; Swanson, W.B.; Hu, J.; Jin, X.; Ma, H.; Zhang, Z.; Liu, Z.; Feng, K.; Feng, G.; Xiao, G.; et al. Pore size directs bone marrow stromal cell fate and tissue regeneration in nanofibrous macroporous scaffolds by mediating vascularization. Acta Biomater. 2018, 82, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Swanson, W.; Omi, M.; Zhang, Z.; Nam, H.K.; Jung, Y.; Wang, G.; Ma, P.X.; Hatch, N.E.; Mishina, Y. Macropore design of tissue engineering scaffolds regulates mesenchymal stem cell differentiation fate. Biomaterials 2021, 272, 120769. [Google Scholar] [CrossRef]
- Seruya, M.; Oh, A.K.; Boyajian, M.J.; Posnick, J.C.; Myseros, J.S.; Yaun, A.L.; Keating, R.F. Long-Term Outcomes of Primary Craniofacial Reconstruction for Craniosynostosis: A 12-Year Experience. Plast. Reconstr. Surg. 2011, 127, 2397–2406. [Google Scholar] [CrossRef]
- McCarthy, J.G.; Warren, S.M.; Bernstein, J.; Burnett, W.; Cunningham, M.L.; Edmond, J.C.; Figueroa, A.A.; Kapp-Simon, K.A.; Labow, B.I.; Peterson-Falzone, S.J.; et al. Parameters of Care for Craniosynostosis. Cleft Palate-Craniofac. J. 2012, 49, 1–24. [Google Scholar] [CrossRef]
- Yu, M.; Ma, L.; Yuan, Y.; Ye, X.; Montagne, A.; He, J.; Ho, T.-V.; Wu, Y.; Zhao, Z.; Maria, N.S.; et al. Cranial Suture Regeneration Mitigates Skull and Neurocognitive Defects in Craniosynostosis. Cell 2021, 184, 243–256.e18. [Google Scholar] [CrossRef]
- Swanson, W.B.; Mishina, Y. The Biology of the Sutures of the Skull. In The Sutures of the Skull; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2021; pp. 171–199. [Google Scholar]
- Majzoub, J.; Barootchi, S.; Tavelli, L.; Wang, C.-W.; Chan, H.-L.; Wang, H.-L. Guided tissue regeneration combined with bone allograft in infrabony defects: Clinical outcomes and assessment of prognostic factors. J. Periodontol. 2020, 91, 746–755. [Google Scholar] [CrossRef]
- Scheyer, E.T.; Heard, R.; Janakievski, J.; Mandelaris, G.; Nevins, M.L.; Pickering, S.R.; Richardson, C.R.; Pope, B.; Toback, G.; Velásquez, D.; et al. A randomized, controlled, multicentre clinical trial of post-extraction alveolar ridge preservation. J. Clin. Periodontol. 2016, 43, 1188–1199. [Google Scholar] [CrossRef]
- Cochran, D.L.; Cobb, C.M.; Bashutski, J.D.; Chun, Y.-H.P.; Lin, Z.; Mandelaris, G.A.; McAllister, B.S.; Murakami, S.; Rios, H.F. Emerging Regenerative Approaches for Periodontal Reconstruction: A Consensus Report From the AAP Regeneration Workshop. J. Periodontol. 2015, 86, S153–S156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skerry, T.M. The response of bone to mechanical loading and disuse: Fundamental principles and influences on osteoblast/osteocyte homeostasis. Arch. Biochem. Biophys. 2008, 473, 117–123. [Google Scholar] [CrossRef] [PubMed]
- McBride, S.H.; Silva, M.J. Adaptive and injury response of bone to mechanical loading. BoneKEy Rep. 2012, 1, 192. [Google Scholar] [CrossRef] [PubMed]
- Sanchis-Moysi, J.; Dorado, C.; Vicente-Rodríguez, G.; Milutinovic, L.; Garces, G.; Calbet, J. Inter-arm asymmetry in bone mineral content and bone area in postmenopausal recreational tennis players. Maturitas 2004, 48, 289–298. [Google Scholar] [CrossRef]
- Menkes, A.; Mazel, S.; Redmond, R.A.; Koffler, K.; Libanati, C.R.; Gundberg, C.M.; Zizic, T.M.; Hagberg, J.M.; Pratley, R.E.; Hurley, B.F. Strength training increases regional bone mineral density and bone remodeling in middle-aged and older men. J. Appl. Physiol. 1993, 74, 2478–2484. [Google Scholar] [CrossRef]
- Zhang, Z.Q.; Yang, J.L. Biomechanical Dynamics of Cranial Sutures during Simulated Impulsive Loading. Appl. Bionics Biomech. 2015, 2015, 596843. [Google Scholar] [CrossRef] [Green Version]
- Katsianou, M.A.; Adamopoulos, C.; Vastardis, H.; Basdra, E.K. Signaling mechanisms implicated in cranial sutures pathophysiology: Craniosynostosis. BBA Clin. 2016, 6, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Qin, L.; Liu, W.; Cao, H.; Xiao, G. Molecular mechanosensors in osteocytes. Bone Res. 2020, 8, 23. [Google Scholar] [CrossRef]
- Xiao, E.; Chen, C.; Zhang, Y. The mechanosensor of mesenchymal stem cells: Mechanosensitive channel or cytoskeleton? Stem Cell Res. Ther. 2016, 7, 140. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Liang, L.; Dou, Y.; Huang, Z.; Mo, H.; Wang, Y.; Yu, B. Mechanical Strain Regulates Osteogenic and Adipogenic Differentiation of Bone Marrow Mesenchymal Stem Cells. BioMed Res. Int. 2015, 2015, 873251. [Google Scholar] [CrossRef]
- Sun, M.; Chi, G.; Xu, J.; Tan, Y.; Xu, J.; Lv, S.; Xu, Z.; Xia, Y.; Li, L.; Li, Y. Extracellular matrix stiffness controls osteogenic differentiation of mesenchymal stem cells mediated by integrin α5. Stem Cell Res. Ther. 2018, 9, 52. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, P.S.; Loboa, E.G. Cytoskeletal and Focal Adhesion Influences on Mesenchymal Stem Cell Shape, Mechanical Properties, and Differentiation Down Osteogenic, Adipogenic, and Chondrogenic Pathways. Tissue Eng. Part B Rev. 2012, 18, 436–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Nakamoto, T.; Dulińska-Molak, I.; Kawazoe, N.; Chen, G. Regulating the stemness of mesenchymal stem cells by tuning micropattern features. J. Mater. Chem. B 2016, 4, 37–45. [Google Scholar] [CrossRef]
- Guo, T.; Yu, L.; Lim, C.G.; Goodley, A.S.; Xiao, X.; Placone, J.K.; Ferlin, K.M.; Nguyen, B.-N.B.; Hsieh, A.H.; Fisher, J.P. Effect of Dynamic Culture and Periodic Compression on Human Mesenchymal Stem Cell Proliferation and Chondrogenesis. Ann. Biomed. Eng. 2015, 44, 2103–2113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Wang, Y.; Zhou, N.; Feng, Y.; Yang, X. Cyclic tensile stress promotes osteogenic differentiation of adipose stem cells via ERK and p38 pathways. Stem Cell Res. 2019, 37, 101433. [Google Scholar] [CrossRef] [PubMed]
- Kearney, E.M.; Farrell, E.; Prendergast, P.J.; Campbell, V.A. Tensile Strain as a Regulator of Mesenchymal Stem Cell Osteogenesis. Ann. Biomed. Eng. 2010, 38, 1767–1779. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, T.; Cosgrove, B.D.; Heo, S.-J.; Shurden, Z.E.; Mauck, R.L. Cytoskeletal to Nuclear Strain Transfer Regulates YAP Signaling in Mesenchymal Stem Cells. Biophys. J. 2015, 108, 2783–2793. [Google Scholar] [CrossRef] [Green Version]
- Pek, Y.S.; Wan, A.C.; Ying, J.Y. The effect of matrix stiffness on mesenchymal stem cell differentiation in a 3D thixotropic gel. Biomaterials 2010, 31, 385–391. [Google Scholar] [CrossRef]
- Baddeley, A.; Turner, R. Spatstat: An R Package for Analyzing Spatial Point Patterns. J. Stat. Softw. 2005, 12, 1–42. [Google Scholar] [CrossRef] [Green Version]
- Bivand, R.S.; Pebesma, E.J.; Gómez-Rubio, V. Spatial Point Pattern Analysis. In Applied Spatial Data Analysis with R; Springer: New York, NY, USA, 2008; pp. 155–190. [Google Scholar]
- Zhao, B.; Li, L.; Wang, L.; Wang, C.-Y.; Yu, J.; Guan, K.-L. Cell detachment activates the Hippo pathway via cytoskeleton reorganization to induce anoikis. Genes Dev. 2012, 26, 54–68. [Google Scholar] [CrossRef] [Green Version]
- Wada, K.-I.; Itoga, K.; Okano, T.; Yonemura, S.; Sasaki, H. Hippo pathway regulation by cell morphology and stress fibers. Development 2011, 138, 3907–3914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rausch, V.; Hansen, C.G. The Hippo Pathway, YAP/TAZ, and the Plasma Membrane. Trends Cell Biol. 2020, 30, 32–48. [Google Scholar] [CrossRef] [PubMed]
- Opperman, L.A.; Rawlins, J.T. The Extracellular Matrix Environment in Suture Morphogenesis and Growth. Cells Tissues Organs 2005, 181, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Regelsberger, J.; Milovanovic, P.; Schmidt, T.; Hahn, M.; Zimmermann, E.A.; Tsokos, M.; Zustin, J.; Ritchie, R.O.; Amling, M.; Busse, B. Changes to the cell, tissue and architecture levels in cranial suture synostosis reveal a problem of timing in bone development. Eur. Cells Mater. 2012, 24, 441–458. [Google Scholar] [CrossRef] [PubMed]
- Schlesinger, P.H.; Blair, H.C.; Stolz, D.B.; Riazanski, V.; Ray, E.C.; Tourkova, I.L.; Nelson, D.J. Cellular and extracellular matrix of bone, with principles of synthesis and dependency of mineral deposition on cell membrane transport. Am. J. Physiol. Physiol. 2020, 318, C111–C124. [Google Scholar] [CrossRef]
- Kegelman, C.D.; Collins, J.M.; Nijsure, M.P.; Eastburn, E.A.; Boerckel, J.D. Gone Caving: Roles of the Transcriptional Regulators YAP and TAZ in Skeletal Development. Curr. Osteoporos. Rep. 2020, 18, 526–540. [Google Scholar] [CrossRef]
- Hoffman, B.D.; Grashoff, C.; Schwartz, M.A. Dynamic molecular processes mediate cellular mechanotransduction. Nature 2011, 475, 316–323. [Google Scholar] [CrossRef]
- Virdi, J.K.; Pethe, P. Biomaterials Regulate Mechanosensors YAP/TAZ in Stem Cell Growth and Differentiation. Tissue Eng. Regen. Med. 2021, 18, 199–215. [Google Scholar] [CrossRef]
- Brusatin, G.; Panciera, T.; Gandin, A.; Citron, A.; Piccolo, S. Biomaterials and engineered microenvironments to control YAP/TAZ-dependent cell behaviour. Nat. Mater. 2018, 17, 1063–1075. [Google Scholar] [CrossRef]
- Wei, G.; Ma, P.X. Macroporous and nanofibrous polymer scaffolds and polymer/bone-like apatite composite scaffolds generated by sugar spheres. J. Biomed. Mater. Res. Part A 2006, 78A, 306–315. [Google Scholar] [CrossRef] [Green Version]
- Piccolo, S.; Dupont, S.; Cordenonsi, M. The Biology of YAP/TAZ: Hippo Signaling and Beyond. Physiol. Rev. 2014, 94, 1287–1312. [Google Scholar] [CrossRef] [PubMed]
- Kegelman, C.D.; Mason, D.E.; Dawahare, J.H.; Horan, D.J.; Vigil, G.D.; Howard, S.S.; Robling, A.G.; Bellido, T.M.; Boerckel, J.D. Skeletal cell YAP and TAZ combinatorially promote bone development. FASEB J. 2018, 32, 2706–2721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Y.; Feinberg, T.; Keller, E.T.; Li, X.-Y.; Weiss, S.J. Snail/Slug binding interactions with YAP/TAZ control skeletal stem cell self-renewal and differentiation. Nat. Cell Biol. 2016, 18, 917–929. [Google Scholar] [CrossRef] [PubMed]
- Frank, V.; Kaufmann, S.; Wright, R.; Horn, P.; Yoshikawa, H.Y.; Wuchter, P.; Madsen, J.; Lewis, A.L.; Armes, S.P.; Ho, A.D.; et al. Frequent mechanical stress suppresses proliferation of mesenchymal stem cells from human bone marrow without loss of multipotency. Sci. Rep. 2016, 6, 24264. [Google Scholar] [CrossRef] [PubMed]
- Mohri, Z.; Hernandez, A.D.R.; Krams, R. The emerging role of YAP/TAZ in mechanotransduction. J. Thorac. Dis. 2017, 9, E507–E509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Zhao, J.; Wang, J.; Sun, L.; Xu, H.; Sun, W.; Pan, Y.; Wang, H.; Zhang, W.-B. ROCK-TAZ signaling axis regulates mechanical tension-induced osteogenic differentiation of rat cranial sagittal suture mesenchymal stem cells. J. Cell. Physiol. 2020, 235, 5972–5984. [Google Scholar] [CrossRef]
- Kovar, H.; Bierbaumer, L.; Radic-Sarikas, B. The YAP/TAZ Pathway in Osteogenesis and Bone Sarcoma Pathogenesis. Cells 2020, 9, 972. [Google Scholar] [CrossRef] [Green Version]
- Bruderer, M.; Richards, R.G.; Alini, M.; Stoddart, M.J. Role and regulation of RUNX2 in osteogenesis. Eur. Cell Mater. 2014, 28, 269–286. [Google Scholar] [CrossRef]
- Xiao, H.; Tong, R.; Yang, B.; Lv, Z.; Du, C.; Peng, C.; Ding, C.; Cheng, S.; Zhou, L.; Xie, H.; et al. TAZ regulates cell proliferation and sensitivity to vitamin D3 in intrahepatic cholangiocarcinoma. Cancer Lett. 2016, 381, 370–379. [Google Scholar] [CrossRef]
- Laird, E.; Ward, M.; McSorley, E.; Strain, J.; Wallace, J. Vitamin D and Bone Health; Potential Mechanisms. Nutrients 2010, 2, 693–724. [Google Scholar] [CrossRef] [Green Version]
- Hankenson, K.D.; Dishowitz, M.; Gray, C.; Schenker, M. Angiogenesis in bone regeneration. Injury 2011, 42, 556–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooglugt, A.; van der Stoel, M.M.; Boon, R.A.; Huveneers, S. Endothelial YAP/TAZ Signaling in Angiogenesis and Tumor Vasculature. Front. Oncol. 2021, 10, 3162. [Google Scholar] [CrossRef]
- Scott, K.E.; Fraley, S.I.; Rangamani, P. A spatial model of YAP/TAZ signaling reveals how stiffness, dimensionality, and shape contribute to emergent outcomes. Proc. Natl. Acad. Sci. USA 2021, 118, e2021571118. [Google Scholar] [CrossRef] [PubMed]
- Costello, P.; Sargent, M.; Maurice, D.; Esnault, C.; Foster, K.; Anjos-Afonso, F.; Treisman, R. MRTF-SRF signaling is required for seeding of HSC/Ps in bone marrow during development. Blood 2015, 125, 1244–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, T.; Gao, B.; Fan, Y.; Liu, Y.; Feng, S.; Cong, Q.; Zhang, X.; Zhou, Y.; Yadav, P.S.; Lin, J.; et al. Piezo1/2 mediate mechanotransduction essential for bone formation through concerted activation of NFAT-YAP1-ß-catenin. eLife 2020, 9, e52779. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Xu, L.; Sun, Y.; Wu, T.; Wang, K.; Li, G. An improved protocol for isolation and culture of mesenchymal stem cells from mouse bone marrow. J. Orthop. Transl. 2015, 3, 26–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amend, S.R.; Valkenburg, K.C.; Pienta, K.J. Murine Hind Limb Long Bone Dissection and Bone Marrow Isolation. J. Vis. Exp. 2016, 110, e53936. [Google Scholar] [CrossRef] [Green Version]
- Palchesko, R.N.; Zhang, L.; Sun, Y.; Feinberg, A.W. Development of Polydimethylsiloxane Substrates with Tunable Elastic Modulus to Study Cell Mechanobiology in Muscle and Nerve. PLoS ONE 2012, 7, e51499. [Google Scholar] [CrossRef] [Green Version]














Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Swanson, W.B.; Omi, M.; Woodbury, S.M.; Douglas, L.M.; Eberle, M.; Ma, P.X.; Hatch, N.E.; Mishina, Y. Scaffold Pore Curvature Influences ΜSC Fate through Differential Cellular Organization and YAP/TAZ Activity. Int. J. Mol. Sci. 2022, 23, 4499. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094499
Swanson WB, Omi M, Woodbury SM, Douglas LM, Eberle M, Ma PX, Hatch NE, Mishina Y. Scaffold Pore Curvature Influences ΜSC Fate through Differential Cellular Organization and YAP/TAZ Activity. International Journal of Molecular Sciences. 2022; 23(9):4499. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094499
Chicago/Turabian StyleSwanson, W. Benton, Maiko Omi, Seth M. Woodbury, Lindsey M. Douglas, Miranda Eberle, Peter X. Ma, Nan E. Hatch, and Yuji Mishina. 2022. "Scaffold Pore Curvature Influences ΜSC Fate through Differential Cellular Organization and YAP/TAZ Activity" International Journal of Molecular Sciences 23, no. 9: 4499. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094499