
Ac/Ds-Induced Receptor-like Kinase Genes Deletion Provides Broad-Spectrum Resistance to Bacterial Blight in Rice


Abstract
:1. Introduction
2. Results
2.1. A Pair of Ds Elements Generate Diverse Chromosomal Rearrangements
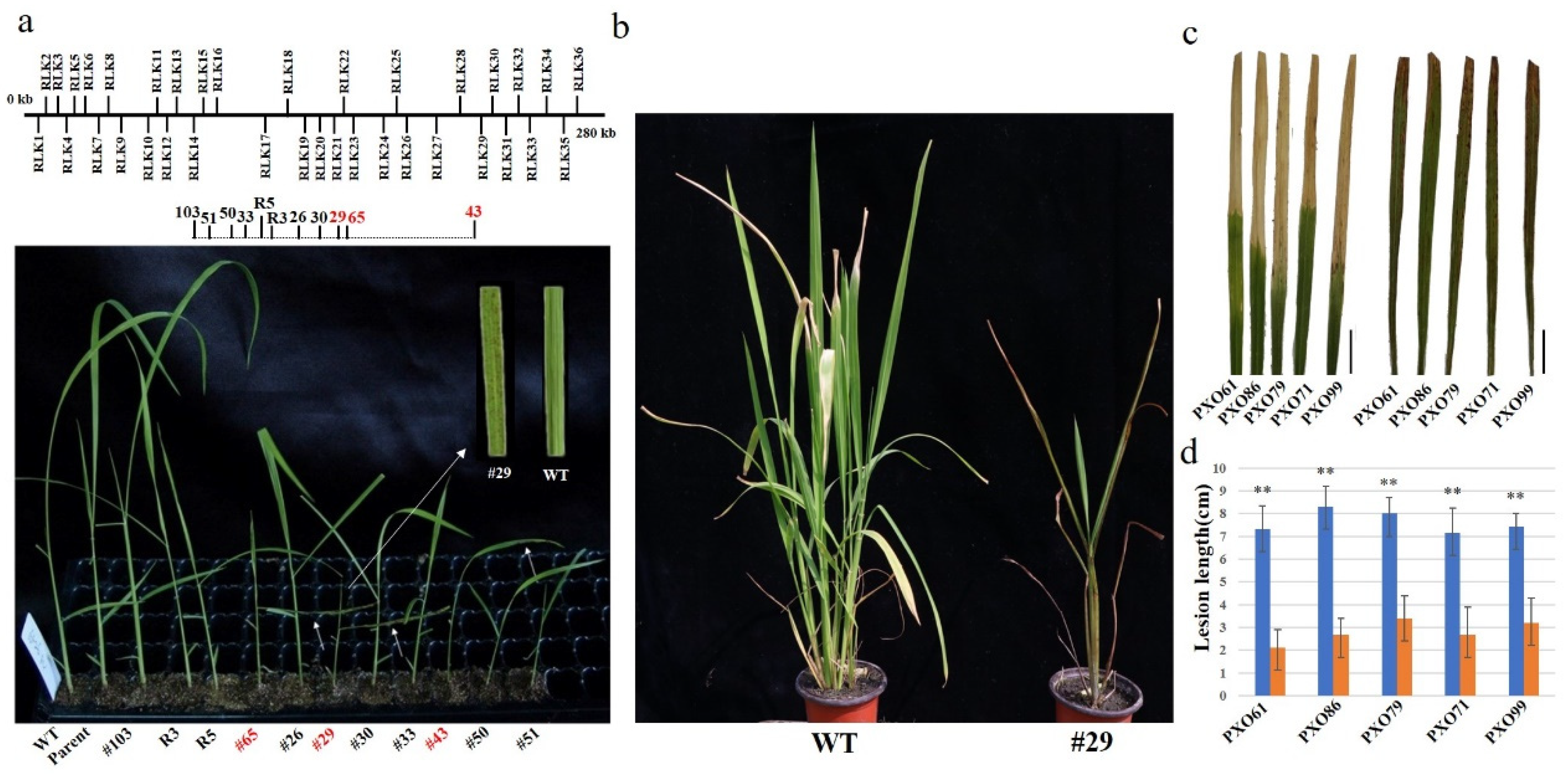
2.2. Ac/Ds-Induced RLK Deletion Mutants Exhibited Broad-Spectrum Resistance to Xoo
2.3. RLK20, RLK21, and RLK22 Regulate the Broad-Spectrum Resistance to Xoo in Rice
2.4. RLK20 Interacts with RBOHB to Modulate ROS Generation
3. Discussion
4. Materials and Methods
4.1. Tissue Culture Regeneration and Transgenic Plant Generation
4.2. Rice Cultivation and Xoo Inoculation
4.3. Determination of H2O2 Content
4.4. qRT-PCR Analysis
4.5. DAB Staining Assay
4.6. Yeast Two-Hybrid Screening
4.7. BiFC Assay
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Niño-Liu, D.O.; Ronald, P.C.; Bogdanove, A.J. Xanthomonas oryzae pathovars: Model pathogens of a model crop. Mol. Plant Pathol. 2006, 7, 303–324. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Liu, H.; Zhou, Y.; Zhang, Q.; Li, X.; Wang, S. Exploring the mechanism and efficient use of a durable gene-mediated resistance to bacterial blight disease in rice. Mol. Breed. 2018, 38, 18. [Google Scholar] [CrossRef]
- Hu, K.M.; Cao, J.B.; Zhang, J.; Xia, F.; Ke, Y.G.; Zhang, H.T.; Xie, W.Y.; Liu, H.B.; Cui, Y.; Cao, Y.L.; et al. Improvement of multiple agronomic traits by a disease resistance gene via cell wall reinforcement. Nat. Plants 2017, 3, 17009. [Google Scholar] [CrossRef] [PubMed]
- Busungu, C.; Taura, S.; Sakagami, J.I.; Ichitani, K. Identification and linkage analysis of a new rice bacterial blight resistance gene from XM14, a mutant line from IR24. Breed. Sci. 2016, 66, 636–645. [Google Scholar] [CrossRef] [Green Version]
- Ji, Z.; Wang, C.; Zhao, K. Rice Routes of Countering Xanthomonas oryzae. Int. J. Mol. Sci. 2018, 19, 3008. [Google Scholar] [CrossRef] [Green Version]
- Jiang, N.; Yan, J.; Liang, Y.; Shi, Y.; He, Z.; Wu, Y.; Zeng, Q.; Liu, X.; Peng, J. Resistance Genes and their Interactions with Bacterial Blight/Leaf Streak Pathogens (Xanthomonas oryzae) in Rice (Oryza sativa L.)—An Updated Review. Rice 2020, 13, 3. [Google Scholar] [CrossRef] [Green Version]
- Xiang, Y.; Cao, Y.; Xu, C.; Li, X.; Wang, S. Xa3, conferring resistance for rice bacterial blight and encoding a receptor kinase-like protein, is the same as Xa26. TAG. Theor. Appl. Genet. Theor. Und Angew. Genet. 2006, 113, 1347–1355. [Google Scholar] [CrossRef]
- Song, W.-Y.; Wang, G.-L.; Chen, L.-L.; Kim, H.-S.; Pi, L.-Y.; Holsten, T.; Gardner, J.; Wang, B.; Zhai, W.-X.; Zhu, L.-H.; et al. A Receptor Kinase-Like Protein Encoded by the Rice Disease Resistance Gene, Xa21. Science 1995, 270, 1804–1806. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Cao, Y.; Yang, Z.; Xu, C.; Li, X.; Wang, S.; Zhang, Q. Xa26, a gene conferring resistance to Xanthomonas oryzae pv. oryzae in rice, encodes an LRR receptor kinase-like protein. Plant J. Cell Mol. Biol. 2004, 37, 517–527. [Google Scholar] [CrossRef]
- Liu, Y.; Cao, Y.; Zhang, Q.; Li, X.; Wang, S. A Cytosolic Triosephosphate Isomerase Is a Key Component in XA3/XA26-Mediated Resistance. Plant Physiol 2018, 178, 923–935. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Zuo, S.; Schwessinger, B.; Chern, M.; Canlas, P.E.; Ruan, D.; Zhou, X.; Wang, J.; Daudi, A.; Petzold, C.J.; et al. An XA21-Associated Kinase (OsSERK2) Regulates Immunity Mediated by the XA21 and XA3 Immune Receptors. Mol. Plant 2014, 7, 874–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luu, D.D.; Joe, A.; Chen, Y.; Parys, K.; Bahar, O.; Pruitt, R.; Chan, L.J.G.; Petzold, C.J.; Long, K.; Adamchak, C.; et al. Biosynthesis and secretion of the microbial sulfated peptide RaxX and binding to the rice XA21 immune receptor. Proc. Natl. Acad. Sci. USA 2019, 116, 8525–8534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Century, K.S.; Lagman, R.A.; Adkisson, M.; Morlan, J.; Tobias, R.; Schwartz, K.; Smith, A.; Love, J.; Ronald, P.C.; Whalen, M.C. Developmental control of Xa21-mediated disease resistance in rice. Plant J. 1999, 20, 231–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruitt, R.N.; Schwessinger, B.; Joe, A.; Thomas, N.; Liu, F.; Albert, M.; Robinson, M.R.; Chan, L.J.G.; Luu, D.D.; Chen, H.; et al. The rice immune receptor XA21 recognizes a tyrosine-sulfated protein from a Gram-negative bacterium. Sci. Adv. 2015, 1, e1500245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwessinger, B.; Li, X.; Ellinghaus, T.L.; Chan, L.J.; Wei, T.; Joe, A.; Thomas, N.; Pruitt, R.; Adams, P.D.; Chern, M.S.; et al. A second-generation expression system for tyrosine-sulfated proteins and its application in crop protection. Integr. Biol. Quant. Biosci. Nano Macro 2016, 8, 542–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, T.; Chern, M.S.; Liu, F.R.; Ronald, P.C. Suppression of bacterial infection in rice by treatment with a sulfated peptide. Mol. Plant Pathol. 2016, 17, 1493–1498. [Google Scholar] [CrossRef] [Green Version]
- Pruitt, R.N.; Joe, A.; Zhang, W.G.; Feng, W.; Stewart, V.; Schwessinger, B.; Dinneny, J.R.; Ronald, P.C. A microbially derived tyrosine-sulfated peptide mimics a plant peptide hormone. New Phytol. 2017, 215, 725–736. [Google Scholar] [CrossRef] [Green Version]
- Leach, J.E.; Vera Cruz, C.M.; Bai, J.; Leung, H. Pathogen fitness penalty as a predictor of durability of disease resistance genes. Annu. Rev. Phytopathol. 2001, 39, 187–224. [Google Scholar] [CrossRef]
- Liu, H.B.; Li, X.H.; Xiao, J.H.; Wang, S.P. A convenient method for simultaneous quantification of multiple phytohormones and metabolites: Application in study of rice-bacterium interaction. Plant Methods 2012, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ke, Y.G.; Liu, H.B.; Li, X.H.; Xiao, J.H.; Wang, S.P. Rice OsPAD4 functions differently from Arabidopsis AtPAD4 in host- pathogen interactions. Plant J. 2014, 78, 619–631. [Google Scholar] [CrossRef]
- Padmavati, M.; Sakthivel, N.; Thara, K.V.; Reddy, A.R. Differential sensitivity of rice pathogens to growth inhibition by flavonoids. Phytochemistry 1997, 46, 499–502. [Google Scholar] [CrossRef]
- Zhang, J.; Yin, Z.; White, F. TAL effectors and the executor R genes. Front. Plant Sci 2015, 6, 641. [Google Scholar] [CrossRef] [PubMed]
- Jacques, M.A.; Arlat, M.; Boulanger, A.; Boureau, T.; Carrère, S.; Cesbron, S.; Chen, N.W.; Cociancich, S.; Darrasse, A.; Denancé, N.; et al. Using Ecology, Physiology, and Genomics to Understand Host Specificity in Xanthomonas. Annu. Rev. Phytopathol. 2016, 54, 163–187. [Google Scholar] [CrossRef] [PubMed]
- Mew, T.W.; Vera Cruz, C.M.; Medalla, E.S. Changes in race frequency of Xanthomonas oryzae pv. oryzae in response to rice cultivars planted in the Philippines. Plant Dis. 1992, 76, 1029–1032. [Google Scholar] [CrossRef]
- Mundt, C.C. Durable resistance: A key to sustainable management of pathogens and pests. Infect. Genet. Evol. 2014, 27, 446–455. [Google Scholar] [CrossRef]
- Xuan, Y.H.; Zhang, J.; Peterson, T.; Han, C.-D. Ac/Ds-induced chromosomal rearrangements in rice genomes. Mob. Genet. Elem. 2012, 2, 67–71. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Peterson, T. Genome rearrangements by nonlinear transposons in maize. Genetics 1999, 153, 1403–1410. [Google Scholar] [CrossRef]
- Zhang, J.; Peterson, T. Transposition of reversed Ac element ends generates chromosome rearrangements in maize. Genetics 2004, 167, 1929–1937. [Google Scholar] [CrossRef] [Green Version]
- Xuan, Y.H.; Piao, H.L.; Je, B.I.; Park, S.J.; Park, S.H.; Huang, J.; Zhang, J.B.; Peterson, T.; Han, C.-d. Transposon Ac/Ds-induced chromosomal rearrangements at the rice OsRLG5 locus. Nucleic. Acids Res. 2011, 39, e149. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, F.; Peterson, T. Transposition of reversed Ac element ends generates novel chimeric genes in maize. PLoS Genet 2006, 2, e164. [Google Scholar] [CrossRef] [Green Version]
- Feuillet, C.; Schachermayr, G.; Keller, B. Molecular cloning of a new receptor-like kinase gene encoded at the Lr10 disease resistance locus of wheat. Plant J. 1997, 11, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Yong, Y.; Qiujun, L.; Xinyu, C.; Weifang, L.; Yuwen, F.; Zhengjin, X.; Yuanhua, W.; Xuming, W.; Jie, Z.; Chulang, Y.; et al. Characterization and Proteomic Analysis of Novel Rice Lesion Mimic Mutant with Enhanced Disease Resistance. Rice Sci. 2021, 28, 466–478. [Google Scholar] [CrossRef]
- Zhao, J.; Liu, P.; Li, C.; Wang, Y.; Guo, L.; Jiang, G.; Zhai, W. LMM5.1 and LMM5.4, two eukaryotic translation elongation factor 1A-like gene family members, negatively affect cell death and disease resistance in rice. J. Genet. Genom. 2017, 44, 107–118. [Google Scholar] [CrossRef]
- Zhang, X.B.; Feng, B.-H.; Wang, H.-M.; Xu, X.; Shi, Y.-F.; He, Y.; Chen, Z.; Sathe, A.P.; Shi, L.; Wu, J.-L. A substitution mutation in OsPELOTA confers bacterial blight resistance by activating the salicylic acid pathway. J. Integr. Plant Biol. 2018, 60, 160–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Z.; Chen, J.; Zeng, L.; Goh, M.; Leung, H.; Khush, G.S.; Wang, G.L. Characterizing Rice Lesion Mimic Mutants and Identifying a Mutant with Broad-Spectrum Resistance to Rice Blast and Bacterial Blight. Mol. Plant Microbe. Interact. 2000, 13, 869–876. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Xiang, X.; Zhang, Y.; Cao, Y.; Wang, B.; Zhang, Y.; Wang, C.; Jiang, M.; Duan, W.; Chen, D.; et al. Disruption of OsPHD1, Encoding a UDP-Glucose Epimerase, Causes JA Accumulation and Enhanced Bacterial Blight Resistance in Rice. Int. J. Mol. Sci. 2022, 23, 751. [Google Scholar] [CrossRef]
- Hu, B.; Zhou, Y.; Zhou, Z.; Sun, B.; Zhou, F.; Yin, C.; Ma, W.; Chen, H.; Lin, Y. Repressed OsMESL expression triggers reactive oxygen species-mediated broad-spectrum disease resistance in rice. Plant Biotechnol. J. 2021, 19, 1511–1522. [Google Scholar] [CrossRef]
- Zhao, X.; Qiu, T.; Feng, H.; Yin, C.; Zheng, X.; Yang, J.; Peng, Y.-L.; Zhao, W. A novel glycine-rich domain protein, GRDP1, functions as a critical feedback regulator for controlling cell death and disease resistance in rice. J. Exp. Bot. 2020, 72, 608–622. [Google Scholar] [CrossRef]
- Nagano, M.; Ishikawa, T.; Fujiwara, M.; Fukao, Y.; Kawano, Y.; Kawai-Yamada, M.; Shimamoto, K. Plasma membrane microdomains are essential for Rac1-RbohB/H-mediated immunity in rice. Plant Cell 2016, 28, 1966–1983. [Google Scholar] [CrossRef] [Green Version]
- Wong, H.L.; Pinontoan, R.; Hayashi, K.; Tabata, R.; Yaeno, T.; Hasegawa, K.; Kojima, C.; Yoshioka, H.; Iba, K.; Kawasaki, T. Regulation of rice NADPH oxidase by binding of Rac GTPase to its N-terminal extension. Plant Cell 2007, 19, 4022–4034. [Google Scholar] [CrossRef] [Green Version]
- Chin, H.G.; Choe, M.S.; Lee, S.H.; Park, S.H.; Koo, J.C.; Kim, N.Y.; Lee, J.J.; Oh, B.G.; Yi, G.H.; Kim, S.C.; et al. Molecular analysis of rice plants harboring an Ac/Ds transposable element-mediated gene trapping system. Plant J. Cell Mol. Biol. 1999, 19, 615–623. [Google Scholar] [CrossRef] [Green Version]
- Park, S.J.; Piao, H.L.; Xuan, Y.-h.; Park, S.H.; Je, B.I.; Kim, C.M.; Lee, E.J.; Park, S.H.; Ryu, B.; Lee, K.H. Analysis of intragenic Ds transpositions and excision events generating novel allelic variation in rice. Mol. Cells Springer Sci. Bus. Media BV 2006, 21, 284–293. [Google Scholar]
- Kim, C.M.; Piao, H.L.; Park, S.J.; Chon, N.S.; Je, B.I.; Sun, B.; Park, S.H.; Park, J.Y.; Lee, E.J.; Kim, M.J. Rapid, large-scale generation of Ds transposant lines and analysis of the Ds insertion sites in rice. Plant J. 2004, 39, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Zhang, C.; Han, X.; Wang, Z.Y.; Ma, L.; Yuan, D.P.; Wu, J.N.; Zhu, X.F.; Liu, J.M.; Li, D.P.; et al. Inhibition of OsSWEET11 function in mesophyll cells improves resistance of rice to sheath blight disease. Mol. Plant Pathol. 2018, 19, 2149–2161. [Google Scholar] [CrossRef] [Green Version]
- Mei, Q.; Yang, Y.; Ye, S.; Liang, W.; Wang, X.; Zhou, J.; Yu, C.; Yan, C.; Chen, J. H2O2 Induces Association of RCA with the Thylakoid Membrane to Enhance Resistance of Oryza meyeriana to Xanthomonas oryzae pv. oryzae. Plants 2019, 8, 351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef]
- Gao, Y.; Xue, C.Y.; Liu, J.M.; He, Y.; Mei, Q.; Wei, S.; Xuan, Y.H. Sheath blight resistance in rice is negatively regulated by WRKY53 via SWEET2a activation. Biochem. Biophys. Res. Commun. 2021, 585, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Thordal-Christensen, H.; Zhang, Z.; Wei, Y.; Collinge, D.B. Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley—Powdery mildew interaction. Plant J. 1997, 11, 1187–1194. [Google Scholar] [CrossRef]
- Kim, T.W.; Guan, S.H.; Sun, Y.; Deng, Z.P.; Tang, W.Q.; Shang, J.X.; Sun, Y.; Burlingame, A.L.; Wang, Z.Y. Brassinosteroid signal transduction from cell-surface receptor kinases to nuclear transcription factors. Nat. Cell Biol. 2009, 11, 1254–1260. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | LOC Number | Direction | Sequence |
|---|---|---|---|
| RLK19 | LOC_Os01g02570 | Forward | AAGCTTATGGCGATTCCTGGAGC |
| Reverse | GGTACCCTAGTTACTAGCGAATTCAATTG | ||
| RLK20 | LOC_Os01g02580 | Forward | GAGCTCATGGCGATCCCTGGTTCG |
| Reverse | GTTAACTCACTCATCCTCCTCTAAGATTTCA | ||
| RLK21 | LOC_Os01g02590 | Forward | AAGCTTATGGCGATTCATGGTGTGTTTC |
| Reverse | GGTACCTCAACAGAAACCTGCAATCATCTTC | ||
| RLK22 | LOC_Os01g02600 | Forward | AAGCTTATGGACTTCACCAACCTTCTTATCA |
| Reverse | GTTAACCTAAATCACAAGTTGATTTTGAGACG |
| Gene | LOC Number | Direction | Sequences |
|---|---|---|---|
| PR1a | LOC_Os07g03710 | Forward Reverse | GTGGGTGTCGGAGAAGCAGTG CGGCGAGTAGTTGCAGGTGAT |
| PBZ1 | LOC_Os12g36880 | Forward Reverse | TGGTCCGGGCACCATCTA CGAGCACATCCGACTTTAGG |
| RLK19 | LOC_Os01g02570 | Forward Reverse | TTGTATCAGACAGGGCATTA CCAGCCATCTCAAGTAGC |
| RLK20 | LOC_Os01g02580 | Forward Reverse | ACGCAATTACTGGAAGATAA TGCCTGGAAGGAGAACAC |
| RLK21 | LOC_Os01g02590 | Forward Reverse | CCGATGACAAGGCTACAA GAAGAGGGCAACTGCTAG |
| RLK22 | LOC_Os01g02600 | Forward Reverse | GTGAGTGGGAGGAGGAAC GCACCATAACGCTACAATA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mei, Q.; Fu, Y.W.; Li, T.M.; Xuan, Y.H. Ac/Ds-Induced Receptor-like Kinase Genes Deletion Provides Broad-Spectrum Resistance to Bacterial Blight in Rice. Int. J. Mol. Sci. 2022, 23, 4561. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094561
Mei Q, Fu YW, Li TM, Xuan YH. Ac/Ds-Induced Receptor-like Kinase Genes Deletion Provides Broad-Spectrum Resistance to Bacterial Blight in Rice. International Journal of Molecular Sciences. 2022; 23(9):4561. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094561
Chicago/Turabian StyleMei, Qiong, Yu Wen Fu, Tian Miao Li, and Yuan Hu Xuan. 2022. "Ac/Ds-Induced Receptor-like Kinase Genes Deletion Provides Broad-Spectrum Resistance to Bacterial Blight in Rice" International Journal of Molecular Sciences 23, no. 9: 4561. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094561