
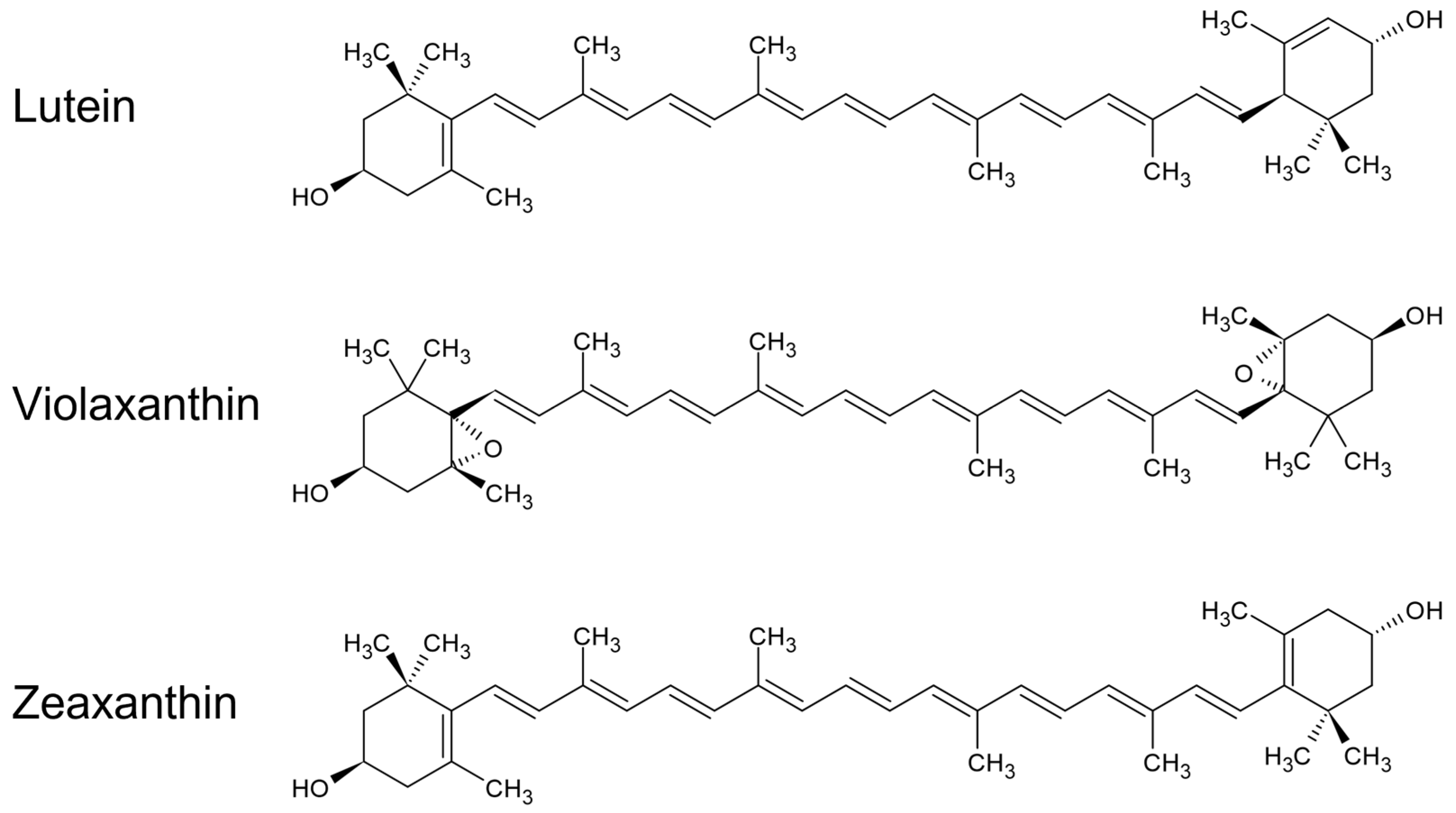
Violaxanthin and Zeaxanthin May Replace Lutein at the L1 Site of LHCII, Conserving the Interactions with Surrounding Chlorophylls and the Capability of Triplet–Triplet Energy Transfer

 , , and
, , and
Abstract
:1. Introduction
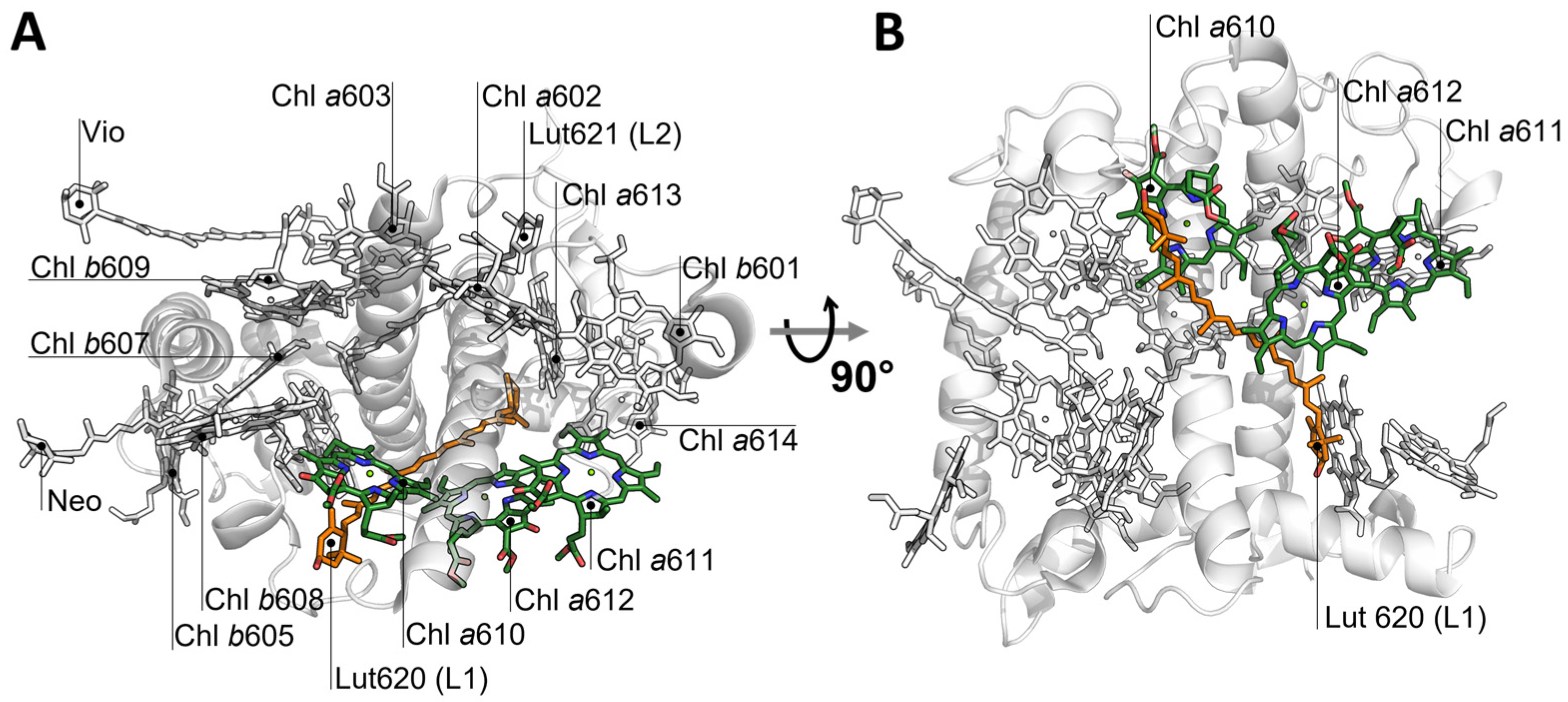
2. Results
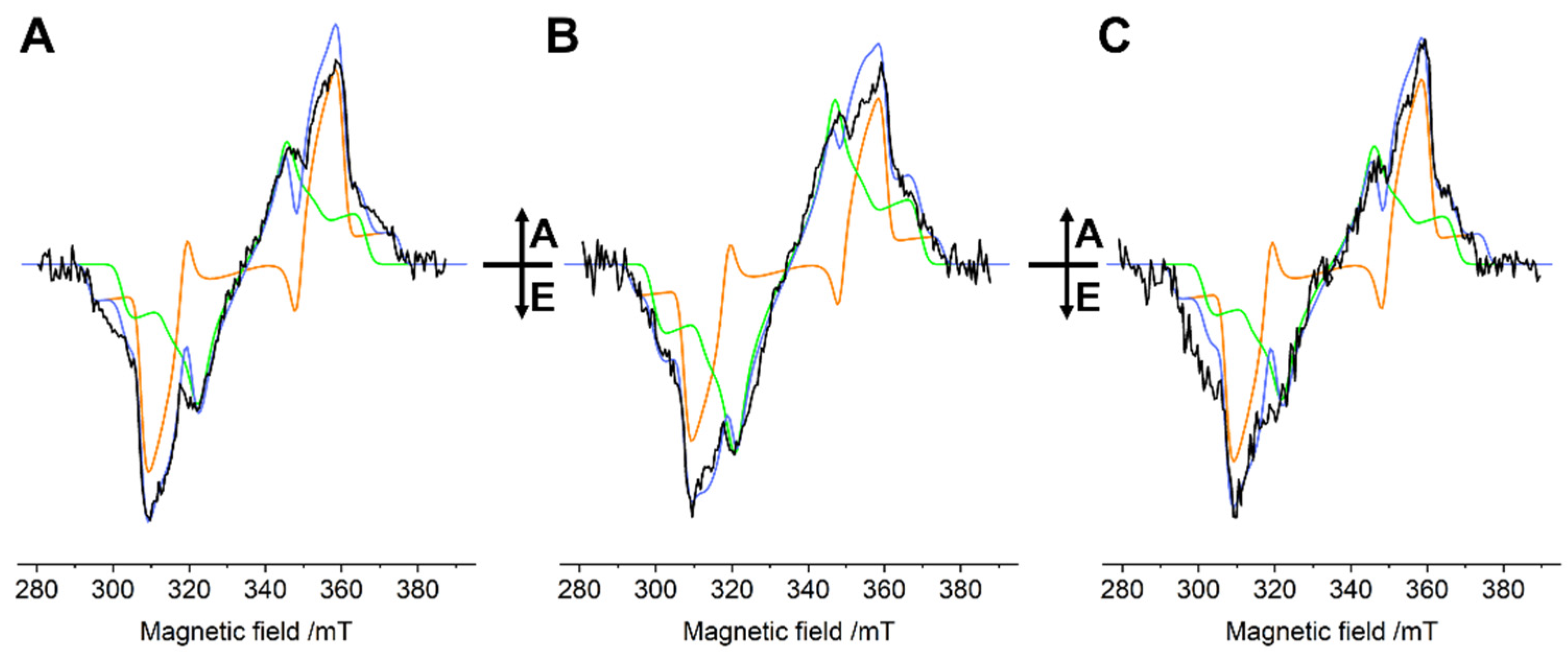
2.1. Time-Resolved Electron Paramagnetic Resonance (TR-EPR)
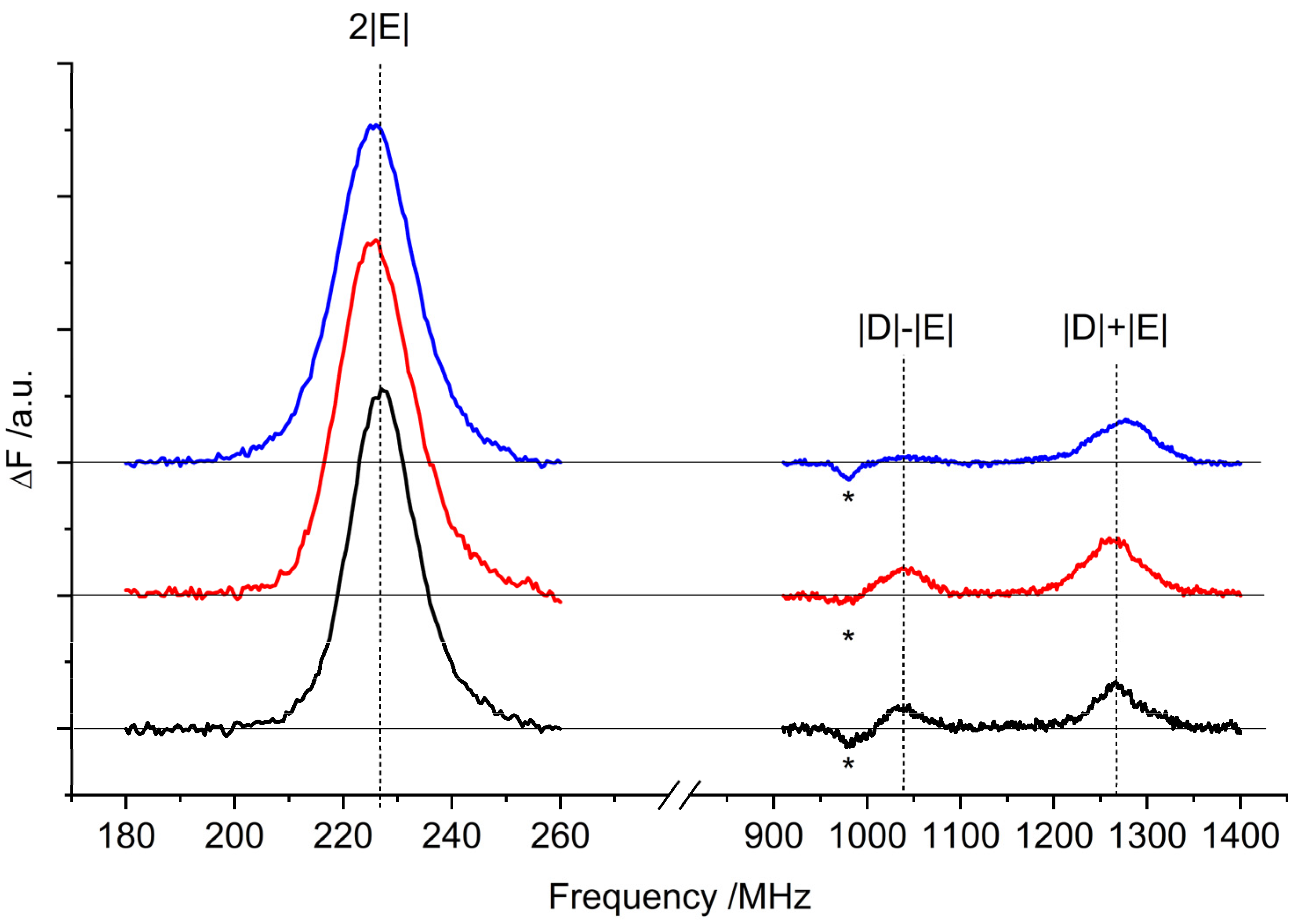
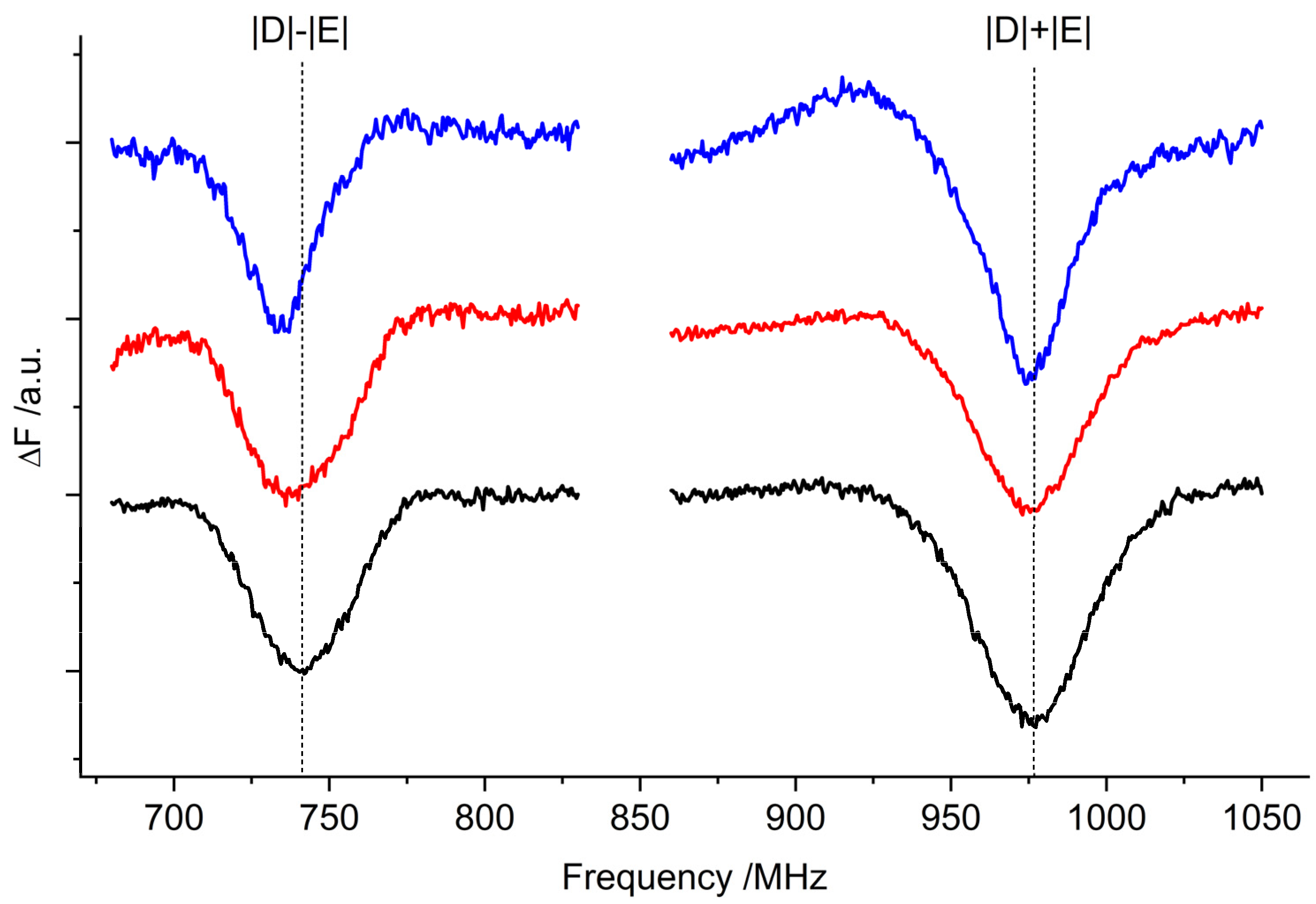
2.2. Optically Detected Magnetic Resonance (ODMR) of 3Car
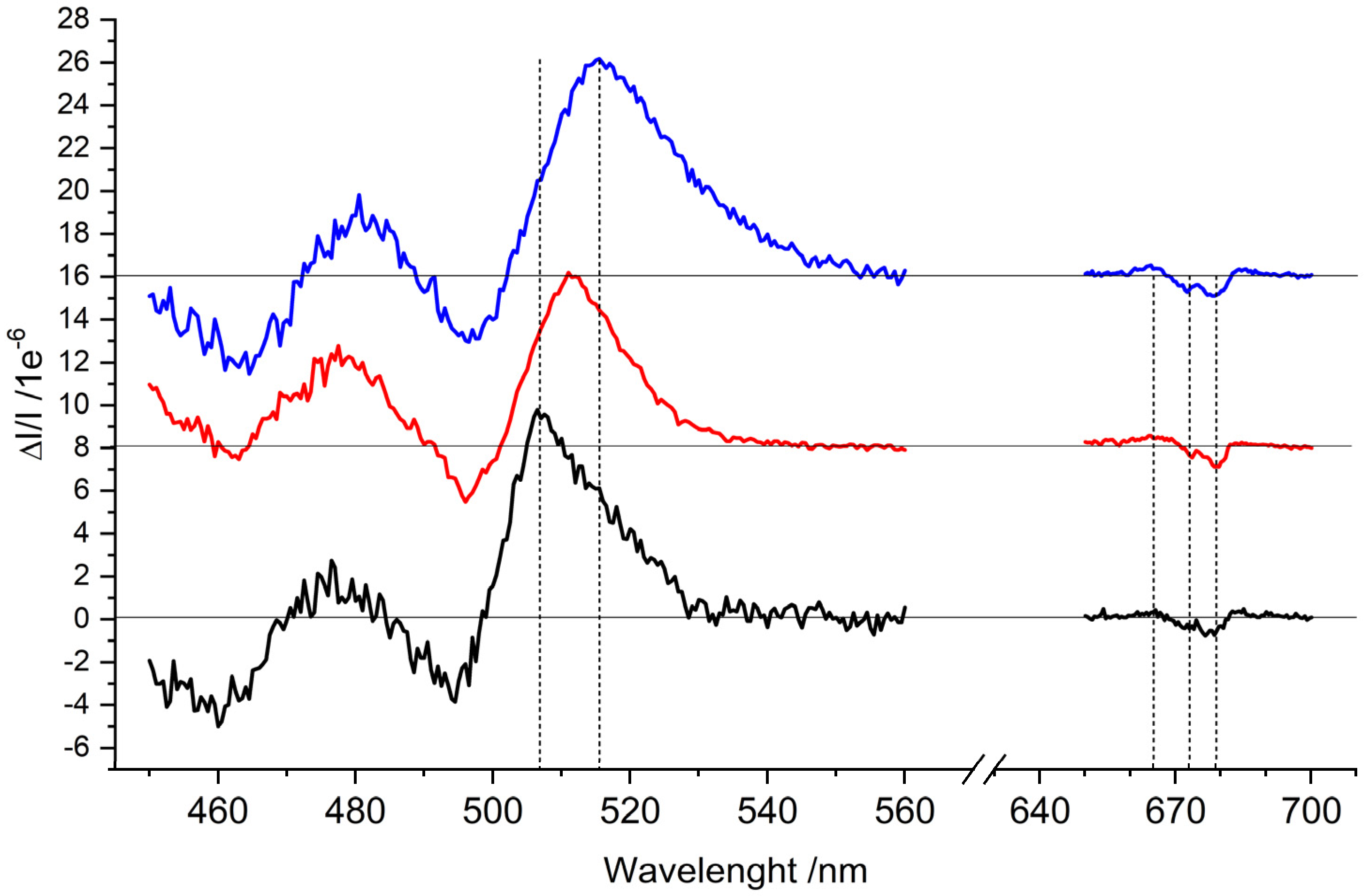
2.3. Optically Detected Magnetic Resonance of 3Chl
3. Discussion
4. Materials and Methods
4.1. Sample Preparation
4.2. ODMR Experiments
4.3. TR-EPR Experiments
4.4. Triplet-State EPR Simulations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Caffarri, S.; Croce, R.; Breton, J.; Bassi, R. The Major Antenna Complex of Photosystem II Has a Xanthophyll Binding Site Not Involved in Light Harvesting. J. Biol. Chem. 2001, 276, 35924–35933. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Yan, H.; Wang, K.; Kuang, T.; Zhang, J.; Gui, L.; An, X.; Chang, W. Crystal structure of spinach major light-harvesting complex at 2.72 Å resolution. Nature 2004, 428, 287–292. [Google Scholar] [CrossRef]
- Rochaix, J.-D. Regulation and Dynamics of the Light-Harvesting System. Annu. Rev. Plant Biol. 2014, 65, 287–309. [Google Scholar] [CrossRef]
- Croce, R.; van Amerongen, H. Light harvesting in oxygenic photosynthesis: Structural biology meets spectroscopy. Science 2020, 369, eaay2058. [Google Scholar] [CrossRef]
- Krieger-Liszkay, A. Singlet oxygen production in photosynthesis. J. Exp. Bot. 2004, 56, 337–346. [Google Scholar] [CrossRef] [Green Version]
- Pinnola, A.; Bassi, R. Molecular mechanisms involved in plant photoprotection. Biochem. Soc. Trans. 2018, 46, 467–482. [Google Scholar] [CrossRef]
- Murchie, E.H.; Ruban, A.V. Dynamic non-photochemical quenching in plants: From molecular mechanism to productivity. Plant J. 2020, 101, 885–896. [Google Scholar] [CrossRef] [Green Version]
- Ruban, A.V. Nonphotochemical Chlorophyll Fluorescence Quenching: Mechanism and Effectiveness in Protecting Plants from Photodamage. Plant Physiol. 2016, 170, 1903–1916. [Google Scholar] [CrossRef] [Green Version]
- Carbonera, D.; Di Valentin, M.; Agostini, G.; Giacometti, G.; Liddell, P.A.; Gust, D.; Moore, A.L.; Moore, T.A. Energy transfer and spin polarization of the carotenoid triplet state in synthetic carotenoporphyrin dyads and in natural antenna complexes. Appl. Magn. Reson. 1997, 13, 487–504. [Google Scholar] [CrossRef]
- Remelli, R.; Varotto, C.; Sandonà, D.; Croce, R.; Bassi, R. Chlorophyll Binding to Monomeric Light-harvesting Complex. J. Biol. Chem. 1999, 274, 33510–33521. [Google Scholar] [CrossRef] [Green Version]
- Agostini, A.; Nicol, L.; Da Roit, N.; Bortolus, M.; Croce, R.; Carbonera, D. Altering the exciton landscape by removal of specific chlorophylls in monomeric LHCII provides information on the sites of triplet formation and quenching by means of ODMR and EPR spectroscopies. Biochim. Biophys. Acta-Bioenerg. 2021, 1862, 148481. [Google Scholar] [CrossRef]
- Formaggio, E.; Cinque, G.; Bassi, R. Functional architecture of the major light-harvesting complex from higher plants. J. Mol. Biol. 2001, 314, 1157–1166. [Google Scholar] [CrossRef]
- Mascoli, V.; Liguori, N.; Cupellini, L.; Elias, E.; Mennucci, B.; Croce, R. Uncovering the interactions driving carotenoid binding in light-harvesting complexes. Chem. Sci. 2021, 12, 5113–5122. [Google Scholar] [CrossRef]
- Saccon, F.; Durchan, M.; Polívka, T.; Ruban, A.V. The robustness of the terminal emitter site in major LHCII complexes controls xanthophyll function during photoprotection. Photochem. Photobiol. Sci. 2020, 19, 1308–1318. [Google Scholar] [CrossRef]
- Saccon, F.; Durchan, M.; Kaňa, R.; Prášil, O.; Ruban, A.V.; Polívka, T. Spectroscopic Properties of Violaxanthin and Lutein Triplet States in LHCII are Independent of Carotenoid Composition. J. Phys. Chem. B 2019, 123, 9312–9320. [Google Scholar] [CrossRef]
- Dall’Osto, L.; Lico, C.; Alric, J.; Giuliano, G.; Havaux, M.; Bassi, R. Lutein is needed for efficient chlorophyll triplet quenching in the major LHCII antenna complex of higher plants and effective photoprotection in vivounder strong light. BMC Plant Biol. 2006, 6, 32. [Google Scholar] [CrossRef] [Green Version]
- Dall’Osto, L.; Holt, N.E.; Kaligotla, S.; Fuciman, M.; Cazzaniga, S.; Carbonera, D.; Frank, H.A.; Alric, J.; Bassi, R. Zeaxanthin Protects Plant Photosynthesis by Modulating Chlorophyll Triplet Yield in Specific Light-harvesting Antenna Subunits. J. Biol. Chem. 2012, 287, 41820–41834. [Google Scholar] [CrossRef] [Green Version]
- Havaux, M.; Dall’Osto, L.; Cuiné, S.; Giuliano, G.; Bassi, R. The Effect of Zeaxanthin as the Only Xanthophyll on the Structure and Function of the Photosynthetic Apparatus in Arabidopsis thaliana. J. Biol. Chem. 2004, 279, 13878–13888. [Google Scholar] [CrossRef] [Green Version]
- Carbonera, D. Optically detected magnetic resonance (ODMR) of photoexcited triplet states. Photosynth. Res. 2009, 102, 403–414. [Google Scholar] [CrossRef]
- Di Valentin, M.; Biasibetti, F.; Ceola, S.; Carbonera, D. Identification of the Sites of Chlorophyll Triplet Quenching in Relation to the Structure of LHC-II from Higher Plants. Evidence from EPR Spectroscopy. J. Phys. Chem. B 2009, 113, 13071–13078. [Google Scholar] [CrossRef]
- Agostini, A.; Palm, D.M.; Paulsen, H.; Carbonera, D. Optically Detected Magnetic Resonance of Chlorophyll Triplet States in Water-Soluble Chlorophyll Proteins from Lepidium virginicum: Evidence for Excitonic Interaction among the Four Pigments. J. Phys. Chem. B 2018, 122, 6156–6163. [Google Scholar] [CrossRef]
- Carbonera, D.; Agostini, A.; Di Valentin, M.; Gerotto, C.; Basso, S.; Giacometti, G.M.; Morosinotto, T. Photoprotective sites in the violaxanthin–chlorophyll a binding Protein (VCP) from Nannochloropsis gaditana. Biochim. Biophys. Acta-Bioenerg. 2014, 1837, 1235–1246. [Google Scholar] [CrossRef]
- Di Valentin, M.; Carbonera, D. The fine tuning of carotenoid–chlorophyll interactions in light-harvesting complexes: An important requisite to guarantee efficient photoprotection via triplet–triplet energy transfer in the complex balance of the energy transfer processes. J. Phys. B At. Mol. Opt. Phys. 2017, 50, 162001. [Google Scholar] [CrossRef]
- Ruban, A.V.; Berera, R.; Ilioaia, C.; van Stokkum, I.H.M.; Kennis, J.T.M.; Pascal, A.A.; van Amerongen, H.; Robert, B.; Horton, P.; van Grondelle, R. Identification of a mechanism of photoprotective energy dissipation in higher plants. Nature 2007, 450, 575–578. [Google Scholar] [CrossRef]
- Di Valentin, M.; Büchel, C.; Giacometti, G.M.; Carbonera, D. Chlorophyll triplet quenching by fucoxanthin in the fucoxanthin–chlorophyll protein from the diatom Cyclotella meneghiniana. Biochem. Biophys. Res. Commun. 2012, 427, 637–641. [Google Scholar] [CrossRef]
- Carbonera, D.; Giacometti, G.; Agostini, G.; Angerhofer, A.; Aust, V. ODMR of carotenoid and chlorophyll triplets in CP43 and CP47 complexes of spinach. Chem. Phys. Lett. 1992, 194, 275–281. [Google Scholar] [CrossRef]
- Peterman, E.J.; Dukker, F.M.; van Grondelle, R.; van Amerongen, H. Chlorophyll a and carotenoid triplet states in light-harvesting complex II of higher plants. Biophys. J. 1995, 69, 2670–2678. [Google Scholar] [CrossRef] [Green Version]
- Carbonera, D.; Giacometti, G.; Agostini, G. FDMR of Carotenoid and Chlorophyll triplets in light-harvesting complex LHCII of spinach. Appl. Magn. Reson. 1992, 3, 859–872. [Google Scholar] [CrossRef]
- van der Vos, R.; Carbonera, D.; Hoff, A.J. Microwave and optical spectroscopy of carotenoid triplets in light-harvesting complex LHC II of spinach by absorbance-detected magnetic resonance. Appl. Magn. Reson. 1991, 2, 179–202. [Google Scholar] [CrossRef]
- Ho, J.; Kish, E.; Méndez-Hernández, D.D.; WongCarter, K.; Pillai, S.; Kodis, G.; Niklas, J.; Poluektov, O.G.; Gust, D.; Moore, T.A.; et al. Triplet–triplet energy transfer in artificial and natural photosynthetic antennas. Proc. Natl. Acad. Sci. USA 2017, 114, E5513–E5521. [Google Scholar] [CrossRef] [Green Version]
- Aust, V.; Angerhofer, A.; Ullrich, J.; von Schütz, J.U.; Wolf, H.C.; Cogdell, R.J. ADMR of carotenoid triplet states in bacterial photosynthetic antenna and reaction center complexes. Chem. Phys. Lett. 1991, 181, 213–221. [Google Scholar] [CrossRef]
- Agostini, A.; Niklas, J.; Schulte, T.; Di Valentin, M.; Bortolus, M.; Hofmann, E.; Lubitz, W.; Carbonera, D. Changing the site energy of per-614 in the Peridinin-chlorophyll a-protein does not alter its capability of chlorophyll triplet quenching. Biochim. Biophys. Acta-Bioenerg. 2018, 1859, 612–618. [Google Scholar] [CrossRef]
- Galinato, M.G.I.; Niedzwiedzki, D.; Deal, C.; Birge, R.R.; Frank, H.A. Cation radicals of xanthophylls. Photosynth. Res. 2007, 94, 67–78. [Google Scholar] [CrossRef]
- Spezia, R.; Aschi, M.; Di Nola, A.; Di Valentin, M.; Carbonera, D.; Amadei, A. The Effect of Protein Conformational Flexibility on the Electronic Properties of a Chromophore. Biophys. J. 2003, 84, 2805–2813. [Google Scholar] [CrossRef] [Green Version]
- Müller, P.; Li, X.-P.; Niyogi, K.K. Non-Photochemical Quenching. A Response to Excess Light Energy. Plant Physiol. 2001, 125, 1558–1566. [Google Scholar] [CrossRef] [Green Version]
- Ruban, A.V.; Johnson, M.P.; Duffy, C.D.P. The photoprotective molecular switch in the photosystem II antenna. Biochim. Biophys. Acta-Bioenerg. 2012, 1817, 167–181. [Google Scholar] [CrossRef] [Green Version]
- Holt, N.E.; Fleming, G.R.; Niyogi, K.K. Toward an Understanding of the Mechanism of Nonphotochemical Quenching in Green Plants. Biochemistry 2004, 43, 8281–8289. [Google Scholar] [CrossRef]
- Dreuw, A.; Fleming, G.R.; Head-Gordon, M. Chlorophyll fluorescence quenching by xanthophylls. Phys. Chem. Chem. Phys. 2003, 5, 3247. [Google Scholar] [CrossRef]
- Dreuw, A.; Fleming, G.R.; Head-Gordon, M. Charge-Transfer State as a Possible Signature of a Zeaxanthin−Chlorophyll Dimer in the Non-photochemical Quenching Process in Green Plants. J. Phys. Chem. B 2003, 107, 6500–6503. [Google Scholar] [CrossRef]
- Cupellini, L.; Calvani, D.; Jacquemin, D.; Mennucci, B. Charge transfer from the carotenoid can quench chlorophyll excitation in antenna complexes of plants. Nat. Commun. 2020, 11, 662. [Google Scholar] [CrossRef]
- Sláma, V.; Cupellini, L.; Mennucci, B. Exciton properties and optical spectra of light harvesting complex II from a fully atomistic description. Phys. Chem. Chem. Phys. 2020, 22, 16783–16795. [Google Scholar] [CrossRef]
- Novoderezhkin, V.I.; Palacios, M.A.; van Amerongen, H.; van Grondelle, R. Excitation Dynamics in the LHCII Complex of Higher Plants: Modeling Based on the 2.72 Å Crystal Structure. J. Phys. Chem. B 2005, 109, 10493–10504. [Google Scholar] [CrossRef] [Green Version]
- Bassi, R.; Rigoni, F.; Barbato, R.; Giacometti, G.M. Light-harvesting chlorophyll a/b proteins (LHCII) populations in phosphorylated membranes. Biochim. Biophys. Acta-Bioenerg. 1988, 936, 29–38. [Google Scholar] [CrossRef]
- Dall’Osto, L.; Caffarri, S.; Bassi, R. A Mechanism of Nonphotochemical Energy Dissipation, Independent from PsbS, Revealed by a Conformational Change in the Antenna Protein CP26. Plant Cell 2005, 17, 1217–1232. [Google Scholar] [CrossRef] [Green Version]
- Dainese, P.; Hoyer-Hansen, G.; Bassi, R. The resolution of chlorophyll a/b binding proteins by a preparative method based on flat bed isoelectric focusing. Photochem. Photobiol. 1990, 51, 693–703. [Google Scholar] [CrossRef]
- Lampoura, S.S.; Barzda, V.; Owen, G.M.; Hoff, A.J.; van Amerongen, H. Aggregation of LHCII Leads to a Redistribution of the Triplets over the Central Xanthophylls in LHCII. Biochemistry 2002, 41, 9139–9144. [Google Scholar] [CrossRef]
- Santabarbara, S.; Agostini, G.; Casazza, A.P.; Syme, C.D.; Heathcote, P.; Böhles, F.; Evans, M.C.W.; Jennings, R.C.; Carbonera, D. Chlorophyll triplet states associated with Photosystem I and Photosystem II in thylakoids of the green alga Chlamydomonas reinhardtii. Biochim. Biophys. Acta-Bioenerg. 2007, 1767, 88–105. [Google Scholar] [CrossRef]
- Carbonera, D.; Collareta, P.; Giacometti, G. The P700 triplet state in an intact environment detected by ODMR. A well resolved triplet minus singlet spectrum. Biochim. Biophys. Acta-Bioenerg. 1997, 1322, 115–128. [Google Scholar] [CrossRef]
- Santabarbara, S.; Bordignon, E.; Jennings, R.C.; Carbonera, D. Chlorophyll Triplet States Associated with Photosystem II of Thylakoids. Biochemistry 2002, 41, 8184–8194. [Google Scholar] [CrossRef]
- Stoll, S.; Schweiger, A. EasySpin, a comprehensive software package for spectral simulation and analysis in EPR. J. Magn. Reson. 2006, 178, 42–55. [Google Scholar] [CrossRef]
- Carbonera, D.; di Valentin, M.; Corvaja, C.; Giacometti, G.; Agostini, G.; Liddell, P.A.; Moore, A.L.; Moore, T.A.; Gust, D. Carotenoid triplet detection by time-resolved EPR spectroscopy in carotenopyropheophorbide dyads. J. Photochem. Photobiol. A Chem. 1997, 105, 329–335. [Google Scholar] [CrossRef]
- Di Valentin, M.; Meneghin, E.; Orian, L.; Polimeno, A.; Büchel, C.; Salvadori, E.; Kay, C.W.M.; Carbonera, D. Triplet–triplet energy transfer in fucoxanthin-chlorophyll protein from diatom Cyclotella meneghiniana: Insights into the structure of the complex. Biochim. Biophys. Acta-Bioenerg. 2013, 1827, 1226–1234. [Google Scholar] [CrossRef]
- Di Valentin, M.; Agostini, G.; Salvadori, E.; Ceola, S.; Giacometti, G.M.; Hiller, R.G.; Carbonera, D. Triplet–triplet energy transfer in Peridinin-Chlorophyll a-protein reconstituted with Chl a and Chl d as revealed by optically detected magnetic resonance and pulse EPR: Comparison with the native PCP complex from Amphidinium carterae. Biochim. Biophys. Acta-Bioenerg. 2009, 1787, 168–175. [Google Scholar] [CrossRef]
- Mozzo, M.; Dall’Osto, L.; Hienerwadel, R.; Bassi, R.; Croce, R. Photoprotection in the Antenna Complexes of Photosystem II. J. Biol. Chem. 2008, 283, 6184–6192. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.P.; Havaux, M.; Triantaphylides, C.; Ksas, B.; Pascal, A.A.; Robert, B.; Davison, P.A.; Ruban, A.V.; Horton, P. Elevated Zeaxanthin Bound to Oligomeric LHCII Enhances the Resistance of Arabidopsis to Photooxidative Stress by a Lipid-protective, Antioxidant Mechanism. J. Biol. Chem. 2007, 282, 22605–22618. [Google Scholar] [CrossRef] [Green Version]
- Morosinotto, T.; Caffarri, S.; Dall’Osto, L.; Bassi, R. Mechanistic aspects of the xanthophyll dynamics in higher plant thylakoids. Physiol. Plant. 2003, 119, 347–354. [Google Scholar] [CrossRef]
- Cazzaniga, S.; Bressan, M.; Carbonera, D.; Agostini, A.; Dall’Osto, L. Differential Roles of Carotenes and Xanthophylls in Photosystem I Photoprotection. Biochemistry 2016, 55, 3636–3649. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2|E| FDMR/MHz | |D| + |E| FDMR/MHz | |D| − |E| FDMR/MHz | |E| a FDMR/MHz | |D| b TR-EPR/MHz | |
|---|---|---|---|---|---|
| WT | 227 ± 1 | 1267 ± 1 | 1037 ± 0.1 | 113 ± 1 | 1152 ± 1 |
| lut2 | 226 ± 1 | 1262 ± 1 | 1040 ± 0.1 | 113 ± 1 | 1151 ± 1 |
| lut2 npq2 | 225 ± 1 | 1275 ± 1 | 1046 ± 0.1 | 112 ± 1 | 1160 ± 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carbonera, D.; Agostini, A.; Bortolus, M.; Dall’Osto, L.; Bassi, R. Violaxanthin and Zeaxanthin May Replace Lutein at the L1 Site of LHCII, Conserving the Interactions with Surrounding Chlorophylls and the Capability of Triplet–Triplet Energy Transfer. Int. J. Mol. Sci. 2022, 23, 4812. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094812
Carbonera D, Agostini A, Bortolus M, Dall’Osto L, Bassi R. Violaxanthin and Zeaxanthin May Replace Lutein at the L1 Site of LHCII, Conserving the Interactions with Surrounding Chlorophylls and the Capability of Triplet–Triplet Energy Transfer. International Journal of Molecular Sciences. 2022; 23(9):4812. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094812
Chicago/Turabian StyleCarbonera, Donatella, Alessandro Agostini, Marco Bortolus, Luca Dall’Osto, and Roberto Bassi. 2022. "Violaxanthin and Zeaxanthin May Replace Lutein at the L1 Site of LHCII, Conserving the Interactions with Surrounding Chlorophylls and the Capability of Triplet–Triplet Energy Transfer" International Journal of Molecular Sciences 23, no. 9: 4812. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094812