
Effects of Concurrent Exposure to Chronic Restraint-Induced Stress and Total-Body Iron Ion Radiation on Induction of Kidney Injury in Mice

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Treatment
2.2. The Histological Observation
2.3. Masson Staining
2.4. TUNEL Assay
2.5. Immunofluorescence
2.6. Immunohistochemistry
2.7. Western Blotting
2.8. Statistical Analysis
3. Results
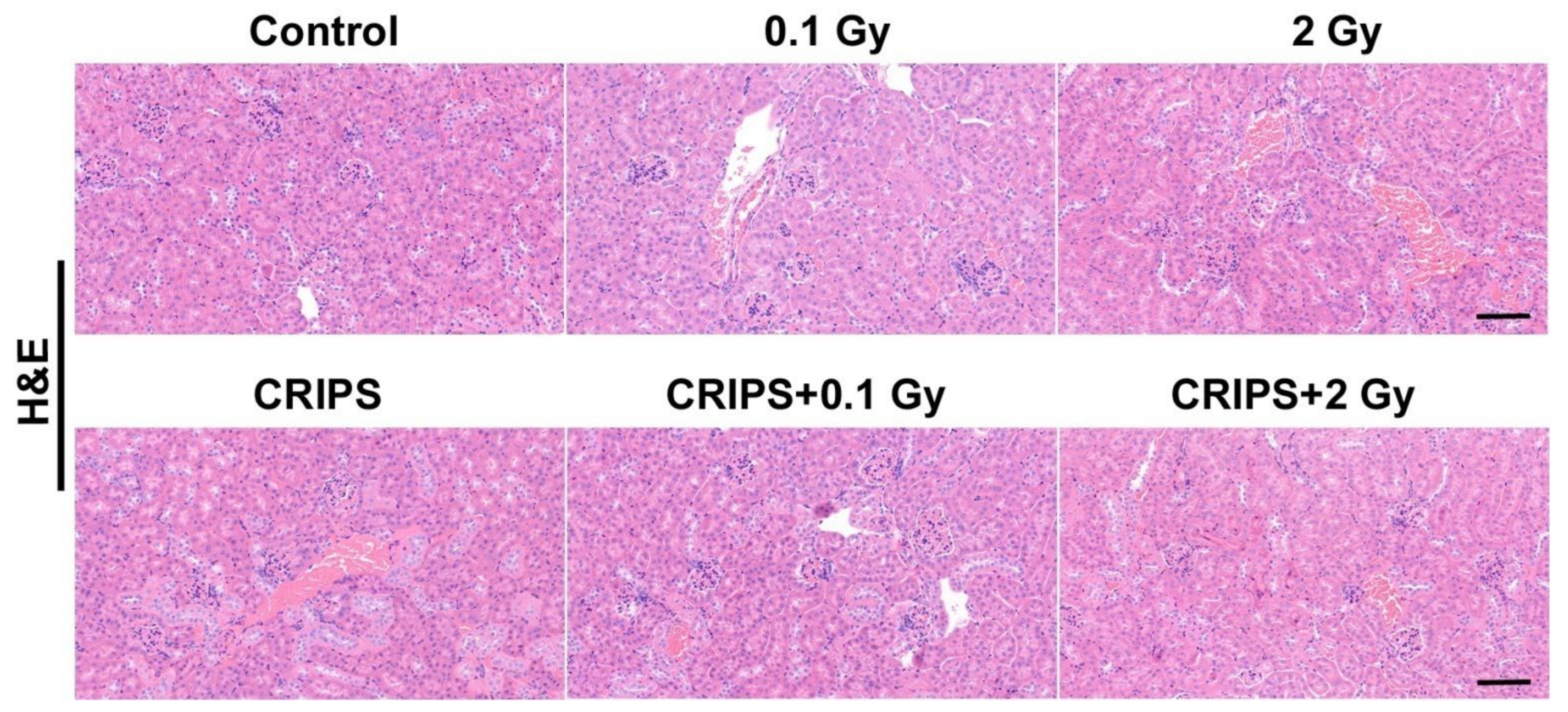
3.1. The Histopathological Observation
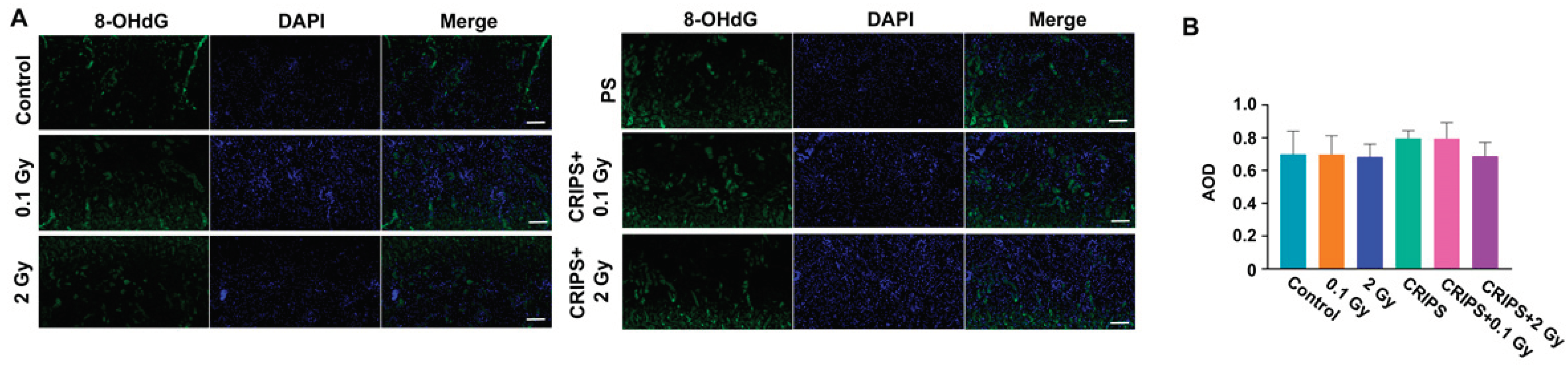
3.2. The 8-OHdG Expression
3.3. The Apoptotic Cells Detection
3.4. The Evaluation of Renal Fibrosis
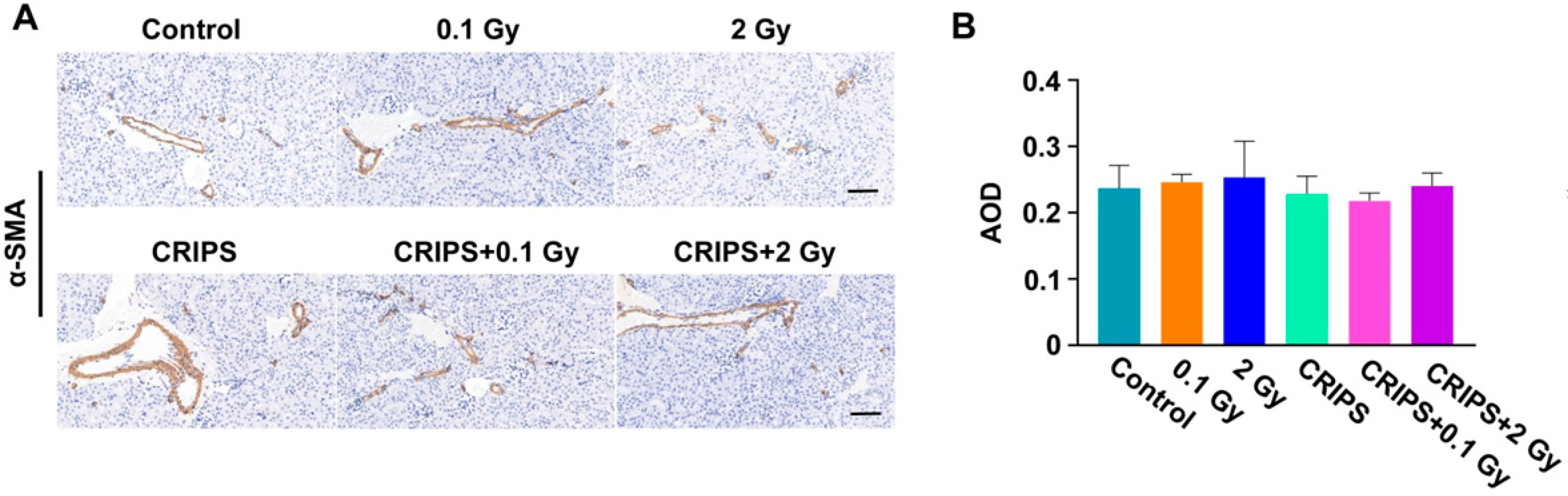
3.5. The FN and α-SMA Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kemeny, M.E.; Schedlowski, M. Understanding the interaction between psychosocial stress and immune-related diseases: A stepwise progression. Brain Behav. Immun. 2007, 21, 1009–1018. [Google Scholar] [CrossRef] [PubMed]
- Schoenfeld, T.J.; McCausland, H.C.; Morris, H.D.; Padmanaban, V.; Cameron, H.A. Stress and Loss of Adult Neurogenesis Differentially Reduce Hippocampal Volume. Biol. Psychiatry 2017, 82, 914–923. [Google Scholar] [CrossRef] [PubMed]
- Pandey, B.N.; Kumar, A.; Tiwari, P.; Mishra, K.P. Radiobiological basis in management of accidental radiation exposure. Int. J. Radiat. Biol. 2010, 86, 613–635. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Gonon, G.; Buonanno, M.; Autsavapromporn, N.; de Toledo, S.M.; Pain, D.; Azzam, E.I. Health Risks of Space Exploration: Targeted and Nontargeted Oxidative Injury by High-Charge and High-Energy Particles. Antioxid. Redox Signal. 2014, 20, 1501–1523. [Google Scholar] [CrossRef] [Green Version]
- Duan, X.; Zhang, H.; Liu, B.; Li, X.-D.; Gao, Q.-X.; Wu, Z.-H. Apoptosis of murine melanoma cells induced by heavy-ion radiation combined with Tp53 gene transfer. Int. J. Radiat. Biol. 2008, 84, 211–217. [Google Scholar] [CrossRef]
- Furukawa, S.; Nagamatsu, A.; Nenoi, M.; Fujimori, A.; Kakinuma, S.; Katsube, T.; Wang, B.; Tsuruoka, C.; Shirai, T.; Nakamura, A.J.; et al. Space Radiation Biology for “Living in Space”. BioMed Res. Int. 2020, 2020, 4703286. [Google Scholar] [CrossRef] [Green Version]
- Madrazo, A.; Schwarz, G.; Churg, J. Radiation nephritis: A review. J. Urol. 1975, 114, 822–827. [Google Scholar] [CrossRef]
- Katsube, T.; Wang, B.; Tanaka, K.; Ninomiya, Y.; Hirakawa, H.; Liu, C.; Maruyama, K.; Vares, G.; Liu, Q.; Kito, S.; et al. Synergistic Effects of Chronic Restraint-Induced Stress and Low-Dose 56Fe-particle Irradiation on Induction of Chromosomal Aberrations in Trp53-Heterozygous Mice. Radiat. Res. 2021, 196, 100–112. [Google Scholar] [CrossRef]
- Wang, B.; Tanaka, K.; Katsube, T.; Ninomiya, Y.; Vares, G.; Liu, Q.; Morita, A.; Nakajima, T.; Nenoi, M. Chronic restraint-induced stress has little modifying effect on radiation hematopoietic toxicity in mice. J. Radiat. Res. 2015, 56, 760–767. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wang, B.; Zhang, H.; Katsube, T.; Xie, Y.; Gan, L. Apoptosis Induction by Iron Radiation via Inhibition of Autophagy in Trp53+/− Mouse Testes: Is Chronic Restraint-Induced Stress a Modifying Factor? Int. J. Biol. Sci. 2018, 14, 1109–1121. [Google Scholar] [CrossRef] [Green Version]
- Liakopoulos, V.; Leivaditis, K.; Eleftheriadis, T.; Dombros, N. The kidney in space. Int. Urol. Nephrol. 2012, 44, 1893–1901. [Google Scholar] [CrossRef]
- Sánchez-Solís, C.N.; Romero, E.C.; Soto-Rodríguez, I.; Arteaga-Castañeda, M.D.L.; De León-Ramírez, Y.M.; Rodríguez-Antolín, J.; Nicolás-Toledo, L. High-sucrose diet potentiates hyperaldosteronism and renal injury induced by stress in young adult rats. Clin. Exp. Pharmacol. Physiol. 2020, 47, 1985–1994. [Google Scholar] [CrossRef]
- Nastase, M.V.; Zeng-Brouwers, J.; Wygrecka, M.; Schaefer, L. Targeting renal fibrosis: Mechanisms and drug delivery systems. Adv. Drug Deliv. Rev. 2018, 129, 295–307. [Google Scholar] [CrossRef]
- Humphreys, B.D. Mechanisms of Renal Fibrosis. Annu. Rev. Physiol. 2018, 80, 309–326. [Google Scholar] [CrossRef]
- Klaus, R.; Niyazi, M.; Lange-Sperandio, B. Radiation-induced kidney toxicity: Molecular and cellular pathogenesis. Radiat. Oncol. 2021, 16, 43. [Google Scholar] [CrossRef]
- Wei, J.; Wang, B.; Wang, H.; Meng, L.; Zhao, Q.; Li, X.; Xin, Y.; Jiang, X. Radiation-Induced Normal Tissue Damage: Oxidative Stress and Epigenetic Mechanisms. Oxidative Med. Cell. Longev. 2019, 2019, 3010342. [Google Scholar] [CrossRef]
- Wyld, M.; Morton, R.; Hayen, A.; Howard, K.; Webster, A. A Systematic Review and Meta-Analysis of Utility-Based Quality of Life in Chronic Kidney Disease Treatments. PLoS Med. 2012, 9, e1001307. [Google Scholar] [CrossRef]
- Wang, A.Y.-M.; Kalantar-Zadeh, K.; Fouque, D.; Wee, P.T.; Kovesdy, C.P.; Price, S.R.; Kopple, J.D. Precision Medicine for Nutritional Management in End-Stage Kidney Disease and Transition to Dialysis. Semin. Nephrol. 2018, 38, 383–396. [Google Scholar] [CrossRef]
- Zhou, Y.; Chai, P.; Wang, J.; Li, L.; Chen, M.H. Wingless/int-1induced secreted protein-1: A new biomarker for renal fibrosis. J. Biol. Regul. Homeost. Agents 2021, 34, 97–103. [Google Scholar] [CrossRef]
- Qi, W.; Chen, X.; Poronnik, P.; Pollock, C.A. The renal cortical fibroblast in renal tubulointerstitial fibrosis. Int. J. Biochem. Cell Biol. 2006, 38, 1–5. [Google Scholar] [CrossRef]
- Huang, C.-Y.; Su, C.-T.; Chung, C.-J.; Pu, Y.-S.; Chu, J.-S.; Yang, H.-Y.; Wu, C.-C.; Hsueh, Y.-M. Urinary total arsenic and 8-hydroxydeoxyguanosine are associated with renal cell carcinoma in an area without obvious arsenic exposure. Toxicol. Appl. Pharmacol. 2012, 262, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, B.; Chen, Z.; He, Y.; Ru, F.; Liu, P.; Chen, X. The long noncoding RNA myocardial infarction-associated transcript modulates the epithelial-mesenchymal transition in renal interstitial fibrosis. Life Sci. 2020, 241, 117187. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-C.; Lin, Y.-Y.; Yang, A.-L.; Kuo, T.-W.; Kuo, C.-H.; Lee, S.-D. Anti-Renal Fibrotic Effect of Exercise Training in Hypertension. Int. J. Mol. Sci. 2018, 19, 613–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.-L.; Qiao, S.-K.; Wang, R.-Y.; Guo, X.-N. NGAL attenuates renal ischemia/reperfusion injury through autophagy activation and apoptosis inhibition in rats. Chem. Biol. Interact. 2018, 289, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Xiang, C.Y.; Yan, Y.; Zhang, D.G. Alleviation of the doxorubicin-induced nephrotoxicity by fasudil in vivo and in vitro. J. Pharmacol. Sci. 2021, 145, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Shang, Q.H.; Chen, C.Y. Effects of short-term high salt diet on tubule epithelial mesenchymal transformation and renal fibrosis in C57BL/6J mice. Chin. J. Hypertens. 2020, 28, 757–766. [Google Scholar] [CrossRef]
- Cohen, E.P.; Lenarczyk, M.; Fish, B.L.; Jia, S.; Hessner, M.J.; E Moulder, J. Evaluation of Genomic Evidence for Oxidative Stress in Experimental Radiation Nephropathy. J. Genet. Disord. Genet. Rep. 2013, 2, 1000101. [Google Scholar] [CrossRef]
- Yan, Y.; Zhang, K.; Zhou, G.; Hu, W. MicroRNAs Responding to Space Radiation. Int. J. Mol. Sci. 2020, 21, 6603. [Google Scholar] [CrossRef]
- Straub, J.M.; New, J.; Hamilton, C.D.; Lominska, C.; Shnayder, Y.; Thomas, S.M. Radiation-induced fibrosis: Mechanisms and implications for therapy. J. Cancer Res. Clin. Oncol. 2015, 141, 1985–1994. [Google Scholar] [CrossRef] [Green Version]
- Pavlakou, P.; Dounousi, E.; Roumeliotis, S.; Eleftheriadis, T.; Liakopoulos, V. Oxidative Stress and the Kidney in the Space Environment. Int. J. Mol. Sci. 2018, 19, 3176. [Google Scholar] [CrossRef] [Green Version]
- Durante, M.; Kronenberg, A. Ground-based research with heavy ions for space radiation protection. Adv. Space Res. 2005, 35, 180–184. [Google Scholar] [CrossRef]
- Turker, M.S.; Grygoryev, D.; Lasarev, M.; Ohlrich, A.; Rwatambuga, F.A.; Johnson, S.; Dan, C.; Eckelmann, B.; Hryciw, G.; Mao, J.-H.; et al. Simulated space radiation-induced mutants in the mouse kidney display widespread genomic change. PLoS ONE 2017, 12, e0180412. [Google Scholar] [CrossRef] [Green Version]
- Stone, H.B.; Moulder, J.E.; Coleman, C.N.; Ang, K.K.; Anscher, M.S.; Barcellos-Hoff, M.H.; Dynan, W.S.; Fike, J.R.; Grdina, D.J.; Greenberger, J.S.; et al. Models for evaluating agents intended for the prophylaxis, mitigation and treatment of radiation injuries. Report of an NCI Workshop, December 3–4 2003. Radiat. Res. 2004, 162, 711–728. [Google Scholar] [CrossRef]
- Robbins, M.E.; Zhao, W.; Davis, C.S.; Toyokuni, S.; Bonsib, S.M. Radiation-induced kidney injury: A role for chronic oxidative stress? Micron 2002, 33, 133–141. [Google Scholar] [CrossRef]
- Emami, B.; Lyman, J.; Brown, A.; Coia, L.; Goitein, M.; Munzenrider, J.E.; Shank, B.; Solin, L.J.; Wesson, M. Tolerance of normal tissue to therapeutic irradiation. Int. J. Radiat. Oncol. Biol. Phys. 1991, 21, 109–122. [Google Scholar] [CrossRef]
- Lenarczyk, M.; Cohen, E.P.; Fish, B.L.; Irving, A.A.; Sharma, M.; Driscoll, C.D.; Moulder, J.E. Chronic Oxidative Stress as a Mechanism for Radiation Nephropathy. Radiat. Res. 2009, 171, 164–172. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Treatment |
|---|---|
| Control (n = 6) | Receiving neither chronic restraint nor TBI with 56Fe irradiation |
| 0.1 Gy (n = 6) | Receiving only 56Fe-TBI at 0.1 Gy |
| 2.0 Gy (n = 6) | Receiving only 56Fe-TBI at 2.0 Gy |
| CRIPS (n = 6) | Receiving only chronic restraint to simulate chronic restraint-induced psychological stress |
| CRIPS + 0.1 Gy (n = 6) | Receiving 0.1 Gy 56Fe-TBI on day 8 of the 28-day restraint regimen |
| CRIPS + 2.0 Gy (n = 6) | Receiving 2.0 Gy 56Fe-TBI on day 8 of the 28-day restraint regimen |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, D.; Li, H.; Katsube, T.; Huang, G.; Liu, J.; Wang, B.; Zhang, H. Effects of Concurrent Exposure to Chronic Restraint-Induced Stress and Total-Body Iron Ion Radiation on Induction of Kidney Injury in Mice. Int. J. Mol. Sci. 2022, 23, 4866. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094866
Xu D, Li H, Katsube T, Huang G, Liu J, Wang B, Zhang H. Effects of Concurrent Exposure to Chronic Restraint-Induced Stress and Total-Body Iron Ion Radiation on Induction of Kidney Injury in Mice. International Journal of Molecular Sciences. 2022; 23(9):4866. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094866
Chicago/Turabian StyleXu, Duling, Hongyan Li, Takanori Katsube, Guomin Huang, Jiadi Liu, Bing Wang, and Hong Zhang. 2022. "Effects of Concurrent Exposure to Chronic Restraint-Induced Stress and Total-Body Iron Ion Radiation on Induction of Kidney Injury in Mice" International Journal of Molecular Sciences 23, no. 9: 4866. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094866